
INTRODUCTION
Lenalidomide (CC-5013, Revlimid) has been approved by the Food and Drug Administration to treat patients with multiple myeloma because it has potent anti-myeloma activities and a better toxicity profile than thalidomide. Lenalidomide's anti-myeloma mechanisms include tumoricidal, immunomodulatory, anti-inflammatory, and anti-angiogenic properties. The direct tumoricidal effect of lenalidomide occurs through caspase activation and apoptosis induction. [1] It also disrupts the stromal cell-enriched tumor microenvironment and inhibits angiogenesis. [2, 3] Lenalidomide enhances immune cell function not only by activating T and NK cells and increasing their cytokine secretion but also by decreasing IL-6 secretion in vitro. [4, 5] Lenalidomide synergistically enhances antibody-dependent cell-mediated cytotoxicity in myeloma cells. [6] All in vitro preclinical studies suggest these dual tumoricidal and immunomodulatory activities for lenalidomide. However, the principal action of lenalidomide in the anti-myeloma immune response in vivo is unclear.
The 5TGM1 myeloma cell line was initially derived from a murine myeloma cell line, 5T, that originated spontaneously from C57BL/KaLwRij mice [7]. After i.v. injection of 5TGM1 cells into C57BL/KaLwRij immunocompetent mice, 5TGM1 myeloma cells thrived and migrated to bone marrow. Similar to myeloma patients, the 5TGM1 myeloma mouse model presented with monoclonal gammopathy and demonstrated marrow replacement, focal osteolytic bone lesions, hind limb paralysis, and occasional hypercalcemia [8].
Our preliminary data showed that 5TGM1 cells were resistant to lenalidomide in vitro and in severe combined immunodeficiency (SCID) mice but were sensitive to lenalidomide in an immune response-dependent manner in immunocompetent C57BL/KaLwRij mice in vivo. Therefore, 5TGM1-bearing C57BL/KaLwRij mice were taken as an ideal model for investigating the anti-myeloma immune action of lenalidomide in vivo. Here, we found that lenalidomide failed to prolong the survival of mice after CD4+ T cell but not CD8+ T cell, B cell, or NK cell depletion and this was associated with a stronger Th1 response. These results suggest that CD4+ T cells are indispensable to a lenalidomide-mediated anti-myeloma immune response in vivo.
RESULTS
5TGM1-bearing C57BL/KaLwRij mouse is an ideal model to study the immunomodulatory effect of lenalidomide in vivo
Due to the complicated antitumor activity of lenalidomide, separating its effects into tumoricidal and immunomodulatory activities is difficult, which hampers study of its direct immunomodulatory effects in vivo. We wanted a murine myeloma model that is resistant to lenalidomide's tumoricidal effect but sensitive to its immunomodulatory effect. Through in vitro treatment with lenalidomide of different myeloma cell lines and analysis of proliferation and apoptosis (data not shown), we decided to focus on 5TGM1 murine myeloma cells. Lenalidomide at concentrations up to 100 μM for 72 hours didn’t induce growth inhibition or apoptosis in 5TGM1 myeloma cells (Figure 1).
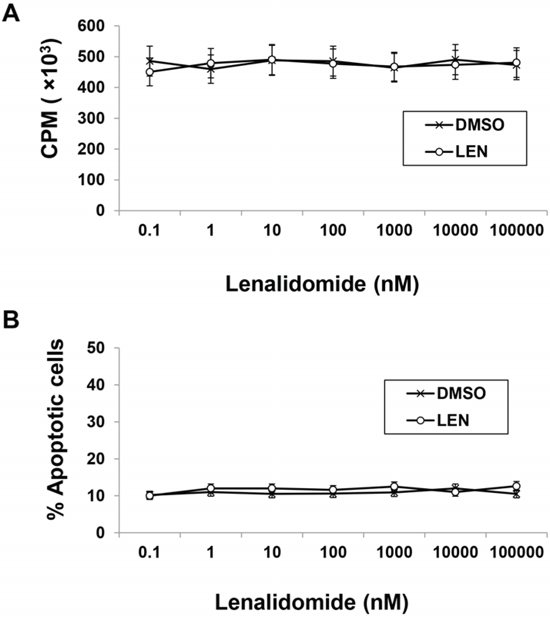
Figure 1: Murine myeloma 5TGM1 cells are resistant to lenalidomide in vitro. 5TGM1 cells were cultured for 72 hours in the presence of the indicated concentrations of lenalidomide or equal volume of DMSO (vehicle control). A. Cell proliferation was assessed by 3H-thymidine incorporation assay. B. Apoptosis was assessed by annexin V and PI staining. Results of three independent experiments are shown. LEN, lenalidomide; DMSO, dimethyl sulfoxide.
