
INTRODUCTION
Gastrointestinal stromal tumors (GISTs) are the most common mesenchymal tumors in the human digestive tract, representing 1–3% of gastrointestinal malignancies [1, 2]. The histogenesis, classification, diagnostic criteria, and biological behavior of GISTs have been the subject of much controversy [2]. Simply, they are typically defined as tumors whose behavior is driven by mutations in the Kit gene or PDGFRA gene [3–5]. The mechanism of activation in part of sporadic GISTs is an alteration of the structure of the receptor's extracellular or cytoplasmic domains caused by somatic mutations of the c-kit gene, which leads to dimerization and autophosphorylation of KIT with subsequent activation of signal transduction cascades in the absence of ligand binding [6–8]. Inhibition of KIT activity by a specific tyrosine kinase inhibitor, imatinib, often results in dramatic clinical responses [8, 9].
In contrast to GISTs associated with somatic mutations, little is known about inherited germline genetic risk factors. The rarity of the disease makes it a difficult subject to conduct population-based genetic research and unbiased assessment of non-genetic risk factors in any study population. An evaluation of the genetic determinants of GISTs is much more feasible, as the germline DNA of individuals does not change over time or in response to disease processes. Recently, O’Brien et al [10] only evaluated the associations between some candidate SNPs and several common types of acquired KIT and PDGFRA somatic mutations in a case-only study for the first time. However, no other research groups have published such evaluations, not to mention germline genetic associations with GISTs.
The TERT and CLPTM1L gene have been identified to be associated with carcinogenesis of at least 15 distinct cancers [11–14]. TERT promoter mutations were also detected in GIST tissues [15]. Recently, six tagSNPs on TERT and CLPTM1L region (five SNPs in the TERT gene: rs7726159, rs2853677, rs2736098, rs13172201, rs10069690; one SNP in CLPTM1L gene: rs451360) were identified, all of which influenced the risk of multiple cancers, including kinds of gastrointestinal cancers [16]. Given this evidence, our main objective was to identify whether these six tagSNPs potentially related to GIST carcinogenesis. Therefore, we first conducted this large population-based, two-stage, case-control study of GIST risk.
RESULTS
A total of 2,000 subjects were included in the current study; 600 were genotyped in Stage I and 1,400 were genotyped in Stage II (Table 1). People in the two genotyping stages were generally comparable. As expected, GIST cases were found to differ from controls in regard to known cancer risk factors: cases were more likely to have a higher education, body mass index (BMI), waist-to-hip ratio (WHR), and more likely to smokers and drinkers. Most of the GISTs were located in stomach (63.8%) or small Intestine (32.5%).
Table 1: Characteristics of the study population
Characteristics |
Stage I (N = 600) |
Stage II (N = 1,400) |
||||
Cases |
Controls |
P value |
Cases |
Controls |
P value |
|
Age (years) |
50.0 ± 4.39 |
50.5 ± 4.19 |
0.193 |
50.2 ± 3.1 |
50.1 ± 3.1 |
0.103 |
Gender (male) |
234 (78.0%) |
219 (73.0%) |
0.154 |
515(73.6%) |
514 (73.4%) |
0.952 |
Education (less than middle school) |
53(17.7%) |
34 (11.3%) |
0.028 |
87(12.4%) |
86(12.6%) |
0.998 |
Body mass index (kg/m2) |
24.1 ± 2.0 |
23.9 ± 2.22 |
0.513 |
24.3 ± 2.4 |
24.0 ± 2.3 |
0.008 |
Waist-to-hip ratio |
0.8223 ± 0.004 |
0.8200 ± 0.004 |
<0.0001 |
0.8223 ± 0.004 |
0.8199 ± 0.003 |
<0.0001 |
Regular physical activity |
83(27.7%) |
99 (33.0%) |
0.155 |
181(25.9%) |
195 (27.9%) |
0.399 |
Ever smokers |
81(27.0%) |
64(21.3%) |
0.105 |
201(28.7%) |
110(15.7%) |
<0.0001 |
Ever drinkers |
88(29.3%) |
50(16.7%) |
<0.0001 |
188(26.9%) |
153(21.9%) |
0.029 |
Tumor Location |
||||||
Stomach |
191(63.6%) |
448(64.0%) |
||||
Small Intestine |
96(32.0%) |
228 (32.6%) |
||||
Rectum |
5 (1.6%) |
9 (1.3%) |
||||
Other |
8 (2.6%) |
15 (2.1%) |
||||
Continuous variables: mean values ± standard deviation, p-value from t-tests;
Categorical variables: numbers and percentages, p-values from x2test
P value in bold means statistically significant.
