
INTRODUCTION
The extended lifespan of the Ames and Snell dwarf mice is attributed to the attenuation of the insulin/IGF-1 signaling pathways [1, 2]. In these mice GH deficiency reduces production and circulating levels of insulin and IGF-1 [1, 3]. Reduced IGF-1 signaling affects insulin sensitivity suggesting that crosstalk occurs between IGF-1 and insulin signaling [4].
Control of mammalian aging by IGF-1is based on the increased longevity of hypopituitary growth hormone (GH)-deficient mice in which reduced IGF-1 expression and peripheral levels are characteristics of increased lifespan [5-7]. Subsequent studies of mice heterozygous for the IGF-1R [IGF-1R(+/-)] provided direct evidence that IGF-1 plays a role in controlling mouse longevity [8, 9]. Low levels of circulating IGF-1 are, therefore, a common feature of several long-lived mouse models (Ames, Snell, and IGF-1R(+/-)). Decreased mRNA and protein pool levels of the hepatic IGF-1 axis in Ames mice suggest that the regulation of genes targeted by the insulin/IGF-1-signaling pathway may contribute to physiological conditions supporting longevity [7]. Thus, in the Ames mouse, IGF-1 also regulates the insulin signaling pathway suggesting the involvement of insulin/IGF-1 crosstalk interactions.
Phosphorylation of serine/threonine (Ser/Thr) or tyrosine (Tyr) residues of the insulin receptor substrate (IRS) proteins regulate insulin signaling [10]. Phosphorylation of the IRS Ser residues inhibits Tyr phosphorylation thereby serving as a physiological negative-feedback control mechanism [11]. Insulin stimulated Ser phosphorylation observed in hyperglycemia [12] or in response to proinflammatory cytokines [13-16] suggests this as the mechanism of acute and chronic stress mediated insulin resistance [17]. Thus, the stimulation of Ser/Thr phosphorylation of IRS-1 (and IRS-2), impairs its association with the insulin receptor (IR) thereby inhibiting insulin-stimulated Tyr-phosphorylation of both IRS-1 and IR [13, 18-21]. Control of IRS-1 signaling is thus achieved by the differential phosphorylation of Ser/Thr and Tyr residues. These phosphorylations are part of the physiological processes of longevity determination as well as the development of insulin and IGF-1 resistance.
Phosphorylation of IRS-1 on Ser307. Ser612, Ser636/639, and Ser1101 negatively regulate several functions of IRS-1 which include: a) phosphorylation of Ser309 which uncouples IRS-1 from the insulin receptor (IR); decreases tyrosine phosphorylation and increases degradation of the IR; b) phosphorylation of Ser612 and Ser636/639 reduces the IRS-1/PI3-kinase association [11].
Although insulin and IGF-1 signaling are initiated by specific receptors there is considerable crosstalk between these pathways [4, 14, 22]. This raises the question of whether insulin/IGF-1 crosstalk involves the phosphorylation of the same IRS-1 Ser residues. By this mechanism, insulin and IGF-1 crosstalk could regulate longevity and the development of insulin and IGF-1 resistance [13].
In past studies we demonstrated that fibroblast cultures derived from young and aged Ames dwarf mice maintain their in vivo characteristics of resistance to mitochondrial generated oxidative stress [23]. Based on these observations we used these cells to address the question of whether: (a) IGF-1 stimulates the phosphorylation of the same IRS-1 Ser residues that are targeted by insulin; (b) the levels of Ser phosphorylation differ in WT vs. Ames dwarf fibroblasts; and (c) aging affects the levels and pattern of IGF-1 stimulated Ser phosphorylations.
We propose that the results of our experiments would provide information on the mechanism by which IGF-1 participates in the regulation of insulin-GH signaling and the determination of longevity.
RESULTS
Multiple physiological functions, including longevity determination and insulin/IGF-1 resistance are regulated by IRS-1signaling involving phosphorylation-dephosphorylation of numerous Ser/Thr and Tyr residues. In these studies we examined whether IGF-1 stimulates the phosphorylation of IRS-1 Ser307, Ser612, Ser636/639 and Ser1101 residues that are known to be phosphorylated in response to insulin treatment, whether these IGF-1 stimulated phosphorylations in Ames fibroblasts are consistent with decreased insulin/IGF-1 pathway activity associated with longevity.
