
Introduction
Most studies on chemotherapy resistance of tumor cells focus on genetic or phenotypic alterations of the cancer cell itself. However, there is growing evidence that the stroma of solid tumors interacts with cancer cells. Tumor-stroma interaction is considered a significant determinant of disease progression and metastasis [1, 2]. Stromal fibroblasts induce epithelial to mesenchymal transition (EMT) in head and neck squamous cell carcinoma (HNSCC) tumor cells [3]. EMT is a reversible cellular process mainly induced by paracrine secretion of small molecules by tumor-associated fibroblasts [4]. Among these small molecules, transforming growth factor-β1 (TGF-β1) is thought to be one of the most relevant mediators [5, 6]. TGF-β1 interacts with TGF-β1 two type I and two type II transmembrane kinase receptors. These receptors activate Smad 2 and 3 signaling pathways, which form a complex with Smad 4, leading to the expression of EMT-activating transcriptional factors such as Snail/Slug [7]. Two major hallmarks of EMT in epithelial tumor cells are increased expression of the intermediate filament vimentin and decreased expression of E-cadherin [3]. We recently reported that EMT in HNSCC-cell lines results in enhanced cell proliferation [8]. In this study, we analyze if EMT increases CDDP (Cisplatin) resistance in two HNSCC cell lines.
SCC-25 cells were orignially isolated from the primary tumor of a patient with tongue carcinoma [9]. SCC-25 cells form tumors in SCID mice but not in athymic nude mice suggesting less aggressive behavior. Moreover, SCC-25 induced tumors do not develop regional or distant metastasis in mouse models [10]. In contrast, Detroit 562 cells grow tumors and develop regional and lung metastases when injected in nude mice [11]. Detroit 562 was isolated from the malignant pleural effusion of a patient with pharyngeal carcinoma [12, 13].
Previously, we had induced EMT by co-cultivation of SCC-25 or Detroit 562 cells and fibroblasts [1]. The resulting co-culture conditioned medium containing EMT-promoting factors was used in this study to induce EMT in pure SCC-25 or Detroit 562 cells. In a second experimental arm, tumor cells were treated with TGF-β1. In a third experimental arm tumor cells were treated with co-culture conditioned medium and a neutralizing dose of anti-TGF-β antibody. SCC-25 and Detroit 562 cells treated with standard cell culture medium served as control in a fourth experimental arm. Changes of vimentin and E-cadherin gene expression and protein synthesis, which were evaluated by quantitative real-time polymerase chain reaction (PCR) and by western blotting were used to confirm EMT. Cell viability was determined with MTT assays and colony-forming ability was investigated using clonogenic assays [14]. Response to CDDP was assessed by cell viability and Clonogenic assays.
Results
Co-culture conditioned medium and TGF-β1 induced EMT in SCC-25 cells
SCC-25 cells were treated with standard medium (control), co-culture conditioned medium or 1 ng/ml TGF-β1 for 3 days. EMT-related gene expression was subsequently measured using real-time PCR and compared with β-actin expression (Figure 1). Relative gene expression of the mesenchymal marker vimentin was higher in SCC-25 cells treated with co-culture conditioned medium (56.6±22.4×10-3) than in control cells (27.1±10.3×10-3; p < 0.001). In contrast, E-cadherin gene expression was down regulated following treatment with co-culture conditioned medium (1.9±1.1×10-3) compared to controls (2.7±0.9×10-3; p < 0.02). EMT-like gene expression changes were also observed in SCC-25 cells treated with 1 ng/ml TGF-β1. TGF-β1 treated cells revealed significantly higher vimentin gene expression (488.1±48 × 10 3) than control cells (74.4±10.1× 10-3; p < 0.001) and lower E-cadherin gene expression levels (0.8±0.3× 10-3) compared to controls (1.6±0.9× 10-3; p < 0.01). At protein level both, co-culture conditioned medium and 1 ng/ml TGF-β1, increased a 46 kD vimentin band. E-cadherin showed a marginal decrease after treatment with co-culture conditioned medium and 1 ng/ml TGF-β1 in SCC-25 cells. Moreover, cell viability of SCC-25 cells treated with co-culture conditioned medium was higher (1.25±0.11) than in untreated controls (1.09±0.23; p < 0.01). In contrast, treatment with TGF-β1 decreased cell viability (0.69±0.007; p < 0.001) compared to controls treated with albumin-containing medium (Figure 3A).
A neutralizing assay with anti-TGF-ß antibody did not reduce the influence of co-culture conditioned medium on cell viability (1.45±0.03; p > 0.1). The number of colonies in clonogenic assays did not differ between standard-medium (3116±654) and co-culture conditioned medium (1805±131; p > 0.5). Furthermore treatment with TGF-β1 (2547±80) or co-culture conditioned medium and anti-TGF-β (1496±119) antibody had no influence on clonogenity of SCC-25 cells (p > 0.1) (Table 1).
