
Introduction
Wnt/β-catenin pathway, also called canonical Wnt pathway, is crucial to embryo development and adult tissue homeostasis [1, 2]. Aberrant activation of this pathway can cause uncontrolled cell growth and cell malignant transformation [1, 2]. This oncogenic pathway is initiated by some secreted glycoproteins, such as Wnt1 and Wnt3a. The binding of these Wnt proteins to their membrane receptor Frizzled and co-receptor lipoprotein receptor-related protein 5/6 (LRP5/6) leads to the dissociation of β-catenin from its degrading complex. Thereafter, β-catenin escapes from phosphorylation by glycogen synthase kinase 3β (GSK3β) and subsequent degradation by ubiquitin-proteasome system (UPS). The accumulated β-catenin in the cytoplasm translocates into the nucleus, and combines with transcription factor T cell factor/lymphocyte enhancer factor (TCF/LEF) (Figure 1).
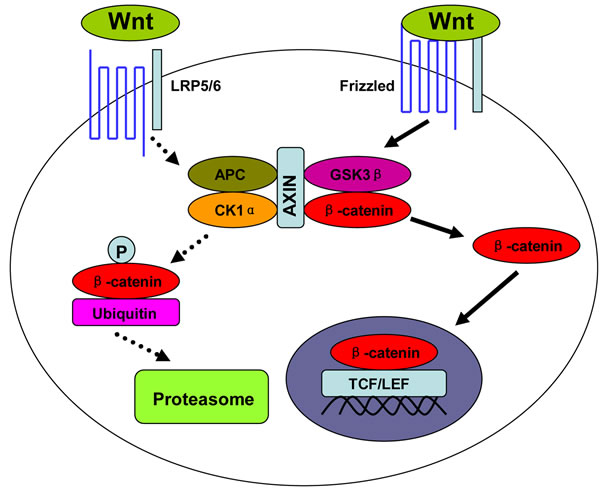
Figure 1: Wnt/β-catenin signal pathway. Upon the binding of Wnt proteins to their receptors, β-catenin dissociates from its degrading complex, which consists of scaffold protein AXIN, casein kinase 1α (CK1α), tumor suppressor adenomatous polyposis coli (APC), and glycogen synthase kinase 3β (GSK3β). The accumulated β-catenin in cytoplasm then translocates into nucleus. P: Phosphorylation.
Helicobacter pylori (H. pylori) infection is a strong risk factor for gastric cancer. The underlying mechanisms include chronic inflammation in gastric mucosa, genetic and epigenetic alterations of tumor suppressor genes, activation of oncogenic signals, and generation of gastric cancer stem cells (CSC) (Figure 2). Chronic inflammation has been recognized as a hallmark of cancer in the recent decade [3, 4]. Aberrant activation of immune cells and overproduction of inflammatory cytokines promote gastric cancer development [5-8]. Infection with H. pylori can induce gastric pre-malignancies by recruiting bone marrow-derived cells (BMDCs) [9, 10]. H. pylori can cause DNA double-strand breaks directly [11], and cause DNA damage indirectly by stimulating the generation of reactive oxygen species (ROS) and reactive nitrogen species (RNS) [12, 13] or by increasing the activity of cytidine deaminase [14]. Hypermethylation as well as subsequent downregulation of tumor suppressor genes is an important epigenetic mechanism in H. pylori-related gastric carcinogenesis [15]. H. pylori induce gastric epithelial cell epithelial-mesenchymal transition (EMT), and generate potential cancer stem cells [16, 17]. H. pylori also stimulate some oncogenic pathways. Activation of epidermal growth factor receptor (EGFR) can resist H. pylori-induced gastric epithelial cell apoptosis [18, 19]. Moreover, increasing evidence has indicated that Wnt/β-catenin pathway is implicated in H. pylori-induced gastric carcinogenesis.
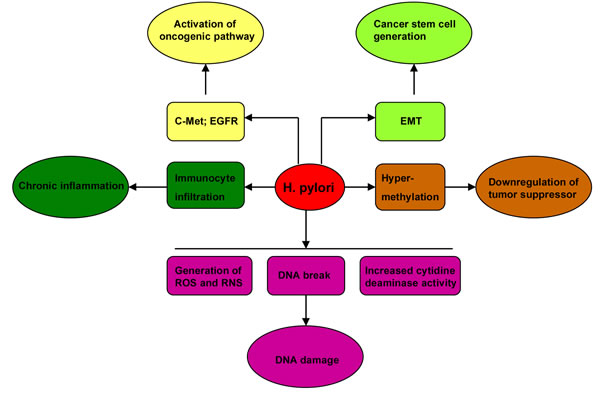
Figure 2: The mechanisms underlying gastric carcinogenesis induced by H. pylori. The mechanisms include chronic inflammation in gastric mucosa, genetic and epigenetic alterations of tumor suppressor genes, activation of oncogenic signals, and generation of gastric cancer stem cells (CSC).
Nuclear β-catenin accumulation in gastric cancers
Investigations on gastric cancer specimens showed that about 20%~30% gastric cancers presented nuclear β-catenin accumulation [20, 21]. Some mutations were identified in β-catenin exon 3 that encodes serine-threonine phosphorylation sites for the GSK3β [20, 21]. These mutations protect β-catenin from phosphorylation by GSK3β and degradation by UPS. However, most of gastric cancer specimens with nuclear β-catenin accumulation did not harbor β-catenin mutation [20, 21]. In colon cancer, APC expression was frequently downregulated, leading to the disassembly of β-catenin degradation complex [22]. Unlike colon cancer, neither APC mutation [23, 24] nor APC methylation [25] seemed to be involved in gastric cancer. These findings suggest that other factors are involved in Wnt/β-catenin activation in gastric cancers.
