
Introduction
Constitutive photomorphogenic 1 (COP1) acts as an E3 ubiquitin ligase and has a pivotal role in light signaling in plants, but its role in mammals is much more complex. Plant COP1 can repress light signaling by targeting transcription factors, such as HY5, HYH, for ubiquitination and degradation [1]. Mammalian COP1 is involved in various cellular functions, such as proliferation and survival, by facilitating the degradation of physiological substrates through ubiquitin-mediated protein degradation [2, 3]. COP1 itself is self-ubiquitinated, and this process is under the regulation of COP9 signalosome subunit 6 (CSN6) [4], a protein involved in MDM2-p53 signaling [5] and Cullin neddylation [6]. So far, mammalian E3 ubiquitin ligase COP1 substrates include c-Jun [7], ETV1 [8], p53 [9], (C/EBPalpha) [10-12], acetyl-CoA carboxylase [13], TORC2 [14], MTA1 [15], 14-3-3 σ [4, 16, 17] and FOXO1 [18], suggesting that COP1 is engaged in many biological activities [3]. COP1 is a RING finger protein with coiled-coil and WD 40 domains that associate with target proteins and participates in various biological functions [1]. However, its many substrates remain to be characterized. COP1 regulates p53 and 14-3-3 σ stability to regulate cell growth, but its role in cell cycle regulation is not fully characterized.
The tumor suppressor p27 governing CDK activity is critical for regulating the cell cycle transition from the G0/G1 to the S phase [19] and is a downstream target of oncogenic signals [20-26]. Levels of p27 are tightly regulated to control cell cycle progression [19, 27], and p27 expression levels are protected by the activity of many tumor suppressors [24, 28]. p27 levels are high during the G0 /G1 phase and get reduced when the cells enter into the S phase. p27 levels are mainly regulated through polyubiquitination [29], and they are downregulated in many types of cancer. However, this deregulation in cancers is not completely understood.
Here, we show that p27 levels are reduced in response to COP1 accumulation during cell cycle progression. We show that COP1 directly associates with p27 and functions as an E3 ligase of p27 to accelerate the ubiquitin-mediated degradation of p27 via its RING domain. Also, COP1-mediated p27 downregulation does not require the participation of known regulators of p27 stability: KPC1, Jab1, PirH2 or Skp2. Importantly, the deregulation of COP1-p27 axis manifests in cell proliferation, transformation, and tumorigenicity.
Results
COP1 directly interacts with p27 during cell cycle progression
COP1 and p27 coeluted as part of the high molecular weight complex in gel filtration studies (Figure 1A). On the basis of these findings, we hypothesized that COP1 and p27 have an interactive or regulatory relationship. Further, co-immunoprecipitation experiments showed endogenous interaction of the two proteins in cells (Figure 1B). To determine whether the interaction was regulated by the cell cycle, we collected cell lysates from synchronized cells at various time points after release from the thymidine-nocodazole block [30] (Figure 1C). COP1 protein was detected in every cell cycle phase, but levels were reduced when cells were cycling from G2/M to G1 (Figure 1C). In addition, p27 accumulated following the reduction of COP1 at G1. Co-immunoprecipitation of p27 and COP1 demonstrated that they interacted during the cell cycle (Figure 1C). These results suggest that COP1 may have an as-yet uncharacterized function in the cell cycle that involves interaction with p27.
Next, we mapped the structural regions of COP1 required for its interaction with p27. Results showed that p27 was bound to the N-terminus of COP1 (aa 1-334 containing RING motif) but not to the C-terminus (aa 401-473; Figure 1D). We also mapped the COP1 binding region on p27 in vitro. A binding assay using TNT products indicated that the C-terminus of p27 (aa 87-198) was responsible for binding COP1 (Figure 1E).
COP1 preferentially binds to target proteins with the VP motifs [31]. We analyzed the p27 peptide sequence and found a putative COP1 binding motif located in p27 (aa 87-198)(Figure 1F). We predicted that abolishing this binding site at aa 101-102 by mutating the VP motif to alanine (VP→AA) would interfere with p27-COP1 binding. Indeed, co-immunoprecipitation showed that the p27 (VP→AA) mutant lost its binding affinity for COP1 (Figure 1F). These results indicate that direct interaction between COP1 and p27 may be critical for cell cycle progression.
