
Basic leucine zipper transcription factor (TF) Batf2 belongs to the activator protein 1 family of transcription factors (TFs), which includes Batf and Batf3 [1-6]. The Batf family members play important functional roles in the development and differentiation of dendritic cells and T lymphocytes, in regulating Th2 cell functions and antibody class switching [7]. For example, Batf3 is critical for CD8α+ dendritic cell development [8] and both Batf and Batf2 can compensate for Batf3 in this process (Figure 1A-1C). Mice deficient in Batf2 have reduced percentage of lung resident CD103+ dendritic cells during intracellular parasite T. gondii infection [9]. Batf is more specific for lymphocytes (Figure 1A), regulating differentiation of Th2 [10], Th9 [11] and Th17 cells [12], follicular helper T cells [10, 13], effector CD8+ T cells [14], adipose resident regulatory T cells [15] and B cell IgG class switching [10, 13]. Batf2 was cloned, characterized and identified as a type 1 IFN (IFN-α/β)-inducible early response gene [5] but seem to be mainly restricted to macrophages and DCs following LPS and IFN-γ stimulation [9]. Since Batf2 is induced by type I IFNs [5], one could speculate that Batf2 may play a fundamental role during viral infection including HIV, however no studies investigated this hypothesis so far.
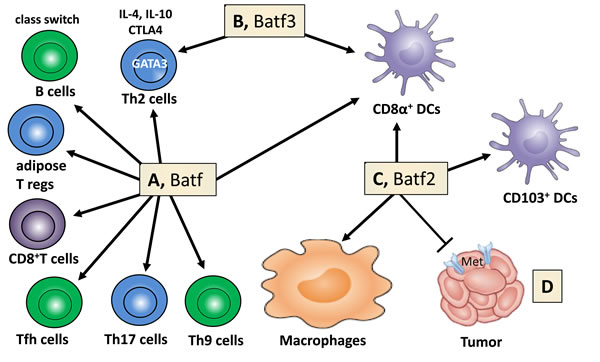
Figure 1: Batf family members regulate cell lineage development, macrophage activation and cancer growth. A. Batf controls the differentiation of Th9, Th17 cells, follicular helper T (Tfh) cells, effector CD8+ T cells and adipose tissue-resident regulatory T cells. Immunoglobulin class switching in B cells, TF (GATA3) and effector factors (IL-4, IL-10, CTLA4) in Th2 cells are regulated by Batf. B. Batf3 contributes to the control of Th2 cell-associated factors and is necessary for the development of CD8α+ dendritic cells. C. Batf2 assists in the lineage development of CD8α+ and CD103+ dendritic cells and controls macrophage activation. D. Batf2 constrains cancer cell growth through MET suppression (adapted and modified from Murphy TL, Tussiwand R, Murphy KM: Nat Rev Immunol 2013, 13(7):499-509).
To further dissect biological roles of Batf family members in different cell types, we composed a mRNA expression atlas of Batf, Batf2 and Batf3 using a large scale genomic analysis, FANTOM (Functional Annotation of the Mammalian Genome) that maps transcription start sites to generate a promoter-level mammalian expression atlas [16] to study the dynamic regulation of enhancers and promoters during mammalian cellular activation and differentiation [17]. The FANTOM consortium utilized the cap analysis gene expression (CAGE) biotechnology [18], which sequences short nucleotide sequence tags from the 5’ end of mRNAs. The CAGE tags are then mapped to the genome to identify transcription start sites and the tag counts are used to quantify the expression of mRNAs. Using this method, RNA Batf family members across a collection of various cancer cell lines (250), human (573) and mouse primary cells (128) were identified (Table 1 and 2), quantified in tags per million (TPM) and normalized by relative log expression. In accordance with the biological role for Batf predominantly in lymphocyte function and development, high Batf expression was found in T and B lymphocytes, as well as in macrophages. In addition, Batf was measured in other cell types that were not previously shown to express Batf (megakaryocytes, endothelial, epithelial and Langerhans cells). Batf2 expression seems to be mainly restricted to macrophages in mouse (12.38 TPM; 56% expression from the dataset collection) and human monocytes/macrophages (185.65 TPM; 76% expression from the dataset collection), but low expression was also found in enterocytes, endothelial cells, adrenal cortex cells, chondroblasts and epithelial cells among others. Batf3 was strongly expressed in human cells, including immature dendritic cells, myeloid, T, NK cells and lower levels in human monocytes and macrophages. Mouse Batf3 showed minimal expression in macrophages and erythroblasts.