Table 1: Co-culture conditioned Medium and TGF-ß1 induced EMT in SCC-25 cells and did not influence the phenotype of Detroit 562 cells
Cell type |
Test |
conditioned Medium |
TGF-ß1 |
SCC-25 |
Vimentin PCR (RNA) |
upregulation*** |
upregulation*** |
E-cadherin PCR (RNA) |
downregulation* |
downregulation** |
|
vimentin Western Blot (protein) |
upregulation (46kD) |
upregulation (46kD) |
|
E-cadherin Western blot (protein) |
downregulation |
downregulation |
|
viability |
increase** |
decrease*** |
|
clonogenity |
no change |
no change |
|
Detroit 562 |
Vimentin PCR (RNA) |
no change |
upregulation*** |
E-cadherin PCR (RNA) |
no change |
no change |
|
Vimentin Western blot (protein) |
no change |
no change |
|
E-cadherin Western blot (protein) |
downregulation |
downregulation |
|
viability |
increase*** |
no change |
|
clonogenity |
no change |
no change |
* p < 0.05, ** p < 0.01, *** p < 0.001
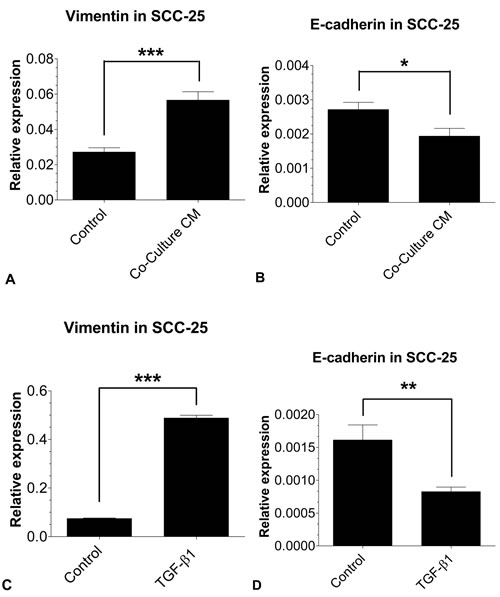
Figure 1: EMT-related gene expression in SCC-25 using real time- PCR: The mRNA expressions of the EMT markers vimentin and E-cadherin in SCC-25 cells treated with co-culture conditioned medium (A, B) or 0.9 ng/ml TGF-β1 (C, D) were quantified relative to SCC-25 control cells. Co-culture conditioned medium treated SCC-25 cells show a significant increase of vimentin A. mRNA expression, while E-cadherin B. mRNA expression was significantly decreased. The treatment with 0.9 ng/ml TGF-β1 led to the same effect to an even greater extent with a highly significant upregulation of vimentin C. and downregulation of E-cadherin D. in SCC-25 cells. Experiments were performed in three replicates with three runs each. *p < 0.05, **p < 0.01, ***p < 0.001.
Conditioned medium did not induce an EMT like phenotype in detroit 562 cells
Treatment with co-culture conditioned medium did not induce significant phenotypic changes in Detroit 562 cells. Vimentin expression (0.59±0.04×10-3 vs. 0.58±0.2×10-3, p > 0.1) and E-cadherin expression (3.6±2.6×10-3 vs. 4.6±0.2×10-3, p > 0.1) did not show significant changes after treatment with co-culture conditioned medium. Treatment with TGF-β1 induced EMT-like phenotypic changes in Detroit 562 cells. TGF-β1 increased vimentin expression about ten fold (5.6±1.4×10-3 vs. 0.58±0.2×10-3, p < 0.0001), but failed to reduce E-cadherin expression (17.5±1×10-3, p > 0.1). At protein level, both, conditioned medium and 1 ng/ml TGF-β1 did not induce changes in vimentin expression (46 kD band) (Figure 2). E-cadherin showed a marginal decrease after treatment with co-culture conditioned medium and 1 ng/ml TGF-β1. Viability increased after treatment with co-culture conditioned medium (2.1±0.04) compared to controls (1.9±0.02, p = 0.001), whereas TGF-β1 had no influence on cell viability (1.9±0.005, p > 0.5). A neutralizing assay with anti-TGF-β antibody did not reduce the effect of co-culture conditioned medium on cell viability (2.2±0.01, p > 0.05). Treatment with co-culture conditioned medium, TGF-β1 and co-culture conditioned medium/anti-TGF-ß antibody had no effect on clonogenity in Detroit 562 cells (standard medium 948±153, CM: 983±38, TGF-β1 876±160; anti-TGF-β: 1000±18; p > 0.1) (Table 1).
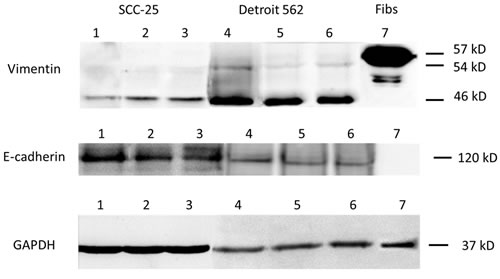
Figure 2: EMT-related protein synthesis in SCC-25 and Detroit 562 cells. Western blots of (supernatants/cell homgenisates), representative sample of three replicates. Lane 1: SCC-25 cells in albumin-containing medium (control), Lane 2: SCC-25 treated with co-culture conditioned medium, Lane 3: SCC-25 treated with 1 ng/ml TGF-β1, Lane 4: Detroit 562 in albumin-containing medium (control), Lane 5: Detroit 562 treated with co-culture conditioned medium, Lane 6: Detroit 562 treated with 1 ng/ml TGF-β1, Lane 7: human gingival fibroblasts. Vimentin was detected in 57, 54 and 46 kD bands. In SCC-25 cells, co-culture conditioned medium and 1 ng/ml TGFβ1 increased the 54 and 46 KD bands. There was a high constitutive expression of 46 kD vimentin in Detroit 562 cells without any treatment. The treatment with co-culture conditioned medium did not increase both vimentin bands. The positive control HGF fibroblasts showed a strong band at 57 kD. E-cadherin was detected at 120 kD. SCC-25 cells at control conditions showed a strong 120 kD band, which was reduced after treatment with co-culture conditioned medium or with 1 ng/ml TGF-β1. Detroit 562 cells showed a light band at 120 kD, which was marginally reduced after treatment with co-culture conditioned medium or with 1 ng/ml TGF-β1 The control HG-fibroblasts did not not express protein at 120 kD with the vimentin antibody. Loading control was done using anti-GAPDH antibody.