A 7-day culture with 100 μM lenalidomide did not affect 5TGM1 growth or apoptosis (data not shown). Because lenalidomide had no direct cytotoxicity on 5TGM1 cells in vitro, we next determined whether lenalidomide had any effect on the tumor cells in vivo. As shown in Figure 2A–2C, lenalidomide inhibited tumor growth and prolonged survival of 5TGM1-bearing C57BL/KaLwRij mice. Notably, 2/12 mice treated with lenalidomide were tumor-free at post-inoculation day 90 (Figure 2B–2C, P < 0.05). However, in immunodeficient B6-SCID mice, which lack T and B cells, lenalidomide treatment failed to inhibit tumor growth (Figure 2D–2E, P > 0.05) or prolong survival of tumor-bearing mice (Figure 2F, P > 0.05). That lenalidomide had no direct tumoricidal effect on 5TGM1 cells in vitro and inhibited myeloma growth in immunocompetent but not immunodeficient mice indicates that the host immune system must play an important role in the anti-myeloma activity of lenalidomide in vivo and this activity can be studied in the 5TGM1-bearing C57BL/KaLwRij model.
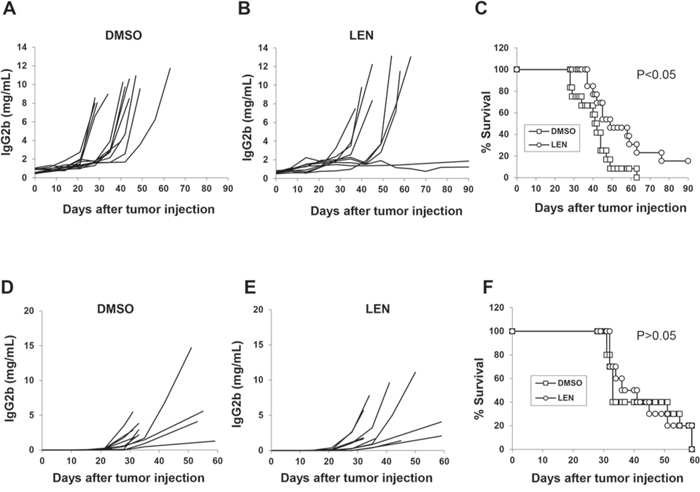
Figure 2: In vivo effect of lenalidomide in myeloma-bearing mice. C57BL/KaLwRij (A–C, 12 mice per group) or B6-SCID (D–F, 10 per group) mice were challenged with 2 × 106 5TGM1 cells via intravenous injection. After 1 week, mice received intraperitoneal injections of lenalidomide (25 mg/kg/day) or equal volume of DMSO for 21 consecutive days. Serum samples were collected weekly, and tumor burden was monitored by measuring circulating IgG2b M-protein. Concentration curves of serum IgG2b M-protein from mice receiving DMSO as vehicle control A and D. or lenalidomide B and E. C and F. Mouse survival curves. LEN, lenalidomide.
NK cells are not the major effector cells for anti-myeloma activity of lenalidomide in vivo
Therefore, we first investigated the importance of NK cells in lenalidomide-mediated anti-myeloma activity in vivo. Our study in B6-SCID mice showed that lenalidomide had no obvious anti-myeloma effects in vivo (Figure 2D–2F). As these SCID mice have functional NK cells but no T and B cells, this result suggested that NK cells may not be important for lenalidomide-mediated anti-myeloma activity in vivo. After NK cells were depleted in 5TGM1-bearing C57BL/KaLwRij mice, lenalidomide still retarded tumor growth and prolonged survival (Figure 3, P < 0.05). Together with the finding that lenalidomide had an anti-myeloma effect in immunocompetent but not in B6-SCID mice, which have NK cells, these results demonstrated that NK cells are not the main effector cells of lenalidomide action in vivo.
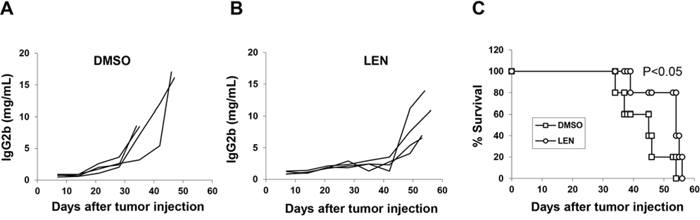
Figure 3: The role of NK cells in the lenalidomide-mediated immunomodulatory effect in 5TGM1-bearing C57BL/KaLwRij mice. Mice were treated as described in Figure 2. After receiving 5TGM1 cells, mice were injected with asialo-GM1 antibody twice per week until the end of drug treatment to deplete NK cells. Concentration curve for serum IgG2b M-protein in NK-depleted C57BL/KaLwRij mice receiving DMSO as vehicle control A. or lenalidomide B, C. Mouse survival curve of NK-depleted C57BL/KaLwRij mice receiving DMSO or lenalidomide (4 mice per group).