IGF-1 stimulates IRS-1 Ser307 phosphorylation in young and aged wild type, and aged Ames dwarf fibroblasts
The longevity of Ames dwarf [Prop1(-/-)] mice is attributed to decreased levels of tissue and circulating insulin and IGF-1[1, 3] and the differential phosphorylation of IRS-1 Ser residues may be part of the mechanism that regulates the altered insulin/IGF-1 signaling that promotes longevity. To address this we examined whether IGF-1 stimulates phosphorylation of the same IRS-1 Ser residues that are insulin-stimulated. The responses by the young Ames fibroblasts and their age-matched WT controls to IGF-1 treatment indicate that Ser307 phosphorylation is stimulated in WT fibroblasts by ~6-fold at 15 minutes and ~10-fold at 30 minutes and remains elevated up to 60 minutes of treatment (Figure 1A and 1B). On the other hand phosphorylation of Ser307 is severely attenuated in the Ames-derived fibroblasts suggesting that these cells maintain the characteristic decreased levels of insulin/IGF-1 signaling associated with Ames longevity.
There is a ~14-fold increase in the endogenous level of IRS-1 Ser307 phosphorylation in aged WT and dwarf fibroblasts (Figure 1C, 1D). Furthermore treatment with IGF-1 did not further enhance phosphorylation of Ser307 in either the aged WT or Ames fibroblasts. Thus, in both cell types the elevated endogenous levels of Ser307 phosphorylation and the loss of response of IGF-1 suggest significantly altered physiological functions. The significant endogenous derepression of Ser307 phosphorylation in both WT and Ames untreated fibroblasts, and loss of response to IGF-1 are indicative of major alterations of metabolic functions.
Interestingly, the aged Ames fibroblasts show a decreased level of Ser307 phosphorylation at 15, 30, and 60 minutes after IGF-1 treatment suggesting that the phosphorylation is responsive whereas the WT fibroblasts are refractive in that they maintain the elevated level of Ser307 phosphorylation (Figure 1C and 1D). These results suggest that functions regulated and inducible by P-Ser307 persist in the WT controls, but are attenuated and transient in the Ames dwarf cells.
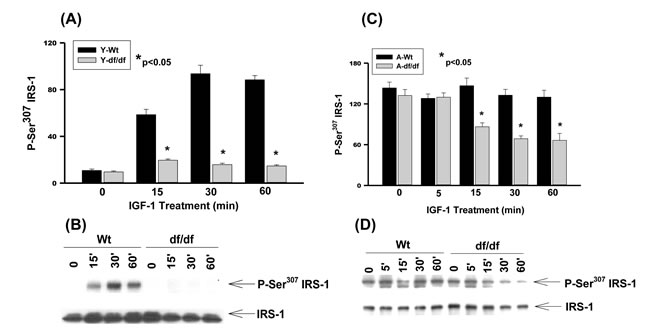
Figure 1: The IGF-1 stimulated phosphorylation of IRS-1 Ser307 in young and aged WT and Ames dwarf mouse tail fibroblasts. A. A bar graph and B. immunoblot analysis of IRS-1 Ser307 phosphorylation in fibroblasts from young (3-6 mos) WT and Ames dwarf mice. C. A bar graph and D. immunoblot analysis of the phosphorylation of IRS-1 Ser307 in fibroblasts from aged (21-24 mos) WT and Ames dwarf mice. Statistical analyses were performed using the two-tailed t-test to show the mean difference between age-matched groups at a significance level of 0.050. The symbols (*) indicate statistical significance for the values represented by the bar or time point.
IGF-1 stimulates IRS-1 Ser612 phosphorylation in young and aged wild type, and aged Ames dwarf fibroblasts
The G protein-coupled receptor kinase-2 (GRK2) is a Ser/Thr kinase that upon endothelin-1 (ET-1) stimulation associates with IRS-1 thereby promoting ET-1 mediated Ser612 phosphorylation and IRS-1 degradation [24]. GRK2 thus plays a role in chronic ET-1 induced insulin resistance by inhibiting IRS-1. Elevated GRK2 functions as a negative regulator of insulin action by interfering with G protein-q/11α subunit signaling. Thus, upon ET-1 activation, GRK2 associates with IRS-1 and promotes ET-1 mediated IRS-1 Ser612 phosphorylation and degradation which is associated with insulin resistance [24]. Furthermore, the phosphorylation of Ser612 is uniquely associated with ET-1 and GRK2 in myocardial ischemic injury [24, 25]; type 2 diabetes [26]. obesity and hypertension [26, 27].