A total of six tagSNPs on TERT and CLPTM1L region were included in the current study; of these, five SNPs were located in the TERT gene (rs7726159, rs2853677, rs2736098, rs13172201, rs10069690) and one SNP was located in neighboring CLPTM1L gene (rs451360). None of the six polymorphisms were found to deviate from HWE. We first evaluated the six tagSNPs in Stage I with 300 cases and 300 controls. The estimates of effect on GIST risk in Stage I, adjusted for age and gender are shown in Table 2. Three SNPs (rs7726159, rs10069690, and rs2736098) were found to have associations of significance with GIST risk. Then they were evaluated in Stage II with 700 cases and 700 controls additionally (Table 3). One SNP (rs2736098) was replicated with significance (P = 7.03 × 10−5). The difference remain significant after Bonferroni correction (P = 7.03 × 10−5 * 6 = 4.2 × 10−4). Compared with individuals with the CC genotype, the age and sex adjusted OR for developing GIST was 1.49 (95% CI 1.18–1.88) among those with the TT genotype. Under the log-additive model, each additional copy of minor allele A was associated with a 1.29-fold increased risk of GIST (OR = 1.29, 95% CI: 1.14–1.47, P = 7.03 × 10−5).
Table 2: Association between tagSNPs on TERT and CLPTM1L region and GIST risk (Stage I)
SNP |
Allelesa |
MAFb |
AB OR (95% CI) |
BB OR (95% CI) |
B vs A OR (95% CI) |
P |
rs7726159 |
C/A |
0.33 |
1.32 (0.93–1.87) |
1.67 (1.04–2.68) |
1.33 (1.05–1.69) |
0.017 |
rs2853677 |
A/G |
0.41 |
0.94 (0.66–1.34) |
1.72 (0.41–1.24) |
0.89 (0.71–1.13) |
0.894 |
rs2736098 |
C/T |
0.37 |
1.49 (1.03–2.14) |
1.78 (1.17–2.72) |
1.44 (1.14–1.81) |
1.92 × 10−3 |
rs13172201 |
C/T |
0.25 |
1.18 (0.82–1.69) |
1.24 (0.75–2.05) |
1.17 (0.90–1.50) |
0.241 |
rs10069690 |
C/T |
0.16 |
1.36 (0.96–1.93) |
2.82 (0.91–8.74) |
1.40 (1.04–1.88) |
0.025 |
rs451360 |
C/A |
0.14 |
1.13 (0.76–1.69) |
1.52 (0.69–3.33) |
1.22 (0.89–1.68) |
0.222 |
aMajor/minor alleles as determined by allele frequency among genotyped controls
bMinor allele frequency among genotyped controls
AA: major allele homozygotes (reference group); AB: heterozygotes; BB: minor allele homozygotes; A: major allele; B: minor allele
P value in bold means statistically significant.