Several studies from a same group demonstrated the attribution of H. pylori to nuclear β-catenin accumulation. Nuclear β-catenin mainly localized in epithelial cells within the proliferative zone in antral glands, and appeared more frequently in H. pylori cytotoxin-associated gene A (CagA)-positive specimens, compared with either CagA-negative or uninfected patients [26]. CagA-positive H. pylori could induce nuclear β-catenin accumulation in vivo and in vitro [26-28]. Recently, the group further revealed that H. pylori promoted gastric epithelial cell proliferation through β-catenin by using gastroids, three-dimensional organ-like structures [29].
Activation of oncogenic c-met and EGFR by H. pylori
Aberrant activation of c-Met receptor occurred commonly in gastric cancers [30]. Infection with CagA-positive H. pylori induced phosphorylation of c-Met and gastric epithelial cell proliferation [31, 32]. Upon translocating into the cytoplasm, CagA combined with c-Met and CD44 to form a functional complex [31, 32]. CD44 deficiency or inhibition blocked H. pylori-induced gastric epithelial cell proliferation and atrophic gastritis [32]. Activation of c-Met triggered phosphatidylinositol 3-kinase (PI3K)/Akt signaling and caused β-catenin accumulation [33]. C-Met-PI3K-β-catenin pathway is also involved in colorectal cancer. Activation of this pathway promoted cell invasion and proliferation, and protected cells from apoptosis [34]. On the contrary, inactivation of c-Met augmented GSK3β activity and β-catenin degradation [35]. Interestingly, β-catenin accumulation could also upregulate c-Met expression [35, 36], indicating a positive feedback between c-Met and β-catenin in carcinogenesis.
EGFR signals another oncogenic pathway in H. pylori-related gastric cancer [18, 19]. Unlike c-Met, activation of EGFR involved vacuolating cytotoxin A (VacA) [37], CagE [38], CagL [39], H. pylori secretory protein HP0175 [40], and outer inflammatory protein A (OipA) [41], whereas not CagA. Indeed, CagA inactivated EGFR by activating SH2 domain-containing protein tyrosine phosphatase (SHP-2) [42]. H. pylori induced EGFR phosphorylation, and then activated PI3K/Akt pathway [19]. Activation of EGFR-PI3K/Akt signaling resulted in GSK3β suppression and β-catenin accumulation via VacA or OipA [41, 43, 44]. These observations indicate that intracellular pathways initiated by EGFR and c-Met converge at PI3K/Akt-GSK3β-β-catenin under H. pylori infection (Figure 3).
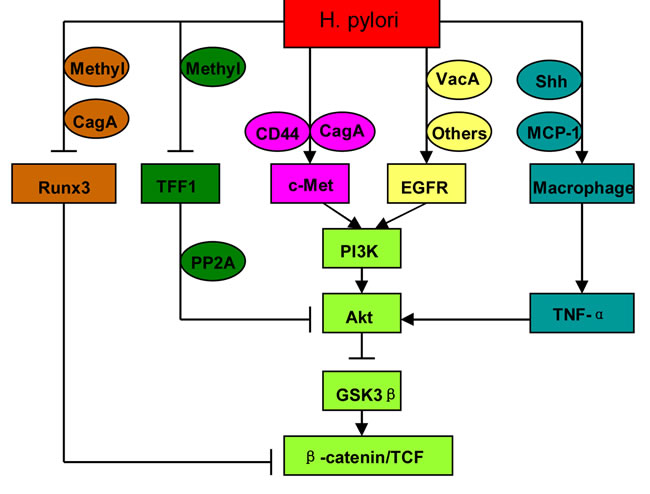
Figure 3: Intracellular signalings mediating the activation of Wnt/β-catenin by H. pylori. Methyl: methylation.
Downregulation of tumor-suppressor Runx3 and TFF1 by H. pylori
Runx3 is an important tumor suppressor for gastric cancer. Runx3 expression was frequently downregulated in gastric cancer cells because of promoter hypermethylation. The clinicopathological analysis on gastric cancers and premalignant lesions showed that Runx3 hypermethylation was correlated with H. pylori infection [45, 46]. In addition to gene hypermethylation, other mechanisms are also involved in H. pylori-induced Runx3 downregulation. CagA could directly associate with Runx3 through a specific recognition of the PY motif of Runx3 by a WW domain of CagA, and result in the ubiquitination and degradation of Runx3 [47]. CagA could also reduce Runx3 mRNA expression by inhibiting Runx3 promoter activity [48]. Runx3 suppressed Wnt/β-catenin pathway by forming a ternary complex with β-catenin/TCF4 [49]. Therefore, Runx3 loss upregulated the expression of Wnt/β-catenin target genes, and induced gastric carcinogenesis [50] (Figure 3).
Trefoil factor 1 (TFF1) was expressed in normal gastric mucosa [51], but frequently downregulated in gastric cancers because of gene mutation [52] and promoter hypermethylation [53]. Recombinant TFF1 protein inhibited gastric epithelial cell proliferation, whereas mutant TFF1 protein lost this effect [54]. Moreover, animals with TFF1 inactivation developed gastric pre-malignant lesions and gastric cancer [55]. These studies indicate that TFF1 is a crucial tumor suppressor for gastric cancer. It is unclear whether H. pylori can induce TFF1 gene mutation, but there is evidence suggesting that H. pylori are responsible for TFF1 gene hypermethylation. TFF1 was significantly downregulated and frequently methylated in H. pylori-positive mucosa, compared with H. pylori-negative mucosa [53, 56]. In N-methyl-N-nitrosourea (MNU)-induced gastric cancers, TFF1 methylation was increased after H. pylori infection [53]. TFF1 inhibited Akt and GSK3β phosphorylation through protein phosphatase 2A (PP2A), and then reduced β-catenin nuclear translocation and TCF transcription activity [57]. On the contrary, TFF1 loss promoted H. pylori-induced oncogenic activation of β-catenin [58] (Figure 3).