Table 1: Batf, Batf2 and Batf3 expression in mouse cell types.
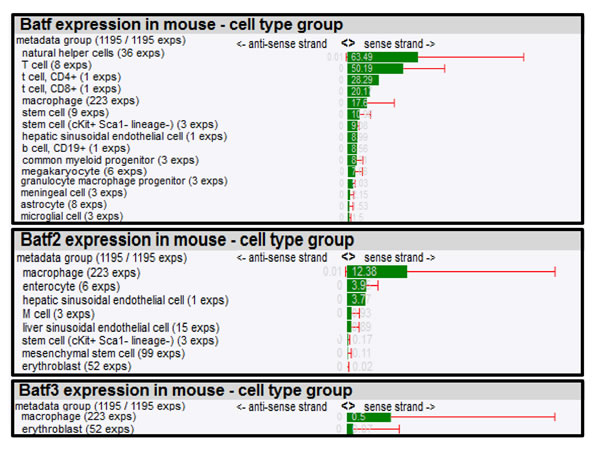
Expression of Batf family members was quantified by CAGE and tags per million normalized by relative log expression are shown. Cell types are ranked according to their highest expression (Exps = experiments).
Table 2: Batf, Batf2 and Batf3 expression in human cell types.
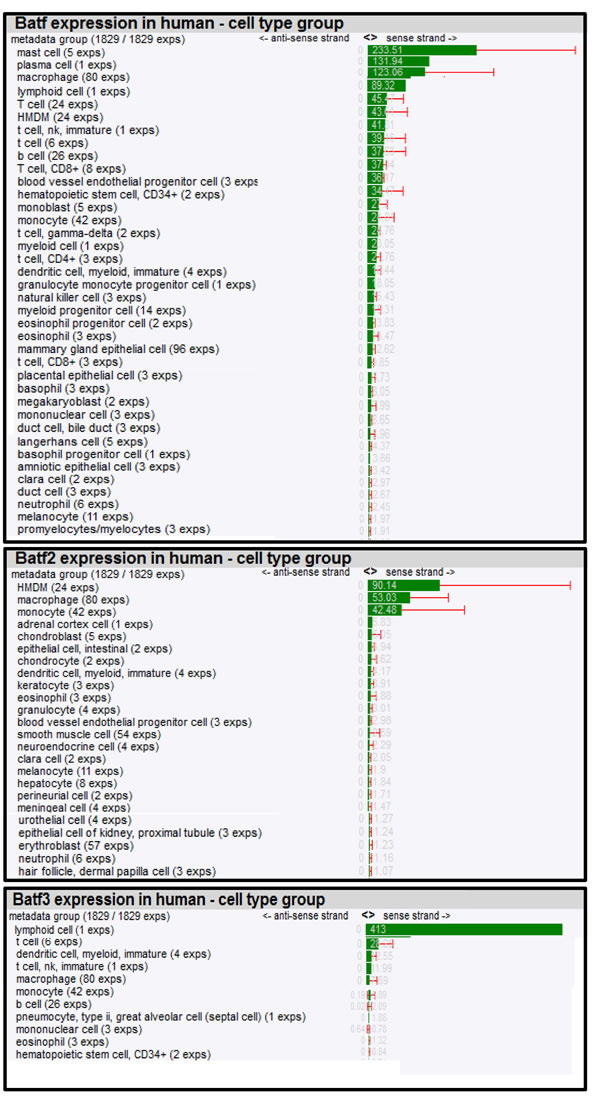
Expression of Batf family members was quantified by CAGE and tags per million normalized by relative log expression are shown. Cell types are ranked according to their highest expression (HMDM= human monocyte-derived macrophages).
