
Introduction
In breast cancer the hormone receptor status is one of the most important predictive factor [1]. Since patients with positive hormone receptor status are eligible for endocrine adjuvant and palliative therapy, assessment of estrogen (ER) and progesterone (PR) receptor is routinely carried out. ER-positive tumors are classified as (highly hormone responsive) luminal A or (high proliferative and less hormone responsive) luminal B tumors. Targeted therapies that inhibit ER or ER-activated pathways include the selective ER modulator tamoxifen and the aromatase inhibitor arimidex. Anti-HER2 (human epidermal growth factor receptor 2) therapies include the administration of trastuzumab and lapatinib. Unfortunately, in spite of receptor positivity, many tumors can develop resistance to therapy, and the expression pattern of ER, PR and HER2 may change over time as tumors progress and metastasize [2].
MicroRNAs (miRs) are evolutionary conserved, small non-coding RNA molecules consisting of approximately 22 nucleotides. MiRs inhibit the gene expression post-transcriptionally by binding specifically to the 3’untranslated-region (UTR) of their target mRNA. Gene silencing can occur through the translational inhibition or cleavage of their target mRNAs depending on the complementary sequence between binding sites of the specific mRNA and miR [3]. Computational analyses indicate that one miR has binding affinity to hundreds of different mRNAs and hence, miRs are involved in the regulation of various cellular processes, such as development, differentiation and proliferation [4].
As miR loci frequently map to fragile chromosomal regions harboring DNA amplifications, deletions or translocations, their expression is often deregulated during tumorigenesis, contributing to tumor progression [5-8]. In this context, they may act as so-called oncomiRs by targeting tumor suppressor genes or as tumor suppressor miRs by inhibiting oncogenes [9]. MiRs are released into the blood circulation by apoptotic and necrotic cells or active secretion [10], and exist either cell-freely, associated with Argonaut proteins, or in exosomes in the blood circulation [11]. To date, numerous miRs have been identified, those transcript levels were dysregulated in the blood of breast cancer patients [12, 13].
In the present study, we focused our analyses on circulating miR-27a, miR-107, miR-130a, miR-132, miR-146a and miR-16, those dysregulated transcript levels were detected in patients with invasive breast cancer by microarray profiling. MiR-27a, that promotes angiogenesis by mediating endothelial differentiation of breast cancer stem like cells [14], has been associated with poor overall survival of breast cancer patients and suggested to be a valuable marker for tumor progression [15]. MiR-107 targets numerous transcripts including estrogen-responsive gene clusters, and its intracellular levels are affected by estrogen [16]. MiR-130a contributes to the growth inhibition induced by retinoic acid that is used as a chemopreventive agent for breast cancer [17]. Overexpression of miR-132 has been reported to inhibit proliferation of breast cancer cells and their colony formation [18]. Transduction of miR-146a may down-regulate expression of epidermal growth factor receptor (EGFR), and inhibit invasion and migration of breast cancer cells [19]. To date, miR-16 has been often used as reference miR to normalize miR data, because this miR showed the smallest variation in the populations, but miR-16 has also been elevated in osteoclast differentiation associated with bone metastasis burden [10, 20].
The aim of this study was to investigate whether the dysregulated levels of circulating miR-27a, miR-107, miR-130a, miR-132, miR-146a and miR-16 are associated with the receptor status of breast cancer. Screening for plasma miRs could provide valuable non-invasive information on the luminal subtype of breast cancer. The effect of adjuvant chemotherapy on the miR levels was also investigated.
Results
Patients
For this retrospective study, we chose 130 patients with invasive breast cancer from the SUCCESS study based on their receptor status, to assemble a collective that showed a well-balanced distribution of established risk factors including hormone receptor status, lymph node status, tumor stages, and grading. Of this cohort, 42% of the patients were triple-negative and/or had lymph node metastases (Figure 1). This highly selected cohort does not represent the population of the SUCCESS study (www.success-studie.de). Blood plasma samples were taken from these patients about 1 or 2 months after surgery before initiation of chemotherapy and additionally, collected after chemotherapy. For our miR analyses, 111 out of 130 patients were eligible. All patients analyzed had histologic proven epithelial cancer. Table 1 summarizes the clinical and histopathologic parameters of the breast cancer patient cohort.
Table 1: Patient characteristics and histopathological parameters at the time after surgery.

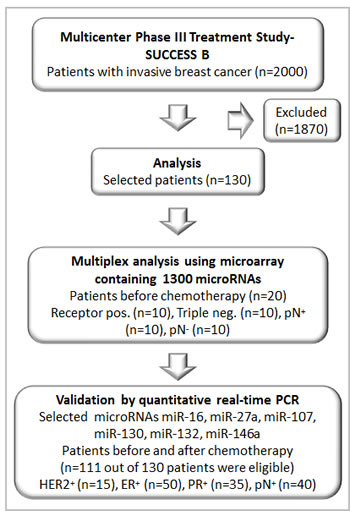
Figure 1: Consort diagram showing the number of patients analyzed from the multicenter study (SUCCESS B).
MiR profiling using a blood-based microarray
For blood-based miR profiling, microfluid biochips containing 1300 different miRs were used to quantify the expression of miRs in the plasma samples of 20 breast cancer patients and 10 healthy women. To detect differentially regulated miRs, the quotation of median, paired Student’s t-test and limma test were assessed. Dysregulated miRs were detected by the highest absolute value of logarithmized bold changes in comparison of breast cancer patients with healthy women. The estimated raw p-values were adjusted for multiple testing, to control the false discovery rate. Table 2 shows 30 most differentially expressed miRs with the adjusted p-values as determined by t-test and limma test. In addition, the normalized median values of healthy women and breast cancer patients are listed, and the comparison of these values in both cohorts indicates the down- or upregulated miRs. A similarity matrix was generated containing all pairwise similarities of the plasma samples of breast cancer patients and healthy women. To detect potential clusters in rows (transcripts) and columns (samples) of the normalized expression matrix, hierarchical clustering was carried out (Figure 2). Based on the highest overall variability derived from these array data (Table 2), we selected miR-16, miR-27a, miR-107 and miR-130a for further validation studies. In addition, we also quantified miR-132 and miR-146a, because of their particular role in breast cancer [18, 21]. MiR-1207 was chosen as reference miR, because it displayed the smallest data variations in the miR profiling.
Table 2: Differentially regulated microRNAs in healthy individuals versus breast cancer patients.