Table 3: Association between tagSNPs on TERT and CLPTM1L region and GIST risk (Stage II)
SNP |
Allelesa |
Stage |
AB OR (95% CI) |
BB OR (95% CI) |
B vs A OR (95% CI) |
P |
rs7726159 |
C/A |
I |
1.32 (0.93–1.87) |
1.67 (1.04–2.68) |
1.33 (1.05–1.69) |
0.017 |
II |
0.94 (0.75–1.19) |
0.95 (0.71–1.29) |
0.96 (0.82–1.13) |
0.630 |
||
rs10069690 |
C/T |
I |
1.36 (0.96–1.93) |
2.82 (0.91–8.74) |
1.40 (1.04–1.88) |
0.025 |
II |
0.92 (0.74–1.15) |
0.83 (0.27–2.49) |
0.93 (0.77–1.13) |
0.489 |
||
rs2736098 |
C/T |
I |
1.49 (1.03–2.14) |
1.78 (1.17–2.72) |
1.44 (1.14–1.81) |
1.92 × 10−3 |
II |
1.27 (1.03–1.60) |
1.38 (1.04–1.83) |
1.23 (1.06–1.44) |
6.63 × 10−3 |
||
Combined |
1.33 (1.09–1.101.62) |
1.49 (1.18–1.88) |
1.29 (1.14–1.47) |
7.03 × 10−5 |
aMajor/minor alleles as determined by allele frequency among genotyped controls
AA: major allele homozygotes (reference group), AB: heterozygotes, BB: minor allele homozygotes;
P value in bold means statistically significant.
The robustness of these findings was evaluated by sensitivity analyses. First, additional adjustments by education, BMI, WHR, physical activity, drinking and smoking were conducted respectively. The results didn’t change materially. To validate our findings of TERT rs2736098 and explore its influence on RTL, we used real-time PCR to measure the RTL in a random sample of 150 GIST cases. In GIST tissues, We found a significant difference in RTLs among the genotype CC, CT, and TT, respectively (Figure 1, P < 0.001).
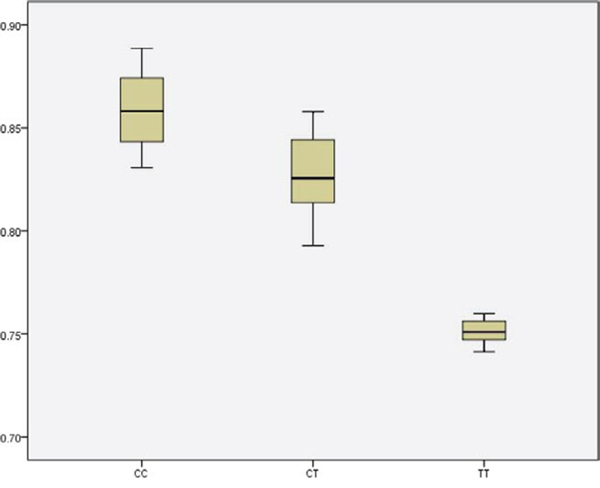
Figure 1: Boxplot for the RTL with different genotype of SNP rs2736098.
DISCUSSION
To the best of our knowledge, this is the first report to attempt an evaluation of the association of six tagSNPs on TERT and CLPTM1L region potentially related to GIST carcinogenesis. In this large population-based, two-stage, case-control study, we identified that the variant rs2736098 was significantly associated with increased risk of GIST, especially for GISTs located in stomach. To validate this finding, real-time PCR showed that the RTL in GIST cells were significantly lower than that of their adjacent normal tissues. And in GIST tissues, carriers of genotype CC have the longest RTL, following by carriers of genotype CT, while carriers of genotype TT have the shortest RTL. These provide evidence to implicate rs2736098 polymorphism as a novel susceptibility factor for GIST risk.
GISTs are the most common soft tissue sarcoma of the gastrointestinal tract, resulting most commonly from KIT or platelet-derived growth factor receptor alpha (PDGFRalpha)-activating mutations [19–21]. However, they have distinct genetic background and gene expression patterns according to localization, genotype and aggressiveness [22, 23]. Chr5p15.33 harbors a unique cancer susceptibility region that contains at least two plausible candidate genes: TERT and CLTPM1L [24–27]. The TERT gene has been mapped to chromosome 5p15.33 and consisted of 16 exons and 15 introns spanning 35kb of genomic DNA [28]. It encodes the catalytic subunit of the telomerase reverse transcriptase, which, in combination with an RNA template (TERC), adds nucleotide repeats to chromosome ends [29, 30]. The CLTPM1L gene, also known as cisplatin resistance-related protein 9 (CRR9p), encodes a protein that is overexpressed in lung and pancreatic cancer, promotes growth and survival, and is required for KRAS driven lung cancer [31, 32]. It confer resistance to apoptosis caused by genotoxic agents in association with up-regulation of the anti-apoptotic protein, Bcl-xL [33]. Studies indicate that the TERT-CLPTM1L region may harbor multiple elements that have the capacity to influence molecular phenotypes in cancer development [16, 34]. Thus, It is possible to study that the interplay between risk variants, multiple biological mechanisms and attributed genes, influence various cancers, including GISTs.
