
INTRODUCTION
Chemokines are small heparin-binding proteins weighing 8–14 kDa, which guide the migration of circulating lymphocytes to specific tissues [1]. Relying on specific receptors, chemokines induce the movement of lymphocytes in a concentration-dependent manner [2]. In cancer patients, the immune cell-infiltration is tightly associated with chemotaxis, and the numbers and types of lymphocytes recruited are determined by the local chemokines secreted [3]. As reported, CCL5 and CXCL10 attract CD8+ T lymphocytes into various tumors [4–7]. In addition, Muthuswamy et al. [8] and Berghuis et al. [9] reported that CCL5 and CXCL10 favor the recruitment of CD8+ T lymphocytes into malignant tissues. However, the expression of CCL5 and CXCL10 and its impact on the migration of CD8+ T lymphocytes remain unknown in patients with esophageal squamous cell carcinoma (ESCC).
As the major histologic subtype of esophageal cancer, ESCC, is one of the most common malignancies with high incidence in China [10, 11]. Although substantial improvements have been made in the diagnosis and treatment of ESCC, the 5-year survival after surgery is about 25%. Considering about 66% of patients are inoperable at diagnosis, the 5-year survival of all ESCC patients is just 10% [12]. Hence, there is a need for more promising therapeutic regimens. T cell or Cytokine-induced killer cell (CIK) therapy could be a great help for ESCC treatment. A prerequisite for effective immune cell therapy is efficient delivery of tumor-killing cells into the tumor lesions. Depending on the specific interaction of chemokines with cognitive receptors, T lymphocytes move across the vascular endothelium and enter tumor tissues, where they are retained [13]. Enhancing the specific interaction would improve T lymphocyte infiltration of tumors [14]. Similarly, the tumor-targeted movement of CIK cells depends on the chemokine-chemokine receptor interaction [15]. Therefore, investigating the chemokines and cognitive receptors harboring T lymphocytes within tumors could be beneficial for the improvement of immune cell-based ESCC therapy.
In this project, we focused on the chemotactic movement of CD8+ T lymphocytes into ESCC, explored the expression of CCL5 and CXCL10, and investigated the corresponding chemokine receptors (CCR5 and CXCR3) on CD8+ T lymphocytes. Our findings show that CCL5-CCR5 and CXCL10-CXCR3 axes are critical for tumor-directed movement of CD8+ T lymphocytes in ESCC.
RESULTS
Expression of CCL5 and CXCL10 is upregulated in tumor tissues
To investigate the expression of CCL5 and CXCL10, we performed immunohistochemistry assay (IHC) on tissue sections from ESCC patients. As shown in Figure 1A, CCL5 and CXCL10 were strongly expressed in the cytoplasm of malignant cells, while moderately stained in marginal tissues. Statistical analysis of those samples indicated that both CCL5 and CXCL10 expression (IHC scores) was significantly higher in tumor lesions versus marginal tissues (1.878 ± 1.679 vs. 0.588 ± 0.543, p < 0.0001; 2.472 ± 1.997 vs. 1.588 ± 1.042, p < 0.005; Figure 1B).
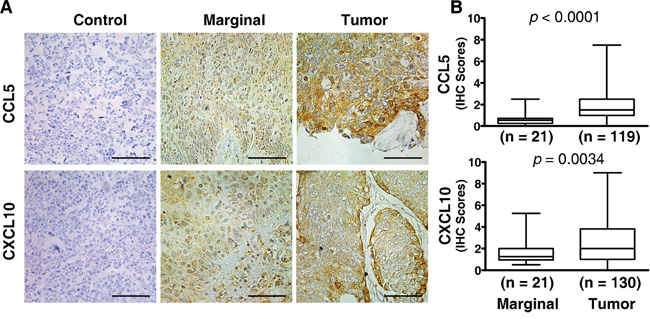
Figure 1: Elevated expression of CCL5 and CXCL10 in ESCC tissue. Both marginal and tumor tissues from ESCC patients were examined by immunohistochemistry for the expression of chemokines of interest, and the immunohistochemical scores were calculated as described in the Materials and Methods. A. Representative photographs of marginal and tumor tissue sections stained with specific primary antibodies or non-immune rabbit IgG are shown. B. Expression scores of CCL5 and CXCL10 were significantly higher in malignant tissues than in marginal tissues. Tumor tissue sections stained with non-immune rabbit IgG were used as a negative control. Scale bar, 50 μM.
Association of CCL5 and CXCL10 with clinical factors and overall survival
Next, we checked the associations of CCL5 and CXCL10 with various clinical and pathologic characteristics in individual ESCC patients. Those parameters included gender (male, female), age (44–84 y), tumor-node-metastasis stage (TNM, stage I–IV), tumor invasion (T1–T4), differentiation (well, poor), lymph node metastasis (negative, positive), and tumor length (0.5–9 cm). We found the expression levels of CCL5 were significantly elevated in T3–T4 stage patients, compared with their T1–T2 stage counterparts (2.196 ± 2.009 vs. 1.555 ± 1.193, p < 0.05) (Table 1). However, the expression of CCL5 was not related to the other clinical factors including gender, age, clinical TNM stage, tumor differentiation, lymph node metastasis, or tumor length (Table 1). Unexpectedly, CXCL10 was also not associated with those parameters (Table 1). In addition, we analyzed the effect of CCL5 and CXCL10 on patients’ survival. Patients with high CCL5-expression had greater survival than those with low CCL5-expression (p < 0.05) (Figure 2A). As shown in Figure 2B, subjects with high CXCL10-expression exhibited increased survival, although the difference was not statistically significant (p > 0.05).