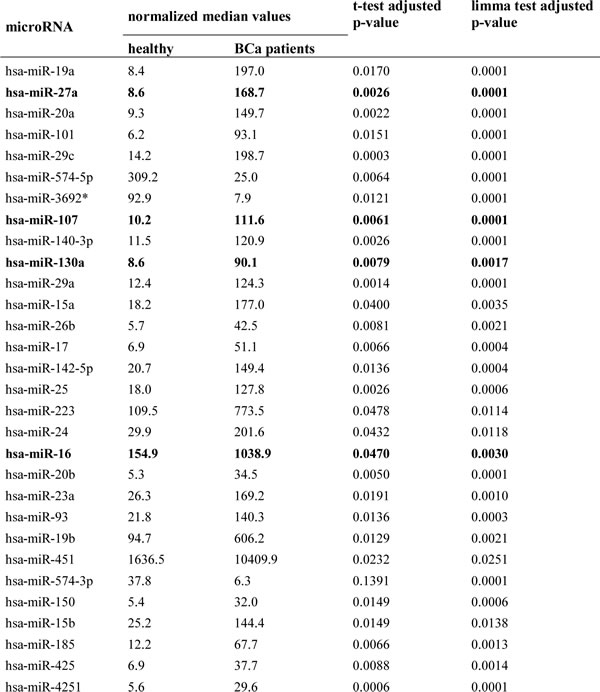
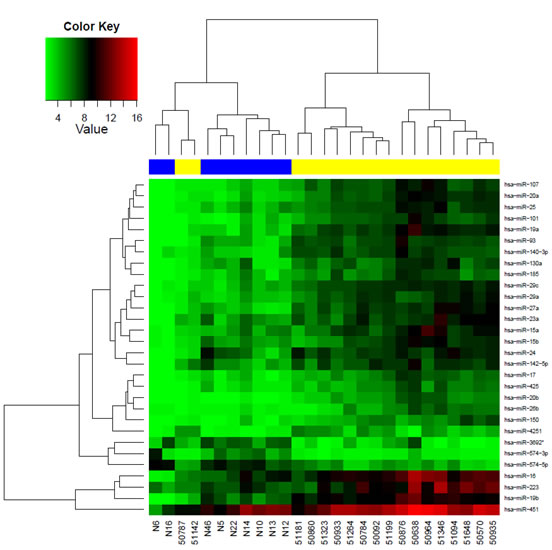
Figure 2: MiR profiling using a blood-based microarray. Hierarchical cluster heat map of miR microarray was performed using microfluid biochips containing 1300 different miRs and postoperative plasma of 20 breast cancer patients before chemotherapy and 10 healthy women. The colored representation of samples and probes is ordered by their similarity with a dendogram on top (clustering of samples) and on the right side (clustering of probes).
Profiling of cell-free miR-16, miR-27a, miR-107, miR-130a, miR-132 and miR-146a in the postoperative plasma of breast cancer patients before and after chemotherapy
Before we quantified the relative transcript levels of our panel of selected miRs in the plasma of 111 patients with invasive breast cancer before and after chemotherapy and 46 age-matched healthy women by quantitative real-time PCR, we measured the total small RNA in the plasma of both cohorts. The plasma RNA levels were higher in patients before chemotherapy than in healthy women (p = 0.0001). The significant increased plasma RNA concentrations detected before chemotherapy significantly decreased after chemotherapy (p = 0.001), but were still higher than the plasma levels in healthy women (p = 0.021, Supplementary Figure S1).
Then, we quantified cell-free miR-16, miR-27a, miR-107, miR-130a, miR-132 and miR-146a by Taqman PCR. As shown in the box plot of Figure 3A, the postoperative plasma levels of miR-16 (p = 0.0001, Mann-Whitney-U test), miR-27a (p = 0.039) and miR-132 (p = 0.020) were significantly higher in breast cancer patients before chemotherapy than in healthy women. The AUC values of miR-16, miR-27a and miR-132 were 0.685 (p = 0.0001), 0.605 (p = 0.039) and 0.618 (p = 0.020), respectively, showing the significant difference of the miR levels between patients before chemotherapy and healthy women (Figure 3B). In contrast, the concentrations of miR-107, miR-130a and miR-146a were similar between the two cohorts. After chemotherapy, the elevated levels of miR-27a (p = 0.035, Wilcoxon test) and miR-132 (p = 0.025) decreased to similar levels as observed in healthy women, whereas the levels of miR-16 remained significantly increased (Figure 3A).
No association of the different treatment arms with the miR levels after chemotherapy was observed. Unfortunately, in the multicenter SUCCESS trial data on endocrine therapy of only 43 patients were available. Of these patients, 17 and 26 patients got tamoxifen and arimidex, respectively. We did not carry out a statistical evaluation, because the patient cohort was too small.
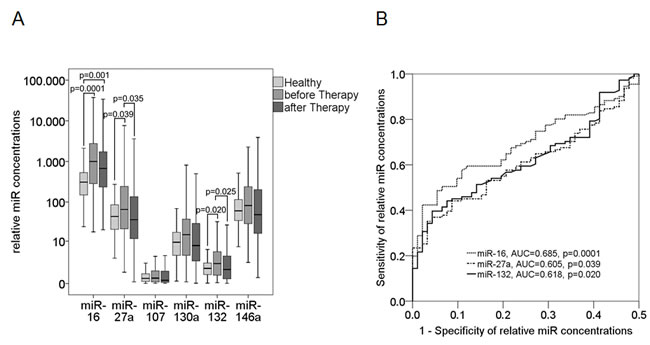
Figure 3: Plasma levels of circulating miR-16, miR-27a, miR-107, miR-130a, miR-132 and miR-146a in breast cancer patients before and after chemotherapy. The box plot compares the miR concentrations in plasma of healthy women (n = 46) with breast cancer patients before (n = 111) and after chemotherapy (n = 111). The relative expression levels were determined by the low cycle threshold (Ct) values. As determined by Mann-Whitney-U and Wilcoxon test, the significant p-values of statistical evaluations are indicated A.. ROC analyses show the profiles of sensitivity and specificity of miR-16, miR-27a and miR-132 to differentiate healthy individuals from breast cancer patients before chemotherapy B..
Association of the plasma levels of miR-16, miR-107, miR-130a and miR-146a with lymph node and receptor status
We compared the relative concentrations of circulating miR-16, miR-27a, miR-107, miR-130a, miR-132 and miR-146a in the postoperative plasma of 111 breast cancer patients before chemotherapy with the clinical and histopathological risk factors of these patients. To determine the diagnostic value of these miRs, the Mann-Whitney-U test was applied. The Supplementary Table S1 summarizes mean and median values, 95% confidence intervals (CI) and p values of the miR variables in the different patient subgroups.
Albeit the significance was only marginal, the plasma levels of miR-16 (Figure 4A, p = 0.019), miR-107 (Figure 4B, p = 0.036), miR-130a (Figure 4C, p = 0.027), and miR-146a (Figure 4D, p = 0.047) could differ between lymph-node negative and positive patients. The elevated plasma levels of miR-16 (Figure 4A, p = 0.0001), miR-130a (Figure 4C, p = 0.006) and miR-146a (Figure 4D, p = 0.023) detected in lymph-node negative patients decreased in lymph-node positive patients to similar levels as observed in healthy women, suggesting that these miRs play rather a role in early breast cancer.