Our results show that IGF-1 stimulates the phosphorylation of Ser612 in WT fibroblasts whereas the Ames fibroblasts do not respond to this treatment (Figure 2A and 2B). On the other hand the level of IGF-1 stimulated Ser612 phosphorylation in aged WT is similar to that of the young fibroblasts whereas phosphorylation of the Ames fibroblasts is significantly derepressed, although the level of expression is lower than that of the WT response (Figure 2C and 2D). The endogenous levels of Ser612 phosphorylation are thus increased in both aged WT and Ames fibroblasts and IGF-1 induces further phosphorylation in both cell types. There are, however, significant differences between WT and Ames fibroblasts in that (a) young Ames fibroblasts are resistant to IGF-1 stimulated phosphorylation and (b) the inducible level of Ser612 phosphorylation in the aged WT is similar to that of the young cells and c) the level of induction of the WT cells remains elevated for 60 minutes, whereas in the Ames cells it peaks in 5 minutes and declines rapidly (Figure 2C and 2D).
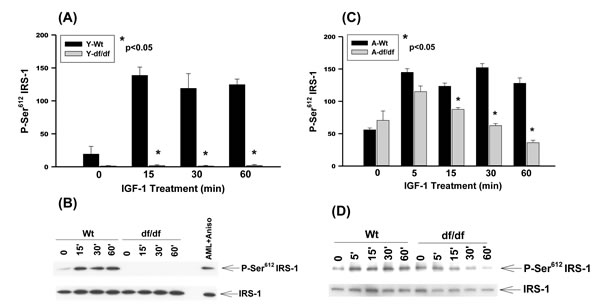
Figure 2: The IGF-1 stimulated phosphorylation of IRS-1 Ser612 in young and aged WT and Ames dwarf mouse tail fibroblasts. A. A bar graph and B. immunoblot analysis of IRS-1 Ser612 phosphorylation in fibroblasts from young (3-6 mos) WT and Ames dwarf mice. C. A bar graph and D. immunoblot analysis of the phosphorylation of IRS-1 Ser612 in fibroblasts derived from aged (21-24 mos) WT and Ames dwarf mice. Statistical analyses were performed using the two-tailed t-test as described in Figure 1 and in Methods.
IGF-1 stimulates Ser636/639 phosphorylation in young and aged WT and aged Ames dwarf fibroblasts
The Ser636/639 residues are involved in the regulation of protein synthesis (mTOR, S6K, ERK) and degradation [28]. Our data show a strong stimulation of Ser636/639 phosphorylation in young WT mice whereas it is virtually non-responsive in dwarf fibroblasts (Figure 3A and 3B). Thus, the Ames fibroblasts are refractory to IGF-1 stimulated phosphorylation of Ser636/639 which could result in the reduced mTOR/S6K1 signaling, attenuation of protein synthesis and decreased insulin resistance. This is a characteristic of longevity in models ranging from nematodes to mice. On the other hand Ser636/639 phosphorylation is strongly induced by IGF-1 in both aged WT and Ames fibroblasts which is a characteristic of insulin resistance (Figure 3C and 3D). The phosphorylation in the dwarf fibroblasts decreases significantly at 30 minutes and 60 minutes whereas such a decrease is delayed in WT cells. The sustained vs. transient phosphorylation of Ser636/639 is indicative of differences in metabolic characteristics such as protein synthesis and insulin resistance.
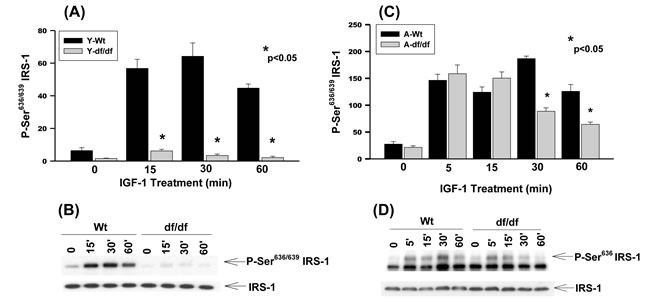
Figure 3: The IGF-1 stimulated phosphorylation of IRS-1 Ser636/639 in young and aged WT and Ames dwarf mouse tail fibroblasts. A. A bar graph and B. immunoblot analysis of IRS-1 Ser636/639 phosphorylation in fibroblasts derived from young (3-6 mos) WT and Ames dwarf mice. C. A bar graph and D. immunoblot analysis of the phosphorylation of IRS-1 Ser636/639 in fibroblasts derived from aged (21-24 mos) WT and Ames dwarf mice. Statistical analyses were performed using the two-tailed t-test as described in Figure 1 and in METHODS.