CD4+ T cells mediate the pivotal anti-myeloma activity of lenalidomide in vivo
Next, we investigated whether the lenalidomide immunomodulatory effect depends on T or B cell activity. Depleting CD4+T cells significantly enhanced tumor growth and shortened survival (Figure 4A, 4B and 4E; P < 0.01, vs. isotype control). Depleting CD8+ T cells or B cells did not significantly affect tumor growth or survival (Figure 4A, 4C, 4D and 4E, P > 0.05, vs. isotype control). These in vivo results demonstrated that CD4+ T cells but not CD8+ or B cells are crucial in the lenalidomide-mediated anti-myeloma immune response in vivo.
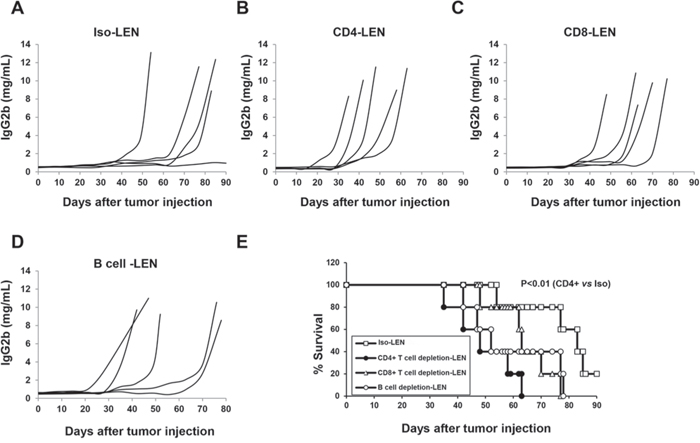
Figure 4: The role of CD4+, CD8+ T and B cells in the lenalidomide-mediated immunomodulatory effect in 5TGM1-bearing C57BL/KaLwRij mice. The same procedures as described in Figures 2 and 3 were used for NK cell depletion, lenalidomide treatment and measurement. CD4+ T cells, CD8+ T cells, and B cells were depleted separately in 5TGM1-bearing C57BL/KaLwRij mice. Concentration curves for serum IgG2b M-protein in 5TGM1-bearing C57BL/KaLwRij mice with isotype IgG A. CD4+ T cell-depleted 5TGM1-bearing C57BL/KaLwRij mice B. CD8+ T cell-depleted 5TGM1-bearing C57BL/KaLwRij mice C. and B cell-depleted 5TGM1-bearing C57BL/KaLwRij mice D, E. Survival curves for CD4+ T cell-, CD8+ T cell- or B cell-depleted 5TGM1-bearing C57BL/KaLwRij mice versus 5TGM1-bearing C57BL/KaLwRij mice without depletion (5 mice per group).
Lenalidomide facilitates a T-cell dependent type I response in 5TGM1-bearing mice
To further understand how lenalidomide affects immune cell function in vivo, 5TGM1-bearing mice were sacrificed 1 week after the final lenalidomide injection and the splenocytes were harvested, analyzed directly or restimulated ex vivo (see below) before assay.
First the percentages of splenic CD4+ T cells, CD8+ T cells, NK cells, and B cells were analyzed by flow cytometry. As Figure 5A shows, the percentages of both CD4+ T cells and CD8+ T cells increased about 2-fold vs. vehicle control (P < 0.01). NK cells and B cells showed no change (P > 0.05).