Although few literature investigating the associations somatic mutations of TERT and GIST risk [15, 35, 36], none has evaluated germline genetic associations with GISTs. In current study, we identified rs2736098 contribute to increased risk of GIST and shorter RTL, using a two-stage, case-control study. This finding is consistent with many previous epidemiological studies with different cancer types, including lung cancer, bladder cancer, pancreatic cancer, gastrointestinal cancers, breast cancer, ovarian cancer, and so on [16, 37–39]. All of evidence above implicate TERT rs2736098 polymorphism as a novel susceptibility factor for carcinogenesis.
Considering rs2736098 polymorphism being a tagSNP, it is possible the association seen with rs2736098 tagSNP is due to one of those linked polymorphisms. We additionally listed the detailed information for these 11 linked polymorphisms of rs2736098 in Table 4. Among them, 9 are intergenic SNPs, and 2 are intron variants. Although rs2736098 was a synonymous SNP, our results showed that carriers of genotype CC have the longest RTL, following by carriers of genotype CT, while carriers of genotype TT have the shortest RTL in GIST tissues (P < 0.001). This evidence yet indicate the functionality of SNP rs2736098. Further fine mapping and sequencing studies may be helpful for the validation the our conclusions.
Table 4: Detailed information for linked SNPs of rs2736098
SNP |
Gene |
Allelesa |
Positions |
Region/Functionality |
rs2736098 |
TERT |
C/T |
chr5:1293836 |
Exon/synonymous |
rs2736109 |
TERT |
A/G |
chr5:1296759 |
intergenic |
rs2736108 |
TERT |
G/A |
chr5: 1297488 |
intergenic |
rs2853672 |
TERT |
C/A |
chr5: 1293233 |
intron |
rs2735940 |
TERT |
G/A |
chr5: 1296736 |
intergenic |
rs2736103 |
TERT |
A/G |
chr5: 1300401 |
intergenic |
rs2735846 |
TERT |
C/G |
chr5: 1299379 |
intergenic |
rs13174919 |
TERT |
C/G |
chr5: 1300112 |
intergenic |
rs4975612 |
TERT |
G/T |
chr5: 1300310 |
intergenic |
rs13174814 |
TERT |
C/G |
chr5: 1300109 |
intergenic |
rs2736099 |
TERT |
C/T |
chr5: 1287340 |
intron |
rs2736105 |
TERT |
A/G |
chr5: 1299756 |
intergenic |
aMajor/minor alleles as determined by allele frequency among genotyped controls
Strengths of the current study include a large population, a two-stage genotyping design to minimize type I error, and good coverage of the genetic variation in the TERT and CLPTM1L region. This study also had several limitations. First, selection bias might have occurred through the selection of control subjects when the sampling is not random within the subpopulations of cancer and cancer-free subjects, though we have try our best to control it through the whole process of the study; since this study was restricted to a Chinese Han population, it is uncertain whether our findings can be replicated by other ethnic groups. Second, in spite of the relatively large sample size, the power to elucidate gene–environment interactions was limited because of the small magnitude of the overall association.
In summary, our findings regarding genetic variation in the TERT and CLPTM1L region and GIST risk add to the growing body of literature suggesting the importance of this genetic region to cancer development. Further research is needed in this area to understand how changes in telomere length over time may influence GIST carcinogenesis in a prospective setting and interaction with a p53 pathway of development.