Treatment with co-culture conditioned medium increased CDDP resistance of SCC-25 cells
Cell cultures were exposed to CDDP ranging from 0 µM to 50 µM and cell viability was assessed with MTT assays. Half-maximal inhibitory CDDP concentration was calculated using 4-parameter nonlinear logistic regression. The IC50 for native SCC-25 cells was 6.24 μM (95% CI 5.43 μM to 7.06 μM). Treatment with conditioned medium significantly increased CDDP-chemoresistance (Figure 3A). It more than doubled the IC50 of SCC-25 cells to 13.05 μM (95% CI 10.35 μM to 15.76 μM; p < 0.005). In addition to viability assays, clonogenic assays following exposure to 5 µM CDDP were performed in control and co-culture conditioned medium treated cells. In control cells, 5 µM CDDP reduced colony number from 3116±654 to 82±8 (91% reduction). In co-culture conditioned medium-treated cells, 5 µM CDDP reduced colony number from 1805±131 to 1106±381 (39% reduction). This means CDDP induced significant less (p = 0.013) reduction of colony numbers in co-culture cells than in standard-culture cells, suggesting an increased CDDP-chemoresistance following exposure to conditioned medium (Figure 4A-4C). In addition, SCC-25 cells were incubated with 1ng/ml TGF-β1 and CDDP to evaluate the specific effect of TGF-β1 on CDDP resistance (Figure 3A). The IC50 following TGF-β1 exposure (6.6 µM; 95% CI 5.47 µM to 7.73 µM) did not differ from the IC50 following standard medium (p > 0.2). TGF-β1 treatment did not alter clonogenity after treatment with 5 µM CDDP (95±11, p > 0.1, 89% reduction) compared to controls (Figure 4C).
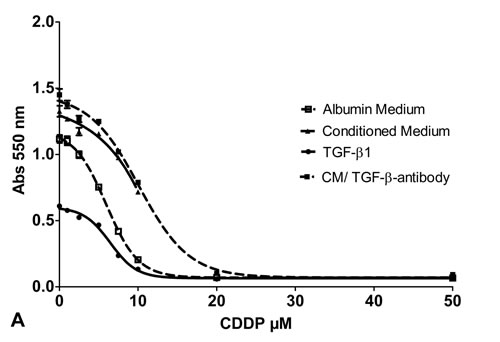
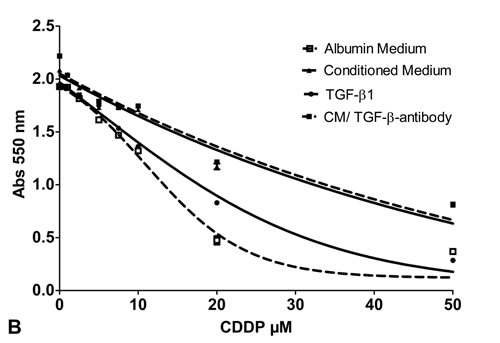
Figure 3: Cell viability assay: Cell viability of SCC-25 cells A. or Detroit 562 cells fat as well B. exposed to increasing doses of CDDP (0- 50 µM) following treatment with albumin containing medium (control; dotted line with white squares), co-culture conditioned medium (solid line with triangles), medium supplemented with TGF-β1 0.9 ng/ml (dotted line with spheres) and co-culture conditioned medium plus anti TGF-β antibody (1.5 µg/ml) (solid line with black squares). Four parameter nonlinear logistic regression model, whiskers indicate standard error of the mean (SEM).
To support the observation that TGF-β1 is not involved in co-culture conditioned medium induced chemoresistance, a neutralizing dose of anti-TGF-β (1.5 µg/ml) was added to the co-culture medium before viability and clonogenity assays were performed. Despite that, the chemoresistance mediating effect of co-culture conditioned medium was preserved. Also with anti-TGF-β, the IC50 of CDDP following administration of co-culture conditioned medium remained significantly higher than in control experiments (10.4 μM, 95% CI 9.2 to 11.5 μM (p < 0.05) (Figure 3A). The effect of co-culture conditioned medium on clonogenity after treatment with 5 μM was not altered by anti-TGF-β antibody treatment (1281±20, 13% reduction, p > 0.5) (Figure 4C).