Table 1: Clinical and pathological profile of ESCC patients
N |
% |
|
Total |
207 |
100 |
Gender |
||
Male |
144 |
69.57 |
Female |
63 |
30.43 |
Age |
||
> 60 |
126 |
60.87 |
≤ 60 |
81 |
39.13 |
Clinical stage |
||
I–IIa |
143 |
69.08 |
IIb–IV |
64 |
30.92 |
Differentiation |
||
Poor (3–4) |
88 |
42.51 |
Well (1–2) |
119 |
57.49 |
Lymph node metastasis |
||
Negative |
150 |
72.46 |
Positive |
57 |
27.54 |
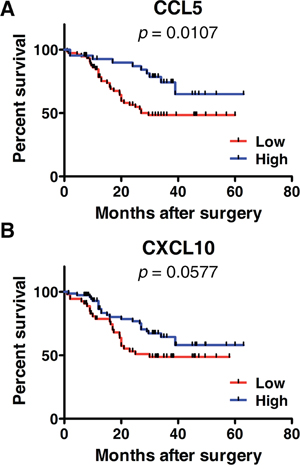
Figure 2: Kaplan-Meier survival curves according to the expressions of CCL5 A. or CXCL10 B. A: A log-rank test was performed to compare the overall survival of patients with high (n = 76) or low CCL5-expression (n = 43). B: A log-rank test was performed to compare the overall survival of patients with high (n = 54) or low CXCL10-expression (n = 76).
Correlation of CCL5 and CXCL10 expression with CD8+ T lymphocyte markers
To check whether CCL5 and CXCL10 exert an impact on the tumoral accumulation of CD8+ T lymphocytes, we performed an RT-PCR assay. In tumor lesions, the local expression of CCL5 and CXCL10 was positively associated with the expression of the CD8+ T lymphocyte markers CD8 and Granzyme B (RCD8 = 0.2338, pCD8 < 0.01, RGranB = 0.5517, pGranB < 0.001; RCD8 = 0.2972, pCD8 < 0.005, RGranB = 0.3204, pGranB < 0.001) (Figure 3). Meanwhile, neither of those two chemokines was correlated with the markers for CD4+ T lymphocytes (CD4) or natural killer (NK) cells (CD56) (RCD4 = 0.1195, pCD4 > 0.05, RCD56 = 0.1496, pCD56 > 0.05; RCD4 = 0.1533, pCD4 > 0.05, RCD56 = 0.1768, pCD56 > 0.05) (Supplementary Figure S1). These data demonstrate that CCL5 and CXCL10 predict the recruitment and retention of CD8+ T lymphocytes in ESCC.
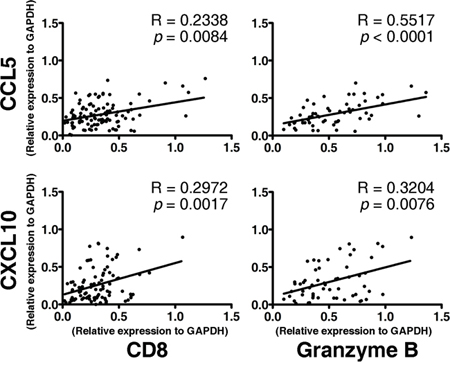
Figure 3: Association between the expression of CD8+ effector T lymphocyte markers (CD8 and Granzyme B) and the intensities of CCL5 (upper panel) and CXCL10 (lower panel) in tumor lesions. A total of 104 fresh tumor tissues from ESCC patients were processed, RNA was extracted, and an RT-PCR assay of interested genes was performed. The Spearman test was performed to determine the correlation between genes.
Enrichment of CCR5+CD8+ and CXCR3+CD8+ T lymphocytes in Tumor-infiltrating Lymphocytes (TILs)
A specific receptor is a key component in chemokine-driven migration of T lymphocytes [4, 5]. Therefore, we detected the expression of CCR5 (CCL5-specific receptor) and CXCR3 (CXCL10-specific receptor) on CD8+ T lymphocytes in TILs and peripheral blood lymphocytes (PBLs). As shown in Figure 4A, CD8+ T lymphocytes (CD3+CD8+) were gated, and the expression of CCR5 and CXCR3 were determined. In 61 paired samples, CCR5 was expressed on a greater fraction of CD8+ T lymphocytes in TILs than in PBLs (47.57 ± 25.91% vs. 21.73 ± 16.25%, p < 0.0001) (Figure 4B). Similarly, the frequency of CXCR3 was significantly higher on tumor-infiltrating CD8+ T lymphocytes than their circulating counterparts (44.52 ± 23.44% vs. 11.62 ± 9.86%, p < 0.0001) (Figure 4A and 4B).
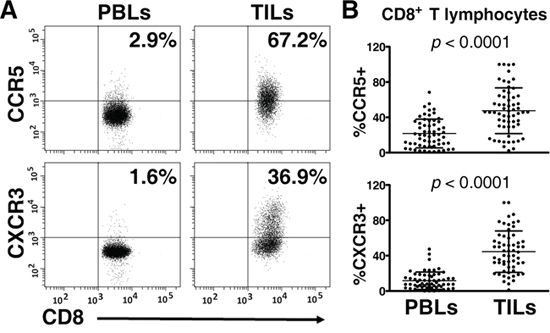
Figure 4: Expression of CCR5 and CXCR3 on CD8+ T lymphocytes in peripheral blood lymphocytes (PBLs) and matched tumor-infiltrating lymphocytes (TILs) (n = 61). Paired PBLs and TILs were stained with antibodies specific for CD3, CD8, CCR5, and CXCR3. CD8+ T lymphocytes (CD3+CD8+) were gated; then, the percentages of CCR5+CD8+ and CXCR3+CD8+ T lymphocytes were determined with multicolor flow cytometry. A. Representative plots of CCR5 and CXCR3 staining on CD8+ T lymphocytes in matched PBLs and TILs, respectively, are shown. B. Paired analysis, for which the paired t test was used, showed that CCR5 and CXCR3 were expressed on greater portions of CD8+ T lymphocytes in TILs, compared with their counterparts in PBLs.