IGF-1 stimulates Ser1101 phosphorylation in young and aged wild type and aged Ames dwarf fibroblasts
Infusion of amino acids into humans leads to the activation of S6K1 phosphorylation of IRS-1 Ser1101, a reduction of IRS-1 function and insulin resistance in skeletal muscle [29]. Mutation of this site blocks the ability of amino acids to suppress IRS-1 Tyr and Akt phosphorylation [29]. Phosphorylation of IRS-1 Ser1101 is increased in livers of obese db/db and WT, but not S6K(-/-) mice maintained on a high protein diet. Nutrient dependent activation of S6K1 is thus associated with insulin resistance in mice and humans in part via Ser1101 phosphorylation [29].
Young WT fibroblasts exhibit a significant level of endogenous phosphorylation of Ser1101 and treatment with IGF-1 results in further increased phosphorylation at this site (Figure 4A, 4B). The peak of phosphorylation occurs ~15 minutes after treatment and decreases by ~60 minutes to the level of untreated cells suggesting a rapid response to and recovery from IGF-1 treatment in young WT fibroblasts.
The endogenous level of Ser1101 phosphorylation in young Ames mice is ~3-fold lower than the corresponding untreated WT cells (Figure 4A and 4B). Furthermore, IGF-1 treatment of the Ames cells showed a slight increase in Ser1101 phosphorylation at 15 minutes compared to the WT and at 30- and 60 minutes the level of phosphorylation decreases to the level of untreated cells.
Similar analyses of the aged WT and dwarf fibroblasts show that the endogenous levels of Ser1101 phosphorylation are significantly lower compared to the young cells, i.e., ~3-fold in WT and ~2-fold in dwarf cells (Figure 4C and 4D). The IGF-1 treated WT and dwarf cells showed a similar response after 5 and 15 minutes of treatment; at 30 minutes, however, the phosphorylation was ~5-fold higher than its endogenous level which is the same as that seen in the young WT fibroblasts. These data indicate that the induction of IGF-1 phosphorylation of Ser1101 is delayed in the WT aged cells. Furthermore, there is a rapid loss of P-Ser1101 by 60 minutes so that the level of phosphorylation by the WT fibroblasts is the same as in the Ames cells. On the other hand there is also a lag period in response to IGF-1 in the aged WT fibroblasts that does not occur in the young cells.
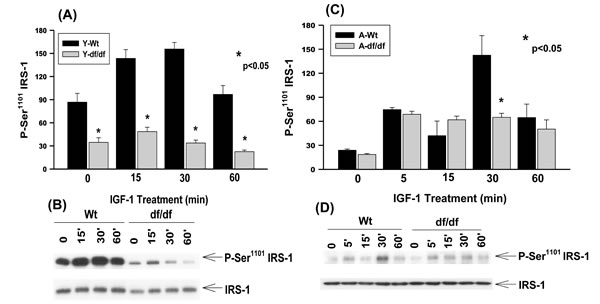
Figure 4: The IGF-1 stimulated phosphorylation of IRS-1 Ser1101 in young and aged WT and Ames dwarf mouse tail fibroblasts. A. A bar graph and B. immunoblot analysis of the phosphorylation of IRS-1 Ser1101 in fibroblasts derived from young (3-6 mos) WT and Ames dwarf mice. C. A bar graph and D. immunoblot analysis of the phosphorylation of IRS-1 Ser1101 in fibroblasts derived from aged (21-24 mos) WT and Ames dwarf mice. Statistical analyses were performed using the two-tailed t-test as described in Figure 1 and in METHODS.