MATERIALS AND METHODS
Subjects
The methods were carried out in “accordance” with the approved guidelines. Also, all experimental protocols were approved by the institutional review boards of liaoning cancer hospital and shengjing hospital, and written informed consent was obtained from all participants. Cases were histopathologically confirmed GIST patients. Controls without clinic evidence of gastrointestinal diseases or tumors were randomly selected from a pool of healthy volunteers who visited the general health checkup center of the same hospital for routine scheduled physical exams. Controls were individually matched to cases on sex, ethnicity (Han), age (±5 years). After giving written consent, participants provided demographic information using a standard interviewer-administered questionnaire. Stage I includes 300 GIST cases and 300 controls, while stage II includes 700 GIST cases and 700 controls. Totally incluede in this study were 1,000 cases and 1,000 controls. Five ml of peripheral blood was obtained for DNA extraction.
SNP selection and genotyping
Totally, five SNPs in the TERT gene (rs7726159, rs2853677, rs2736098, rs13172201, rs10069690) and one SNP in neighboring CLPTM1L gene (rs451360) were selected in this study (details in Table 5 and supplementary table 1), according to the previous literature [16]. 5′-Nuclease TaqMan® assays were used to genotype the polymorphisms in 96-well plates on an ABI PRISM 7900HT Sequence Detection system (Applied BioSystems, Foster City, CA, USA). The primers and probes for the TaqMan® assays were designed using Primer Express Oligo Design software v2.0 (ABI PRISM) and are available upon request as TaqMan® Pre-Designed SNP Genotyping Assays. Samples from matched case-control pairs were handled identically and genotyped in the same batch in a blinded fashion. All included SNPs had concordance rates of 100% among duplicates within each platform, and laboratory personnel were blinded to the case–control and QC status of all samples.
Table 5: Basic information for each tagSNP
SNP |
Gene |
Allelesa |
Positions |
Region/Functionality |
rs7726159 |
TERT |
C/A |
chr5:1282069 |
intron |
rs2853677 |
TERT |
A/G |
chr5:1286944 |
intron |
rs2736098 |
TERT |
C/T |
chr5:1293836 |
Exon/synonymous |
rs13172201 |
TERT |
C/T |
chr5:1269156 |
intron |
rs10069690 |
TERT |
C/T |
chr5:1279540 |
intron |
rs451360 |
CLPTM1L |
C/A |
chr5:1319680 |
intron |
aMajor/minor alleles as determined by allele frequency among genotyped controls
Relative telomere length (RTL) determination
RTL of GIST cells were measured using quantitative real-time polymerase chain reaction (PCR) as described earlier in a random sample of 150 GIST cases [17]. In short, telomeres and a single-copy gene (β2-globin) were amplified including an internal reference control cell line (CCRF-CEM) to which all samples were compared. The ΔΔCt method was used for calculation of RTL values and a standard curve was created in each PCR run to monitor the PCR efficiency.
Statistical Analyses
Hardy–Weinberg equilibrium (HWE) was tested by comparing observed and expected genotype frequencies among controls (x2 test). Odds ratios (ORs) and corresponding 95% confidence intervals (CIs) were determined by logistic regression analyses using models that included adjustment for age and gender. Linkage disequilibrium (LD) was assessed by Haploview [18]. Differences of RTLs among different groups were compared by One-Way ANOVA method. All statistical analyses were conducted with SAS version 9.2 (SAS Institute Inc.). All statistical tests were 2-tailed, and P < 0.05 was interpreted as statistically significant unless otherwise indicated.
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest.
Author contributions
S.C. and Z.R. designed the experiments; S.C., Z.R., Z.J., X.J., L.F., X.Y., B.X. and D.C. performed the investigations and experiments; S.C., Z.R., Z.J., X.J., analyzed the data; S.C., Z.R., Z.J., X.J., wrote the manuscript; all authors reviewed the manuscript.
REFERENCES
1. Hirota S, Isozaki K, Moriyama Y, Hashimoto K, Nishida T, Ishiguro S, Kawano K, Hanada M, Kurata A, Takeda M, Muhammad Tunio G, Matsuzawa Y, Kanakura Y, Shinomura Y, Kitamura Y. Gain-of-function mutations of c-kit in human gastrointestinal stromal tumors. Science. 1998; 279:577–580.