CCL5 and CXCL10 are important for the chemotactic migration of CD8+ T lymphocytes
To ascertain the regulatory effects of CCL5 and CXCL10 on CD8+ T lymphocyte migration, transwell assays were performed. Fresh ESCC tissues were cultured in DMEM media supplied with 10% FBS, and the supernatants were collected 48 h later. In addition, CD8+ T lymphocytes were magnetically isolated from fresh TILs. The purity of T lymphocytes used was greater than 90% (Data not shown). Compared with DMEM media supplemented with 10% FBS, the supernatants derived from primary tumor tissues robustly enhanced the movement of T lymphocytes (p < 0.001) (Figure 5). However, the neutralizing antibodies for CCL5 or CXCL10 significantly hampered the migration induced by tumor supernatant (pCCL5 < 0.05; pCXCL10 < 0.01) (Figure 5), and the migration of T lymphocytes was further inhibited by the combined use of anti-CCL5 and anti-CXCL10 neutralizing antibodies (p < 0.001) (Figure 5). Similar results of inhibited movement of CD8+ T lymphocytes by CCL5- and/or CXCL10-specific antibodies were obtained in five other independent experiments (Supplementary Figure S2). These results indicated that both CCL5 and CXCL10 are important in attracting CD8+ T lymphocytes towards ESCC.
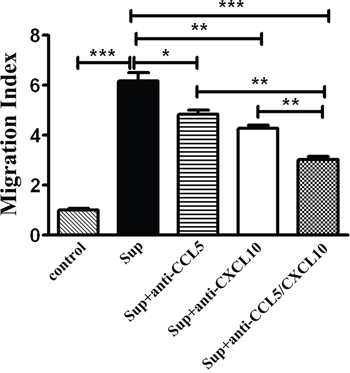
Figure 5: CCL5 and CXCL10 attracting CD8+ T lymphocytes in vitro. Supernatants of primary tumor tissues were added, alone or with neutralizing antibodies specific for CCL5 (1 μg/mL) and/or CXCL10 (5 μg/mL), as indicated. DMEM media with 10% FBS were used as a control. Then, purified CD8+ T lymphocytes (Purity > 90%) from the TILs of an ESCC patient were placed into transwell inserts (5-μm pore size). After incubation, cells that migrated into the lower chamber were collected and counted. The migration index was calculated by dividing the number of cells that migrated in indicated groups by the number migrating in control groups. Data shown represent mean ± SD of data from 1 representative experiment of 6 independent experiments. *p < 0.05; **p < 0.01; ***p < 0.001.
DISCUSSION
The natural infiltration of T lymphocytes is common in tumors [16–19]; the chemotaxis-induced accumulation of T lymphocytes has great effects on tumor differentiation, metastasis, disease progress, and patient survival [20–24]. Although several studies have examined the expression of chemokine receptors by esophageal carcinoma cells [25, 26], the expressions of ESCC-associated chemokines in relation to local T lymphocytes accumulation had not been investigated. In this study, we sought to determine CCL5 and CXCL10 expression and their effects on CD8+ T lymphocyte trafficking to ESCC. Several studies indicated that the tumoral expression of CCL5 is tightly related with CD8+ T lymphocytes infiltrating in colorectal carcinoma [8, 27], melanoma [28], and sarcoma [9]. CXCL10 is involved in the recruitment of CD8+ T lymphocytes in breast cancer [29], colorectal carcinoma [8], melanoma [28], and hepatocellular carcinoma [30]. Consistent with those reports, we found the expression of CCL5 and CXCL10 in ESCC tissues was a) strongly elevated, compared to surrounding healthy tissues, and b) positively correlated with the local expression of CD8+ T lymphocyte markers, including CD8 and Granzyme B. These observations strongly indicate that CCL5 and CXCL10 are involved in the local recruitment of CD8+ T lymphocytes to ESCC lesions and their local retention.