DISCUSSION
Using genetically identical young and aged WT and Ames dwarf fibroblasts we have identified specific Ser residues of IRS-1 that are substrates for IGF-1 stimulated phosphorylations and are the same as those for insulin stimulated Ser phosphorylation. We thus propose that this may be a crosstalk mechanism by which circulating IGF-1 participates in the regulation of insulin sensitivity and plays an important role in the hormonal balance between GH and insulin thus enabling them to regulate the same metabolic pathways [4]. Importantly, the levels of endogenous and inducible phosphorylation of Ser307, Ser612, Ser636/639, and Ser1101 are significantly higher in the WT fibroblasts suggesting that these epigenetic alterations in the Ames mice control the unique serine phosphorylation patterns that determine the physiological conditions of IRS-1 signaling associated with longevity. We thus propose that epigenetic changes associated with the hormonal deficiencies of the Prop1(-/-) mutation are stabilized and maintained in fibroblasts derived from these mice [23, 30, 31]
The IGF-1 mediated phosphorylation of IRS-1 Ser307, Ser612, Ser636/639, and Ser1101 is attenuated in Ames cells. Since these phosphorylations regulate multiple signaling pathways and metabolic processes the overall reduced levels of phosphorylation will, for example, affect levels of protein synthesis, fatty acid metabolism, glucose metabolism and stress response activity characteristic of the long-lived Ames mice [30, 31]. This is consistent with the reported specificities of Ser phosphorylation that include both normal and disease-associated metabolic functions. An interesting example is the phosphorylation of Ser612 which is virtually absent in the fibroblasts from young Ames mice. Since phosphorylation of IRS-1 Ser612 is a physiological characteristic of cardiovascular pathology [25, 32], type 2 diabetes [26], obesity and hypertension [26, 27], our data suggest that the high levels of phosphorylation of Ser612 in young WT mice may promote cardiovascular pathology and also serve as a marker of this disease as well as of aging. Alternatively, the virtual absence of Ser612 phosphorylation in young Ames fibroblasts and rapid loss of P-Ser612-IRS-1 by aged Ames fibroblasts suggests a decreased risk of cardiovascular disease associated with longevity. The fact that phosphorylation levels are significantly elevated in fibroblasts from aged WT and dwarfs suggest that this increased endogenous activity and strong stimulation of phosphorylation may represent major age-associated changes in the IRS-1 targeted metabolic functions.
There is a significant difference between the induction of Ser307, Ser612, and Ser636/639 phosphorylation in aged WT vs. aged Ames fibroblasts. For example, the level of phosphorylation in aged WT fibroblasts remains elevated up to 60 minutes whereas the phosphorylation declines after 15-30 minutes in the age-matched Ames fibroblasts. Thus, the WT-specific, sustained and elevated level of phosphorylated IRS-1 Ser residues may be part of the mechanism of inhibition of IRS-1 activity associated with resistance to insulin/IRS-1. On the other hand, although the Ames fibroblasts respond strongly to IGF-1, the decrease of Ser phosphorylation suggests this rapid recovery may be the mechanism that protects against the consequences of IRS-1 hyperphosphorylation.
The insulin/IGF-1 and mammalian mTOR pathways are highly conserved physiological processes that protect against the development of the aging phenotype. The multiple physiological consequences of IRS-1 phosphorylations are seen in increased activation of the mTOR pathway along with enhanced phosphorylation of IRS-1 at Ser307 and Ser636/639 that occur in animal models of insulin resistance [30]. Thus, activation of the mTOR pathway leading to the down-regulation of IRS-mediated signals is a negative feedback mechanism which may be involved in the development of insulin resistance [33, 34]. Reduced insulin/IGF-1 signaling extends lifespan in both invertebrates and vertebrates [35, 36]. Thus, low levels of insulin and/or IGF-1 signaling (together with high insulin and IGF-1 sensitivity) are physiological characteristics that support extended lifespan of the Ames mutants [37]. That deletion of IRS-1 extends lifespan and increases resistance to several age-related pathologies in male and female mice is further support of this mechanism [9, 36]. These studies suggest that reduced IRS-1 dependent signaling is a robust mechanism for the modulation of mammalian lifespan.
It has been shown that the haploinsufficient IGF-1R(+/-) mouse down regulates the principle pathways stimulated by IGF-1 [8]. Embryonic fibroblasts derived from Igf1r(+/-)mice showed a 50% reduction in IGF-1R levels and a marked reduction in IGF-1 induced tyrosine phosphorylation of its substrate, IRS-1. This suggests that the down-regulation of IGF-1R in haploinsufficient mice down-regulates the principal pathways stimulated by IGF-1 and that the general decrease of IGF-1 via a decrease in IGF-1R can increase mammalian lifespan [8].
Phosphorylation of IRS-1 Ser307 exemplifies a common site that integrates heterologous inhibition of insulin signaling by multiple factors (See Ref.15 for a detailed Table). For example, Ser307 phosphorylation is stimulated by insulin, IGF-1, TNFα [38], anisomycin [36], fatty acids [39], amino acids [10] and C-reactive protein [40]. Our studies suggest that IGF-1 may act synergistically with these factors to promote Ser307 phosphorylation [41] and attenuation of IRS-1 and its targeted downstream pathways, e.g., PI3K and MAPK pathways [40, 42].