Macrophages connecting inflammation with Wnt/β-catenin activation
It has been well accepted that tumor-associated macrophages (TAMs) promote cancer development. H. pylori infection recruited macrophages via monocyte chemoattractant protein-1 (MCP-1) [7, 59] or Sonic Hedgehog (Shh) [60] in gastric mucosa. These macrophages produced pro-inflammatory cytokines, such as TNF-α and IL-1β. TNF-α could activate Wnt/β-catenin via Akt-GSK3β signaling in gastric cancer [6, 7] (Figure 3). Macrophage-derived IL-1β inhibited GSK3β activity and β-catenin degradation, and enhanced TCF transcription activity in colon cancers [61]. The suppression of GSK3β by IL-1β depended on NF-κB and Akt activation [62]. Macrophages are also involved in Wnt/β-catenin activation in cholangiocarcinoma [63]. These observations demonstrate macrophages as important linkers between chronic inflammation and Wnt/β-catenin activation.
MicroRNAs: potential linkers between H. pylori infection and Wnt/β-catenin activation
MicroRNAs (miRs) are small noncoding RNAs that can up- or downregulate the expression of oncogenes and tumor suppressors. Some miRs, such as miR-101, mir-124a, miR-203, miR-210 and miR-320, were downregulated by H. pylori. MiR-101 and miR-320 were inhibited by H. pylori through CagA [64, 65]. Hypermethylation was responsible for miR-124a, miR-203 and miR-210 downregulation [66-68]. The reduction in expression of these miRs activated Wnt/β-catenin pathway in different cells or tissues [69-73], indicating that these miRs functioned as tumor suppressors. On the contrary, miR-21, miR-155, and miR-222 were upregulated by H. pylori [74-76]. These miRs stimulated Wnt/β-catenin pathway, and functioned as oncogenes or tumor-promoters [77-79]. The implication of these miRs in Wnt/β-catenin pathway in gastric cancer remains unknown. Actually, these miRs are candidates linking H. pylori infection with Wnt/β-catenin activation in gastric cancer (Table 1).
Table 1: Potential microRNAs linking H. pylori infection with Wnt/β-catenin activation
miR |
Regulation by H. pylori |
Reference |
Effect on Wnt/β-catenin |
Cell/Tissue |
Reference |
21 101 124a 155 203 210 222 320 |
up down down up down down up down |
74 64 66 75 67 68 76 65 |
activation inhibition inhibition activation inhibition inhibition activation inhibition |
colorectal cancer colorectal cancer glioma hepatocellular carcinoma breast cancer adipose breast cancer prostate cancer |
77 69 71 78 72 73 79 70 |
The effects of H. pylori on upstream molecules in Wnt/β-catenin pathway
Some evidence indicates that H. pylori may activate Wnt/β-catenin pathway by affecting Wnt ligands, receptors or antagonists. H. pylori and TNF-α could induce Wnt10a and Wnt10b expression in gastric cancer cells [80, 81]. H. pylori infection could also activate Wnt co-receptor LRP6, and result in nuclear β-catenin accumulation [82]. Secreted Frizzled-related proteins (SFRPs) can combine with Wnt ligands or receptors to interfere Wnt signaling. These Wnt antagonists were frequently downregulated in gastric cancers due to gene promoter hypermethylation [83]. Actually, SFRP4 and SFRP5 methylation was found to be positively correlated with H. pylori infection [84]. In addition, Wnt3 [85], Wnt7a [86], Wnt7b [87] and Wnt receptor Frizzled [88], were also expressed in gastric cancer cells. The effects of H. pylori on these molecules are still unclear.
The role of Wnt/β-catenin pathway in H. pylori-induced gastric stem cell generation and expansion
Gastric stem cells are implicated in gastric cancer initiation and progression. Via CagA, H. pylori colonized stomach gland epithelium, and promoted stem cell-related gene expression and Lgr5(+) stem cell proliferation [89]. H. pylori also induced gastric epithelial cell EMT to generate gastric cancer stem cells, and this process was also via CagA [16]. The molecular mechanisms underlying H. pylori-induced EMT and stem cell generation remain largely unknown. Wnt/β-catenin pathway was important for gastrointestinal progenitor cell proliferation and differentiation [90, 91]. Activation of this pathway could induce EMT in gastric cancer [92, 93]. Recently, it was revealed that CagA induced EMT by inhibiting GSK-3 activity [94]. Moreover, Wnt/β-catenin target CD44 was observed to be needed in H. pylori-induced gastric stem cell proliferation [95]. These findings indicate that H. pylori induce gastric stem cell generation and proliferation at least partly via Wnt/β-catenin pathway.
Conclusion
Increasing evidence demonstrates Wnt/β-catenin as a crucial pathway stimulated by H. pylori in gastric carcinogenesis. H. pylori can upregulate Wnt/β-catenin activator c-Met and EGFR, and downregulate Wnt/β-catenin suppressor TFF1 and RUNX3. H. pylori can also activate Wnt/β-catenin pathway by recruiting tumor-associated macrophages. Importantly, via Wnt/β-catenin pathway, H. pylori induced gastric stem cell generation and expansion, promoting gastric cancer initiation and progression.
However, there are still some questions need to be answered. Which signal molecule plays a dominant role in Wnt/β-catenin activation under H. pylori infection, c-Met, EGFR, TFF1, Runx3, or else? Can dysregulations of these molecules synergize in gastric cancer development? Which virulent factor of H. pylori plays a dominant role in Wnt/β-catenin activation, CagA, VacA or else? Given the complexities of H. pylori strains and host factors, more work should be done to find the answers. In addition, the effects of H. pylori on Wnt ligands, receptors and antagonists, and the roles of miRs in Wnt/β-catenin activation in gastric cancer, need to be further investigated.