To further verify the effects of CCL5 and CXCL10 on tumoral infiltration of CD8+ T lymphocytes, we checked the expression of their specific receptors on T lymphocytes. Upon activating specific receptors, chemokines recruit the cells expressing such receptors into tumors [31–33]. Therefore, studying CCR5 (specifically recognizing CCL5) and CXCR3 (specifically recognizing CXCL10) helps to elucidate the molecular basis of CD8+ T lymphocyte homing to specific sites. The CCR5-CCL5 axis has been reported to induce the tumoral accumulation of CD8+ T lymphocytes in renal cell carcinoma [4], nasopharyngeal carcinoma [5], and colorectal carcinoma [27]. The CXCR3-CXCL10 interaction induces the infiltration of CD8+ T lymphocytes in renal cell carcinoma [4] and colorectal carcinoma [27]. In this project, we performed a flow cytometry assay to detect the frequencies of CCR5 and CXCR3 on CD8+ T lymphocytes in TILs and PBLs, separately. The percentages of CCR5+CD8+ and CXCR3+CD8+ T lymphocytes were 47.57 ± 25.91% and 44.52 ± 23.44% in TILs, respectively, whereas the proportions of those subgroups in T lymphocytes were just 21.73 ± 16.25% and 11.62 ± 9.86% in PBLs. The differences were statistically significant (pCCR5 < 0.0001;pCXCR3 < 0.0001). This phenomenon suggests that CCR5 and CXCR3 are activated by their ligands CCL5 and CXCL10 and then deliver CD8+ T lymphocytes into malignant tissues, where CCL5 and CXCL10 are produced and the concentrations of chemokines are higher. To consolidate the roles of CCL5 and CXCL10 in attracting T lymphocytes, transwell assays were performed. As expected, the migration of CD8+ T lymphocytes induced by tumor supernatant was partially but significantly inhibited by CCL5- and/or CXCL10-neutralizing antibodies. Taking the data together, it can be inferred that the CCL5-CCR5 and CXCL10-CXCR3 axes are critical for the movement of CD8+ T lymphocytes to tumor tissues in ESCC patients.
Tumoral infiltration of cytolytic CD8+ T lymphocytes is associated with a favorable prognosis and increased survival [34–36]. In our study, CCL5 and CXCL10 were positively correlated with ESCC patient survival (Figure 2). Patients with high CCL5- or CXCL10-expression exhibited better overall survival. The impact of chemokines on survival is likely associated with the accumulation of CD8+ T lymphocytes. Consistent with our observations, CCL5 and CXCL10 are reported to recruit antitumor CD8+ T lymphocytes into malignancies and are positively associated with the survival of patients with colorectal cancer or hepatocellular carcinoma [30, 37].
Understanding the molecular basis of T lymphocytes accumulation in tumors is important for the improvement of immune cell-based therapy. As the tissue-specific migration of both T and CIK cells is dependent on chemotaxis, the movement of these tumor-killing cells to tumor sites would be enhanced when the interactions between chemokine ligand and receptor are employed by tumor-infiltrating T cells [14, 15, 38]. Recent technical advances in gene modification make it easy to introduce specific receptors, including chemokine receptors into T and CIK cells [14, 39, 40]. Therefore, the key for efficient delivery of tumor-killing cells is to determine which chemokines are involved in immune cells in vivo, especially for T lymphocyte trafficking. Considering that immune cells are prone to migrate to sites with higher concentrations of chemokines [2, 3], the chemokines highly expressed in tumor sites are good candidates. The expression of chemokines in tumors is complex. Some chemokines are upregulated in tumor tissues [41–43], while others are downregulated [44, 45]. In our study, CCL5 and CXCL10 were more highly expressed in cancerous tissues than in marginal sites; the chemokines’ expression was not affected by patients’ age, gender, tumor differentiation, or clinical TNM stages, except that CCL5 was further upregulated in clinical T3-T4 stages (Table 1). These data imply that CCL5 and CXCL10 are ideal candidates for the effective delivery of T and CIK cells.
CCL5 and CXCL10, together with CCR5 and CXCR3, recruit CD8+ T lymphocytes into ESCC tissue. Moreover, CCL5 and CXCL10 are overexpressed in malignant lesions, suggesting that CCR5 and CXCR3 induction would enhance the tumoral delivery of infused T and CIK cells and improve the curative efficacy of such adoptive therapy.
MATERIALS AND METHODS
Clinical samples
A total of 207 patients with esophageal squamous cell carcinoma were enrolled in this study, which was approved by the Ethics Committee Board of the First Affiliated Hospital of Zhengzhou University. All patients signed informed consent forms. Tumor stage and differentiation was graded according to the classification system of the American Joint Committee on Cancer (AJCC, 6th Edition). The detailed information of patients is listed in Table 2. During surgery from June 2008 through March 2014 in the First Affiliated Hospital of Zhengzhou University, 207 tumors and 61 paired peripheral blood samples were collected. In addition, 21 marginal tissues were sampled.
Table 2: Association of CCL5 and CXCL10 scores with clinicopathological factors
CCL5 |
CXCL10 |
|||||
N |
Mean ± SD |
p-value |
N |
Mean ± SD |
p-value |
|
Gender |
||||||
Male |
86 |
1.904 ± 1.748 |
0.7871 |
94 |
2.384 ± 1.905 |
0.4189 |
Female |
33 |
1.811 ± 1.508 |
36 |
2.701 ± 2.234 |
||
Age |
||||||
≤ 60 |
47 |
1.888 ± 2.018 |
0.9578 |
53 |
2.486 ± 2.046 |
0.9463 |
> 60 |
72 |
1.872 ± 1.432 |
77 |
2.462 ± 1.977 |
||
Clinical stage |
||||||
I–IIa |
82 |
1.902 ± 1.712 |
0.8155 |
92 |
2.395 ± 2.081 |
0.4963 |
IIb–IV |
37 |
1.824 ± 1.625 |
38 |
2.658 ± 1.793 |
||
T stage |
||||||
T1–T2 |
59 |
1.555 ± 1.193 |
0.0369 |
65 |
2.254 ± 1.617 |
0.3487 |
T3–T4 |
60 |
2.196 ± 2.009 |
65 |
2.566 ± 2.133 |
||
Differentiation |
||||||
Poor |
59 |
1.747 ± 1.203 |
0.4022 |
62 |
2.424 ± 1.802 |
0.7975 |
Well |
60 |
2.007 ± 2.045 |
68 |
2.515 ± 2.173 |
||
Lymph node metastasis |
||||||
Negative |
87 |
1.977 ± 1.716 |
0.2916 |
97 |
2.364 ± 2.049 |
0.2940 |
Positive |
32 |
1.609 ± 1.570 |
33 |
2.788 ± 1.832 |
||
Tumor length |
||||||
≤ 3 cm |
57 |
2.013 ± 1.771 |
0.4027 |
62 |
2.477 ± 1.971 |
0.9781 |
> 3 cm |
62 |
1.754 ± 1.595 |
68 |
2.467 ± 2.036 |
||
Isolation of TILs and PBLs
Tumor tissues were rinsed with Roswell Park Memorial Institute 1640 (RPMI-1640) media (Invitrogen Life Technologies, Carlsbad, CA, USA) to remove traces of blood and then cut into small pieces. The tissues were incubated with 300 μg/mL collagenase (Roche, Indianapolis, IN, USA) and 50 μg/mL deoxyribonuclease I (Sigma-Aldrich, St. Louis, MO, USA) for 2 h at 37°C. Following that, the samples were mechanically disaggregated. TILs were separated by centrifugation at 2,000 rpm for 20 min on Ficoll-Paque Plus (Sigma-Aldrich, St. Louis, MO, USA). PBLs were isolated from heparinized blood samples by Ficoll-Paque Plus density centrifugation.