We propose that crosstalk between insulin and IGF-1 signaling is mediated by the phosphorylation/dephosphorylation of Ser/Thr and Tyr residues, and that this is the mechanism of regulation of the major pathways that target physiological functions associated with longevity determination.
materials and METHODS
Isolation and treatment of Ames mouse tail fibroblasts
The isolation of fibroblasts from young (3-6 mos) and aged (21-24 mos) wild-type and Ames dwarf mice has been described [23]. In this study, the fibroblasts at passage 4 or 5 were plated in 100 mm2 cell culture dishes (2x105 cells/dish) and cultured for 3 days in DMEM medium containing 15% FBS. The day before treatment, the medium was replaced with a medium containing 0.5% FBS. The recombinant human IGF-1 (Sigma) was prepared in distilled H2O and filter sterilized. The IGF-1 stock solution (25 ng/ml) was diluted in DMEM medium containing 0.5% FBS and the fibroblasts were treated with IGF-1 at a final concentration of 2 ng/ml. The cells were harvested and cytoplasmic and nuclear extracts were prepared [23]. The protease and phosphatase inhibitors were added to the extraction buffers prior to use [23]. The protein concentration of the extracts was determined using Bradford reagent (Bio-Rad).
Western blot analyses and immunoprecipitation assays
Western blot analyses and immunoprecipitation assays were performed as described [23]. Antibodies used for immunoblot analysis of phosphorylated proteins were from Cell Signaling Technology: anti-phospho Ser307 IRS-1, anti-phospho Ser612 IRS-1, anti-phospho Ser636/639 IRS-1 and anti-phospho Ser1101 IRS-1. The antibody was from Sigma.
Statistical analyses
Statistical analyses were performed for age-matched comparisons, the single dependent variable being the Ames dwarf mutants. The normalized values of protein and phosphorylated protein bands were analyzed using the 2-tailed t-test to test the mean difference between age-matched groups at a significance level of 0.05. The symbols (*) indicate statistical significance for the values represented by the bar or time-point.
ACKNOWLEDGMENTS
This publication was supported by USPHS grant 1P01 AG021830 awarded by the National Institute on Aging; by 1 P30 AG024832 Claude D. Pepper Older Americans Independence Center grant and by the Sealy Center on Aging. J.P. thanks the Bertha and Robert Bucksch Distinguished Research Professor of Aging endowment for their support of this study. We thank Dr. Karon P. Cassidy for her rigorous review of the manuscript.
CONFLICTS OF INTEREST
The authors have no conflicts of interest relevant to this manuscript.
REFERENCES
1. Bartke A. Minireview: role of the growth hormone/insulin-like growth factor system in mammalian aging. Endocrinology. 2005; 146:3718-3723.
2. Hsieh CC and Papaconstantinou J. The effect of aging on p38 signaling pathway activity in the mouse liver and in response to ROS generated by 3-nitropropionic acid. Mechanisms of ageing and development. 2002; 123:1423-1435.
3. Masternak MM, Panici JA, Wang F, Wang Z and Spong A. The effects of growth hormone (GH) treatment on GH and insulin/IGF-1 signaling in long-lived Ames dwarf mice. The journals of gerontology Series A, Biological sciences and medical sciences. 2010; 65:24-30.
4. Dominici FP, Argentino DP, Munoz MC, Miquet JG, Sotelo AI and Turyn D. Influence of the crosstalk between growth hormone and insulin signalling on the modulation of insulin sensitivity. Growth hormone & IGF research : official journal of the Growth Hormone Research Society and the International IGF Research Society. 2005; 15:324-336.
5. Coschigano KT, Clemmons D, Bellush LL and Kopchick JJ. Assessment of growth parameters and life span of GHR/BP gene-disrupted mice. Endocrinology. 2000; 141:2608-2613.
6. Flurkey K, Papaconstantinou J, Miller RA and Harrison DE. Lifespan extension and delayed immune and collagen aging in mutant mice with defects in growth hormone production. Proceedings of the National Academy of Sciences of the United States of America. 2001; 98:6736-6741.
7. Papaconstantinou J, Deford JH, Gerstner A, Hsieh CC, Boylston WH, Guigneaux MM, Flurkey K and Harrison DE. Hepatic gene and protein expression of primary components of the IGF-I axis in long lived Snell dwarf mice. Mechanisms of ageing and development. 2005; 126:692-704.
8. Holzenberger M, Dupont J, Ducos B, Leneuve P, Geloen A, Even PC, Cervera P and Le Bouc Y. IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature. 2003; 421:182-187.
9. Selman C, Lingard S, Choudhury AI, Batterham RL, Claret M, Clements M, Ramadani F, Okkenhaug K, Schuster E, Blanc E, Piper MD, Al-Qassab H, Speakman JR, et al. Evidence for lifespan extension and delayed age-related biomarkers in insulin receptor substrate 1 null mice. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 2008; 22:807-818.