We recently reported that Batf2 was significantly induced in macrophages following LPS or IFN-γ stimulation [19]. Indeed, alternatively activated or non-stimulated macrophages showed low or no expression but classical activation M(IFN-γ) highly induced Batf2.Interestingly, Batf2 knockdown experiments in IFN-γ or LPS-stimulated macrophages using shRNA resulted in reduced expression of host protective genes, such as Nos2, Tnf, Ccl5, IL-12b and Socs1. These genes are involved in controlling inflammatory cell recruitment and/or the activation of bactericidal defense mechanisms (Figure 2). As the Batf family lack DNA binding domains [5], we further demonstrated that Batf2 directly interacts with Irf1 by immunoprecipitation. Hence, Batf2/Irf1is likely to cooperatively regulate these immune effector genes, which is well consistent with that the other family member Batf associates with Irf4 and Irf8 to mediate downstream gene activation [9, 20, 21]. Importantly, Batf2 was also induced during M. tuberculosis (Mtb, Beijing strain HN878) infection in classical activated macrophages and shRNA-mediated down-regulation of Batf2 resulted in decreased expression Nos2, Tnf, Ccl5 and IL-12b in heat-killed Mtb-stimulated macrophages (Figure 2). We currently investigate the consequence of Batf2 deficiency in mice during infection with M. tuberculosis and Listeria monocytogenes. Together, these results highlight the importance of Batf2 in controlling macrophage activation during IFN-γ, LPS and mycobacterial infection. Hence, Batf2 may be an important transcription factor to control the switch of inflammatory responses during certain immune processes.We currently started infection studies in Batf2 deficient mice, and depending on the biological outcome, Batf2 might be an interesting biomarker and possible candidate for host directed therapy against tuberculosis (TB).
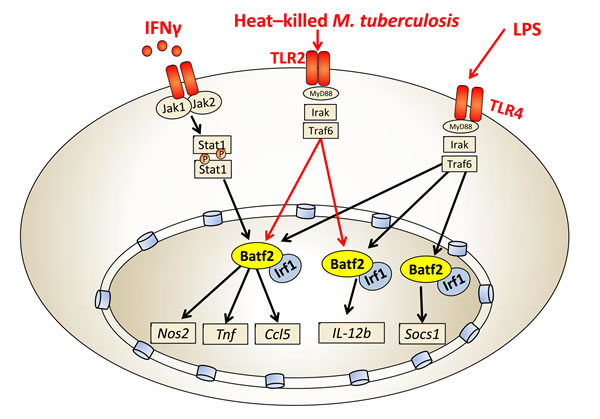
Figure 2: Batf2/Irf1 controls macrophage-specific inflammatory responses. Batf2/Irf1 induces inflammatory responses (Nos2, Tnf, Ccl5, IL-12b and Socs1) in IFN-γ, heat-killed Mtb and LPS-stimulated macrophages.
In recent years, it became evident that adjunctive host-directed drug therapy in combination with current first and second line treatments with antibiotics could develop into a promising innovative approach to treat drug resistant Mtb strains by reducing tissue pathology and possibly shorten the duration of current treatments [22-38]. The existing selection of potential host-directed drug candidates against TB disease are extensive and do include FDA approved drugs that are currently used for treatments of other diseases and conditions. This includes Gefitinib [39], Fluoxetine [39], Metformin [40, 41], Nitazoxanide [42], ProchlorperazineEdisylate [43], Nortriptyline [43], Haloperidol [43], Lithium [44], Imatinib [45, 46], Rapamycin [47, 48], high-dose immunoglobulin [49], TNF blockers [50-52], thalidomide analog [53], Ibuprofen [54, 55], leukotriene inhibitors [56], statins [57, 58], PPARγ antagonists [59], Vitamin D [59-61], IFNγ [62], phosphodiesterase inhibitors [63], metalloproteinase inhibitors [64], autologous mesenchymal stromal cell infusion [65], and corticosteroids [66, 67], among others. We suggest to include Batf2 in the search of new targets for host-directed drug therapies against tuberculosis due to its important regulation of inflammation and macrophage killing effector functions and its specific expression to macrophage/DC cells, the primary target cells of Mtb.
We believe that large scale genomic projects consortium are initial steps for the identification of potential drug targets, which is certainly of utter importance. Indeed, pathogens successfully exploit and modulate the host epigenome for their survival and persistence, including TFs like Stat1, Daxx or ZNF23 [68]. Hence, we identified TFs differentially expressed between classical and alternative activated macrophages [69], building on the hypothesis that intracellular pathogens avoid classical activation, while persisting in alternative activated or non-stimulated macrophages [70]. Functional characterization of these selected TFs may direct us to the identification of host-directed drug targets to increase immunity of the infected host.
We also suggest to include Batf2 as therapeutic target against cancer as Batf2 has been shown as a novel tumor suppresser gene, inhibiting growth of cancer cells [5, 71-73] through repression of hepatocyte growth factor receptor / MET signaling (Figure 1D) [74]. Low Batf2 expression, in patients with colorectal cancer [74], hepatocellular carcinoma (HCC) [75] or oral tongue squamous cell carcinoma [76] do have significant increased mortality when compared to cancer patients with high Batf2 expression and overexpression of Batf2 [5] promotes growth inhibition and apoptosis in cancer cells, but not in normal cells.