In addition, the plasma levels of miR-107 could distinguish between ER-negative and -positive status and were significantly elevated in ER-negative patients (Figure 4B, p = 0.035). The plasma concentrations of miR-130a (Figure 4C, p = 0.001) and miR-146a (Figure 4D, p = 0.025) could differ between HER2-negative and -positive tumors, and were significantly elevated in HER2-positive patients.
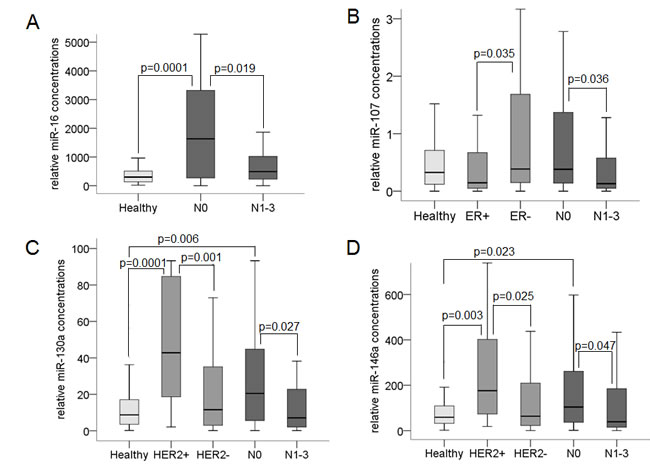
Figure 4: Association of cell-free miR-16, miR-107, miR-130a and miR-146a with lymph node and receptor status. The box plots compare the plasma concentrations of miR-16 in lymph node-negative (n = 71) with those in lymph node-positive patients (n = 40) A., of miR-107 in estrogen-positive (n = 50) with those in estrogen-negative patients (n = 61) and in lymph node-negative (n = 71) with those in lymph node-positive patients (n = 40) B., and of miR-130a C. and miR-146a D. in HER2-positive (n = 15) with those in HER2-negative patients (n = 96) and in lymph node-negative (n = 71) with those in lymph node-positive patients (n = 40). As determined by the Mann-Whitney-U test, the significant p-values of statistical evaluations are indicated.
Expression levels of miR-16, miR-27a, miR-107, miR-130a, miR-132 and miR-146a in MCF-7 and MDA-MB-231 cells
The relative expression levels of our miR panel were determined in non-invasive, ER-positive breast adenocarcinoma MCF-7 cells and invasive, ER-negative MDA-MB-231 cells [22] by quantitative real-time PCR (Figure 5). These breast adenocarcinoma cell lines showed a heterogeneous miR pattern. The transcript levels of miR-16, miR-27a, miR-107 and miR-132 were similar in both cell lines, while miR-130a (p = 0.0001) and miR-146a (p = 0.0001) were significantly higher expressed in MDA-MB-231 cells than MCF-7 cells.
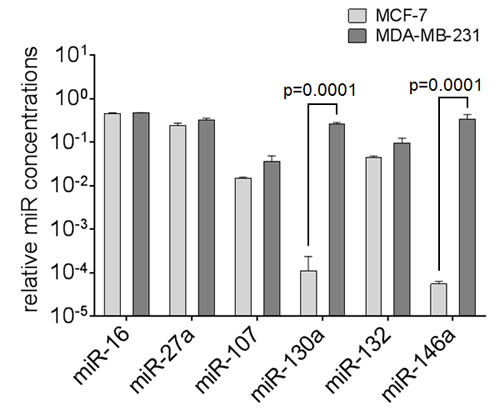
Figure 5: Expression levels of circulating miR-16, miR-27a, miR-107, miR-130a, miR-132 and miR-146a in breast cancer cell lines. The bar chart compares the miR concentrations in non-invasive MCF-7 cells with those in invasive MDA-MB-231. The relative expression levels were determined by the low cycle threshold (Ct) values. The significant p-values as determined by the one-way ANOVA test and standard deviations from triplicate experiments are indicated.
MiR-107 does not affect the protein expression of ER
Since our findings suggest that miR-107 is associated with ER-negative breast cancer, we examined whether ER is a target of miR-107. Therefore, we screened the miR databases DIANA-microT-CDS [23], microRNA.org [24] and TargetScanHuman [25], and detected two potential binding sites of miR-107 in the 3’UTR of ER (Supplementary Figure S2A). To examine whether expression of ER is regulated by miR-107, we performed transfections of non-invasive, ER-positive MCF-7 cells. We transiently transfected MCF-7 cells with mimics and inhibitors of miR-107 and with an expression plasmid encoding for miR-107. The mimics are double-stranded RNA molecules which mimic the endogenous, mature miR-107, whereas the inhibitors are single-stranded, modified RNA molecules which after transfection, specifically binds to mimics and endogenous miR-107, and inhibits its function. The transfection with scramble miR served as a negative control. We confirmed the overexpression of miR-107 in the transfected cells by real-time PCR (data not shown). The data of the quantitative real-time PCR using gene-specific primers and Western blots using protein-specific antibodies specific for ER showed no effect of the mimics and inhibitors on the ER RNA and protein levels, respectively (Figure S2B and S2C). These findings suggest that miR-107 is not involved in the repression of ER in MCF-7 cells.
MiR-130a affects cell proliferation in MCF-7 cells
The effects of miR-16, miR-107 and miR-130a were also investigated on apoptosis and proliferation of MCF-7 and MDA-MB-231 cells. We transiently transfected these cells with mimics and inhibitors of miR-16, miR-107 and miR-130a. To induce apoptosis, the transfected cells were treated with topoisomerase I inhibitor camptothecin. FACS analyses did not show any effect on camptothecin mediated apoptosis by overexpression or inhibition of these miRs in the cell lines (Supplementary Figure S3).
Additionally, the effects of miR-16, miR-107 and miR-130a on cell proliferation were evaluated by a MTT assay. However, we only observed an effect on cell proliferation by miR-130a in MCF-7 cells, but not by miR-16 and miR-107 and in MDA-MB-231 cells. As shown in Figure 6, transfection of miR-130a inhibitor reduced the cell proliferation in non-invasive MCF-7 cells compared with analogous basal cells (p = 0.0001), whereas surprisingly, administration of miR-130a mimic had no effect on cell proliferation in this cell line. This could be explained that the endogenous expression level of miR-130a is too high in MCF-7 cells (albeit much lower than in MDA-MB-231), so that exogenously introduced miR-130a is not effective. Even using different concentrations of mimics and inhibitors for transfection of MCF-7 and MDA-MB-231 cells similar results were observed (Supplementary Figure S4).