2. Barnett CM, Corless CL, Heinrich MC. Gastrointestinal stromal tumors: molecular markers and genetic subtypes. Hematol Oncol Clin North Am. 2013; 27:871–888.
3. Miettinen M, Lasota J. Gastrointestinal stromal tumors: review on morphology, molecular pathology, prognosis, and differential diagnosis. Archives of pathology & laboratory medicine. 2006; 130:1466–1478.
4. Janeway KA, Liegl B, Harlow A, Le C, Perez-Atayde A, Kozakewich H, Corless CL, Heinrich MC, Fletcher JA. Pediatric KIT wild-type and platelet-derived growth factor receptor alpha-wild-type gastrointestinal stromal tumors share KIT activation but not mechanisms of genetic progression with adult gastrointestinal stromal tumors. Cancer Res. 2007; 67:9084–9088.
5. Daum O, Grossmann P, Vanecek T, Sima R, Mukensnabl P, Michal M. Diagnostic morphological features of PDGFRA-mutated gastrointestinal stromal tumors: molecular genetic and histologic analysis of 60 cases of gastric gastrointestinal stromal tumors. Ann Diagn Pathol. 2007; 11:27–33.
6. Andre C, Martin E, Cornu F, Hu WX, Wang XP, Galibert F. Genomic organization of the human c-kit gene: evolution of the receptor tyrosine kinase subclass III. Oncogene. 1992; 7:685–691.
7. Cho S, Kitadai Y, Yoshida S, Tanaka S, Yoshihara M, Yoshida K, Chayama K. Genetic and pathologic characteristics of gastrointestinal stromal tumors in extragastric lesions. Int J Mol Med. 2006; 18:1067–1071.
8. Heinrich MC, Rubin BP, Longley BJ, Fletcher JA. Biology and genetic aspects of gastrointestinal stromal tumors: KIT activation and cytogenetic alterations. Hum Pathol. 2002; 33:484–495.
9. Miettinen M, Sarlomo-Rikala M, Sobin LH, Lasota J. Gastrointestinal stromal tumors and leiomyosarcomas in the colon: a clinicopathologic, immunohistochemical, and molecular genetic study of 44 cases. Am J Surg Pathol. 2000; 24:1339–1352.
10. O’Brien KM, Orlow I, Antonescu CR, Ballman K, McCall L, Dematteo R, Engel LS. Gastrointestinal stromal tumors: a case-only analysis of single nucleotide polymorphisms and somatic mutations. Clinical sarcoma research. 2013; 3:12.
11. Schilling G, Penas EM, Janjetovic S, Oliveira-Ferrer L, Braig M, Behrmann P, Bokemeyer C, Dierlamm J. Molecular characterization of chromosomal band 5p15.33: a recurrent breakpoint region in mantle cell lymphoma involving the TERT-CLPTM1L locus. Leuk Res. 2013; 37:280–286.
12. Zhao Y, Chen G, Song X, Chen H, Mao Y, Lu D. Fine-mapping of a region of chromosome 5p15.33 (TERT-CLPTM1L) suggests a novel locus in TERT and a CLPTM1L haplotype are associated with glioma susceptibility in a Chinese population. Int J Cancer. 2012; 131:1569–1576.
13. Haiman CA, Chen GK, Vachon CM, Canzian F, Dunning A, Millikan RC, Wang X, Ademuyiwa F, Ahmed S, Ambrosone CB, Baglietto L, Balleine R, Bandera EV, Beckmann MW, Berg CD, Bernstein L, et al. A common variant at the TERT-CLPTM1L locus is associated with estrogen receptor-negative breast cancer. Nat Genet. 2011; 43:1210–1214.