In conclusion, for a host-directed drug therapy against TB, we recommend targeting Batf2 specifically in macrophages and dendritic cells to suppress inflammation and limit pathology. Antagonizing Batf2 might be useful for other immune-related diseases where inflammation induces tissue destruction and pathology. In cancer, Batf2 could be used as a biomarker for cancer prognosis and a promising therapeutic target against cancer, by augmenting Batf2 in malignant cells.
Acknowledgments
This work was supported by National Research Foundation (NRF) of South Africa, the Department of Science and Technology, South African Research Chair Initiative, NRF Competitive Programme for Unrated Researchers, South African Medical Research Council (SAMRC) and the Strategic International Research Cooperative Program in Japan.
Conflicts of Interest
There is no conflict of interest.
References
1. Bower KE, Zeller RW, Wachsman W, Martinez T and McGuire KL. Correlation of transcriptional repression by p21(SNFT) with changes in DNA.NF-AT complex interactions. J Biol Chem. 2002; 277:34967-34977.
2. Dorsey MJ, Tae HJ, Sollenberger KG, Mascarenhas NT, Johansen LM and Taparowsky EJ. B-ATF: a novel human bZIP protein that associates with members of the AP-1 transcription factor family. Oncogene. 1995; 11:2255-2265.
3. Hasegawa H, Utsunomiya Y, Kishimoto K, Tange Y, Yasukawa M and Fujita S. SFA-2, a novel bZIP transcription factor induced by human T-cell leukemia virus type I, is highly expressed in mature lymphocytes. Biochem Biophys Res Commun. 1996; 222:164-170.
4. Iacobelli M, Wachsman W and McGuire KL. Repression of IL-2 promoter activity by the novel basic leucine zipper p21SNFT protein. J Immunol. 2000; 165:860-868.
5. Su ZZ, Lee SG, Emdad L, Lebdeva IV, Gupta P, Valerie K, Sarkar D and Fisher PB. Cloning and characterization of SARI (suppressor of AP-1, regulated by IFN). Proc Natl Acad Sci U S A. 2008; 105:20906-20911.
6. Williams KL, Nanda I, Lyons GE, Kuo CT, Schmid M, Leiden JM, Kaplan MH and Taparowsky EJ. Characterization of murine BATF: a negative regulator of activator protein-1 activity in the thymus. Eur J Immunol. 2001; 31:1620-1627.
7. Murphy TL, Tussiwand R and Murphy KM. Specificity through cooperation: BATF-IRF interactions control immune-regulatory networks. Nat Rev Immunol. 2013; 13:499-509.
8. Hildner K, Edelson BT, Purtha WE, Diamond M, Matsushita H, Kohyama M, Calderon B, Schraml BU, Unanue ER, Diamond MS, Schreiber RD, Murphy TL and Murphy KM. Batf3 deficiency reveals a critical role for CD8alpha+ dendritic cells in cytotoxic T cell immunity. Science. 2008; 322:1097-1100.
9. Tussiwand R, Lee WL, Murphy TL, Mashayekhi M, Kc W, Albring JC, Satpathy AT, Rotondo JA, Edelson BT, Kretzer NM, Wu X, Weiss LA, Glasmacher E, Li P, Liao W, Behnke M, et al. Compensatory dendritic cell development mediated by BATF-IRF interactions. Nature. 2012; 490:502-507.
10. Betz BC, Jordan-Williams KL, Wang C, Kang SG, Liao J, Logan MR, Kim CH and Taparowsky EJ. Batf coordinates multiple aspects of B and T cell function required for normal antibody responses. J Exp Med. 2010; 207:933-942.
11. Jabeen R, Goswami R, Awe O, Kulkarni A, Nguyen ET, Attenasio A, Walsh D, Olson MR, Kim MH, Tepper RS, Sun J, Kim CH, Taparowsky EJ, Zhou B and Kaplan MH. Th9 cell development requires a BATF-regulated transcriptional network. J Clin Invest. 2013; 123:4641-4653.
12. Schraml BU, Hildner K, Ise W, Lee WL, Smith WA, Solomon B, Sahota G, Sim J, Mukasa R, Cemerski S, Hatton RD, Stormo GD, Weaver CT, Russell JH, Murphy TL and Murphy KM. The AP-1 transcription factor Batf controls T(H)17 differentiation. Nature. 2009; 460:405-409.
13. Ise W, Kohyama M, Schraml BU, Zhang T, Schwer B, Basu U, Alt FW, Tang J, Oltz EM, Murphy TL and Murphy KM. The transcription factor BATF controls the global regulators of class-switch recombination in both B cells and T cells. Nat Immunol. 2011; 12:536-543.