Flow cytometry
TILs and PBLs were suspended in flow buffer (Phosphate-buffered saline containing 2% fetal bovine serum) and incubated with anti-CD3-APC (HIT3a), anti-CD8-APC/Cy7 (HIT8a), anti-CCR5-PE/Cy7 (J418F1), anti-CXCR3-PE (G025H7), or isotype-matched antibodies (Biolegend, San Diego, CA, USA) for 30 min at 4°C. After incubation, cells were rinsed twice. Then, cells were analyzed using a BD CantoII flow cytometer (Becton Dickinson, San Jose, CA, USA).
RT-PCR
Tumor tissues were cut into 20 mm pieces and mechanically minced. Then, total RNA was extracted using Trizol solution (Invitrogen Life Technologies, Carlsbad, CA, USA). Subsequently, RNA from each sample was separately reverse-transcribed using the PrimeScript RT Reagent Kit (Takara Bio, Otsu, Shiga, Japan). Then, the genes of interest were tested. Primers used were listed in Table 3. The initial step was performed at 95°C for 30 s and the amplification was performed for 30 cycles of 95°C for 15 s, 58°C for 30 s, and 72°C for 30s. PCR products were separated on a 1.5% agarose gel and recorded. Targeted bands were analyzed using ImageJ software (National Institute of Health, USA) and optical densities were calculated. Then, the relative expression of genes was determined. The house-keeping gene GAPDH was used as reference.
Table 3: The sequences of primers used
Gene |
Sequence |
GAPDH |
Sense: 5′-GGAGCCAAAAGGGTCATCATCTC-3′ |
Anti-sense: 5′-GAGGGGCCATCCACAGTCTTCT-3′ |
|
CD8 |
Sense: 5′-CGCTGTCAGATCCCCTTTGT-3′ |
Anti-sense: 5′-GAGGAAGGACCCTCTCCCTT-3′ |
|
Granzyme B |
Sense: 5′-GCAGGAAGATCGAAAGTGCG-3′ |
Anti-sense: 5′-TACAGCGGGGGCTTAGTTTG-3′ |
|
CCL5 |
Sense: 5′-CAGTCGTCTTTGTCACCCGA-3′ |
Anti-sense: 5′-TGTAACTGCTGCTGTGTGGT-3′ |
|
CXCL10 |
Sense: 5′-AACTGTACGCTGTACCTGCAT-3′ |
Anti-sense: 5′-GCATCGATTTTGCTCCCCTC-3′ |
Immunohistochemistry
Formalin-fixed paraffin-embedded sections (3 μm) were de-paraffinized in xylene, rehydrated through graded alcohol, and washed briefly in tap water. Endogenous peroxidase was blocked with methanol containing 0.3% hydrogen peroxide for 30 min. To retrieve antigenicity, sections were boiled in 10 mM citrate buffer (pH 5.8) for 30 min in a microwave oven (800 W). Next, sections were incubated with goat serum diluted in PBS (pH 7.4) for 30 min at room temperature. Subsequently, sections were incubated at 4°C overnight with the primary antibodies specific for CCL5 (dilution 1: 200) or CXCL10 (dilution 1:200) (Abcam, Cambridge, UK). Then, sections were rinsed in fresh PBS and incubated with horseradish peroxidase-linked secondary antibodies at room temperature for 60 min. Finally, sections were stained with 3, 3′-diaminobenzidine (DAB) substrate (Dako, Carpinteria, CA, USA) and counterstained with Mayer's hematoxylin. Non-immune rabbit IgG at the same dilution was used as negative control. Photos were recorded under a microscope (Leica, Wetzlar, Germany).
Evaluation of immunohistochemical staining
All sections were assessed at 20 × magnification by 2 experienced observers. The staining was evaluated based on the intensity (weak = 1, moderate = 2, and high = 3) of chemokine immunostaining and the density (0% = 0, 1–40% = 1, 41–75% = 2, > 76% = 3) of positive tumor cells. The final score of each sample was multiplied by the intensity and density. If the two evaluations did not agree, the specimens were re-evaluated and then classified according to the assessment given most frequently by the observers. For CCL5 and CXCL10, samples with a score > 1.50 were regarded as having high expression.