Recently, a series of therapies antagonizing Wnt/β-catenin pathway have entered clinical trials. As Wnt/β-catenin pathway is essential for tissue homeostasis, it remains elusive about their clinical efficacy and safety [96]. H. pylori eradication can reduce the risk of gastric cancer, but it can not completely prevent H. pylori-related gastric carcinogenesis. One of the reasons is that the activation of oncogenic pathway, such as Wnt/β-catenin, has happened before H. pylori eradication.
ACKNOWLEDGMENTS
This work was supported by National Natural Science Foundation of China (No. 81370517).
CONFLICTS OF INTEREST
There is no financial conflict of interest concerning this study.
REFERENCES
1. Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol. 2004;20:781-810.
2. Clevers H. Wnt/beta-catenin signaling in development and disease. Cell. 2006;127:469-480.
3. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646-674.
4. Diakos CI, Charles KA, McMillan DC, Clarke SJ. Cancer-related inflammation and treatment effectiveness. Lancet Oncol. 2014;15:e493-503.
5. Tu S, Bhagat G, Cui G, Takaishi S, Kurt-Jones EA, Rickman B, Betz KS, Penz-Oesterreicher M, Bjorkdahl O, Fox JG, Wang TC. Overexpression of interleukin-1beta induces gastric inflammation and cancer and mobilizes myeloid-derived suppressor cells in mice. Cancer Cell. 2008;14:408-419.
6. Oguma K, Oshima H, Aoki M, Uchio R, Naka K, Nakamura S, Hirao A, Saya H, Taketo MM, Oshima M. Activated macrophages promote Wnt signalling through tumour necrosis factor-alpha in gastric tumour cells. EMBO J. 2008;27:1671-1681.
7. Oshima H, Hioki K, Popivanova BK, Oguma K, Van Rooijen N, Ishikawa TO, Oshima M. Prostaglandin E₂ signaling and bacterial infection recruit tumor-promoting macrophages to mouse gastric tumors. Gastroenterology. 2011;140:596-607.e7.
8. Zhuang Y, Peng LS, Zhao YL, Shi Y, Mao XH, Chen W, Pang KC, Liu XF, Liu T, Zhang JY, Zeng H, Liu KY, Guo G, Tong WD, Shi Y, Tang B, Li N, Yu S, Luo P, Zhang WJ, Lu DS, Yu PW, Zou QM. CD8(+) T cells that produce interleukin-17 regulate myeloid-derived suppressor cells and are associated with survival time of patients with gastric cancer. Gastroenterology. 2012;143:951-962.e8.
9. Houghton J, Stoicov C, Nomura S, Rogers AB, Carlson J, Li H, Cai X, Fox JG, Goldenring JR, Wang TC. Gastric cancer originating from bone marrow-derived cells. Science. 2004;306:1568-1571.
10. Varon C, Dubus P, Mazurier F, Asencio C, Chambonnier L, Ferrand J, Giese A, Senant-Dugot N, Carlotti M, Mégraud F. Helicobacter pylori infection recruits bone marrow-derived cells that participate in gastric preneoplasia in mice. Gastroenterology. 2012;142:281-291.
11. Toller IM, Neelsen KJ, Steger M, Hartung ML, Hottiger MO, Stucki M, Kalali B, Gerhard M, Sartori AA, Lopes M, Müller A. Carcinogenic bacterial pathogen Helicobacter pylori triggers DNA double-strand breaks and a DNA damage response in its host cells. Proc Natl Acad Sci U S A. 2011;108:14944-14949.
12. Obst B, Wagner S, Sewing KF, Beil W. Helicobacter pylori causes DNA damage in gastric epithelial cells. Carcinogenesis. 2000;21:1111-1115.
13. Katsurahara M, Kobayashi Y, Iwasa M, Ma N, Inoue H, Fujita N, Tanaka K, Horiki N, Gabazza EC, Takei Y. Reactive nitrogen species mediate DNA damage in Helicobacter pylori-infected gastric mucosa. Helicobacter. 2009;14:552-558.
14. Shimizu T, Marusawa H, Matsumoto Y, Inuzuka T, Ikeda A, Fujii Y, Minamiguchi S, Miyamoto S, Kou T, Sakai Y, Crabtree JE, Chiba T. Accumulation of somatic mutations in TP53 in gastric epithelium with Helicobacter pylori infection. Gastroenterology. 2014;147:407-417.e3.
15. Zhao C, Bu X. Promoter methylation of tumor-related genes in gastric carcinogenesis. Histol Histopathol. 2012;27:1271-1282.
16. Bessède E, Staedel C, Acuña Amador LA, Nguyen PH, Chambonnier L, Hatakeyama M, Belleannée G, Mégraud F, Varon C. Helicobacter pylori generates cells with cancer stem cell properties via epithelial-mesenchymal transition-like changes. Oncogene. 2014;33:4123-4131.
17. Lee DG, Kim HS, Lee YS, Kim S, Cha SY, Ota I, Kim NH, Cha YH, Yang DH, Lee Y, Park GJ, Yook JI, Lee YC. Helicobacter pylori CagA promotes Snail-mediated epithelial-mesenchymal transition by reducing GSK-3 activity. Nat Commun. 2014;5:4423.
18. Chaturvedi R, Asim M, Piazuelo MB, Yan F, Barry DP, Sierra JC, Delgado AG, Hill S, Casero RA Jr, Bravo LE, Dominguez RL, Correa P, Polk DB, Washington MK, Rose KL, Schey KL, Morgan DR, Peek RM Jr, Wilson KT. Activation of EGFR and ERBB2 by Helicobacter pylori results in survival of gastric epithelial cells with DNA damage. Gastroenterology. 2014;146:1739-1751.e14.
19. Yan F, Cao H, Chaturvedi R, Krishna U, Hobbs SS, Dempsey PJ, Peek RM Jr, Cover TL, Washington MK, Wilson KT, Polk DB. Epidermal growth factor receptor activation protects gastric epithelial cells from Helicobacter pylori-induced apoptosis. Gastroenterology. 2009 ;136:1297-1307, e1-3.