14. Kurachi M, Barnitz RA, Yosef N, Odorizzi PM, DiIorio MA, Lemieux ME, Yates K, Godec J, Klatt MG, Regev A, Wherry EJ and Haining WN. The transcription factor BATF operates as an essential differentiation checkpoint in early effector CD8+ T cells. Nat Immunol. 2014; 15:373-383.
15. Vasanthakumar A, Moro K, Xin A, Liao Y, Gloury R, Kawamoto S, Fagarasan S, Mielke LA, Afshar-Sterle S, Masters SL, Nakae S, Saito H, Wentworth JM, Li P, Liao W, Leonard WJ, et al. The transcriptional regulators IRF4, BATF and IL-33 orchestrate development and maintenance of adipose tissue-resident regulatory T cells. Nat Immunol. 2015; 16:276-285.
16. Forrest AR, Kawaji H, Rehli M, Baillie JK, de Hoon MJ, Haberle V, Lassman T, Kulakovskiy IV, Lizio M, Itoh M, Andersson R, Mungall CJ, Meehan TF, Schmeier S, Bertin N, Jorgensen M, et al. A promoter-level mammalian expression atlas. Nature. 2014; 507:462-470.
17. Arner E, Daub CO, Vitting-Seerup K, Andersson R, Lilje B, Drablos F, Lennartsson A, Ronnerblad M, Hrydziuszko O, Vitezic M, Freeman TC, Alhendi AM, Arner P, Axton R, Baillie JK, Beckhouse A, et al. Gene regulation. Transcribed enhancers lead waves of coordinated transcription in transitioning mammalian cells. Science. 2015; 347:1010-1014.
18. de Hoon M and Hayashizaki Y. Deep cap analysis gene expression (CAGE): genome-wide identification of promoters, quantification of their expression, and network inference. Biotechniques. 2008; 44:627-628, 630, 632.
19. Roy S, Guler R, Parihar SP, Schmeier S, Kaczkowski B, Nishimura H, Shin JW, Negishi Y, Ozturk M, Hurdayal R, Kubosaki A, Kimura Y, de Hoon MJ, Hayashizaki Y, Brombacher F and Suzuki H. Batf2/Irf1 Induces Inflammatory Responses in Classically Activated Macrophages, Lipopolysaccharides, and Mycobacterial Infection. J Immunol. 2015; 194:6035-6044.
20. Ciofani M, Madar A, Galan C, Sellars M, Mace K, Pauli F, Agarwal A, Huang W, Parkurst CN, Muratet M, Newberry KM, Meadows S, Greenfield A, Yang Y, Jain P, Kirigin FK, et al. A validated regulatory network for Th17 cell specification. Cell. 2012; 151:289-303.
21. Glasmacher E, Agrawal S, Chang AB, Murphy TL, Zeng W, Vander Lugt B, Khan AA, Ciofani M, Spooner CJ, Rutz S, Hackney J, Nurieva R, Escalante CR, Ouyang W, Littman DR, Murphy KM, et al. A genomic regulatory element that directs assembly and function of immune-specific AP-1-IRF complexes. Science. 2012; 338:975-980.
22. Bento CF, Empadinhas N and Mendes V. Autophagy in the fight against tuberculosis. DNA Cell Biol. 2015; 34:228-242.
23. Hawn TR, Shah JA and Kalman D. New tricks for old dogs: countering antibiotic resistance in tuberculosis with host-directed therapeutics. Immunol Rev. 2015; 264:344-362.
24. Iannaccone M, Dorhoi A and Kaufmann SH. Host-directed therapy of tuberculosis: what is in it for microRNA? Expert Opin Ther Targets. 2014; 18:491-494.
25. Schwegmann A and Brombacher F. Host-directed drug targeting of factors hijacked by pathogens. Sci Signal. 2008; 1:re8.
26. Wallis RS and Hafner R. Advancing host-directed therapy for tuberculosis. Nat Rev Immunol. 2015; 15:255-263.
27. Tobin DM. Host-Directed Therapies for Tuberculosis. Cold Spring Harb Perspect Med. 2015.
28. Hmama Z, Pena-Diaz S, Joseph S and Av-Gay Y. Immunoevasion and immunosuppression of the macrophage by Mycobacterium tuberculosis. Immunol Rev. 2015; 264:220-232.
29. Cambier CJ, Falkow S and Ramakrishnan L. Host evasion and exploitation schemes of Mycobacterium tuberculosis. Cell. 2014; 159:1497-1509.