14. Rafnar T, Sulem P, Stacey SN, Geller F, Gudmundsson J, Sigurdsson A, Jakobsdottir M, Helgadottir H, Thorlacius S, Aben KK, Blondal T, Thorgeirsson TE, Thorleifsson G, Kristjansson K, Thorisdottir K, Ragnarsson R, et al. Sequence variants at the TERT-CLPTM1L locus associate with many cancer types. Nat Genet. 2009; 41:221–227.
15. Papathomas TG, Oudijk L, Zwarthoff EC, Post E, Duijkers FA, van Noesel MM, Hofland LJ, Pollard PJ, Maher ER, Restuccia DF, Feelders RA, Franssen GJ, Timmers HJ, Sleijfer S, de Herder WW, de Krijger RR, et al. Telomerase reverse transcriptase promoter mutations in tumors originating from the adrenal gland and extra-adrenal paraganglia. Endocr Relat Cancer. 2014; 21:653–661.
16. Wang Z, Zhu B, Zhang M, Parikh H, Jia J, Chung CC, Sampson JN, Hoskins JW, Hutchinson A, Burdette L, Ibrahim A, Hautman C, Raj PS, et al. Imputation and subset-based association analysis across different cancer types identifies multiple independent risk loci in the TERT-CLPTM1L region on chromosome 5p15.33. Hum Mol Genet. 2014; 23:6616–6633.
17. Nordfjall K, Larefalk A, Lindgren P, Holmberg D, Roos G. Telomere length and heredity: Indications of paternal inheritance. Proc Natl Acad Sci U S A. 2005; 102:16374–16378.
18. Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 2005; 21:263–265.
19. Haller F, Zhang JD, Moskalev EA, Braun A, Otto C, Geddert H, Riazalhosseini Y, Ward A, Balwierz A, Schaefer IM, Cameron S, Ghadimi BM, Agaimy A, et al. Combined DNA methylation and gene expression profiling in gastrointestinal stromal tumors reveals hypomethylation of SPP1 as an independent prognostic factor. Int J Cancer. 2015; 136:1013–1023.
20. Maynard MA, Huang SA. Thyroid hormone inactivation in gastrointestinal stromal tumors. N Engl J Med. 2014; 371:86–87.
21. von Mehren M, Randall RL, Benjamin RS, Boles S, Bui MM, Casper ES, Conrad EU 3rd, DeLaney TF, Ganjoo KN, George S, Gonzalez RJ, Heslin MJ, Kane JM 3rd, Mayerson J, McGarry SV, Meyer C, et al. Gastrointestinal stromal tumors, version 2. Journal of the National Comprehensive Cancer Network : JNCCN. 2014; 12:853–862.
22. Corless CL. Gastrointestinal stromal tumors: what do we know now. Mod Pathol. 2014; 27:S1–16.
23. Schaefer IM, Delfs C, Cameron S, Gunawan B, Agaimy A, Ghadimi BM, Haller F. Chromosomal aberrations in primary PDGFRA-mutated gastrointestinal stromal tumors. Hum Pathol. 2014; 45:85–97.
24. Yin Z, Cui Z, Ren Y, Zhang H, Yan Y, Zhao Y, Ma R, Wang Q, He Q, Zhou B. Genetic polymorphisms of TERT and CLPTM1L, cooking oil fume exposure, and risk of lung cancer: a case-control study in a Chinese non-smoking female population. Med Oncol. 2014; 31:114.
25. Walsh KM, Codd V, Smirnov IV, Rice T, Decker PA, Hansen HM, Kollmeyer T, Kosel ML, Molinaro AM, McCoy LS, Bracci PM, Cabriga BS, Pekmezci M, Zheng S, Wiemels JL, Pico AR, et al. Variants near TERT and TERC influencing telomere length are associated with high-grade glioma risk. Nat Genet. 2014; 46:731–735.
26. Zhang Y, Zhao M, Shen L, Ren Y, Su L, Li X, Yin Z, Zhou B. Genetic polymorphisms of TERT and CLPTM1L and risk of lung cancer: a case-control study in northeast Chinese male population. Med Oncol. 2014; 31:18.