20. Woo DK, Kim HS, Lee HS, Kang YH, Yang HK, Kim WH. Altered expression and mutation of beta-catenin gene in gastric carcinomas and cell lines. Int J Cancer. 2001;95:108-113.
21. Clements WM, Wang J, Sarnaik A, Kim OJ, MacDonald J, Fenoglio-Preiser C, Groden J, Lowy AM. beta-Catenin mutation is a frequent cause of Wnt pathway activation in gastric cancer. Cancer Res. 2002;62:3503-3506.
22. Segditsas S, Tomlinson I. Colorectal cancer and genetic alterations in the Wnt pathway. Oncogene. 2006;25:7531-7537.
23. Powell SM, Cummings OW, Mullen JA, Asghar A, Fuga G, Piva P, Minacci C, Megha T, Tosi P, Jackson CE. Characterization of the APC gene in sporadic gastric adenocarcinomas. Oncogene. 1996;12:1953-1959.
24. Sud R, Talbot IC, Delhanty JD. Infrequent alterations of the APC and MCC genes in gastric cancers from British patients. Br J Cancer. 1996;74:1104-1108.
25. Tsuchiya T, Tamura G, Sato K, Endoh Y, Sakata K, Jin Z, Motoyama T, Usuba O, Kimura W, Nishizuka S, Wilson KT, James SP, Yin J, Fleisher AS, Zou T, Silverberg SG, Kong D, Meltzer SJ. Distinct methylation patterns of two APC gene promoters in normal and cancerous gastric epithelia. Oncogene. 2000;19:3642-3646.
26. Franco AT, Israel DA, Washington MK, Krishna U, Fox JG, Rogers AB, Neish AS, Collier-Hyams L, Perez-Perez GI, Hatakeyama M, Whitehead R, Gaus K, O’Brien DP, Romero-Gallo J, Peek RM Jr. Activation of beta-catenin by carcinogenic Helicobacter pylori. Proc Natl Acad Sci U S A. 2005;102:10646-10651.
27. Franco AT, Johnston E, Krishna U, Yamaoka Y, Israel DA, Nagy TA, Wroblewski LE, Piazuelo MB, Correa P, Peek RM Jr. Regulation of gastric carcinogenesis by Helicobacter pylori virulence factors. Cancer Res. 2008;68:379-387.
28. Nagy TA, Wroblewski LE, Wang D, Piazuelo MB, Delgado A, Romero-Gallo J, Noto J, Israel DA, Ogden SR, Correa P, Cover TL, Peek RM Jr. β-Catenin and p120 mediate PPARδ-dependent proliferation induced by Helicobacter pylori in human and rodent epithelia. Gastroenterology. 2011;141:553-564.
29. Wroblewski LE, Piazuelo MB, Chaturvedi R, Schumacher M, Aihara E, Feng R, Noto JM, Delgado A, Israel DA, Zavros Y, Montrose MH, Shroyer N, Correa P, Wilson KT, Peek RM Jr. Helicobacter pylori targets cancer-associated apical-junctional constituents in gastroids and gastric epithelial cells. Gut. 2015;64:720-730.
30. Hack SP, Bruey JM, Koeppen H. HGF/MET-directed therapeutics in gastroesophageal cancer: a review of clinical and biomarker development. Oncotarget. 2014;5:2866-2880.
31. McCracken KW, Catá EM, Crawford CM, Sinagoga KL, Schumacher M, Rockich BE, Tsai YH, Mayhew CN, Spence JR, Zavros Y, Wells JM. Modelling human development and disease in pluripotent stem-cell-derived gastric organoids. Nature. 2014;516:400-404.
32. Bertaux-Skeirik N, Feng R, Schumacher MA, Li J, Mahe MM, Engevik AC, Javier JE, Peek RM Jr, Ottemann K, Orian-Rousseau V, Boivin GP, Helmrath MA, Zavros Y. CD44 plays a functional role in Helicobacter pylori-induced epithelial cell proliferation. PLoS Pathog. 2015;11:e1004663.
33. Suzuki M, Mimuro H, Kiga K, Fukumatsu M, Ishijima N, Morikawa H, Nagai S, Koyasu S, Gilman RH, Kersulyte D, Berg DE, Sasakawa C. Helicobacter pylori CagA phosphorylation-independent function in epithelial proliferation and inflammation. Cell Host Microbe. 2009;5:23-34.
34. Rasola A, Fassetta M, De Bacco F, D’Alessandro L, Gramaglia D, Di Renzo MF, Comoglio PM. A positive feedback loop between hepatocyte growth factor receptor and beta-catenin sustains colorectal cancer cell invasive growth. Oncogene. 2007;26:1078-1087.
35. Tuynman JB, Vermeulen L, Boon EM, Kemper K, Zwinderman AH, Peppelenbosch MP, Richel DJ. Cyclooxygenase-2 inhibition inhibits c-Met kinase activity and Wnt activity in colon cancer. Cancer Res. 2008;68:1213-1220.
36. Boon EM, van der Neut R, van de Wetering M, Clevers H, Pals ST. Wnt signaling regulates expression of the receptor tyrosine kinase met in colorectal cancer. Cancer Res. 2002;62:5126-5128.
37. Caputo R, Tuccillo C, Manzo BA, Zarrilli R, Tortora G, Blanco Cdel V, Ricci V, Ciardiello F, Romano M. Helicobacter pylori VacA toxin up-regulates vascular endothelial growth factor expression in MKN 28 gastric cells through an epidermal growth factor receptor-, cyclooxygenase-2-dependent mechanism. Clin Cancer Res. 2003;9:2015-2021.