30. Huynh KK, Joshi SA and Brown EJ. A delicate dance: host response to mycobacteria. Curr Opin Immunol. 2011; 23:464-472.
31. Rajaram MV, Ni B, Dodd CE and Schlesinger LS. Macrophage immunoregulatory pathways in tuberculosis. Semin Immunol. 2014; 26:471-485.
32. Maeurer M and Zumla A. The host battles drug-resistant tuberculosis. Sci Transl Med. 2014; 6:263fs247.
33. Kaufmann SH, Lange C, Rao M, Balaji KN, Lotze M, Schito M, Zumla AI and Maeurer M. Progress in tuberculosis vaccine development and host-directed therapies—a state of the art review. Lancet Respir Med. 2014; 2:301-320.
34. Zumla A, Rao M, Parida SK, Keshavjee S, Cassell G, Wallis R, Axelsson-Robertsson R, Doherty M, Andersson J and Maeurer M. Inflammation and tuberculosis: host-directed therapies. J Intern Med. 2015; 277:373-387.
35. Hawn TR, Matheson AI, Maley SN and Vandal O. Host-directed therapeutics for tuberculosis: can we harness the host? Microbiol Mol Biol Rev. 2013; 77:608-627.
36. Rayasam GV and Balganesh TS. Exploring the potential of adjunct therapy in tuberculosis. Trends Pharmacol Sci. 2015.
37. Parida SK, Madansein R, Singh N, Padayatchi N, Master I, Naidu K, Zumla A and Maeurer M. Cellular therapy in tuberculosis. Int J Infect Dis. 2015; 32:32-38.
38. Uhlin M, Andersson J, Zumla A and Maeurer M. Adjunct immunotherapies for tuberculosis. J Infect Dis. 2012; 205 Suppl 2:S325-334.
39. Stanley SA, Barczak AK, Silvis MR, Luo SS, Sogi K, Vokes M, Bray MA, Carpenter AE, Moore CB, Siddiqi N, Rubin EJ and Hung DT. Identification of host-targeted small molecules that restrict intracellular Mycobacterium tuberculosis growth. PLoS Pathog. 2014; 10:e1003946.
40. Vashisht R and Brahmachari SK. Metformin as a potential combination therapy with existing front-line antibiotics for Tuberculosis. J Transl Med. 2015; 13:83.
41. Singhal A, Jie L, Kumar P, Hong GS, Leow MK, Paleja B, Tsenova L, Kurepina N, Chen J, Zolezzi F, Kreiswirth B, Poidinger M, Chee C, Kaplan G, Wang YT and De Libero G. Metformin as adjunct antituberculosis therapy. Sci Transl Med. 2014; 6:263ra159.
42. Lam KK, Zheng X, Forestieri R, Balgi AD, Nodwell M, Vollett S, Anderson HJ, Andersen RJ, Av-Gay Y and Roberge M. Nitazoxanide stimulates autophagy and inhibits mTORC1 signaling and intracellular proliferation of Mycobacterium tuberculosis. PLoS Pathog. 2012; 8:e1002691.
43. Sundaramurthy V, Barsacchi R, Samusik N, Marsico G, Gilleron J, Kalaidzidis I, Meyenhofer F, Bickle M, Kalaidzidis Y and Zerial M. Integration of chemical and RNAi multiparametric profiles identifies triggers of intracellular mycobacterial killing. Cell Host Microbe. 2013; 13:129-142.
44. Sohn H, Kim K, Lee KS, Choi HG, Lee KI, Shin AR, Kim JS, Shin SJ, Song CH, Park JK and Kim HJ. Lithium inhibits growth of intracellular Mycobacterium kansasii through enhancement of macrophage apoptosis. J Microbiol. 2014; 52:299-306.
45. Napier RJ, Rafi W, Cheruvu M, Powell KR, Zaunbrecher MA, Bornmann W, Salgame P, Shinnick TM and Kalman D. Imatinib-sensitive tyrosine kinases regulate mycobacterial pathogenesis and represent therapeutic targets against tuberculosis. Cell Host Microbe. 2011; 10:475-485.
46. Yogalingam G and Pendergast AM. Abl kinases regulate autophagy by promoting the trafficking and function of lysosomal components. J Biol Chem. 2008; 283:35941-35953.
47. Romagnoli A, Etna MP, Giacomini E, Pardini M, Remoli ME, Corazzari M, Falasca L, Goletti D, Gafa V, Simeone R, Delogu G, Piacentini M, Brosch R, Fimia GM and Coccia EM. ESX-1 dependent impairment of autophagic flux by Mycobacterium tuberculosis in human dendritic cells. Autophagy. 2012; 8:1357-1370.