10. Boura-Halfon S and Zick Y. Serine kinases of insulin receptor substrate proteins. Vitamins and hormones. 2009; 80:313-349.
11. Gual P, Le Marchand-Brustel Y and Tanti JF. Positive and negative regulation of insulin signaling through IRS-1 phosphorylation. Biochimie. 2005; 87:99-109.
12. Nakajima K, Yamauchi K, Shigematsu S, Ikeo S, Komatsu M, Aizawa T and Hashizume K. Selective attenuation of metabolic branch of insulin receptor down-signaling by high glucose in a hepatoma cell line, HepG2 cells. The Journal of biological chemistry. 2000; 275:20880-20886.
13. Kanety H, Feinstein R, Papa MZ, Hemi R and Karasik A. Tumor necrosis factor alpha-induced phosphorylation of insulin receptor substrate-1 (IRS-1). Possible mechanism for suppression of insulin-stimulated tyrosine phosphorylation of IRS-1. The Journal of biological chemistry. 1995; 270:23780-23784.
14. Liu YF, Herschkovitz A, Boura-Halfon S, Ronen D, Paz K, Leroith D and Zick Y. Serine phosphorylation proximal to its phosphotyrosine binding domain inhibits insulin receptor substrate 1 function and promotes insulin resistance. Molecular and cellular biology. 2004; 24:9668-9681.
15. Paz K, Hemi R, LeRoith D, Karasik A, Elhanany E, Kanety H and Zick Y. A molecular basis for insulin resistance. Elevated serine/threonine phosphorylation of IRS-1 and IRS-2 inhibits their binding to the juxtamembrane region of the insulin receptor and impairs their ability to undergo insulin-induced tyrosine phosphorylation. The Journal of biological chemistry. 1997; 272:29911-29918.
16. Venters HD, Tang Q, Liu Q, VanHoy RW, Dantzer R and Kelley KW. A new mechanism of neurodegeneration: a proinflammatory cytokine inhibits receptor signaling by a survival peptide. Proceedings of the National Academy of Sciences of the United States of America. 1999; 96:9879-9884.
17. Draznin B. Molecular mechanisms of insulin resistance: serine phosphorylation of insulin receptor substrate-1 and increased expression of p85alpha: the two sides of a coin. Diabetes. 2006; 55:2392-2397.
18. Hotamisligil GS, Peraldi P, Budavari A, Ellis R, White MF and Spiegelman BM. IRS-1-mediated inhibition of insulin receptor tyrosine kinase activity in TNF-alpha- and obesity-induced insulin resistance. Science. 1996; 271:665-668.
19. Peraldi P, Hotamisligil GS, Buurman WA, White MF and Spiegelman BM. Tumor necrosis factor (TNF)-alpha inhibits insulin signaling through stimulation of the p55 TNF receptor and activation of sphingomyelinase. The Journal of biological chemistry. 1996; 271:13018-13022.
20. Tanti JF, Gremeaux T, van Obberghen E and Le Marchand-Brustel Y. Serine/threonine phosphorylation of insulin receptor substrate 1 modulates insulin receptor signaling. The Journal of biological chemistry. 1994; 269:6051-6057.
21. Mothe I and Van Obberghen E. Phosphorylation of insulin receptor substrate-1 on multiple serine residues, 612, 632, 662, and 731, modulates insulin action. The Journal of biological chemistry. 1996; 271:11222-11227.
22. Papaconstantinou J. Insulin/IGF-1 and ROS signaling pathway cross-talk in aging and longevity determination. Molecular and cellular endocrinology. 2009; 299:89-100.
23. Hsieh CC and Papaconstantinou J. Dermal fibroblasts from long-lived Ames dwarf mice maintain their in vivo resistance to mitochondrial generated reactive oxygen species (ROS). Aging (Albany NY). 2009; 1:784-802.
24. Usui I, Imamura T, Babendure JL, Satoh H, Lu JC, Hupfeld CJ and Olefsky JM. G protein-coupled receptor kinase 2 mediates endothelin-1-induced insulin resistance via the inhibition of both Galphaq/11 and insulin receptor substrate-1 pathways in 3T3-L1 adipocytes. Molecular endocrinology. 2005; 19:2760-2768.
25. Ciccarelli M, Chuprun JK, Rengo G, Gao E, Wei Z, Peroutka RJ, Gold JI, Gumpert A, Chen M, Otis NJ, Dorn GW, 2nd, Trimarco B, Iaccarino G, et al. G protein-coupled receptor kinase 2 activity impairs cardiac glucose uptake and promotes insulin resistance after myocardial ischemia. Circulation. 2011; 123:1953-1962.