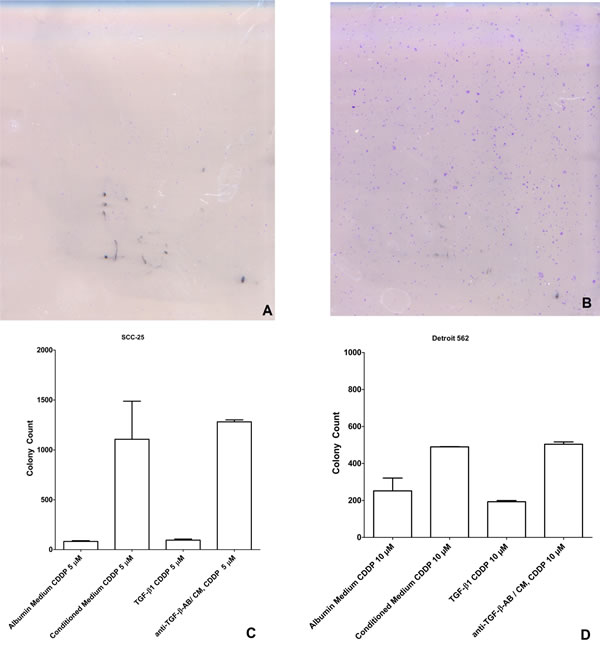
Figure 4: Clonogenic assay after treatment with albumin medium/ conditioned medium and CDDP 0 µM, 5 µM, 10 µM: A. Picture of the clonogenic assay of the control SCC-25 cells after treatment with 5 µM CDDP. B. Picture of the clonogenic assay of the SCC-25 cells cultured in co-culture conditioned medium after treatment with 5 µM CDDP. C. Number of colonies as a function of treatment. This demonstrates the significant increased colony forming ability of the SCC-25 cells after treatment with co-culture conditioned medium or co-culture conditioned medium/ anti-TGF-β-antibody (1.5 µg/ml) and CDDP 5 µM compared to controls or cells treated with TGF-β1 1ng/ml. D. Number of colonies as a function of treatment. This demonstrates the significant increased colony forming ability of the Detroit 562 cells after treatment with co-culture conditioned medium or co-culture conditioned medium/ anti-TGF-β-antibody (1.5 µg/ml) and CDDP 10 µM compared to the controls or cells treated with TGF-β1 1ng/ml.
Treatment with co-culture conditioned medium increased CDDP resistance of detroit 562 cells
The same experiments were repeated in Detroit 562 cells. The IC50 of CDDP for native Detroit 562 cells was 13.1 μM (95% CI 9.8 to 16.5 μM). Treatment with co-culture conditioned medium more than doubled the IC50 to 26.8 μM (95% CI 12.7 to 41.0 μM) (p < 0.01) (Figure 3B). In control cells 10 μM CDDP reduced the colony number to 252±69 (73% reduction). In co-culture conditioned medium treated cells 10 μM CDDP reduced the colony number to 490±1 (50% reduction) (p < 0.05) (Figure 4D). Treatment with 1 ng/ml TGF-β1 had no influence on the IC50 of CDDP in Detroit 562 cells, the IC50 remained stable with 16.9 μM (95% CI 14.9 to 19 μM, p > 0.05) (Figure 3B). TGF-β1 treatment did not alter clonogenity after treatment with 10 µM CDDP (193±6, p > 0.3) compared to controls cells (Figure 4D). Accordingly, treatment with co-culture conditioned medium containing a neutralizing dose of anti-TGF-β1-antibody (1.5 µg/ml) did not change the IC50 of CDDP in Detroit 562 cells compared to co-culture conditioned medium treated Detroit 562 cells (IC50 27.5 μM, 95% CI 18.5 to 36.3 μM, p > 0.05) (Figure 3B). Treatment with anti-TGF-ß-antibody/co-culture conditioned medium and 10 µM CDDP did not alter the effect of conditioned medium on clonogenity of Detroit 562 cells (504±6, p > 0.05) (Figure 4D) (Table 2).
Table 2: Treatment with co-culture conditioned medium resulted in increased CDDP resistance (measured with cell viability and clonogenity) in both cell lines
Cell type |
Parameter |
Cond. Medium |
TGF-ß1 |
Cond.Medium + anti TGF-ß |
SCC-25 |
IC50 |
increased *** |
no change |
increased*** |
Clonogenity after 5µM CDDP |
increased** |
no change |
increased*** |
|
Detroit 562 |
IC50 in µM |
increased** |
no change |
increased** |
Clonogenity after 10µM CDDP |
increased* |
no change |
increased*** |
TGF-ß1 treatment did not influence CDDP resistance. A neutralizing assay performed with anti-TGF-ß antibody supported this concept because it did not alter the CDDP resistance induced by co-culture conditioned medium.
* p < 0.05, ** p < 0.01, *** p < 0.001.
Discussion
EMT is a reversible cellular process caused by the interaction of epithelial tumor cells and tumor-associated fibroblasts. During EMT, epithelial tumor cells acquire characteristics of a mesenchymal phenotype. It is thought to be a relevant mechanism of cancer progression [5, 6]. In this study, we were interested in the effect of EMT on CDDP resistance of HNSCC tumor cells in vitro. Co-cultures of epithelial and mesenchymal cells are frequently used to model EMT in vitro. Here we used a direct fibroblast-tumor cell co-culture conditioned medium to induce EMT in the two human papilloma virus-negative HNSCC cell lines SCC-25 and Detroit 562.
Co-culture conditioned medium induced EMT and increased cell viability
Co-culture conditioned medium upregulated vimentin expression and downregulated E-cadherin expression in SCC-25 cells. As expected for an epithelial cell line, baseline vimentin expression was low and baseline E-cadherin expression was high. Similar phenotypic changes were induced by treatment with 1 ng/ml
TGF-β1. This TGF-β1 concentration has previously been measured in co-culture conditioned medium using enzyme-linked immunoassays (data not shown). TGF-β1 is considered a major factor for phenotypic changes in EMT [15].
Especially 46 kD vimentin protein levels increased in SCC-25 cells after treatment with co-culture conditioned medium and TGF-β1. 46 kD vimentin had been previously identified in epithelial cells under stress conditions [16]. On the protein level, co-culture conditioned medium and TGF-β1 slightly reduced the E-cadherin expression. The fact that EMT can be induced by cell free medium supports the concept that EMT mainly depends on paracrine signaling [8, 17, 18]. Beyond upregulation of mesenchymal and downregulation of epithelial markers, the co-culture conditioned medium increased cell viability in SCC 25 cells (p < 0.01). EMT-associated increase of cell viability is in line with several recent reports [4, 19].