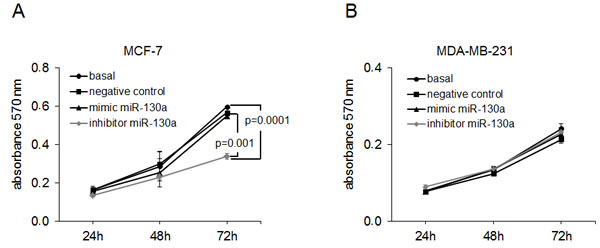
Figure 6: MiR-130a affects proliferation of MCF-7 cells. MCF-7 A. and MDA-MB-231 B. cells were transfected with mimic or inhibitor of miR-130a and treated with MTT. Transfection with scramble miR served as a negative control. Cell proliferation after overexpression and inhibition of miR-130a for 24 h, 48 h and 72 h. Inhibition of endogenous miR-130a reduced cell proliferation in MCF-7 cells as compared with basal MCF-7 cells and the negative control. The significant p-value as determined by the one-way ANOVA test and standard deviations from triplicate experiments are indicated.
MiR-107 affects migration and invasion of MCF-7 and MDA-MB-231 cells
Our current findings show that low levels of miR-107 correlated with a positive ER (p = 0.035) and lymph node status (p = 0.036). To examine whether miR-107 has an inhibitory effect on cell migration and invasion, respective assays were carried out using non-invasive MCF-7 and invasive MDA-MB-231 cells. As shown in Figure 7A, the overexpression of miR-107 significantly reduced the ability of MCF-7 (p = 0.001) and MDA-MB-231 (p = 0.01) cells to migrate compared with the negative control. The inhibition of miR-107 could increase migration of MCF-7 (p = 0.002), but no significant impact of MDA-MB-231 cells could be observed. Administration of miR-107 mimic also reduced the invasiveness of MCF-7 (p = 0.003) and MDA-MB-231 (p = 0.01) cells (Figure 7B). Transfection of miR-107 inhibitor stimulated the invasion of MCF-7 (p = 0.02) and MDA-MB-231 (p = 0.0001) cells through matrigel-coated transwell membranes. These findings show that alterations of the miR-107 level could change the migration and invasion behavior of cells. Therefore, miR-107 could play an important role in tumor progression.
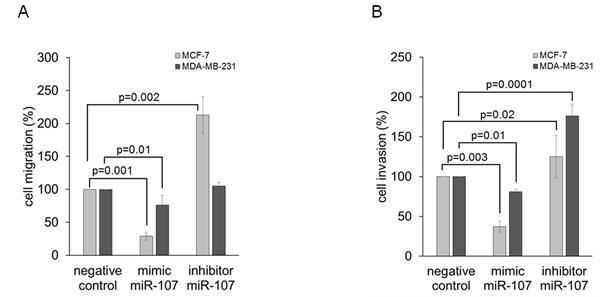
Figure 7: MiR-107 affects migration and invasion of MCF-7 and MDA-MB-231 cells. Cell migration A. and invasion B. of MCF-7 and MDA-MB-231 cells were analyzed by transwell assays. Cells were transfected with mimic or inhibitor of miR-107 prior to serum starvation for 24 h. Cells were added to the upper transwell chamber that was uncoated for the migration assay and BME-coated for the invasion assay, and allowed to migrate or invade for 24 h. Migrated or invaded cells were stained with Calcein-AM, and fluorescence (excitation: 485 nm, emission: 535 nm) was measured. Overexpression of miR-107 decreased migration and invasion in MCF-7 and MDA-MB-231 cells as compared with the negative control (transfection with scramble miR). Inhibition of endogenous miR-107 increased invasion in both cell lines, and migration in MCF-7 cells but not in MDA-MB-231 cells. The significant p-values as determined by the one-way ANOVA test and standard deviations from triplicate experiments are indicated.
Discussion
Based on our microarray data and their particular characteristics, we quantified six miRs (miR-16, miR-27a, miR-107, miR-130a, miR-132 and miR-146a) in postoperative plasma of breast cancer patients before and after chemotherapy and healthy women. As far as we know, only miR-16 and miR-146a of our panel of selected miRs have been quantified in the plasma or serum of breast cancer patients by other studies. The quantification of circulating miR-16 was often used as a normalization control [10]. The levels of miR-146a were measured in the plasma of Indian breast cancer patients and were significantly higher than in healthy women, indicating the diagnostic potential of this miR in breast cancer [26].
In our study, the plasma levels of miR-27a and miR-132 were increased in breast cancer patients before chemotherapy and decreased to normal (wild type) levels after chemotherapy, whereas the increased levels of miR-16 were not affected by chemotherapy. No correlation of the plasma levels of these miRs with different patient treatment arms could be observed. The levels of miR-16, miR-107, miR-130a and miR-146a were significantly higher in lymph-node negative patients than in their counterparts. Moreover, the concentrations of miR-107 were significantly increased in ER-negative patients compared with ER-positive patients, suggesting a suppressive effect on ER expression in vivo. However, our experimental studies on MCF-7 cancer cells showed that miR-107 could not downregulate ER protein expression in vitro. Finally, the plasma levels of miR-130a and miR-146a correlated with the HER2 status and were significantly increased in HER2-positive patients. Inhibition of miR-130a reduced the cell proliferation in non-invasive MCF-7 cells. Overexpression of miR-107 decreased migration and invasion of MCF-7 and MDA-MB-231 cells, while inhibition of miR-107 stimulated invasion of MCF-7 and MDA-MB-231 cells and migration of MCF-7 cells.
Discrepant data on miR-16 exist in numerous publications that describe this miR as oncogene, tumor suppressor gene or reference miR [27-29]. MiR-16 is most frequently used as endogenous control for data normalization, because this miR is highly expressed in plasma or serum, and has been described as being relatively invariant across diverse blood samples [30]. On the other hand, miR-16 expression has also been reported to be a potential therapeutic target and clinical biomarker of bone metastasis, because it is elevated in osteoclast differentiation and bone metastasis [20]. In our study, the levels of miR-16 were significantly increased in lymph node-negative patients and decreased to normal levels in patients with lymph node metastases. These findings point to a wavelike transcription of miR-16 during tumor development and progression, but also to a varying expression pattern of miR-16 in different tumors. However, to interpret more exactly the different transcript levels of miR-16, large patient populations should be investigated and most importantly, a standardized reference for normalization of the data should be established. To better comprehend the different features of miR-16, we looked for their mRNA targets, and found that miR-16 controls the intrinsic apoptosis pathway in breast cancer cell lines. Overexpression of miR-16 downregulated expression of the apoptosis inhibitor Bcl-2 at the protein level in MCF-7 cells, whereas suppression of miR-16 increased expression of Bcl-2 in MDA-MB-231 cells [31]. However, in our present study we could not observe such an apoptotic effect in MCF-7 and MDA-MB-231 cells. It is difficult to explain these discrepant data that could be rather caused by different experimental procedures. Moreover, we detected that in contrast to the other miRs of our panel, chemotherapy did not affect the increased levels of miR-16 in postoperative plasma of the breast cancer patients. This could be explained by inflammatory processes which release increasingly miR-16 into the blood circulation.