27. Zhao MM, Zhang Y, Shen L, Ren YW, Li XL, Yin ZH, Zhou BS. Genetic variations in TERT-CLPTM1L genes and risk of lung cancer in a Chinese population. Asian Pac J Cancer Prev. 2014; 15:2809–2813.
28. Wick M, Zubov D, Hagen G. Genomic organization and promoter characterization of the gene encoding the human telomerase reverse transcriptase (hTERT). Gene. 1999; 232:97–106.
29. Weinrich SL, Pruzan R, Ma L, Ouellette M, Tesmer VM, Holt SE, Bodnar AG, Lichtsteiner S, Kim NW, Trager JB, Taylor RD, Carlos R, Andrews WH, Wright WE, Shay JW, Harley CB, et al. Reconstitution of human telomerase with the template RNA component hTR and the catalytic protein subunit hTRT. Nat Genet. 1997; 17:498–502.
30. Cong YS, Wen J, Bacchetti S. The human telomerase catalytic subunit hTERT: organization of the gene and characterization of the promoter. Hum Mol Genet. 1999; 8:137–142.
31. James MA, Vikis HG, Tate E, Rymaszewski AL, You M. CRR9/CLPTM1L regulates cell survival signaling and is required for Ras transformation and lung tumorigenesis. Cancer Res. 2014; 74:1116–1127.
32. Jia J, Bosley AD, Thompson A, Hoskins JW, Cheuk A, Collins I, Parikh H, Xiao Z, Ylaya K, Dzyadyk M, Cozen W, Hernandez BY, Lynch CF, Loncarek J, Altekruse SF, Zhang L, et al. CLPTM1L promotes growth and enhances aneuploidy in pancreatic cancer cells. Cancer Res. 2014; 74:2785–2795.
33. James MA, Wen W, Wang Y, Byers LA, Heymach JV, Coombes KR, Girard L, Minna J, You M. Functional characterization of CLPTM1L as a lung cancer risk candidate gene in the 5p15.33 locus. PLoS One. 2012; 7:e36116.
34. Heidenreich B, Rachakonda PS, Hosen I, Volz F, Hemminki K, Weyerbrock A, Kumar R. TERT promoter mutations and telomere length in adult malignant gliomas and recurrences. Oncotarget. 2015; 6:10617–10633.
35. Wang K, Liu T, Ge N, Liu L, Yuan X, Liu J, Kong F, Wang C, Ren H, Yan K, Hu S, Xu Z, Bjorkholm M, Fan Y, Zhao S, Liu C, et al. TERT promoter mutations are associated with distant metastases in upper tract urothelial carcinomas and serve as urinary biomarkers detected by a sensitive castPCR. Oncotarget. 2014; 5:12428–12439.
36. Killela PJ, Pirozzi CJ, Healy P, Reitman ZJ, Lipp E, Rasheed BA, Yang R, Diplas BH, Wang Z, Greer PK, Zhu H, Wang CY, Carpenter AB, Friedman H, Friedman AH, Keir ST, et al. Mutations in IDH1, IDH2, and in the TERT promoter define clinically distinct subgroups of adult malignant gliomas. Oncotarget. 2014; 5:1515–1525.
37. Ngeow J, Eng C. TERT and BRAF in Thyroid Cancer: Teaming Up for Trouble. J Clin Oncol. 2014; 32:2683–2684.
38. Zhong R, Liu L, Zou L, Zhu Y, Chen W, Zhu B, Shen N, Rui R, Long L, Ke J, Lu X, Zhang T, Zhang Y, Wang Z, Sun Y, Cheng L, et al. Genetic variations in TERT-CLPTML locus are associated with risk of lung cancer in Chinese population. Mol Carcinog. 2013; 52:E118–126.
39. Bojesen SE, Pooley KA, Johnatty SE, Beesley J, Michailidou K, Tyrer JP, Edwards SL, Pickett HA, Shen HC, Smart CE, Hillman KM, Mai PL, Lawrenson K, Stutz MD, Lu Y, Karevan R, et al. Multiple independent variants at the TERT locus are associated with telomere length and risks of breast and ovarian cancer. Nat Genet. 2013; 45:371–384.