This effect was not mediated by TGF-β1, since TGF-β1 reduced cell viability in SCC-25 cells (p < 0.001). It was demonstrated that Smad 4 is active in SCC-25 cells, which might enable a tumor suppressive effect of TGF-β1 [20]. Apparently, other factors than TGF-β1 are responsible for increased viability in SCC-25 cells exposed to co-culture conditioned medium and these factors are even able to override TGF-β1 induced viability reduction in SCC-25 cells. Moreover, the discrepancy of co-culture conditioned medium and TGF-β1 induced effects on EMT and cell viability implies that observed changes in cell viability are not caused by the acquisition of a mesenchymal phenotype (EMT), but rather an EMT associated effect of epithelial-mesenchymal crosstalk in the tumor microenvironment.
Despite lack of phenotypic changes, co-culture conditioned medium increased cell viability of Detroit 562 cells (p = 0.001), supporting the concept that EMT-induction and changes in cell viability may be caused by different factors. Consistent with the observations in SCC-25 cells, TGF-β1 had no influence on cell viability and clonogenity in Detroit 562 cells.
Co-culture conditioned medium induced CDDP chemoresistance
The main goal of this study was to determine if EMT is associated with increased CDDP chemoresistance. Medium from an epithelial-mesenchymal co-culture more than doubled the IC50 of CDDP in MTT assays in both cell lines (p < 0.001) (Figure 2). These results are consistent with the outcome of clonogenic assays. The anti-clonogenic effect of CDDP was significantly reduced after pretreatment with co-culture conditioned medium (p < 0.05) (Table 2). Hazlehurst et al. observed in 2001 that tumor microenvironment is involved in cell cycle arrest and drug resistance. This so called microenvironment related drug resistance is mediated through survival pathways activated as a result of cell-cell-adhesion (cell-adhesion mediated drug resistance) and extracellular matrix derived adhesive signals, which reduce the cytotoxic effect of CDDP [21]. Niessner et al. reported that the release of paracrine signaling factors produced by carcinoma-associated fibroblasts stimulate survival pathways such as the Akt-pathway and reduce cytotoxic effects of chemotherapies [22]. We treated HNSCC cell lines with a cell-free conditioned medium, suggesting the latter being responsible for the increase in CDDP resistance in our experimental setting. Similar results have been demonstrated in lung cancer cells. In these cells treatment with fibroblast-conditioned medium increased Paclitaxel resistance by restraining Paclitaxel induced apoptosis by activation of both extracellular signal-regulated kinases (Erk) 1/2 and Akt kinase. In this study, treatment with fibroblast-conditioned medium did not alter the CDDP resistance of lung cancer cells in vitro [23].
No effect of TGF-β1 on CDDP sensitivity
TGF-β1 did not increase the IC50 of CDDP of HNSCC cells in vitro. The IC50 of control and TGF-β1 treated cells was remarkably similar. Consistently, treatment with TGF-β1 had no influence on colony forming ability following CDDP-treatment. To support this observation, an additional experiment with co-culture conditioned medium supplemented with a neutralizing anti-TGF-β antibody was performed. Anti-TGF-β did not antagonize chemoresistance induction by co-culture conditioned medium in both cell lines. This supports the concept that increased CDDP-resistance is not caused, but merely associated with the acquisition of a mesenchymal phenotype. The factors leading to increased CDDP resistance following epithelial-mesenchymal interaction are not yet identified, but it doesn’t seem to be TGF-β1. Gilbert et al. reported that IL-6 (Interleukin-6) and TIMP (Tissue Inhibitor of metalloproteinase) could be possible tumor stroma-derived factors, which promote the survival of cancer cells [24]. Other fibroblast-produced mediators which might mediate EMT-associated chemoresistance, are HGF (human hepatocyte growth factor), MED12 (mediator complex subunit 12) and PGE-2 (Prostaglandine-E2) [15].
Materials and Methods
Cell lines
Human gingival fibroblasts (HGF) cells were purchased from Cell Line Service, Eppelheim, Germany [1, 25]. They were cultured in DMEM-low glucose (PAA, Pasching, Austria) supplemented with 10% foetal bovine serum (FBS) (PAA), 2 mM l-glutamine, 100 units/ml penicillin, and 100 μg/ml streptomycin. SCC-25 were purchased from German Collection of Microorganisms (DSMZ, Braunschweig, Germany) and Detroit 562 cells from the Cell Lines Service (Eppelheim, Germany), and were cultured in DMEM/F12 (PAA) supplemented with 10% FBS (PAA), 2 mM l-glutamine, 100 units/ml penicillin, and 100 μg/ml streptomycin [1].
Co-culture conditioned medium
For the production of co-culture conditioned medium, 4 x 104 /ml SCC-25 or Detroit 562 cells and 1 x 104 HGF cells/ml were plated in 250 ml cell culture flasks and cultured for 72 hours in 15 ml foetal bovine serum-containing medium (1:1 mix of DMEM/F12 (PAA) and DMEM-low glucose (PAA) supplemented with 10% foetal bovine serum (FBS) (PAA), 2 mM l-glutamine, 100 units/ml penicillin, and 100 μg/ml streptomycin). Then the cells were washed twice with Dulbecco’s Phosphate-Buffered Saline (DPBS) (Biowhittaker®, Belgium) and the serum-containing medium was replaced by 15 ml albumin-containing medium (7,5 ml DMEM/F12 (PAA) and 7,5 ml DMEM-low glucose (PAA) supplemented with bovine serum albumin (BSA, PAA) (0.4 g albumin/ 100 ml medium) replacing the protein content of 10% FBS, 2 mM l-glutamine, 100 units/ml penicillin, and 100 μg/ml streptomycin). Albumin-containing medium was left 48 hours on the co-culture allowing interacting epithelial cells and fibroblasts to secrete EMT-related factors into the medium. Afterwards, the co-culture conditioned medium was collected and cells were counted. The co-culture conditioned medium was portioned according to cell numbers as described by Hassona et al. [18]. The co-culture conditioned medium was sterile-filtered and stored at -80°C.