Purification of CD8+ T lymphocytes
TILs were isolated from fresh ESCC tissues as mentioned above. Then, CD8+ T lymphocytes were further purified by positive selection using CD8 microbeads (Miltenyi Biotec, Auburn, CA, USA) according to the manufacturer's protocol. Briefly, 107 TILs were suspended in magnetic-activated cell sorting (MACS) buffer, incubated with 20 μL CD8 microbeads at 4°C for 15 min, and washed once. Then, CD8+ T lymphocytes were magnetically isolated.
Transwell assay
The chemotactic migration of CD8+ T lymphocytes was evaluated in 24-well plates with 5-μm pore size polycarbonate filters (Corning Inc, Coring, NY, USA) [8]. First, ESCC tumor tissues were cut into small pieces and cultured in Dulbecco's Modified Eagle Medium (DMEM) with 10% fetal bovine serum (FBS) for 48 h. Then, 600 μL of tumor supernatants were placed into the lower chambers of transwell plates. Anti-CCL5 (1 μg/mL) and/or anti-CXCL10 (5 μg/mL) neutralizing antibodies were added as indicated. DMEM media with 10% FBS were used as a control. Purified CD8+ T lymphocytes from TILs (Purity > 90%) were counted. Then 5 × 105 CD8+ T lymphocytes were added into the upper chambers and incubated at 37°C in a 5% CO2 atmosphere for 2 h. Cells in the bottom chambers were counted using a limited 60-second analysis on a flow cytometer.
Statistical analysis
Analyses were performed using GraphPad Prism 5 software (GraphPad Software, La Jolla, CA, USA). Data were expressed as mean ± SD. The Student's t test and one-way ANOVA were conducted to compare the differences between variables. The Spearman test was adapted to determine the correlation between chemokine genes and CD8+ T lymphocyte-related markers. Kaplan-Meier curves in combination with log-rank tests were used to compare the survival of patients in different groups. Values with p < 0.05 (two-tailed) were considered significant.
ACKNOWLEDGMENTS
This study was supported by grants from the China-US (NFSC-NIH) Program for Biomedical Collaborative Research (Grant no.812111102), the National Natural Science Foundation of China (Grant no.81171986), the Ministry of Public Health (Grant no.20110110001), and the Basic and Advanced Technology Research Foundation from Science and Technology Department of Henan Province (Grant nos.112300410153 and 122300410155).
CONFLICTS OF INTEREST
All authors declare that they have no conflicts of interest.
REFERENCES
1. Charo IF, Ransohoff RM. The many roles of chemokines and chemokine receptors in inflammation. N Engl J Med. 2006; 354:610–621.
2. Masopust D, Schenkel JM. The integration of T cell migration, differentiation and function. Nat Rev Immunol. 2013; 13:309–320.
3. Balkwill F. Cancer and the chemokine network. Nat Rev Cancer. 2004; 4:540–550.
4. Oldham KA, Parsonage G, Bhatt RI, Wallace DM, Deshmukh N, Chaudhri S, Adams DH, Lee SP. T lymphocyte recruitment into renal cell carcinoma tissue: a role for chemokine receptors CXCR3, CXCR6, CCR5, and CCR6. Eur Urol. 2012; 61:385–394.
5. Parsonage G, Machado LR, Hui JW, McLarnon A, Schmaler T, Balasothy M, To KF, Vlantis AC, van Hasselt CA, Lo KW, Wong WL, Hui EP, Chan AT, Lee SP. CXCR6 and CCR5 localize T lymphocyte subsets in nasopharyngeal carcinoma. Am J Pathol. 2012; 180:1215–1222.
6. Kim HJ, Song DE, Lim SY, Lee SH, Kang JL, Lee SJ, Benveniste EN, Choi YH. Loss of the promyelocytic leukemia protein in gastric cancer: implications for IP-10 expression and tumor-infiltrating lymphocytes. PLoS One. 2011; 6:e26264.
7. Zumwalt TJ, Arnold M, Goel A, Boland CR. Active secretion of CXCL10 and CCL5 from colorectal cancer microenvironments associates with GranzymeB+ CD8+ T-cell infiltration. Oncotarget. 2015; 6:2981–2991.
8. Muthuswamy R, Berk E, Junecko BF, Zeh HJ, Zureikat AH, Normolle D, Luong TM, Reinhart TA, Bartlett DL, Kalinski P. NF-kappaB hyperactivation in tumor tissues allows tumor-selective reprogramming of the chemokine microenvironment to enhance the recruitment of cytolytic T effector cells. Cancer Res. 2012; 72:3735–3743.
9. Berghuis D, Santos SJ, Baelde HJ, Taminiau AH, Egeler RM, Schilham MW, Hogendoorn PC, Lankester AC. Pro-inflammatory chemokine-chemokine receptor interactions within the Ewing sarcoma microenvironment determine CD8(+) T-lymphocyte infiltration and affect tumour progression. J Pathol. 2011; 223:347–357.
10. Zhao JS, Li WJ, Ge D, Zhang PJ, Li JJ, Lu CL, Ji XD, Guan DX, Gao H, Xu LY, Li EM, Soukiasian H, Koeffler HP, Wang XF, Xie D. Tumor initiating cells in esophageal squamous cell carcinomas express high levels of CD44. PLoS One. 2011; 6:e21419.
11. Hang D, Dong HC, Ning T, Dong B, Hou DL, Xu WG. Prognostic value of the stem cell markers CD133 and ABCG2 expression in esophageal squamous cell carcinoma. Dis Esophagus. 2012; 25:638–644.