In our study, the plasma levels of circulating miR-107 were significantly higher in lymph node-negative and ER-negative tumors than in their respective positive counterparts. Though, the association of increased levels of miR-107 with ER-negative tumor is not caused by a downregulation of ER by miR-107, as shown by our in vitro data on MCF-7 cells. Nevertheless, the detection of decreased amounts of miR-107 in our subpopulation of ER-positive patients is supported by a previous study. This study reported an inhibitory effect of estrogen on the intracellular levels of miR-107 and its down-regulation in estrogen treated MCF-7 cells [16]. These and our present results point to that estrogen may negatively regulate the gene expression of miR-107. Moreover, we found that inhibition of miR-107 may stimulate migration of MCF-7 cells and invasion of MCF-7 and MDA-MB-231 cells. Overexpression of miR-107 inhibits the migratory and invasive ability of both cell lines. Our findings are supported by investigations in other tumor types such as glioma or cervical cancer. Enhanced miR-107 expression significantly inhibited invasion of glioma stem cells and reduced matrix metalloproteinase-12 expression [32]. It was shown for cervical cancer cells that miR-107 directly targeted MCL1 and activated ATR/Chk1 pathway to inhibit proliferation, migration and invasiveness of cervical cancer cells. Overexpression of miR-107 resulted in a significant reduction of migratory and invasive potential of the cells. Inhibition of miR-107 led to significant increase in invasive potential of HeLa cells [33]. As far as we know, only one study has investigated the role of miR-107 in breast cancer cells. It was reported that miR-107 negatively regulated the expression of CDK8, that is involved in the regulation of cell cycle and cell growth, and inhibited the proliferation and migration of MDA-MB-231 cell line [34]. Our results additionally demonstrate the potential role of miR-107 in migration and invasion of MCF-7 cells by overexpression and inhibition of this miR. These data may explain the high miR-107 transcript levels detected in patient with negative lymph node stage.
Moreover, we detected that increased plasma levels of miR-130a and miR-146a correlated with lymph node-negative and HER2-positive status of the patients. In contrast to our findings showing that miR-130a affects cell proliferation, but has no apoptotic effect, miR-130a was described as an inhibitor of apoptosis. Its expression may be directly regulated by the oncogene product c-Myc. Based on the frequent amplification of c-Myc in human cancers, high levels of c-Myc protein may provoke high expression of miR-130a, which may prevent apoptosis of cancer cells [17]. MiR-146a binds to the 3’UTR of BRCA1 and down-regulates its expression. This down-regulation was accompanied by an increased proliferation and a reduced homologous recombination rate, two processes controlled by BRCA1 [21]. Transfection of miR-146a into MDA-MB-231 inhibited invasion and migration in vitro and breast cancer metastasis [19]. Further work is needed to better understand the complex role of miR-130a and miR-146a.
In our panel of miRs, chemotherapy had an effect on the plasma levels of miR-27a and miR-132. The elevated amounts before chemotherapy decreased to normal levels after chemotherapy. As far as we know, there are no analyses on cell-free miR-27a and miR-132 in the blood circulation of breast cancer patients. Functional analyses showed that miR-27a promotes angiogenesis by mediating endothelial differentiation [14], and its expression in primary tumor tissues was associated with poor overall survival of breast cancer patients [15]. Conversely, the expression of miR-132 was found to be significantly deregulated in ductal carcinoma in situ (DCIS) [18].
In conclusion, clinically relevant, quantitative changes in the transcript levels of miRs can be detected in the blood circulation of cancer patients. Their screening in plasma of patients with invasive breast cancer may provide information on the aberrant signaling pathway that could be blocked by the chosen targeted therapy. The clinical use of these “liquid biopsies” could, therefore, contribute to the understanding of the molecular mechanisms underlying breast cancer development and progression. Our findings demonstrate the modulations of circulating miR-16, miR-27a, miR-107, miR-130a, miR-132 and miR-146a in the plasma of patients with invasive breast cancer and their association with specific breast cancer subtypes and biological behavior. Prospective studies on larger cohorts of patients are required to substantiate their diagnostic role.
Materials and methods
Study populations
Within a multicenter study (SUCCESS), which includes 251 German centers, postoperative blood plasma samples were collected from patients with invasive breast cancer. Plasma samples of 111 eligible patients participating at this study were analyzed before (July 2008 to May 2011) and after chemotherapy (November 2008 to September 2011, Figure 1). In addition, plasma samples were collected from 46 age-matched healthy women with no history of cancer and in good health based on self-report. All patients and healthy controls gave their informed consent. The study was approved by the ethic committee of the Ludwig-Maximilians University Munich and conducted in accordance with the Declaration of Helsinki.
The tumor stage at primary diagnosis of the patients was classified according to the revised American Joint Committee on Cancer tumor-node-metastasis classification. Histopathologic grading of the primary tumors was conducted according to the Bloom-Richardson system. For the diagnosis of lymph node metastasis, single embedded lymph nodes were screened up to 3 levels. External beam radiation therapy was administered in all patients treated with breast conservation. Chest wall irradiation following mastectomy was conducted in patients with more than 3 involved lymph nodes or T3 and T4 stage tumors. The adjuvant therapy included 3 cycles of 5-Fluorouracil 500 mg/m² i.v. body surface area and Epirubicin 100 mg/m² i.v. and Cyclophosphamide 500 mg/m² i.v., (FEC100), each administered on day 1, repeated on day 22, subsequently followed by 3 cycles of Docetaxel 75 mg/m² body surface area i.v., and Gemcitabine 1000 mg/m² i.v. (30 min infusion), administered on day 1, followed by Gemcitabine 1000 mg/m² i.v. (30 min infusion) on day 8, repeated on day 22. After chemotherapy, women with hormone receptor- positive disease received endocrine treatment (tamoxifen and arimidex). As determined by immunohistochemistry (IHC), a cut-off level of more than 10% was used for positive hormone receptor status. For the expression of HER2, the DAKO score from 0 to 3b was determined by IHC (+++) and FISH (+) analyses, according to the recommendations of American Society of Clinical Oncology. Women with HER2-positive disease received a therapy with trastuzumab.
MicroRNA profiling
MicroRNA profiling was performed with plasma samples derived from 20 breast cancer patients and 10 healthy women. Small RNA was isolated from 600 µL plasma using NucleoSpin miRNA Plasma Kit (Macherey-Nagel, Düren, Germany) and according to the manufacturer’s protocol. Blood-based microarray profiling was performed with SurePrintG3 Human miRNA (8x60K) microarrays (Agilent Technologies; Febit Biomed GmbH, Heidelberg, Germany) containing 1300 human and viral miRs derived from Sanger miRBase release 16.0 (April 2011). Samples were labeled with pCp-Cy3 using T4 ligase and hybridized on 8x60K format Agilent human miRNA array. Data were acquired using Agilent Feature Extraction software version 10.7.3.1.