26. Takahashi K, Ghatei MA, Lam HC, O’Halloran DJ and Bloom SR. Elevated plasma endothelin in patients with diabetes mellitus. Diabetologia. 1990; 33:306-310.
27. Ferri C, Bellini C, Desideri G, Di Francesco L, Baldoncini R, Santucci A and De Mattia G. Plasma endothelin-1 levels in obese hypertensive and normotensive men. Diabetes. 1995; 44:431-436.
28. Tanti JF and Jager J. Cellular mechanisms of insulin resistance: role of stress-regulated serine kinases and insulin receptor substrates (IRS) serine phosphorylation. Current opinion in pharmacology. 2009; 9:753-762.
29. Tremblay F, Brule S, Hee Um S, Li Y, Masuda K, Roden M, Sun XJ, Krebs M, Polakiewicz RD, Thomas G and Marette A. Identification of IRS-1 Ser-1101 as a target of S6K1 in nutrient- and obesity-induced insulin resistance. Proceedings of the National Academy of Sciences of the United States of America. 2007; 104:14056-14061.
30. Copps KD and White MF. Regulation of insulin sensitivity by serine/threonine phosphorylation of insulin receptor substrate proteins IRS1 and IRS2. Diabetologia. 2012; 55:2565-2582.
31. Hancer NJ, Qiu W, Cherella C, Li Y, Copps KD and White MF. Insulin and metabolic stress stimulate multisite serine/threonine phosphorylation of insulin receptor substrate 1 and inhibit tyrosine phosphorylation. The Journal of biological chemistry. 2014; 289:12467-12484.
32. Brinks H, Das A and Koch WJ. A role for GRK2 in myocardial ischemic injury: indicators of a potential future therapy and diagnostic. Future cardiology. 2011; 7:547-556.
33. Manning BD. Balancing Akt with S6K: implications for both metabolic diseases and tumorigenesis. The Journal of cell biology. 2004; 167:399-403.
34. Harrington LS, Findlay GM and Lamb RF. Restraining PI3K: mTOR signalling goes back to the membrane. Trends in biochemical sciences. 2005; 30:35-42.
35. Piper MD, Selman C, McElwee JJ and Partridge L. Separating cause from effect: how does insulin/IGF signalling control lifespan in worms, flies and mice? Journal of internal medicine. 2008; 263:179-191.
36. Selman C, Partridge L and Withers DJ. Replication of extended lifespan phenotype in mice with deletion of insulin receptor substrate 1. PloS one. 2011; 6:e16144.
37. Masternak MM, Panici JA, Bonkowski MS, Hughes LF and Bartke A. Insulin sensitivity as a key mediator of growth hormone actions on longevity. The journals of gerontology Series A, Biological sciences and medical sciences. 2009; 64:516-521.
38. Aguirre V, Uchida T, Yenush L, Davis R and White MF. The c-Jun NH(2)-terminal kinase promotes insulin resistance during association with insulin receptor substrate-1 and phosphorylation of Ser(307). The Journal of biological chemistry. 2000; 275:9047-9054.
39. Dresner A, Laurent D, Marcucci M, Griffin ME, Dufour S, Cline GW, Slezak LA, Andersen DK, Hundal RS, Rothman DL, Petersen KF and Shulman GI. Effects of free fatty acids on glucose transport and IRS-1-associated phosphatidylinositol 3-kinase activity. The Journal of clinical investigation. 1999; 103:253-259.
40. D’Alessandris C, Lauro R, Presta I and Sesti G. C-reactive protein induces phosphorylation of insulin receptor substrate-1 on Ser307 and Ser 612 in L6 myocytes, thereby impairing the insulin signalling pathway that promotes glucose transport. Diabetologia. 2007; 50:840-849.
41. Rui L, Aguirre V, Kim JK, Shulman GI, Lee A, Corbould A, Dunaif A and White MF. Insulin/IGF-1 and TNF-alpha stimulate phosphorylation of IRS-1 at inhibitory Ser307 via distinct pathways. The Journal of clinical investigation. 2001; 107:181-189.
42. Yu C, Chen Y, Cline GW, Zhang D, Zong H, Wang Y, Bergeron R, Kim JK, Cushman SW, Cooney GJ, Atcheson B, White MF, Kraegen EW, et al. Mechanism by which fatty acids inhibit insulin activation of insulin receptor substrate-1 (IRS-1)-associated phosphatidylinositol 3-kinase activity in muscle. The Journal of biological chemistry. 2002; 277:50230-50236.