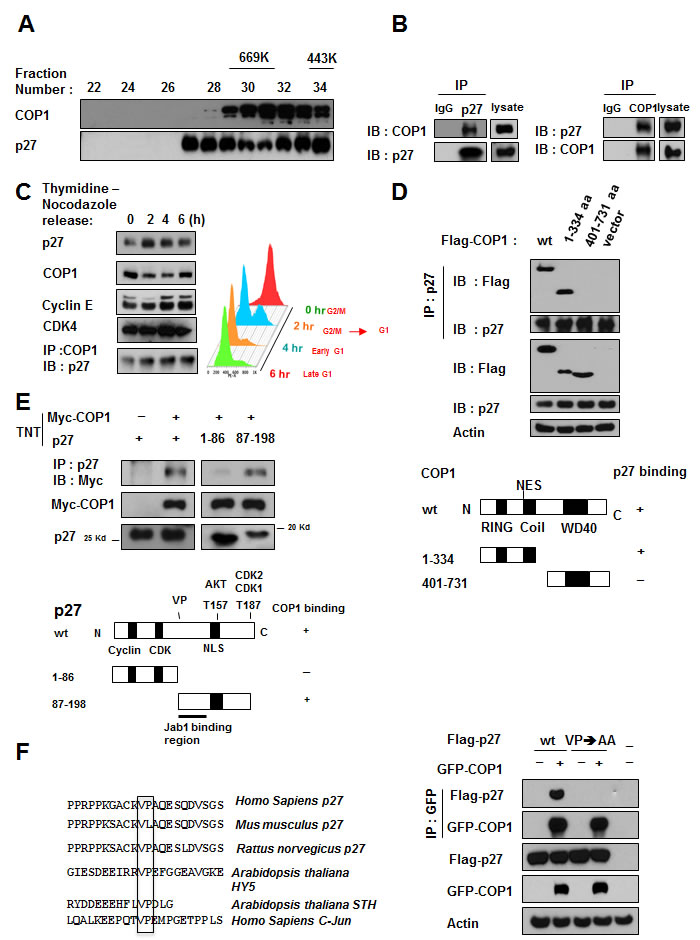
Figure 1: COP1 directly interacts with p27. (A) COP1 and p27 coeluted as part of the high molecular weight complex. Gel filtration and elution profiles analysis of COP1 and p27. The distributions of these proteins were analyzed by gel filtration chromatography (Superose 6). Immunoblots of the fractions for indicated proteins are shown in U2OS cells. Molecular size of eluted fraction is indicated above. (B) Endogenous interaction of COP1 and p27 was observed. Lysates of U2OS cells were prepared and equal amounts of cell lysates were analyzed by immunoprecipitation (IP) with either control mouse IgG or p27 and analyzed by immunoblotting (IB) with anti-COP1. Lysates were also analyzed by IP with the indicated antibodies and IB with anti-p27. (C) p27 and COP1 interacted during the cell cycle. U2OS cells were synchronized to the G2/M phase using treatment with thymidine-nocodazole. Lysates of synchronized cells were analyzed by IB with the indicated antibodies. COP1-p27 interaction at various phases of the cell cycle was detected by IP with the COP1 antibody followed by IB with anti-p27 (left). Cell samples at labeled time points after release of nocodazole were stained with propidium iodide and analyzed by FACS for DNA content. DNA content histograms are shown for the time points indicated (right). (D) p27 bound to the N-terminus of COP1 but not to the C-terminus. Wild-type (wt; aa 1-731), N-terminal (aa 1-334), or C-terminal (aa 401-731) Flag-COP1 was transfected into HeLa cells. Cells were treated with MG132, and cell lysates were analyzed by IP with anti-p27 and IB with anti-Flag. (E) COP1 bound to the C-terminus of p27. Myc-COP1 and PET-p27 were transcribed and translated in vitro (TNT). COP1 and p27 proteins were incubated overnight and analyzed by IP with anti-p27 followed by IB with anti-Myc. (F) The interaction of COP1 and p27 was mediated by the conserved VP sequence on p27. p27 has the VP motif for COP1 binding. Consensus COP1 binding motif, highlighted in sequences of p27 in human and other species’ DNA for comparison. HY5, STH, and c-Jun proteins are known COP1 binding proteins with VP motif. 293T cells were transfected with the indicated plasmids and treated with MG132. Cell lysates were analyzed by IP with anti-GFP and IB with anti-Flag.
COP1 negatively regulates p27 protein stability
We found that COP1 and p27 levels were inversely correlated in synchronized cells after synchronization release (Figure 2A). We showed that mRNA levels of p27 were not affected by COP1 overexpression in a real-time quantitative PCR analysis (Figure 2B) and that COP1-mediated p27 downregulation can be rescued by the proteasome inhibitor MG132 (Figure 2C). These results suggest that COP1 downregulates p27 at the posttranscriptional level.