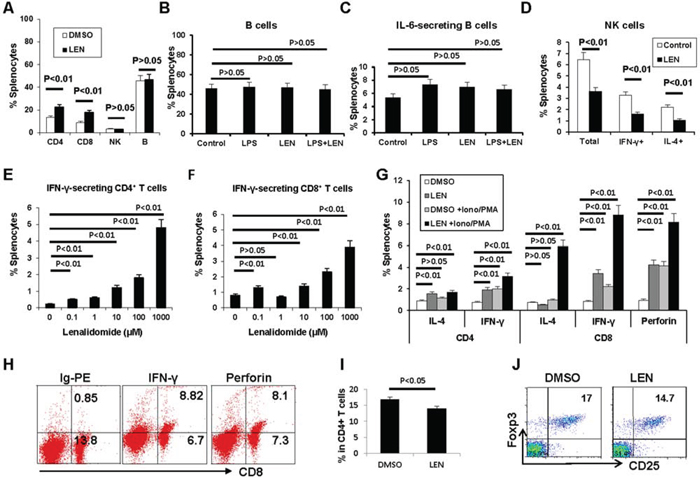
Figure 5: Lenalidomide promotes the expansion of T cells in 5TGM1-bearing C57BL/KaLwRij mice. Splenocytes from myeloma-bearing C57BL/KaLwRij mice were analyzed directly (A) or restimulated ex vivo for 72 hours (B–J) Percentages of A. CD4+ T cells, CD8+ T cells, NK cells, and B cells, B-C. B cells and IL-6 secreting B cells, D. NK cells including IFN-γ-secreting and IL-4-secreting NK cells, E. IFN-γ-secreting CD4+ T cells, and F. IFN-γ-secreting CD8+ T cells. G. The synergistic effect of lenalidomide with PMA/ionomycin on the activation of CD4+ T cells and CD8+ T cells. H. Representative flow cytometry results showing CD8+ T cell activation. I. Percentage of CD25+Foxp3+ T cells after restimulation with lenalidomide. J. Representative flow cytometry results for CD25+Foxp3+ T cells. PMA, phorbol 12-myristate 13-acetate; Ion, ionomycin; LPS, lipopolysaccharide; Ig-PE, R-phycoerythrin-conjugated rat immunoglobulin.
After restimulation of splenocytes with lenalidomide for 72 hours ex vivo, the B cell population showed no significant change even when treatment was combined with LPS (Figure 5B–5C; P > 0.05). The total number of NK cells, including IFN-γ-secreting and IL-4-secreting NK cells, decreased (Figure 5D; P < 0.01).
We found that lenalidomide significantly increased IFN-γ secretion in both CD4+ and CD8+ T cells in 5TGM1-bearing mice (Figure 5E–5F). When lenalidomide was combined with PMA/ionomycin, synergistic effects on IFN-γ production by CD4+ and CD8+ T cells and expression of perforin by CD8+ T cells were observed (Figure 5G–5H). Interestingly, lenalidomide and PMA/ionomycin also stimulated CD8+ but not CD4+ T cells to secrete significantly more IL-4 (Figure 5G). Lenalidomide alone slightly decreased the percentage of CD25+Foxp3+CD4+ regulatory T (Treg) cells (Figure 5I). Taken together, these data clearly showed that lenalidomide enhances the type I anti-myeloma CD4+ and CD8+ T cell responses in vivo.
DISCUSSION
Lenalidomide, a second-generation immunomodulatory drug, exhibits antitumor activity. Lenalidomide-containing therapeutic regimens resulted in higher response and improved outcomes in the patients with MM relapse or with refractory MM [10, 11]. Previous studies demonstrated that lenalidomide had a direct tumoricidal role by inducing apoptosis and blocking tumor cell proliferation [12, 13]. Recent in vitro studies showed that lenalidomide bound to cereblon and then selectively degraded specific transcription factors — the Ikaros family zinc finger proteins 1 and 3 (IKZF1 and IKZF3) — which were necessary for myeloma survival [14, 15]. However, it is essential to investigate how lenalidomide modulates the immune response in vivo. Considering the complicated effects of lenalidomide on tumor cells and immune cells, the mechanistic study of immunomodulation is limited by the availability of a suitable tumor model that can separate these two effects. In the current study, we found that a murine myeloma cell line, 5TGM1, was totally resistant to lenalidomide toxicity in vitro. Most importantly, we found that lenalidomide inhibited tumor growth and prolonged the survival of 5TGM1-bearing C57BL/KaLwRij mice; however, it failed to inhibit tumor growth of 5TGM1-bearing SCID mice. Those results demonstrated that lenalidomide affect 5TGM1-bearing mice survival in an immune response-dependent manner only. This 5TGM1 myeloma model facilitates study of the immunomodulatory role without the direct tumoricidal effect of lenalidomide in vivo.
By depleting different cell populations in the 5TGM1-bearing immunocompetent mice separately, we found that CD4+ T cells play a critical role in the lenalidomide-mediated antitumor immune response. Under different types of stimulation, CD4+ T cells will differentiate into different populations, including Th1, Th2, Th9, Th17, TfH, and regulatory T cells, to mediate different immune responses. Lenalidomide potentiated T cell activation in a costimulatory manner in vitro [16]. IL-2 is T cell activation hallmark and its transcription was suppressed by IKZF3, which bound to the IL-2 promoter directly [17]. Lenalidomide promoted IKZF3 degradation, thus releasing the suppressive effect of IKZF3 on IL-2 transcription and promoting T cell activation [14, 15].