38. Keates S, Keates AC, Nath S, Peek RM Jr, Kelly CP. Transactivation of the epidermal growth factor receptor by cag+ Helicobacter pylori induces upregulation of the early growth response gene Egr-1 in gastric epithelial cells. Gut. 2005;54:1363-1369.
39. Wiedemann T, Hofbaur S, Tegtmeyer N, Huber S, Sewald N, Wessler S, Backert S, Rieder G. Helicobacter pylori CagL dependent induction of gastrin expression via a novel αvβ5-integrin-integrin linked kinase signalling complex. Gut. 2012;61:986-996.
40. Basu S, Pathak SK, Chatterjee G, Pathak S, Basu J, Kundu M. Helicobacter pylori protein HP0175 transactivates epidermal growth factor receptor through TLR4 in gastric epithelial cells. J Biol Chem. 2008;283:32369-32376.
41. Tabassam FH, Graham DY, Yamaoka Y. Helicobacter pylori activate epidermal growth factor receptor- and phosphatidylinositol 3-OH kinase-dependent Akt and glycogen synthase kinase 3beta phosphorylation. Cell Microbiol. 2009;11:70-82.
42. Bauer B, Pang E, Holland C, Kessler M, Bartfeld S, Meyer TF. The Helicobacter pylori virulence effector CagA abrogates human β-defensin 3 expression via inactivation of EGFR signaling. Cell Host Microbe. 2012;11:576-586.
43. Sokolova O, Bozko PM, Naumann M. Helicobacter pylori suppresses glycogen synthase kinase 3beta to promote beta-catenin activity. J Biol Chem. 2008283:29367-29374.
44. Nakayama M, Hisatsune J, Yamasaki E, Isomoto H, Kurazono H, Hatakeyama M, Azuma T, Yamaoka Y, Yahiro K, Moss J, Hirayama T. Helicobacter pylori VacA-induced inhibition of GSK3 through the PI3K/Akt signaling pathway. J Biol Chem. 2009;284:1612-1619.
45. Kitajima Y, Ohtaka K, Mitsuno M, Tanaka M, Sato S, Nakafusa Y, Miyazaki K. Helicobacter pylori infection is an independent risk factor for Runx3 methylation in gastric cancer. Oncol Rep. 2008;19:197-202.
46. Li WQ, Pan KF, Zhang Y, Dong CX, Zhang L, Ma JL, Zhou T, Li JY, You WC. RUNX3 methylation and expression associated with advanced precancerous gastric lesions in a Chinese population. Carcinogenesis. 2011;32:406-410.
47. Tsang YH, Lamb A, Romero-Gallo J, Huang B, Ito K, Peek RM Jr, Ito Y, Chen LF. Helicobacter pylori CagA targets gastric tumor suppressor RUNX3 for proteasome-mediated degradation. Oncogene. 2010;29:5643-5650.
48. Liu Z, Xu X, Chen L, Li W, Sun Y, Zeng J, Yu H, Chen C, Jia J. Helicobacter pylori CagA inhibits the expression of Runx3 via Src/MEK/ERK and p38 MAPK pathways in gastric epithelial cell. J Cell Biochem. 2012;113:1080-1086.
49. Ito K, Lim AC, Salto-Tellez M, Motoda L, Osato M, Chuang LS, Lee CW, Voon DC, Koo JK, Wang H, Fukamachi H, Ito Y. RUNX3 attenuates beta-catenin/T cell factors in intestinal tumorigenesis. Cancer Cell. 2008;14:226-237.
50. Ito K, Chuang LS, Ito T, Chang TL, Fukamachi H, Salto-Tellez M, Ito Y. Loss of Runx3 is a key event in inducing precancerous state of the stomach. Gastroenterology. 2011;140:1536-1546.e8.
51. Rio MC, Bellocq JP, Daniel JY, Tomasetto C, Lathe R, Chenard MP, Batzenschlager A, Chambon P. Breast cancer-associated pS2 protein: synthesis and secretion by normal stomach mucosa. Science. 1988;241:705-708.
52. Park WS, Oh RR, Park JY, Lee JH, Shin MS, Kim HS, Lee HK, Kim YS, Kim SY, Lee SH, Yoo NJ, Lee JY. Somatic mutations of the trefoil factor family 1 gene in gastric cancer. Gastroenterology. 2000;119:691-698.
53. Tomita H, Takaishi S, Menheniott TR, Yang X, Shibata W, Jin G, Betz KS, Kawakami K, Minamoto T, Tomasetto C, Rio MC, Lerkowit N, Varro A, Giraud AS, Wang TC. Inhibition of gastric carcinogenesis by the hormone gastrin is mediated by suppression of TFF1 epigenetic silencing. Gastroenterology. 2011;140:879-891.
54. Yio X, Diamond M, Zhang JY, Weinstein H, Wang LH, Werther L, Itzkowitz S. Trefoil factor family-1 mutations enhance gastric cancer cell invasion through distinct signaling pathways. Gastroenterology. 2006;130:1696-1706.
55. Lefebvre O, Chenard MP, Masson R, Linares J, Dierich A, LeMeur M, Wendling C, Tomasetto C, Chambon P, Rio MC. Gastric mucosa abnormalities and tumorigenesis in mice lacking the pS2 trefoil protein. Science. 1996;274:259-262.
56. Oh JH, Rhyu MG, Jung SH, Choi SW, Kim SI, Hong SJ. Slow overmethylation of housekeeping genes in the body mucosa is associated with the risk for gastric cancer. Cancer Prev Res (Phila). 2014;7:585-595.
57. Soutto M, Peng D, Katsha A, Chen Z, Piazuelo MB, Washington MK, Belkhiri A, Correa P, El-Rifai W. Activation of β-catenin signalling by TFF1 loss promotes cell proliferation and gastric tumorigenesis. Gut. 2015;64:1028-1039.