48. Gutierrez MG, Master SS, Singh SB, Taylor GA, Colombo MI and Deretic V. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell. 2004; 119:753-766.
49. Roy E, Stavropoulos E, Brennan J, Coade S, Grigorieva E, Walker B, Dagg B, Tascon RE, Lowrie DB, Colston MJ and Jolles S. Therapeutic efficacy of high-dose intravenous immunoglobulin in Mycobacterium tuberculosis infection in mice. Infect Immun. 2005; 73:6101-6109.
50. Wallis RS, van Vuuren C and Potgieter S. Adalimumab treatment of life-threatening tuberculosis. Clin Infect Dis. 2009; 48:1429-1432.
51. Skerry C, Harper J, Klunk M, Bishai WR and Jain SK. Adjunctive TNF inhibition with standard treatment enhances bacterial clearance in a murine model of necrotic TB granulomas. PLoS One. 2012; 7:e39680.
52. Bourigault ML, Vacher R, Rose S, Olleros ML, Janssens JP, Quesniaux VF and Garcia I. Tumor necrosis factor neutralization combined with chemotherapy enhances Mycobacterium tuberculosis clearance and reduces lung pathology. Am J Clin Exp Immunol. 2013; 2:124-134.
53. Tsenova L, Mangaliso B, Muller G, Chen Y, Freedman VH, Stirling D and Kaplan G. Use of IMiD3, a thalidomide analog, as an adjunct to therapy for experimental tuberculous meningitis. Antimicrob Agents Chemother. 2002; 46:1887-1895.
54. Canan CH, Gokhale NS, Carruthers B, Lafuse WP, Schlesinger LS, Torrelles JB and Turner J. Characterization of lung inflammation and its impact on macrophage function in aging. J Leukoc Biol. 2014; 96:473-480.
55. Vilaplana C, Marzo E, Tapia G, Diaz J, Garcia V and Cardona PJ. Ibuprofen therapy resulted in significantly decreased tissue bacillary loads and increased survival in a new murine experimental model of active tuberculosis. J Infect Dis. 2013; 208:199-202.
56. Mayer-Barber KD, Andrade BB, Oland SD, Amaral EP, Barber DL, Gonzales J, Derrick SC, Shi R, Kumar NP, Wei W, Yuan X, Zhang G, Cai Y, Babu S, Catalfamo M, Salazar AM, et al. Host-directed therapy of tuberculosis based on interleukin-1 and type I interferon crosstalk. Nature. 2014; 511:99-103.
57. Parihar SP, Guler R, Khutlang R, Lang DM, Hurdayal R, Mhlanga MM, Suzuki H, Marais AD and Brombacher F. Statin therapy reduces the mycobacterium tuberculosis burden in human macrophages and in mice by enhancing autophagy and phagosome maturation. J Infect Dis. 2014; 209:754-763.
58. Skerry C, Pinn ML, Bruiners N, Pine R, Gennaro ML and Karakousis PC. Simvastatin increases the in vivo activity of the first-line tuberculosis regimen. J Antimicrob Chemother. 2014; 69:2453-2457.
59. Rajaram MV, Brooks MN, Morris JD, Torrelles JB, Azad AK and Schlesinger LS. Mycobacterium tuberculosis activates human macrophage peroxisome proliferator-activated receptor gamma linking mannose receptor recognition to regulation of immune responses. J Immunol. 2010; 185:929-942.
60. Salahuddin N, Ali F, Hasan Z, Rao N, Aqeel M and Mahmood F. Vitamin D accelerates clinical recovery from tuberculosis: results of the SUCCINCT Study [Supplementary Cholecalciferol in recovery from tuberculosis]. A randomized, placebo-controlled, clinical trial of vitamin D supplementation in patients with pulmonary tuberculosis’. BMC Infect Dis. 2013; 13:22.
61. Daley P, Jagannathan V, John KR, Sarojini J, Latha A, Vieth R, Suzana S, Jeyaseelan L, Christopher DJ, Smieja M and Mathai D. Adjunctive vitamin D for treatment of active tuberculosis in India: a randomised, double-blind, placebo-controlled trial. Lancet Infect Dis. 2015; 15:528-534.
62. Gao XF, Yang ZW and Li J. Adjunctive therapy with interferon-gamma for the treatment of pulmonary tuberculosis: a systematic review. Int J Infect Dis. 2011; 15:e594-600.