Extraction of small RNA and conversion into cDNA
Small RNA was extracted from 300 µL plasma using the NucleoSpin miRNA Plasma Kit (Macherey-Nagel) and according to the manufacturer’s instructions. For extraction efficiency, 20 fmol of synthetic non-human cel-miR-39 was added as an exogenous spike in control. The RNA was quantified on a NanoDrop ND-1000 Spectrophotometer (Thermo Scientific, Wilmington, Delaware, USA) and stored at -80°C until they were reverse transcribed into cDNA.
Reverse transcription was performed by the TaqMan MicroRNA Reverse Transcription Kit (Life Technologies, New York, USA). The 10 μL-reverse transcription reaction contained 0.1 μL 100 mM dNTPs, 0.66 μL MultiScribe Reverse Transcriptase (50 U/μL), 1 μL 10× Reverse Transcription Buffer, 0.13 μL RNase Inhibitor (20 U/μL), 2 µL 5x TaqMan RT Primer, nuclease-free water and 2 μL RNA. The reaction was carried out at 16°C for 30 min., 42°C for 30 min. and 85°C for 5 min. on a MJ Research PTC-200 Peltier Thermal Cycler (Global Medical Instrumentation, Ramsey, Minnesota, USA). The cDNA samples were stored at -20°C until further use.
Quantitative real-time PCR of miR-16, miR-27a, miR-107, miR-130a, miR-132 and miR-146a
For quantitative real-time PCR, the miR-specific TaqMan MicroRNA Assays (Life Technologies) for miR-1207 (reference miR), miR-16, miR-27a, miR-107, miR-130a, miR-132 and miR-146a were used. In a 10 μL-reaction, 1 μL cDNA were mixed with 5 μL TaqMan Gene Expression Master Mix and 0.5 μL 20x miR-specific TaqMan MicroRNA Assay on a twin-tec real-time PCR plate (Eppendorf, Hamburg, Germany). The quantitative real-time PCR reaction was performed at 95°C for 10 min. and for 40 cycles at 95°C for 15 s and 60°C for 60 s on a Mastercycler Realplex (Eppendorf).
As there is no consensus concerning the normalization of circulating miRs, we chose miR-1207 as reference to normalize our miR data, because this miR showed the smallest coefficient variation of the populations as measured by the blood-based microarray (0.262). In the plasma samples of breast cancer patients, we calculated a mean value of 29.67 (SD = 1.75) before therapy and of 29.42 (SD = 1.79) after therapy. The mean value in healthy controls was 30.51 (SD = 1.46). These findings show the relative constant expression of miR-1207 in the different cohorts. The obtained data of the miR expression levels were calculated and evaluated by the ΔCt method as follows: ΔCt = mean value Ct (reference miR-1207) - mean value Ct (miR of interest). The relative expression of the miR of interest corresponded to the 2^(ΔCt) value.
To avoid quantifying our miR panel in hemolytic plasma samples that may influence our results, we excluded such plasma samples that color changed in red or dark red. In addition, we also examined plasma samples for contamination by hemolysis using miR-451 and miR-23a. As described by Blondel et al., if the ratio between these miRs is higher than 5, it is an indicator of possible erythrocyte miR contamination, and a ΔCt of 7–8 or more indicates a high risk of hemolysis affecting the data obtained [35]. Our data for our measurements of the ratio between miR-451 and miR-23a showed a mean value of 3.19 with a standard deviation of 1.27, a median value of 3.73 and a range between 0.32 and 5.26, indicating that the plasma samples were not contaminated.
Transient transfection of miR-16, miR-107 and miR-130a
The breast adenocarcinoma cell lines MCF-7 and MDA-MB-231 (ATCC) were cultured in DMEM (Invitrogen, Karlsruhe, Germany) supplemented with 10% FCS (fetal calf serum; PAA Laboratories, Cölbe, Germany) and 2 mM L-glutamin (Life Technologies) under standard conditions (37°C, 10% CO2, humidified atmosphere). Using Multiplex Cell Authentication (Multiplexion, Heidelberg, Germany) they were authenticated within six months of use. 3*105 of cells were seeded on 6-well plates (NUNC, Roskilde, Denmark) and transfected with 1 µg of an expression plasmid encoding for miR-107 or the double-stranded miScript miRNA mimics miR-16, miR-107 and miR-130a at final concentration of 10 nM (Qiagen, Hilden, Germany) or the analogous single-stranded miScript miRNA inhibitors at final concentration of 50 nM (Qiagen) with 2 µL X-tremeGENE HP DNA Transfection Reagent (Roche Diagnostics, Mannheim, Germany). As a negative control served the transfection with miScript scramble at final concentration of 50 nM (Qiagen). For titration experiments the cells were transfected with miR-16, miR-107 and miR-130a mimics at final concentration of 5, 10, 20, or 30 nM and with inhibitors at final concentration of 25, 50, 100, or 150 nM.
The expression (pcDNA3.1) plasmid encoding for miR-107 was constructed by annealing the DNA sequences of 5’-TGCAGAATTCCTCTCTGCTTTCAG CTTCTTTACAGTGTTGCCTTGTGGCATGGAGTTCA -3’ and 5’-AGTATCTCGAGTCTGTGCTTTGATAGCCC TGTACAATGCTGCTTGAACTCCATGCCACA-3’. The 5’ overhanging ends were filled in by the Klenow fragment DNA polymerase (Thermo Scientific). The double-stranded DNA was cut by the restriction enzymes EcoRI and XhoI (New England Biolabs, Frankfurt, Germany) and cloned into the multiple cloning site of the pcDNA 3.1 vector (Life Technologies). The cloned expression plasmid was verified by DNA sequencing.
Quantitative real-time PCR and western blot of ER expression
After 48 hour incubation of untreated and transfected MCF-7 and MDA-MB-231 cells, total RNA and protein were extracted using peqGOLD TriFast (Peqlab, Erlangen, Germany) according to the manufacturer`s instructions.
To determine mRNA expression of ER, a total of 200 ng RNA from basal and transfected cells was reverse transcribed using the First strand cDNA synthesis kit (Thermo Scientific). The mRNA expression levels were subsequently quantified by real-time PCR using the Maxima SYBR Green/ROX qPCR Master Mix (Thermo Scientific) and the following primers for ER (forward: 5’-GCATTCTACAGGCCAAATTCA-3’ and reverse: 5’-TCCTTGGCAGATTCCATAGC-3’). GAPDH (forward: 5’-CCTGCACCACCAACTGCTTAG-3’ and reverse: 5’-TGGCATGGACTGTGGTCATG-3’) served as reference gene. Protein levels of ER in basal and transfected MCF-7 cells were investigated by Western blot analysis. Thirty µg of cell lysates were electrophoretically separated and blotted onto a PVDF membrane (Millipore, Billerica, USA) which was subsequently incubated with antibodies specific for ER (Thermo Scientific) and GAPDH (Santa Cruz, Heidelberg, Germany) overnight. Detection of the proteins was carried out using peroxidase-conjugated secondary antibodies (Dako, Glostrup, Denmark) and the chemiluminescence ECL detection solution (Sigma-Aldrich St.Louis, Missouri, USA).