Our data showed that lenalidomide enhanced IFN-γ production in CD4+ T cell in vitro and in vivo, suggesting an increased Th1 response after lenalidomide treatment. Th1 polarized immune responses are associated with CD8+ T cell-mediated cytotoxicity against tumor cells and induce tumor regression. After lenalidomide treatment, CD8+ T cells produced more IFN-γ and perforin, which is consistent with the enhanced Th1 response. The cytotoxic effect of CD8+ T cells can be inhibited through recruitment and/or conversion of natural or adaptive Tregs. Our data showed that the number of Tregs was somewhat decreased in tumor-bearing C57BL/KaLwRij mice after lenalidomide treatment. An in vitro study by Luptakova et al demonstrated that lenalidomide resulted in T cell polarization toward a Th1 phenotype characterized by increased IFN-γ secretion of T cells while decreasing the number of Tregs [18], which is consistent with our in vivo data.
The exact molecular mechanism by which lenalidomide regulates CD4+ T cell activation and polarization is not clear. In the absence of CD28 stimulation, lenalidomide induced IL-2, IFN-γ and TNF-α secretion [19], suggesting that lenalidomide may activate T cells by mimicking the costimulatory signaling. Furthermore, LeBlance et al found that lenalidomide increased tyrosine phosphorylation in the CD28 receptor intracellular domain in the absence of costimulatory molecules [20].
Although cereblon is essential for lenalidomide-mediated direct tumoricidal activity against myeloma, it also mediated T cell activation by lenalidomide or its analogues because knockdown of cereblon in primary human T cells abrogates drug-induced IL-2 expression [21]. Furthermore, lenalidomide promoted the cereblon-dependent destruction of Ikaros proteins [15]. IKZF1 (Ikaros) directly bound to the promoter region of T-bet, the master regulator of Th1 cell differentiation, and suppressed IFN-γ production and promoted Th2 polarization [22]. IKZF2 (Helios) directly bound to Foxp3 promoter and up-regulated Foxp3 expression, which results Treg differentiation [23]. IL-2 transcription was suppressed by IKZF3 through binding of its promoter. Thus, although it is not known whether lenalidomide can degrade IZKF2 as well as IZKF1 and IZKF3, it seems that lenalidomide can promote T cell activation by down-regulation of IZKF1, polarize Th1 response by degradation of IZKF3, and may slightly decrease Treg by down-regulation of the Ikaros family.
Previous studies showed that lenalidomide activated NK cells and enhanced NK cell function in MM patients, and its effects on NK cells were dependent on CD4+T cells and IL-2 secretion [24, 25]. Our results demonstrated that NK cells were not a therapeutic target of lenalidomide. Therefore, lenalidomide may affect NK cell function in the presence of CD4+ T cells.
Our study demonstrates that CD4+ T cells are critical to the lenalidomide-mediated antitumor immune response in vivo and the number of CD4+ T cells was significantly increased. Therefore it is necessary to analyze whether there is a positive correlation between the number of CD4+ T cells and response to lenalidomide in myeloma patients.
MATERIALS AND METHODS
Mice and cell lines
Male C57BL/KaLwRij mice were purchased from Harlan, and CPB.B6.CB17-PrkdcSCID/Szj (B6 SCID) mice were purchased from Jackson Laboratory. The Institutional Animal Care and Use Committee of the University of Texas M.D. Anderson Cancer Center approved the study. The 5TGM1 myeloma cell line was derived from 5T murine myeloma that spontaneously arose in C57BL/KaLwRij mice and was cultured in Iscove's modified Dulbecco's medium (Invitrogen) supplemented with 10% heat-inactivated fetal bovine serum (Atlanta Biologicals), 100 units/mL penicillin-streptomycin, and 2 mmol/L L-glutamine (Invitrogen).
Drugs and reagents
Lenalidomide was provided by Celgene (Summit, NJ) and dissolved in dimethyl sulfoxide (DMSO) at a stock concentration of 0.5 M. FITC-conjugated annexin V was purchased from Caltag Laboratories (Burlingame, CA). Propidium iodide (PI), lipopolysaccharide (LPS), ionomycin, and phorbol 12-myristate 13-acetate (PMA) were purchased from Sigma-Aldrich (St. Louis, MO). Fetal bovine serum was purchased from Atlanta Biologicals (Norcross, CA). 3H-thymidine was purchased from Amersham (Arlington Heights, IL).