Stimulation of SCC-25/ Detroit 562 cells with co-culture conditioned medium and TGF-β1
To induce EMT in the first experimental arm, SCC-25/ Detroit 562 cells were treated with 7 ml co-culture conditioned medium per 50 ml cell culture flask for 72 hours. The medium was changed daily. To assess the effects of TGF-β1 in the second experimental arm, SCC-25/ Detroit 562 cells were cultivated in albumin-containing medium supplemented with TGF-β1 1 ng/ml (R&D Systems®, Minneapolis, US). Exposure conditions were the same as in the first experimental group, i.e., TGF-β1 supplemented medium was used over a period of 72 hours and media were changed daily. A TGF-β1 neutralizing assay was performed in the third experimental arm. SCC-25 cells and Detroit 562 were therefore treated with anti- TGF-β 1,-2,-3 antibody (R&D SystemsTM, Biomedica, Vienna; Austria) 1.5 µg/ml medium, as neutralizing dose assayed by the provider and 7 ml co-culture conditioned medium per 50 ml cell culture flask for 72 hours. Exposure conditions were the same as in the other experimental groups. In the fourth experimental arm, standard medium was used with identical media changes. At the end of the stimulation period, cells were used for RNA extraction, protein isolation, MTT assays and clonogenic assays, respectively.
RNA extraction, reverse transcription and real-time RT-PCR
A fraction of SCC-25/ Detroit 562 cells was harvested and RNA was isolated using TRIzol® reagent following the manufacturer’s instructions (Ambion®, Life technologiesTM, Thermo Fisher Scientific Inc., Waltham, MA, USA). RNA concentrations were determined by photometric measurements (BioPhotometer plus 6132, Eppendorf, Germany). Total RNA was reverse transcribed by M-MuLV Reverse Transcriptase (GeneON, Ludwigshafen, Germany) according to the manufacturer’s instructions in a MyiQTM cycler (BIO-RAD Laboratories, Inc., US). Real-Time quantitative PCR (qPCR) of copy-DNA transcripts was performed in a MyiQTM cycler (BIO-RAD Laboratories Inc., Hercules, CA, US) using iTaqTM Universal SYBR® Green Supermix (BIO-RAD Laboratories, Inc., Hercules, CA, US). β-Actin primers were purchased from InvitrogenTM (Darmstadt, Germany), while E-cadherin and vimentin primers were provided by Eurofins MWG Operon, Inc. (Ebersberg, Germany). β-Actin functioned well as a housekeeping gene and did not show significant changes across the three treatment conditions. Moreover, the size of the Real-Time PCR products was confirmed by agarose gel electrophoresis analysis.
Protein isolation and western blotting
Following treatments SCC-25 and Detroit 562 cells were washed twice with DPBS and collected with Accutase (PAA), followed by cell counting and harvesting by centrifugation at 290 g, 5 minutes, 4 °C. The resulting cell pellets were resuspended in 100 µl RIPA-buffer ( 50 mM Tris HCl/pH:8.0, 2 mM EDTA, 1 mM EGTA, 1 % Triton X-100, 0.25 % sodium deoxycholate, 0.1 % sodium dodecylsulfate, 150 mM NaCl, 10 mM NaF, 1 mM PMSF) / 106 cells. The cell suspension was vortexed and incubated 3-times for 15 minutes on ice, homogenized in 22G needles and centrifuged at 15000 g, 15 minutes, 4 °C. The cleared supernatant was subjected to protein concentration measurement using the Pierce 660 nm protein assay (Pierce, Rochford, IL, USA) according to the instructions of the manufacturer. 15 µg protein from all samples was subsequently processed for western blot analysis published previously [17], using primary antibodies: mouse monoclonal anti-vimentin (clone VI-10) IgM (Exbio, Prague, Czech Republic) at 1:200, or mouse monoclonal anti-E-cadherin (clone 36) IgG2a (BD Transduction Laboratories, BD Austria, Vienna, Austria) at 1: 2500, or anti-GAPDH (clone 6C5) IgG1 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 1: 200. For signal detection horseradish peroxidase coupled matched secondary antibodies and chemiluminescent substrate of Thermo Fisher Scientific (Vienna, Austria) were used in conditions suggested by the manufacturer. The chemilumescence signal was imaged by an Azure C500 documentation system (Biomedica, Vienna, Austria).
Treatment of the cells with CDDP and IC50 determination
Following stimulation SCC-25/ Detroit 562 cells were treated with increasing doses of CDDP (0 μM, 1 μM, 2.5 μM, 5 μM, 7.5 μM, 10 μM, 20 μM, 50 μM, 100 μM) for three days. The medium was changed daily. Cell viability was plotted against CDDP-concentration and IC50 was calculated employing a four-parameter nonlinear regression model [26]. For the clonogenic assays, 2 × 104
cells were plated in 250ml cell culture flasks. After stimulation they were treated with 5 μM CDDP for three days. The media were changed daily.