58. Soutto M, Romero-Gallo J, Krishna U, Piazuelo MB, Washington MK, Belkhiri A, Peek RM Jr, El-Rifai W. Loss of TFF1 promotes Helicobacter pylori-induced β-catenin activation and gastric tumorigenesis. Oncotarget. 2015;6:17911-17922.
59. Kuroda T, Kitadai Y, Tanaka S, Yang X, Mukaida N, Yoshihara M, Chayama K. Monocyte chemoattractant protein-1 transfection induces angiogenesis and tumorigenesis of gastric carcinoma in nude mice via macrophage recruitment. Clin Cancer Res. 2005;11:7629-7636.
60. Schumacher MA, Donnelly JM, Engevik AC, Xiao C, Yang L, Kenny S, Varro A, Hollande F, Samuelson LC, Zavros Y. Gastric Sonic Hedgehog acts as a macrophage chemoattractant during the immune response to Helicobacter pylori. Gastroenterology. 2012;142:1150-1159.e6.
61. Kaler P, Augenlicht L, Klampfer L. Macrophage-derived IL-1beta stimulates Wnt signaling and growth of colon cancer cells: a crosstalk interrupted by vitamin D3. Oncogene. 2009;28:3892-3902.
62. Kaler P, Godasi BN, Augenlicht L, Klampfer L. The NF-κB/AKT-dependent Induction of Wnt Signaling in Colon Cancer Cells by Macrophages and IL-1β. Cancer Microenviron. 2009;2:69-80.
63. Boulter L, Guest RV, Kendall TJ, Wilson DH, Wojtacha D, Robson AJ, Ridgway RA, Samuel K, Van Rooijen N, Barry ST, Wigmore SJ, Sansom OJ, Forbes SJ. WNT signaling drives cholangiocarcinoma growth and can be pharmacologically inhibited. J Clin Invest. 2015;125:1269-1285.
64. Hayashi Y, Tsujii M, Wang J, Kondo J, Akasaka T, Jin Y, Li W, Nakamura T, Nishida T, Iijima H, Tsuji S, Kawano S, Hayashi N, Takehara T. CagA mediates epigenetic regulation to attenuate let-7 expression in Helicobacter pylori-related carcinogenesis. Gut. 2013;62:1536-1546.
65. Noto JM, Piazuelo MB, Chaturvedi R, Bartel CA, Thatcher EJ, Delgado A, Romero-Gallo J, Wilson KT, Correa P, Patton JG, Peek RM Jr. Strain-specific suppression of microRNA-320 by carcinogenic Helicobacter pylori promotes expression of the antiapoptotic protein Mcl-1. Am J Physiol Gastrointest Liver Physiol. 2013;305:G786-796.
66. Ando T, Yoshida T, Enomoto S, Asada K, Tatematsu M, Ichinose M, Sugiyama T, Ushijima T. DNA methylation of microRNA genes in gastric mucosae of gastric cancer patients: its possible involvement in the formation of epigenetic field defect. Int J Cancer. 2009;124:2367-2374.
67. Zhou X, Xu G, Yin C, Jin W, Zhang G. Down-regulation of miR-203 induced by Helicobacter pylori infection promotes the proliferation and invasion of gastric cancer by targeting CASK. Oncotarget. 2014;5:11631-11640.
68. Kiga K, Mimuro H, Suzuki M, Shinozaki-Ushiku A, Kobayashi T, Sanada T, Kim M, Ogawa M, Iwasaki YW, Kayo H, Fukuda-Yuzawa Y, Yashiro M, Fukayama M, Fukao T, Sasakawa C. Epigenetic silencing of miR-210 increases the proliferation of gastric epithelium during chronic Helicobacter pylori infection. Nat Commun. 2014;5:4497.
69. Strillacci A, Valerii MC, Sansone P, Caggiano C, Sgromo A, Vittori L, Fiorentino M, Poggioli G, Rizzello F, Campieri M, Spisni E. Loss of miR-101 expression promotes Wnt/β-catenin signalling pathway activation and malignancy in colon cancer cells. J Pathol. 2013;229:379-389.
70. Hsieh IS, Chang KC, Tsai YT, Ke JY, Lu PJ, Lee KH, Yeh SD, Hong TM, Chen YL. MicroRNA-320 suppresses the stem cell-like characteristics of prostate cancer cells by downregulating the Wnt/beta-catenin signaling pathway. Carcinogenesis. 2013;34:530-538.
71. Lu SH, Jiang XJ, Xiao GL, Liu DY, Yuan XR. miR-124a restoration inhibits glioma cell proliferation and invasion by suppressing IQGAP1 and β-catenin. Oncol Rep. 2014;32:2104-2110.
72. Taube JH, Malouf GG, Lu E, Sphyris N, Vijay V, Ramachandran PP, Ueno KR, Gaur S, Nicoloso MS, Rossi S, Herschkowitz JI, Rosen JM, Issa JP, Calin GA, Chang JT, Mani SA. Epigenetic silencing of microRNA-203 is required for EMT and cancer stem cell properties. Sci Rep. 2013;3:2687.
73. Qin L, Chen Y, Niu Y, Chen W, Wang Q, Xiao S, Li A, Xie Y, Li J, Zhao X, He Z, Mo D. A deep investigation into the adipogenesis mechanism: profile of microRNAs regulating adipogenesis by modulating the canonical Wnt/beta-catenin signaling pathway. BMC Genomics. 2010;11:320.
74. Zhang Z, Li Z, Gao C, Chen P, Chen J, Liu W, Xiao S, Lu H. miR-21 plays a pivotal role in gastric cancer pathogenesis and progression. Lab Invest. 2008;88:1358-1366.
75. Link A, Schirrmeister W, Langner C, Varbanova M, Bornschein J, Wex T, Malfertheiner P. Differential expression of microRNAs in preneoplastic gastric mucosa. Sci Rep. 2015;5:8270.