In vivo therapeutic effect of lenalidomide
Mice were challenged intravenously with 2 × 106 5TGM1 myeloma cells. Three weeks later, when myeloma growth was established, mice received intraperitoneal lenalidomide (25 mg/kg) or equal volume vehicle control daily for 21 consecutive days. Tumor burden was monitored by measuring serum IgG2b M-protein by ELISA. Mice were euthanized when moribund or when hind-leg paralysis developed.
Cell proliferation assay
The inhibitory growth effect of lenalidomide on myeloma cells was assessed by 3H-thymidine incorporation assay. Briefly, cells were plated in 96-well plates at a density of 5 × 104 cells/well and treated with different concentrations of lenalidomide for 72 hours. 3H-thymidine, 1 μCi, was added to each well, and the cells were incubated for an additional 16 hours. The cells were washed, and the radioactivity was measured using a scintillation beta-counter (PerkinElmer Life and Analytical Sciences, Shelton, CT). The data are expressed as the percentage of the DMSO control value.
Apoptosis assay
An annexin V-binding assay was used to detect the induction of apoptosis. Cells were seeded in 48-well plates with different concentrations of lenalidomide for 72 hours. To quantify the percentage of cells undergoing apoptosis, cells were washed twice with cold phosphate-buffered saline (PBS) and then resuspended in 1X binding buffer at 1 × 106 cells/mL. Subsequently, 100 μl cell suspension solution was transferred into a 5-mL tube and 5 μl annexin V and PI were added. The tubes were gently vortexed and incubated for 15 minutes at room temperature in the dark. Then 300 μl 1X binding buffer was added and analyzed immediately using a FACScan flow cytometer (Becton Dickinson, San Jose, CA). The number of apoptotic cells was determined by counting annexin V-positive cells.
Enzyme-linked immunosorbent assay (ELISA)
5TGM1 myeloma cells secrete mouse immunoglobulin IgG2b Id protein. The level of mouse circulating IgG2b Id protein was used to monitoring tumor burden. As described previously [9], mouse serum was collected at the indicated times, and the amounts of secreted IgG2b Id protein in mouse serum were quantified by ELISA.
Flow cytometry
An aliquot of freshly isolated cells from the spleen of lenalidomide/DMSO treated C57BL/KaLwRij mice was incubated with fluorescent isothiocyanate (FITC)-, phycoerythrin (PE)-, or allophycocyanin (APC)-conjugated monoclonal antibodies (mAbs) against CD4, CD8, B220, or NK1.1 (eBioscience, San Diego, CA) for 30 minutes at 4°C, and then washed twice and analyzed by flow cytometry. Another aliquot of freshly isolated cells was incubated with 1 μM lenalidomide or DMSO (vehicle control) with and without LPS (100 ng/mL) or ionomycin (1 μg/mL) plus PMA (300 ng/mL) at 37°C for 72 hours. These cells were then subjected to CD4, CD8, B220, or NK1.1 cell surface staining, and then treated using a Cytofix/Cytoperm kit (BD Biosciences) for intracellular staining for IL-4, IFN-γ, perforin or Foxp3 (eBioscience, San Diego, CA).
In vivo T cell, B cell, and NK cell depletion
Hybridomas producing anti-CD4 mAb GK1.5 (rat IgG2b) and anti-CD8 mAb 2.43 (rat IgG2b) were obtained from the American Type Culture Collection to deplete CD4 and CD8 cells. Antibodies were purified using HiTrap Protein D columns (GE Healthcare). Anti-mouse B220 mAb was purchased from R&D Systems to deplete B cells. Anti-mouse asialo-ganglio N-tetraoglyceramide (asislo-GM1) polyclonal antibody (Cedarlane, Canada) was used to deplete NK cells. Cell depletion was performed by intraperitoneal injection of 100 μg of antibody against CD8, CD4, B220, or asislo-GM1 twice per week before and during treatment. The efficiency of cell depletion was assessed by staining peripheral blood mononuclear cells with CD4-PE, CD8-FITC, B220-PE, or NK1.1-FITC (eBioscience).
Statistical analysis
All assays were performed in triplicate and expressed as mean ± SE. Statistical significance was determined by Student's t test. Mouse overall survival was determined using the Kaplan-Meier method. P values < 0.05 were considered significant.
ACKNOWLEDGMENTS AND FUNDING
We thank Celgene for providing the research drug and funding. We thank C. Talerico, PhD, for providing editorial suggestions and comments.
CONFLICTS OF INTEREST
The authors declare no competing financial interests.
GRANT SUPPORT
This work was supported by National Cancer Institute R01 CA138402, R01 CA138398, R01 CA163881, and P50 CA142509 to Dr. Qing Yi, and NSFC81372545 to Dr. Zheng.