MTT- assay
Cell viability was evaluated by MTT-assays using the tetrazolium salt method. The MTT-assay is a quantitative colorimetric method used to determine metabolic activity [27]. After three days of CDDP treatment, 10 μl of 5 mg/ml MTT salt (in DMEM/F12 (PAA) supplemented with 10% FBS (PAA), 2 mM l-glutamine, 100 units/ml penicillin, and 100 μg/ml streptomycin) was administered to the cells (100 μl). Cells were incubated for 4h at 37°C and then the formazan reaction product was dissolved using 10% sodium dodecylsulphate in 10 mM HCl at 37°C for 12 hours. Absorbance at 550 nm was measured with a microtiter plate reader (Athos 2010, Salzburg, Austria). The MTT-tests were performed in four independent sets containing at least six biological repeats.
Clonogenic assay
For analysis of the anti-clonogenic effect of CDDP on the tumor cells we used the modified clonogenic assay described by Phuk and coworkers [14]. SCC-25/ Detroit 562 cells were washed with PBS and cultured in 250 ml tissue culture flasks in DMEM/F12 (PAA) supplemented with 10% FBS (PAA), 2 mM l-glutamine, 100 units/ml penicillin, and 100 μg/ml streptomycin for 14 days. After 14 days, cultures were fixed and stained in 0.5% gentian violet dissolved in methanol. Subsequently, the stained flasks were scanned in 1200 dpi resolution using a commercial flatbed scanner. Based on the resulting micrographs, colonies were counted and occupied areas were measured semi-automatically using a macro written in imageJ/FIJI macro language [28]. Background subtraction was performed on single images using rolling-ball-algorithm [29]. Micrographs were subjected to color deconvolution [30] and filtered using a Fourier band-pass filter. Colonies were segmented using auto-thresholding algorithms [31-35]. Segmented colonies were counted and occupied areas were measured. The clonogenic assays were performed in three independent sets.
Data analysis
Data were presented as mean +/- standard deviation (SD) unless indicated otherwise. The IC50 +/- 95% confidence intervals (CI) of CDDP were calculated with four-parameter nonlinear logistic regression using CurveExpert Professional (Daniel Hyams, Hixson, TN, USA). The results of real time PCR analysis were analyzed with GraphPad Prism 4.03 (GraphPad Software Inc, San Diego, CA, USA). Mean values among groups were compared with unpaired t-tests. MTT-changes in co-culture conditioned medium treated cells vs. controls were tested with unpaired t-test. For evaluation of clonogenic assays, a two-factorial analysis of variance with colony number as the response parameter was used. Cell culture medium (co-culture vs. standard) and CCDP exposure (0 µM vs 5 µM or 10 µM) served as factors. The interaction term served to indicate cell-culture mediated differences in CDDP sensitivity. For this data analysis, SPSS 22 was used (IBM Corporation, Armonk, NY, USA)
Conflicts of Interest
The authors declare that they have no competing interests
Grant Support
This work was supported by Austrian Science Fund [FWF P 25869-B13]. Role of the Funding Source: The funding source has no influence on the direction and the outcome of the study, it is an independent granting.
References
1. Dudas J, Bitsche M, Schartinger V, Falkeis C, Sprinzl GM, Riechelmann H. Fibroblasts produce brain-derived neurotrophic factor and induce mesenchymal transition of oral tumor cells. Oral oncology. 2011;47:98-103.
2. Radisky DC. Fibroblasts act as co-conspirators for chemotherapy resistance. Cancer biology & therapy. 2008;7:1348-9.
3. Smith A, Teknos TN, Pan Q. Epithelial to mesenchymal transition in head and neck squamous cell carcinoma. Oral oncology. 2013;49:287-92.
4. Orimo A, Weinberg RA. Stromal fibroblasts in cancer: a novel tumor-promoting cell type. Cell cycle. 2006;5:1597-601.
5. Fullar A, Kovalszky I, Bitsche M, Romani A, Schartinger VH, Sprinzl GM, Riechelmann H, Dudas J. Tumor cell and carcinoma-associated fibroblast interaction regulates matrix metalloproteinases and their inhibitors in oral squamous cell carcinoma. Experimental cell research. 2012;318:1517-27.
6. Xu LN, Xu BN, Cai J, Yang JB, Lin N. Tumor-associated fibroblast-conditioned medium promotes tumor cell proliferation and angiogenesis. Genetics and molecular research : GMR. 2013;12:5863-71.
7. Katsuno Y, Lamouille S, Derynck R. TGF-beta signaling and epithelial-mesenchymal transition in cancer progression. Current opinion in oncology. 2013;25:76-84.
8. Dudas J, Fullar A, Romani A, Pritz C, Kovalszky I, Hans Schartinger V, Sprinzl GM, Riechelmann H. Curcumin targets fibroblast-tumor cell interactions in oral squamous cell carcinoma. Experimental cell research. 2013;319:800-9.
9. Rheinwald JG, Beckett MA. Tumorigenic keratinocyte lines requiring anchorage and fibroblast support cultured from human squamous cell carcinomas. Cancer research. 1981;41:1657-63.
10. Chen YW, Lin GJ, Chia WT, Lin CK, Chuang YP, Sytwu HK. Triptolide exerts anti-tumor effect on oral cancer and KB cells in vitro and in vivo. Oral oncology. 2009;45:562-8.
11. Sano D, Xie TX, Ow TJ, Zhao M, Pickering CR, Zhou G, Sandulache VC, Wheeler DA, Gibbs RA, Caulin C, Myers JN. Disruptive TP53 mutation is associated with aggressive disease characteristics in an orthotopic murine model of oral tongue cancer. Clinical cancer research. 2011;17:6658-70.