76. Li N, Tang B, Zhu ED, Li BS, Zhuang Y, Yu S, Lu DS, Zou QM, Xiao B, Mao XH. Increased miR-222 in H. pylori-associated gastric cancer correlated with tumor progression by promoting cancer cell proliferation and targeting RECK. FEBS Lett. 2012;586:722-728.
77. Lin PL, Wu DW, Huang CC, He TY, Chou MC, Sheu GT, Lee H. MicroRNA-21 promotes tumour malignancy via increased nuclear translocation of β-catenin and predicts poor outcome in APC-mutated but not in APC-wild-type colorectal cancer. Carcinogenesis. 2014;35:2175-2182.
78. Zhang Y, Wei W, Cheng N, Wang K, Li B, Jiang X, Sun S. Hepatitis C virus-induced up-regulation of microRNA-155 promotes hepatocarcinogenesis by activating Wnt signaling. Hepatology. 2012;56:1631-1640.
79. Rao X, Di Leva G, Li M, Fang F, Devlin C, Hartman-Frey C, Burow ME, Ivan M, Croce CM, Nephew KP. MicroRNA-221/222 confers breast cancer fulvestrant resistance by regulating multiple signaling pathways. Oncogene. 2011;30:1082-1097.
80. Kirikoshi H, Sekihara H, Katoh M. Up-regulation of WNT10A by tumor necrosis factor alpha and Helicobacter pylori in gastric cancer. Int J Oncol. 2001;19:533-536.
81. Saitoh T, Kirikoshi H, Mine T, Katoh M. Proto-oncogene WNT10B is up-regulated by tumor necrosis factor alpha in human gastric cancer cell line MKN45. Int J Oncol. 2001;19:1187-1192.
82. Gnad T, Feoktistova M, Leverkus M, Lendeckel U, Naumann M. Helicobacter pylori-induced activation of beta-catenin involves low density lipoprotein receptor-related protein 6 and Dishevelled. Mol Cancer. 2010;9:31.
83. Nojima M, Suzuki H, Toyota M, Watanabe Y, Maruyama R, Sasaki S, Sasaki Y, Mita H, Nishikawa N, Yamaguchi K, Hirata K, Itoh F, Tokino T, Mori M, Imai K, Shinomura Y. Frequent epigenetic inactivation of SFRP genes and constitutive activation of Wnt signaling in gastric cancer. Oncogene. 2007;26:4699-4713.
84. Kang GH, Lee S, Cho NY, Gandamihardja T, Long TI, Weisenberger DJ, Campan M, Laird PW. DNA methylation profiles of gastric carcinoma characterized by quantitative DNA methylation analysis. Lab Invest. 2008;88:161-170.
85. Katoh M. Molecular cloning and characterization of human WNT3. Int J Oncol. 2001;1):977-982.
86. Kirikoshi H, Katoh M. Expression of WNT7A in human normal tissues and cancer, and regulation of WNT7A and WNT7B in human cancer. Int J Oncol. 2002;21:895-900.
87. Kirikoshi H, Sekihara H, Katoh M. Molecular cloning and characterization of human WNT7B. Int J Oncol. 2001;19:779-783.
88. Kirikoshi H, Sekihara H, Katoh M. Expression profiles of 10 members of Frizzled gene family in human gastric cancer. Int J Oncol. 2001;19:767-771.
89. Sigal M, Rothenberg ME, Logan CY, Lee JY, Honaker RW, Cooper RL, Passarelli B, Camorlinga M, Bouley DM, Alvarez G, Nusse R, Torres J, Amieva MR. Helicobacter pylori Activates and Expands Lgr5(+) Stem Cells Through Direct Colonization of the Gastric Glands. Gastroenterology. 2015;148:1392-1404.e21.
90. Ireland H, Kemp R, Houghton C, Howard L, Clarke AR, Sansom OJ, Winton DJ. Inducible Cre-mediated control of gene expression in the murine gastrointestinal tract: effect of loss of beta-catenin. Gastroenterology. 2004;126:1236-1246.
91. Mao J, Fan S, Ma W, Fan P, Wang B, Zhang J, Wang H, Tang B, Zhang Q, Yu X, Wang L, Song B, Li L. Roles of Wnt/β-catenin signaling in the gastric cancer stem cells proliferation and salinomycin treatment. Cell Death Dis. 2014;5:e1039.
92. Huang J, Xiao D, Li G, Ma J, Chen P, Yuan W, Hou F, Ge J, Zhong M, Tang Y, Xia X, Chen Z. EphA2 promotes epithelial-mesenchymal transition through the Wnt/β-catenin pathway in gastric cancer cells. Oncogene. 2014;33:2737-2747.
93. Tong X, Li L, Li X, Heng L, Zhong L, Su X, Rong R, Hu S, Liu W, Jia B, Liu X, Kou G, Han J, Guo S, Hu Y, Li C, Tao Q, Guo Y. SOX10, a novel HMG-box-containing tumor suppressor, inhibits growth and metastasis of digestive cancers by suppressing the Wnt/β-catenin pathway. Oncotarget. 2014;5:10571-10583.
94. Lee DG, Kim HS, Lee YS, Kim S, Cha SY, Ota I, Kim NH, Cha YH, Yang DH, Lee Y, Park GJ, Yook JI, Lee YC. Helicobacter pylori CagA promotes Snail-mediated epithelial-mesenchymal transition by reducing GSK-3 activity. Nat Commun. 2014;5:4423.
95. Khurana SS, Riehl TE, Moore BD, Fassan M, Rugge M, Romero-Gallo J, Noto J, Peek RM Jr, Stenson WF, Mills JC. The hyaluronic acid receptor CD44 coordinates normal and metaplastic gastric epithelial progenitor cell proliferation. J Biol Chem. 2013;288:16085-16097.
96. Kahn M. Can we safely target the WNT pathway? Nat Rev Drug Discov. 2014;13:513-532.