Apoptosis assay and flow cytometry
Transfected (24 hour transfection) MCF-7 cells (on a 6-well plate) were treated with 6 µM of the topoisomerase I inhibitor camptothecin (Biovision, Milpitas, USA) for 4 hours. Twenty-four hours after induction of the camptothecin-mediated apoptosis, the cells were stained for flow cytometry using the FITC Annexin V Apoptosis Detection Kit (BD Biosciences, San Jose, California, USA) according to the manufacturer`s instructions. The cells were then analyzed on a FACS CantoII flow cytometer (BD Biosciences).
MTT assay
To measure the cell proliferation, non-transfected and transfected cells were incubated with 20 µL 5 mg/ml MTT (thiazolyl blue tetrazolium bromide, Sigma-Aldrich) in PBS on a 96-well plate at 37°C for 3 hours. Following incubation, the cells were lysed with lysis buffer (4 mM HCl, 0.1% NP40 in isopropanol). A microplate reader (Tecan, Männedorf, Switzerland) was used to measure the OD values at 540 nm. Each experimental group contained three replicate wells, and the experiment was repeated three times.
Cell migration and invasion assay
Cell migration and invasion were measured using 8-µm pore uncoated or BME (basement membrane extract)-coated transwell inserts, respectively (Trevigen, City of Gaithersburg, Maryland, USA). Briefly, 8 h after transfection with 20 nM mimic, 100 nM inhibitor of miR-107, MCF-7 and MDA-MB-231 cells were grown in starvation medium (DMEM with 0.5% FCS and 2 mM L-glutamine) for 24 h prior to their detachment. Cells were resuspended in 400 µL starvation medium, and 100,000 cells per well were added to the top chamber. Medium containing 10% FCS was added to the bottom chamber, and cells were allowed to migrate and invade for 24 h at 37°C. Inserts and wells were washed with 1x washing buffer (Trevigen), and migrated or invaded cells were incubated in 500 µL dissociation buffer containing 0.83 µg Calcein-AM (Trevigen) for 1 h. Fluorescence (excitation: 485 nm, emission: 535 nm) was measured by the LB 940 Mithras microplate reader (Berthold Technologies, Bad Wildbach, Germany). In order to refer relative fluorescence units (RFU) to cell number, separate standard curves for each cell line were performed. On a 24-well plate, a serial dilution of 50,000, 25,000, 12,500, 6,250, 3,125 cells per well was carried out in triplicate. Fluorescence was measured after 1 hour incubation using 500 µL dissociation buffer containing 0.83 µg Calcein-AM (Trevigen). Mean values were plotted for standard curve and the line equation of the trend line was used to calculate the number of migrated or invaded cells.
Statistical analysis
The statistical analyses were performed using the SPSS software package, version 18.0 (SPSS Inc. Chicago, IL). Because of the skewed distribution of the miRs concentrations, differences in group levels for nonparametric comparisons were bivariately assessed by univariate analyses of the Mann–Whitney-U test of two independent variables and Wilcoxon test of two dependent variables. Diagnostic power of the miRs was analyzed by receiver operating characteristic (ROC) curves. Areas under the curves (AUC) were calculated, assuming nonparametric distribution. Additionally, univariate binary logistical regression was carried out for analyzing combined miRs with the clinicopatholgical data or combined hormone receptor statuses with the variables. For cell lines, the one-way ANOVA with Dunnett and Tukey test was used for all pairwise comparisons that correct for experiment-wise error rate. Missing data were handled by pairwise deletion. A p-value <0.05 was considered as statistically significant. All p-values are two-sided. Due to the explorative nature of the study no formal adjustment for multiple testing was performed.
Acknowledgements
We thank Mrs. Bettina Steinbach for their excellent technical assistance. The Core Unit FACS-Sorting of the University Hospital Hamburg-Eppendorf for the helpful assistance to carry out flow cytometry and PD Dr. Zahner from the Center for Internal Medicine of the University Hospital Hamburg-Eppendorf for providing us the LB 940 Mithras microplate reader. We are also grateful to ERC (European Research Council) for supporting this study.
Conflicts of Interest
No potential conflicts of interest were disclosed.
Grant Support
This present study was supported by ERC (European Research Council) Advanced Investigator Grant (ERC-2010 AsG 20100317 DISSECT).
References
1. Strehl JD, Wachter DL, Fasching PA, Beckmann MW and Hartmann A. Invasive Breast Cancer: Recognition of Molecular Subtypes. Breast Care (Basel). 2011; 6:258-264.
2. Pritchard KI. Endocrine therapy: is the first generation of targeted drugs the last? Journal of internal medicine. 2013; 274:144-152.
3. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009; 136:215-233.
4. Cheng AM, Byrom MW, Shelton J and Ford LP. Antisense inhibition of human miRNAs and indications for an involvement of miRNA in cell growth and apoptosis. Nucleic Acids Res. 2005; 33:1290-1297.
5. Heneghan HM, Miller N, Lowery AJ, Sweeney KJ and Kerin MJ. MicroRNAs as Novel Biomarkers for Breast Cancer. J Oncol. 2009; 2009:950201.
6. Perilli L, Vicentini C, Agostini M, Pizzini S, Pizzi M, D’Angelo E, Bortoluzzi S, Mandruzzato S, Mammano E, Rugge M, Nitti D, Scarpa A, Fassan M and Zanovello P. Circulating miR-182 is a biomarker of colorectal adenocarcinoma progression. Oncotarget. 2014; 5:6611-6619.
7. Shen J, Hu Q, Schrauder M, Yan L, Wang D, Medico L, Guo Y, Yao S, Zhu Q, Liu B, Qin M, Beckmann MW, Fasching PA, Strick R, Johnson CS, Ambrosone CB, et al. Circulating miR-148b and miR-133a as biomarkers for breast cancer detection. Oncotarget. 2014; 5:5284-5294.
8. Ye SB, Li ZL, Luo DH, Huang BJ, Chen YS, Zhang XS, Cui J, Zeng YX and Li J. Tumor-derived exosomes promote tumor progression and T-cell dysfunction through the regulation of enriched exosomal microRNAs in human nasopharyngeal carcinoma. Oncotarget. 2014; 5:5439-5452.