12. Mariette C, Piessen G, Triboulet JP. Therapeutic strategies in oesophageal carcinoma: role of surgery and other modalities. Lancet Oncol. 2007; 8:545–553.
13. Franciszkiewicz K, Boissonnas A, Boutet M, Combadiere C, Mami-Chouaib F. Role of chemokines and chemokine receptors in shaping the effector phase of the antitumor immune response. Cancer Res. 2012; 72:6325–6332.
14. Asai H, Fujiwara H, An J, Ochi T, Miyazaki Y, Nagai K, Okamoto S, Mineno J, Kuzushima K, Shiku H, Inoue H, Yasukawa M. Co-Introduced Functional CCR2 Potentiates In Vivo Anti-Lung Cancer Functionality Mediated by T Cells Double Gene-Modified to Express WT1-Specific T-Cell Receptor. PLoS One. 2013; 8:e56820.
15. Zou Y, Li F, Hou W, Sampath P, Zhang Y, Thorne SH. Manipulating the expression of chemokine receptors enhances delivery and activity of cytokine-induced killer cells. Br J Cancer. 2014; 110:1992–1999.
16. Sugiyama D, Nishikawa H, Maeda Y, Nishioka M, Tanemura A, Katayama I, Ezoe S, Kanakura Y, Sato E, Fukumori Y, Karbach J, Jager E, Sakaguchi S. Anti-CCR4 mAb selectively depletes effector-type FoxP3+CD4+ regulatory T cells, evoking antitumor immune responses in humans. Proc Natl Acad Sci U S A. 2013; 110:17945–17950.
17. Sherwood AM, Emerson RO, Scherer D, Habermann N, Buck K, Staffa J, Desmarais C, Halama N, Jaeger D, Schirmacher P, Herpel E, Kloor M, Ulrich A, Schneider M, Ulrich CM, Robins H. Tumor-infiltrating lymphocytes in colorectal tumors display a diversity of T cell receptor sequences that differ from the T cells in adjacent mucosal tissue. Cancer Immunol Immunother. 2013; 62:1453–1461.
18. Salerno EP, Olson WC, McSkimming C, Shea S, Slingluff CL Jr. T cells in the human metastatic melanoma microenvironment express site-specific homing receptors and retention integrins. Int J Cancer. 2014; 134:563–574.
19. Wehler TC, Graf C, Altherr K, Zimmermann T, Brenner W, Thuroff JW, Biesterfeld S, Gockel I, Theobald M, Galle PR, Schimanski CC. SDF1beta expression in renal cell carcinoma correlates with grading and infiltration by CD8+ T-cells. Anticancer Res. 2011; 31:2797–2803.
20. Gu-Trantien C, Loi S, Garaud S, Equeter C, Libin M, de Wind A, Ravoet M, Le Buanec H, Sibille C, Manfouo-Foutsop G, Veys I, Haibe-Kains B, Singhal SK, Michiels S, Rothe F, Salgado R, et al. CD4+ follicular helper T cell infiltration predicts breast cancer survival. J Clin Invest. 2013; 123:2873–2892.
21. Winkler AE, Brotman JJ, Pittman ME, Judd NP, Lewis JS Jr, Schreiber RD, Uppaluri R. CXCR3 enhances a T-cell-dependent epidermal proliferative response and promotes skin tumorigenesis. Cancer Res. 2011; 71:5707–5716.
22. Liu J, Zhang N, Li Q, Zhang W, Ke F, Leng Q, Wang H, Chen J. Tumor-associated macrophages recruit CCR+ regulatory T cells and promote the development of colorectal cancer via enhancing CCL20 production in mice. PLoS One. 2011; 6:e19495.
23. Chen KJ, Lin SZ, Zhou L, Xie HY, Zhou WH, Taki-Eldin A, Zheng SS. Selective recruitment of regulatory T cell through CCR6-CCL20 in hepatocellular carcinoma fosters tumor progression and predicts poor prognosis. PLoS One. 2011; 6:e24671.
24. Zhang T, Somasundaram R, Berking C, Caputo L, Van Belle P, Elder D, Czerniecki B, Hotz S, Schuchter L, Spitz FR, Berencsi K, Rani P, Marincola F, Qiu R, Herlyn D. Preferential involvement of CX chemokine receptor 4 and CX chemokine ligand 12 in T-cell migration toward melanoma cells. Cancer Biol Ther. 2006; 5:1304–1312.
25. Tachezy M, Zander H, Gebauer F, von Loga K, Pantel K, Izbicki JR, Bockhorn M. CXCR7 expression in esophageal cancer. J Transl Med. 2013; 11:238.
26. Verbeke H, Geboes K, Van Damme J, Struyf S. The role of CXC chemokines in the transition of chronic inflammation to esophageal and gastric cancer. Biochim Biophys Acta. 2012; 1825:117–129.
27. Musha H, Ohtani H, Mizoi T, Kinouchi M, Nakayama T, Shiiba K, Miyagawa K, Nagura H, Yoshie O, Sasaki I. Selective infiltration of CCR5(+)CXCR3(+) T lymphocytes in human colorectal carcinoma. Int J Cancer. 2005; 116:949–956.
28. Harlin H, Meng Y, Peterson AC, Zha Y, Tretiakova M, Slingluff C, McKee M, Gajewski TF. Chemokine expression in melanoma metastases associated with CD8+ T-cell recruitment. Cancer Res. 2009; 69:3077–3085.