We then found that overexpression of COP1 increased the endogenous ubiquitination level of p27 (Figure 2D), whereas COP1 knockdown reduced the endogenous ubiquitination level of p27 (Figure 2D). Similar results were obtained for ubiquitination levels of exogenous p27 (Figure 2E). Our data indicate that COP1 is involved in p27 ubiquitination.
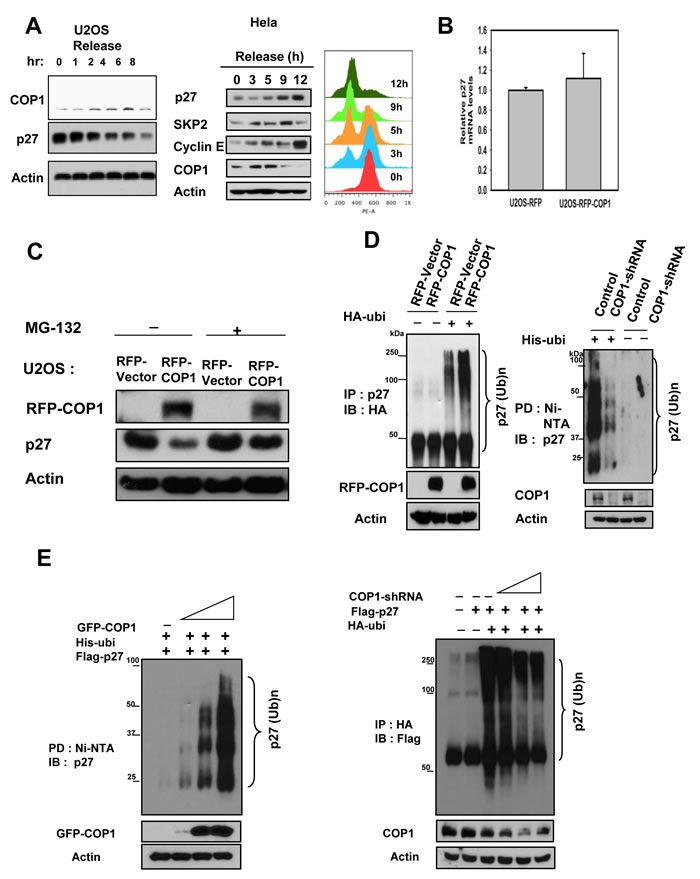
Figure 2: COP1 increases p27 polyubiquitination. (A) COP1 and p27 levels were inversely correlated in synchronized cells. U2OS cells were synchronized to the G1 phase using treatment with serum starvation, followed by refeeding with the serum. Lysates of synchronized cells after serum refeeding at labeled time points were analyzed by immunoblotting (IB) with the indicated antibodies. HeLa cells were synchronized to the G2/M phase using treatment with thymidine-nocodazole. Lysates of synchronized cells were analyzed by immunoblotting (IB) with the indicated antibodies. Cell samples at labeled time points after release of nocodazole were stained with propidium iodide and analyzed by FACS for DNA content. DNA content histograms are shown for the indicated time points. (B) mRNA levels of p27 were not affected by overexpression of COP1. Real-time quantitative PCR analysis of p27 in RFP-COP1–overexpressing U2OS stable transfectants and vector control transfectants showed no obvious difference in p27 mRNA levels.(C) MG132 rescued COP1-mediated p27 downregulation. RFP-COP1–overexpressing U2OS stable transfectants and vector control transfectants were treated with or without the proteasome inhibitor MG132 before lysates were collected. Lysates were analyzed by immunoblotting with the indicated antibodies. (D-E) COP1 regulated endogenous p27 polyubiquitination. Indicated transfected cells were treated with 5 µg/ml MG132 (Sigma) for 6 hours before they were harvested. Cells were lysed in guanidine–HCl containing buffer. The cell lysates then underwent pull down (PD) with nickel beads and analyzed by IB with anti-p27 or was immunoprecipitated (IP) with anti-HA and analyzed by IB with anti-Flag.
COP1-mediated p27 downregulation requires RING domain and physical binding
COP1-mediated p27 polyubiquitination was found to occur through the K48 link (Figure 3A). We also performed an in vitro ubiquitination assay to confirm that COP1 E3 ligase is capable of triggering polyubiquitination of p27 (Figure 3B). We then showed that the RING domain of COP1 is responsible for regulating p27 degradation because the COP1 RING mutant (C136S/C139S) was not able to downregulate steady-state expression of p27 (Figure 3C). Congruently, COP1 not only increased the turnover rate of p27 in the presence of the de novo protein synthesis inhibitor cycloheximide (Figure 3D), but also facilitated the ubiquitination of p27 (Figure 3E), whereas the COP1 RING mutant (C136S/C139S) failed to increase turnover of p27 and subsequent p27 ubiquitination (Figures 3D & 3E). In line with the binding requirement for COP1-mediated p27 downregulation, the p27 (VP→AA) mutant, which failed to bind COP1, showed slower turnover in the presence of COP1 compared with wt p27 (Figure 3F) and was resistant to COP1-mediated p27 ubiquitination (Figure 3G). In summary, these results demonstrated that COP1-mediated downregulation of p27 occurs through protein-protein interaction on the p27 VP motif and is triggered by COP1 E3 ligase for polyubiquitination through the RING motif.