9. Esquela-Kerscher A and Slack FJ. Oncomirs - microRNAs with a role in cancer. Nat Rev Cancer. 2006; 6(4):259-269.
10. Schwarzenbach H, Hoon DS and Pantel K. Cell-free nucleic acids as biomarkers in cancer patients. Nat Rev Cancer. 2011; 11:426-437.
11. Simpson RJ, Lim JW, Moritz RL and Mathivanan S. Exosomes: proteomic insights and diagnostic potential. Expert Rev Proteomics. 2009; 6:267-283.
12. Cortez MA and Calin GA. MicroRNA identification in plasma and serum: a new tool to diagnose and monitor diseases. Expert Opin Biol Ther. 2009; 9:703-711.
13. Blenkiron C, Goldstein LD, Thorne NP, Spiteri I, Chin SF, Dunning MJ, Barbosa-Morais NL, Teschendorff AE, Green AR, Ellis IO, Tavare S, Caldas C and Miska EA. MicroRNA expression profiling of human breast cancer identifies new markers of tumor subtype. Genome biology. 2007; 8:R214.
14. Tang W, Yu F, Yao H, Cui X, Jiao Y, Lin L, Chen J, Yin D, Song E and Liu Q. miR-27a regulates endothelial differentiation of breast cancer stem like cells. Oncogene. 2013.
15. Tang W, Zhu J, Su S, Wu W, Liu Q, Su F and Yu F. MiR-27 as a prognostic marker for breast cancer progression and patient survival. PloS one. 2012; 7:e51702.
16. Cicatiello L, Mutarelli M, Grober OM, Paris O, Ferraro L, Ravo M, Tarallo R, Luo S, Schroth GP, Seifert M, Zinser C, Chiusano ML, Traini A, De Bortoli M and Weisz A. Estrogen receptor alpha controls a gene network in luminal-like breast cancer cells comprising multiple transcription factors and microRNAs. Am J Pathol. 2010; 176:2113-2130.
17. Yang F, Miao L, Mei Y and Wu M. Retinoic acid-induced HOXA5 expression is co-regulated by HuR and miR-130a. Cell Signal. 2013; 25:1476-1485.
18. Li S, Meng H, Zhou F, Zhai L, Zhang L, Gu F, Fan Y, Lang R, Fu L, Gu L and Qi L. MicroRNA-132 is frequently down-regulated in ductal carcinoma in situ (DCIS) of breast and acts as a tumor suppressor by inhibiting cell proliferation. Pathol Res Pract. 2013; 209:179-183.
19. Hurst DR, Edmonds MD, Scott GK, Benz CC, Vaidya KS and Welch DR. Breast cancer metastasis suppressor 1 up-regulates miR-146, which suppresses breast cancer metastasis. Cancer research. 2009; 69:1279-1283.
20. Ell B, Mercatali L, Ibrahim T, Campbell N, Schwarzenbach H, Pantel K, Amadori D and Kang Y. Tumor-Induced Osteoclast miRNA Changes as Regulators and Biomarkers of Osteolytic Bone Metastasis. Cancer Cell. 2013; 24:542-556.
21. Garcia AI, Buisson M, Bertrand P, Rimokh R, Rouleau E, Lopez BS, Lidereau R, Mikaelian I and Mazoyer S. Down-regulation of BRCA1 expression by miR-146a and miR-146b-5p in triple negative sporadic breast cancers. EMBO molecular medicine. 2011; 3:279-290.
22. Nagaraja GM, Othman M, Fox BP, Alsaber R, Pellegrino CM, Zeng Y, Khanna R, Tamburini P, Swaroop A and Kandpal RP. Gene expression signatures and biomarkers of noninvasive and invasive breast cancer cells: comprehensive profiles by representational difference analysis, microarrays and proteomics. Oncogene. 2006; 25:2328-2338.
23. Maragkakis M, Reczko M, Simossis VA, Alexiou P, Papadopoulos GL, Dalamagas T, Giannopoulos G, Goumas G, Koukis E, Kourtis K, Vergoulis T, Koziris N, Sellis T, Tsanakas P and Hatzigeorgiou AG. DIANA-microT web server: elucidating microRNA functions through target prediction. Nucleic Acids Res. 2009; 37(Web Server issue):W273-276.
24. Betel D, Wilson M, Gabow A, Marks DS and Sander C. The microRNA.org resource: targets and expression. Nucleic Acids Res. 2008; 36(Database issue):D149-153.
25. Lewis BP, Burge CB and Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell. 2005; 120:15-20.
26. Kumar S, Keerthana R, Pazhanimuthu A and Perumal P. Overexpression of circulating miRNA-21 and miRNA-146a in plasma samples of breast cancer patients. Indian journal of biochemistry & biophysics. 2013; 50:210-214.
27. Hu Z, Dong J, Wang LE, Ma H, Liu J, Zhao Y, Tang J, Chen X, Dai J, Wei Q, Zhang C and Shen H. Serum microRNA profiling and breast cancer risk: the use of miR-484/191 as endogenous controls. Carcinogenesis. 2012; 33:828-834.
28. Schwarzenbach H, Nishida N, Calin GA and Pantel K. Clinical relevance of circulating cell-free microRNAs in cancer. Nature reviews Clinical oncology. 2014.
29. Schwarzenbach H. Circulating nucleic acids as biomarkers in breast cancer. Breast cancer research : BCR. 2013; 15:211.
30. Kroh EM, Parkin RK, Mitchell PS and Tewari M. Analysis of circulating microRNA biomarkers in plasma and serum using quantitative reverse transcription-PCR (qRT-PCR). Methods. 2010; 50:298-301.
31. Zhang N, Wang X, Huo Q, Li X, Wang H, Schneider P, Hu G and Yang Q. The oncogene metadherin modulates the apoptotic pathway based on the tumor necrosis factor superfamily member TRAIL (Tumor Necrosis Factor-related Apoptosis-inducing Ligand) in breast cancer. The Journal of biological chemistry. 2013; 288:9396-9407.
32. Chen L, Chen XR, Chen FF, Liu Y, Li P, Zhang R, Yan K, Yi YJ, Xu ZM and Jiang XD. MicroRNA-107 inhibits U87 glioma stem cells growth and invasion. Cellular and molecular neurobiology. 2013; 33:651-657.
33. Zhou C, Li G, Zhou J, Han N, Liu Z and Yin J. miR-107 activates ATR/Chk1 pathway and suppress cervical cancer invasion by targeting MCL1. PloS one. 2014; 9:e111860.
34. Li XY, Luo QF, Wei CK, Li DF, Li J and Fang L. MiRNA-107 inhibits proliferation and migration by targeting CDK8 in breast cancer. International journal of clinical and experimental medicine. 2014; 7:32-40.
35. Blondal T, Jensby Nielsen S, Baker A, Andreasen D, Mouritzen P, Wrang Teilum M and Dahlsveen IK. Assessing sample and miRNA profile quality in serum and plasma or other biofluids. Methods. 2013; 59:S1-6.