Authors’ contributions
QY, LZ, and EB initiated the work, designed the studies, and wrote the paper. LZ, EB, SH, and JQ performed the experiments and statistical analyses. MW and CZ provided critical suggestions.
REFERENCES
1. Tai YT, Li XF, Catley L, Coffey R, Breitkreutz I, Bae J, Song W, Podar K, Hideshima T, Chauhan D, Schlossman R, Richardson P, Treon SP, Grewal IS, Munshi NC, Anderson KC. Immunomodulatory drug lenalidomide (CC-5013, IMiD3) augments anti-CD40 SGN-40-induced cytotoxicity in human multiple myeloma: clinical implications. Cancer research. 2005; 65:11712–11720.
2. Breitkreutz I, Raab MS, Vallet S, Hideshima T, Raje N, Mitsiades C, Chauhan D, Okawa Y, Munshi NC, Richardson PG, Anderson KC. Lenalidomide inhibits osteoclastogenesis, survival factors and bone-remodeling markers in multiple myeloma. Leukemia. 2008; 22:1925–1932.
3. De Luisi A, Ferrucci A, Coluccia AM, Ria R, Moschetta M, de Luca E, Pieroni L, Maffia M, Urbani A, Di Pietro G, Guarini A, Ranieri G, Ditonno P, Berardi S, Caivano A, Basile A, et al. Lenalidomide restrains motility and overangiogenic potential of bone marrow endothelial cells in patients with active multiple myeloma. Clinical cancer research: an official journal of the American Association for Cancer Research. 2011; 17:1935–1946.
4. Lee BN, Gao H, Cohen EN, Badoux X, Wierda WG, Estrov Z, Faderl SH, Keating MJ, Ferrajoli A, Reuben JM. Treatment with lenalidomide modulates T-cell immunophenotype and cytokine production in patients with chronic lymphocytic leukemia. Cancer. 2011; 117:3999–4008.
5. Wu L, Adams M, Carter T, Chen R, Muller G, Stirling D, Schafer P, Bartlett JB. lenalidomide enhances natural killer cell and monocyte-mediated antibody-dependent cellular cytotoxicity of rituximab-treated CD20+ tumor cells. Clinical cancer research: an official journal of the American Association for Cancer Research. 2008; 14:4650–4657.
6. Tai YT, Horton HM, Kong SY, Pong E, Chen H, Cemerski S, Bernett MJ, Nguyen DH, Karki S, Chu SY, Lazar GA, Munshi NC, Desjarlais JR, Anderson KC, Muchhal US. Potent in vitro and in vivo activity of an Fc-engineered humanized anti-HM1.24 antibody against multiple myeloma via augmented effector function. Blood. 2012; 119:2074–2082.
7. Radl J, De Glopper ED, Schuit HR, Zurcher C. Idiopathic paraproteinemia. II. Transplantation of the paraprotein-producing clone from old to young C57BL/KaLwRij mice. Journal of immunology. 1979; 122:609–613.
8. Asosingh K, Radl J, Van Riet I, Van Camp B, Verkerken K. The 5TMM series: a useful in vivo mouse model of human multiple myeloma. The hematology journal: the official journal of the European Haematology Association / EHA. 2000; 1:351–356.
9. Wang S, Hong S, Wezeman M, Qian J, Yang J, Yi Q. Dendritic cell vaccine but not idiotype-KLH protein vaccine primes therapeutic tumor-specific immunity against multiple myeloma. Frontiers in bioscience: a journal and virtual library. 2007; 12:3566–3575.
10. Palumbo A, Attal M, Roussel M. Shifts in the therapeutic paradigm for patients newly diagnosed with multiple myeloma: maintenance therapy and overall survival. Clinical cancer research: an official journal of the American Association for Cancer Research. 2011; 17:1253–1263.
11. Palumbo A, Hajek R, Delforge M, Kropff M, Petrucci MT, Catalano J, Gisslinger H, Wiktor-Jedrzejczak W, Zodelava M, Weisel K, Cascavilla N, Iosava G, Cavo M, Kloczko J, Blade J, Beksac M, et al. Continuous lenalidomide treatment for newly diagnosed multiple myeloma. The New England journal of medicine. 2012; 366:1759–1769.
12. Verhelle D, Corral LG, Wong K, Mueller JH, Moutouh-de Parseval L, Jensen-Pergakes K, Schafer PH, Chen R, Glezer E, Ferguson GD, Lopez-Girona A, Muller GW, Brady HA, Chan KW. Lenalidomide and CC-4047 inhibit the proliferation of malignant B cells while expanding normal CD34+ progenitor cells. Cancer research. 2007; 67:746–755.