29. Mulligan AM, Raitman I, Feeley L, Pinnaduwage D, Nguyen LT, O’Malley FP, Ohashi PS, Andrulis IL. Tumoral Lymphocytic Infiltration and Expression of the Chemokine CXCL10 in Breast Cancers from the Ontario Familial Breast Cancer Registry. Clin Cancer Res. 2013; 19:336–346.
30. Chew V, Chen J, Lee D, Loh E, Lee J, Lim KH, Weber A, Slankamenac K, Poon RT, Yang H, Ooi LL, Toh HC, Heikenwalder M, Ng IO, Nardin A, Abastado JP. Chemokine-driven lymphocyte infiltration: an early intratumoural event determining long-term survival in resectable hepatocellular carcinoma. Gut. 2012; 61:427–438.
31. Zlotnik A, Yoshie O. The chemokine superfamily revisited. Immunity. 2012; 36:705–716.
32. Lanca T, Costa MF, Goncalves-Sousa N, Rei M, Grosso AR, Penido C, Silva-Santos B. Protective role of the inflammatory CCR2/CCL2 chemokine pathway through recruitment of type 1 cytotoxic gammadelta T lymphocytes to tumor beds. J Immunol. 2013; 190:6673–6680.
33. Chen D, Jiang R, Mao C, Shi L, Wang S, Yu L, Hu Q, Dai D, Xu H. Chemokine/chemokine receptor interactions contribute to the accumulation of Th17 cells in patients with esophageal squamous cell carcinoma. Hum Immunol. 2012; 73:1068–1072.
34. Rooney MS, Shukla SA, Wu CJ, Getz G, Hacohen N. Molecular and genetic properties of tumors associated with local immune cytolytic activity. Cell. 2015; 160:48–61.
35. Sato E, Olson SH, Ahn J, Bundy B, Nishikawa H, Qian F, Jungbluth AA, Frosina D, Gnjatic S, Ambrosone C, Kepner J, Odunsi T, Ritter G, Lele S, Chen YT, Ohtani H, et al. Intraepithelial CD8+ tumor-infiltrating lymphocytes and a high CD8+/regulatory T cell ratio are associated with favorable prognosis in ovarian cancer. Proc Natl Acad Sci U S A. 2005; 102:18538–18543.
36. Djenidi F, Adam J, Goubar A, Durgeau A, Meurice G, de Montpreville V, Validire P, Besse B, Mami-Chouaib F. CD8+CD103+ Tumor-Infiltrating Lymphocytes Are Tumor-Specific Tissue-Resident Memory T Cells and a Prognostic Factor for Survival in Lung Cancer Patients. J Immunol. 2015; 194:3475–3486.
37. Mlecnik B, Tosolini M, Charoentong P, Kirilovsky A, Bindea G, Berger A, Camus M, Gillard M, Bruneval P, Fridman WH, Pages F, Trajanoski Z, Galon J. Biomolecular network reconstruction identifies T-cell homing factors associated with survival in colorectal cancer. Gastroenterology. 2010; 138:1429–1440.
38. Thanarajasingam U, Sanz L, Diaz R, Qiao J, Sanchez-Perez L, Kottke T, Thompson J, Chester J, Vile RG. Delivery of CCL21 to metastatic disease improves the efficacy of adoptive T-cell therapy. Cancer Res. 2007; 67:300–308.
39. Urbanska K, Lanitis E, Poussin M, Lynn RC, Gavin BP, Kelderman S, Yu J, Scholler N, Powell DJ Jr. A universal strategy for adoptive immunotherapy of cancer through use of a novel T-cell antigen receptor. Cancer Res. 2012; 72:1844–1852.
40. Hombach AA, Rappl G, Abken H. Arming Cytokine-induced Killer Cells With Chimeric Antigen Receptors: CD28 Outperforms Combined CD28-OX40 “Super-stimulation”. Mol Ther. 2013; 21:2268–2277.
41. Miyake M, Lawton A, Goodison S, Urquidi V, Gomes-Giacoia E, Zhang G, Ross S, Kim J, Rosser CJ. Chemokine (C-X-C) ligand 1 (CXCL1) protein expression is increased in aggressive bladder cancers. BMC Cancer. 2013; 13:322.
42. Zeng J, Yang X, Cheng L, Liu R, Lei Y, Dong D, Li F, Lau QC, Deng L, Nice EC, Xie K, Huang C. Chemokine CXCL14 is associated with prognosis in patients with colorectal carcinoma after curative resection. J Transl Med. 2013; 11:6.
43. Panse J, Friedrichs K, Marx A, Hildebrandt Y, Luetkens T, Barrels K, Horn C, Stahl T, Cao Y, Milde-Langosch K, Niendorf A, Kroger N, Wenzel S, Leuwer R, Bokemeyer C, Hegewisch-Becker S, et al. Chemokine CXCL13 is overexpressed in the tumour tissue and in the peripheral blood of breast cancer patients. Br J Cancer. 2008; 99:930–938.
44. Hu C, Lin F, Zhu G, Xue X, Ding Y, Zhao Z, Zhang L, Shen X. Abnormal hypermethylation of promoter region downregulates chemokine CXC ligand 14 expression in gastric cancer. Int J Oncol. 2013; 43:1487–1494.
45. Pivarcsi A, Muller A, Hippe A, Rieker J, van Lierop A, Steinhoff M, Seeliger S, Kubitza R, Pippirs U, Meller S, Gerber PA, Liersch R, Buenemann E, Sonkoly E, Wiesner U, Hoffmann TK, et al. Tumor immune escape by the loss of homeostatic chemokine expression. Proc Natl Acad Sci U S A. 2007; 104:19055–19060.