12. Peterson WD, Jr., Stulberg CS, Simpson WF. A permanent heteroploid human cell line with type B glucose-6-phosphate dehydrogenase. Proceedings of the Society for Experimental Biology and Medicine Society for Experimental Biology and Medicine. 1971;136:1187-91.
13. Peterson WD, Jr., Stulberg CS, Swanborg NK, Robinson AR. Glucose-6-phosphate dehydrogenase isoenzymes in human cell cultures determined by sucrose-agar gel and cellulose acetate zymograms. Proceedings of the Society for Experimental Biology and Medicine Society for Experimental Biology and Medicine. 1968;128:772-6.
14. Puck TT, Marcus PI. Action of x-rays on mammalian cells. The Journal of experimental medicine. 1956;103:653-66.
15. Paraiso KH, Smalley KS. Fibroblast-mediated drug resistance in cancer. Biochemical pharmacology. 2013;85:1033-41.
16. Buchmaier BS, Bibi A, Muller GA, Dihazi GH, Eltoweissy M, Kruegel J, Dihazi H. Renal cells express different forms of vimentin: the independent expression alteration of these forms is important in cell resistance to osmotic stress and apoptosis. PloS one. 2013;8:e68301.
17. Dudas J, Fullar A, Bitsche M, Schartinger V, Kovalszky I, Sprinzl GM, Riechelmann H. Tumor-produced, active interleukin-1beta regulates gene expression in carcinoma-associated fibroblasts. Experimental cell research. 2011;317:2222-9.
18. Hassona Y, Cirillo N, Heesom K, Parkinson EK, Prime SS. Senescent cancer-associated fibroblasts secrete active MMP-2 that promotes keratinocyte dis-cohesion and invasion. British journal of cancer. 2014;111:1230-7.
19. Liu Y, Zhang M, Qian J, Bao M, Meng X, Zhang S, Zhang L, Zhao R, Li S, Cao Q, Li P, Ju X, Lu Q, et al. miR-134 Functions as a Tumor Suppressor in Cell Proliferation and Epithelial-to-Mesenchymal Transition by Targeting KRAS in Renal Cell Carcinoma Cells. DNA and cell biology. 2015.
20. Martin D, Abba MC, Molinolo AA, Vitale-Cross L, Wang Z, Zaida M, Delic NC, Samuels Y, Lyons JG, Gutkind JS. The head and neck cancer cell oncogenome: a platform for the development of precision molecular therapies. Oncotarget. 2014;5:8906-23.
21. Flach EH, Rebecca VW, Herlyn M, Smalley KS, Anderson AR. Fibroblasts contribute to melanoma tumor growth and drug resistance. Molecular pharmaceutics. 2011;8:2039-49.
22. Niessner H, Forschner A, Klumpp B, Honegger JB, Witte M, Bornemann A, Dummer R, Adam A, Bauer J, Tabatabai G, Flaherty K, Sinnberg T, Beck D et al. Targeting hyperactivation of the AKT survival pathway to overcome therapy resistance of melanoma brain metastases. Cancer medicine. 2013;2:76-85.
23. Bartling B, Hofmann HS, Silber RE, Simm A. Differential impact of fibroblasts on the efficient cell death of lung cancer cells induced by paclitaxel and cisplatin. Cancer biology & therapy. 2008;7:1250-61.
24. Gilbert LA, Hemann MT. DNA damage-mediated induction of a chemoresistant niche. Cell. 2010;143:355-66.
25. Docheva D, Padula D, Popov C, Weishaupt P, Pragert M, Miosge N, Hickel R., Bocker W., Clausen-Schaumann H., Schieker M. Establishment of immortalized periodontal ligament progenitor cell line and its behavioural analysis on smooth and rough titanium surface. European cells & materials. 2010;19:228-41.
26. Goutelle S, Maurin M, Rougier F, Barbaut X, Bourguignon L, Ducher M, Maire P. The Hill equation: a review of its capabilities in pharmacological modelling. Fundamental & clinical pharmacology. 2008;22:633-48.
27. Schartinger VH, Galvan O, Riechelmann H, Dudas J. Differential responses of fibroblasts, non-neoplastic epithelial cells, and oral carcinoma cells to low-level laser therapy. Supportive care in cancer. 2012;20:523-9.
28. Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, Tinevez JY, White DJ, Hartenstein V, et al. Fiji: an open-source platform for biological-image analysis. Nature methods. 2012;9:676-82.
29. Sternberg B. Exploring the potential of computer-assisted medical illustration. Biomedical communications. 1983;11:28-31.
30. Ruifrok AC, Johnston DA. Quantification of histochemical staining by color deconvolution. Analytical and quantitative cytology and histology / the International Academy of Cytology [and] American Society of Cytology. 2001;23:291-9.
31. Zack GW, Rogers WE, Latt SA. Automatic measurement of sister chromatid exchange frequency. The journal of histochemistry and cytochemistry. 1977;25:741-53.
32. Li CH, Lee C.K. . Minimum Croos Entropy Thresholding. Pattern Recognition. 1993;26:617-25.
33. Kapur JN, Sahoo PK, Wong AKC. A new method for gray-level picture thresholding using the entropy of the histogram. Computer Vision, Graphics, and Image Processing. 1985;29:273-85.
34. N. O. A treshold Selection Method from Gray-Level Histograms. IEEE Transactions on Systems, Man, and Cybernetics. 1979;1:62-6.
35. Shanbhag AG. Utilization of Information Measure as a Means of Image Thresholding. CVGIP: Graphical Models and Image Processing. 1994;56:414-9.