13. Gandhi AK, Kang J, Capone L, Parton A, Wu L, Zhang LH, Mendy D, Lopez-Girona A, Tran T, Sapinoso L, Fang W, Xu S, Hampton G, Bartlett JB, Schafer P. Dexamethasone synergizes with lenalidomide to inhibit multiple myeloma tumor growth, but reduces lenalidomide-induced immunomodulation of T and NK cell function. Current cancer drug targets. 2010; 10:155–167.
14. Kronke J, Udeshi ND, Narla A, Grauman P, Hurst SN, McConkey M, Svinkina T, Heckl D, Comer E, Li X, Ciarlo C, Hartman E, Munshi N, Schenone M, Schreiber SL, Carr SA, et al. Lenalidomide causes selective degradation of IKZF1 and IKZF3 in multiple myeloma cells. Science. 2014; 343:301–305.
15. Lu G, Middleton RE, Sun H, Naniong M, Ott CJ, Mitsiades CS, Wong KK, Bradner JE, Kaelin WG Jr. The myeloma drug lenalidomide promotes the cereblon-dependent destruction of Ikaros proteins. Science. 2014; 343:305–309.
16. Haslett PA, Corral LG, Albert M, Kaplan G. Thalidomide costimulates primary human T lymphocytes, preferentially inducing proliferation, cytokine production, and cytotoxic responses in the CD8+ subset. The Journal of experimental medicine. 1998; 187:1885–1892.
17. Quintana FJ, Jin H, Burns EJ, Nadeau M, Yeste A, Kumar D, Rangachari M, Zhu C, Xiao S, Seavitt J, Georgopoulos K, Kuchroo VK. Aiolos promotes TH17 differentiation by directly silencing Il2 expression. Nature immunology. 2012; 13:770–777.
18. Luptakova K, Rosenblatt J, Glotzbecker B, Mills H, Stroopinsky D, Kufe T, Vasir B, Arnason J, Tzachanis D, Zwicker JI, Joyce RM, Levine JD, Anderson KC, Kufe D, Avigan D. Lenalidomide enhances anti-myeloma cellular immunity. Cancer immunology, immunotherapy: CII. 2013; 62:39–49.
19. Schafer PH, Gandhi AK, Loveland MA, Chen RS, Man HW, Schnetkamp PP, Wolbring G, Govinda S, Corral LG, Payvandi F, Muller GW, Stirling DI. Enhancement of cytokine production and AP-1 transcriptional activity in T cells by thalidomide-related immunomodulatory drugs. The Journal of pharmacology and experimental therapeutics. 2003; 305:1222–1232.
20. LeBlanc R, Hideshima T, Catley LP, Shringarpure R, Burger R, Mitsiades N, Mitsiades C, Cheema P, Chauhan D, Richardson PG, Anderson KC, Munshi NC. Immunomodulatory drug costimulates T cells via the B7-CD28 pathway. Blood. 2004; 103:1787–1790.
21. Lopez-Girona A, Mendy D, Ito T, Miller K, Gandhi AK, Kang J, Karasawa S, Carmel G, Jackson P, Abbasian M, Mahmoudi A, Cathers B, Rychak E, Gaidarova S, Chen R, Schafer PH, et al. Cereblon is a direct protein target for immunomodulatory and antiproliferative activities of lenalidomide and pomalidomide. Leukemia. 2012; 26:2326–2335.
22. Thomas RM, Chen C, Chunder N, Ma L, Taylor J, Pearce EJ, Wells AD. Ikaros silences T-bet expression and interferon-gamma production during T helper 2 differentiation. The Journal of biological chemistry. 2010; 285:2545–2553.
23. Getnet D, Grosso JF, Goldberg MV, Harris TJ, Yen HR, Bruno TC, Durham NM, Hipkiss EL, Pyle KJ, Wada S, Pan F, Pardoll DM, Drake CG. A role for the transcription factor Helios in human CD4(+)CD25(+) regulatory T cells. Molecular immunology. 2010; 47:1595–1600.
24. Hayashi T, Hideshima T, Akiyama M, Podar K, Yasui H, Raje N, Kumar S, Chauhan D, Treon SP, Richardson P, Anderson KC. Molecular mechanisms whereby immunomodulatory drugs activate natural killer cells: clinical application. British journal of haematology. 2005; 128:192–203.
25. Hsu AK, Quach H, Tai T, Prince HM, Harrison SJ, Trapani JA, Smyth MJ, Neeson P, Ritchie DS. The immunostimulatory effect of lenalidomide on NK-cell function is profoundly inhibited by concurrent dexamethasone therapy. Blood. 2011; 117:1605–1613.