63. Maiga M, Ammerman NC, Maiga MC, Tounkara A, Siddiqui S, Polis M, Murphy R and Bishai WR. Adjuvant host-directed therapy with types 3 and 5 but not type 4 phosphodiesterase inhibitors shortens the duration of tuberculosis treatment. J Infect Dis. 2013; 208:512-519.
64. Walker NF, Clark SO, Oni T, Andreu N, Tezera L, Singh S, Saraiva L, Pedersen B, Kelly DL, Tree JA, D’Armiento JM, Meintjes G, Mauri FA, Williams A, Wilkinson RJ, Friedland JS, et al. Doxycycline and HIV infection suppress tuberculosis-induced matrix metalloproteinases. Am J Respir Crit Care Med. 2012; 185:989-997.
65. Skrahin A, Ahmed RK, Ferrara G, Rane L, Poiret T, Isaikina Y, Skrahina A, Zumla A and Maeurer MJ. Autologous mesenchymal stromal cell infusion as adjunct treatment in patients with multidrug and extensively drug-resistant tuberculosis: an open-label phase 1 safety trial. Lancet Respir Med. 2014; 2:108-122.
66. Wallis RS. Corticosteroid effects on sputum culture in pulmonary tuberculosis: a meta-regression analysis. Open Forum Infect Dis. 2014; 1:ofu020.
67. Mayanja-Kizza H, Jones-Lopez E, Okwera A, Wallis RS, Ellner JJ, Mugerwa RD and Whalen CC. Immunoadjuvant prednisolone therapy for HIV-associated tuberculosis: a phase 2 clinical trial in Uganda. J Infect Dis. 2005; 191:856-865.
68. Silmon de Monerri NC and Kim K. Pathogens hijack the epigenome: a new twist on host-pathogen interactions. Am J Pathol. 2014; 184:897-911.
69. Roy S, Schmeier S, Arner E, Alam T, Parihar SP, Ozturk M, Tamgue O, Kawaji H, de Hoon MJ, Itoh M, Lassmann T, Carninci P, Hayashizaki Y, Forrest AR, Bajic VB, Guler R, et al. Redefining the transcriptional regulatory dynamics of classically and alternatively activated macrophages by deepCAGE transcriptomics. Nucleic Acids Res. 2015.
70. Guler R, Parihar SP, Savvi S, Logan E, Schwegmann A, Roy S, Nieuwenhuizen NE, Ozturk M, Schmeier S, Suzuki H and Brombacher F. IL-4Ralpha-Dependent Alternative Activation of Macrophages Is Not Decisive for Mycobacterium tuberculosis Pathology and Bacterial Burden in Mice. PLoS One. 2015; 10:e0121070.
71. Zhou RJ, Shi Z, Zhou K, Wang HD, Zhang GQ, Li XT and Xu JP. Decreased SARI expression predicts poor prognosis of Chinese patients with non-small cell lung cancer. Int J Clin Exp Pathol. 2013; 6:2056-2063.
72. Dash R, Su ZZ, Lee SG, Azab B, Boukerche H, Sarkar D and Fisher PB. Inhibition of AP-1 by SARI negatively regulates transformation progression mediated by CCN1. Oncogene. 2010; 29:4412-4423.
73. Wang C, Su Y, Zhang L, Wang M, You J, Zhao X, Zhang Z, Liu J and Hao X. The function of SARI in modulating epithelial-mesenchymal transition and lung adenocarcinoma metastasis. PLoS One. 2012; 7:e38046.
74. Liu Z, Wei P, Yang Y, Cui W, Cao B, Tan C, Yu B, Bi R, Xia K, Chen W, Wang Y, Zhang Y, Du X and Zhou X. BATF2 Deficiency Promotes Progression in Human Colorectal Cancer via Activation of HGF/MET Signaling: A Potential Rationale for Combining MET Inhibitors with IFNs. Clin Cancer Res. 2015; 21:1752-1763.
75. Ma H, Liang X, Chen Y, Pan K, Sun J, Wang H, Wang Q, Li Y, Zhao J, Li J, Chen M and Xia J. Decreased expression of BATF2 is associated with a poor prognosis in hepatocellular carcinoma. Int J Cancer. 2011; 128:771-777.
76. Wen H, Chen Y, Hu Z, Mo Q, Tang J and Sun C. Decreased expression of BATF2 is significantly associated with poor prognosis in oral tongue squamous cell carcinoma. Oncol Rep. 2014; 31:169-174.