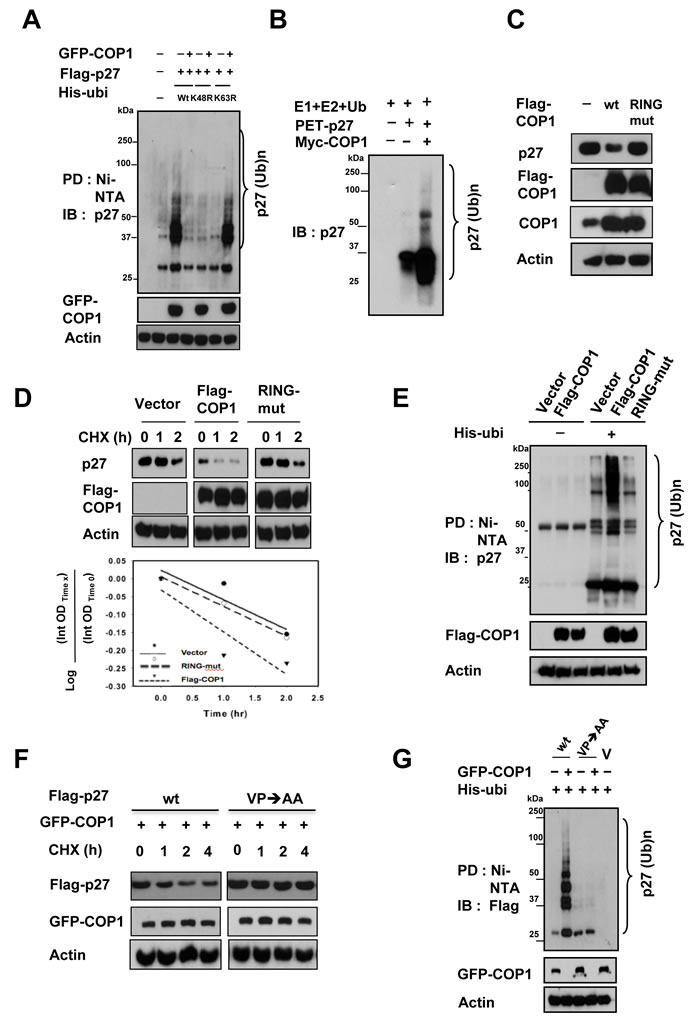
Figure 3: COP1-mediated p27 ubiquitination requires RING domain and physical binding. (A) COP1-mediated p27 polyubiquitination occurred through the K48 link. The K48R mutant of His-ubi disrupted COP1-mediated p27 polyubiquitination. 293T cells were co-transfected with the indicated plasmids. Cells were treated with 5 µg/ml MG132 (Sigma) for 6 hours before they were harvested. Cells were lysed in guanidine–HCl containing buffer. The cell lysates then underwent pull down (PD) with nickel beads and were analyzed by IB with anti-p27. Equal amounts of cell lysates were analyzed by IB with the indicated antibodies. (B) COP1 induced ubiquitination of p27 in an in vitro ubiquitination assay. Myc-COP1 and PET-p27 were prepared by in vitro transcription and translation using the TNT system. p27 was incubated with or without COP1 in the presence of E1, E2, His-Ubiquitin, and ATP as indicated. The ubiquitinated p27 was detected by IB with anti-p27 antibodies. (C) COP1 C136S/C139S (RING mutant) had no impact on p27 levels. HeLa cells were transfected with the indicated expression vectors. Equal amounts of protein from cell lysates were analyzed by IB with anti-p27, or anti-Flag-COP1. (D) COP1 C136S/C139S (RING mutant) had no effect on the turnover rate of p27. HeLa cells were transfected with the indicated expression vectors. Forty-eight hours after transfection, the cells were treated with cycloheximide (CHX; 100 µg/ml) for the indicated times. Cell lysates were analyzed by IB with anti-p27, anti-Flag, or anti-Actin. (E) Flag-COP1 C136S/C139S (RING mutant) compromised polyubiquitination of p27. HeLa cells were transfected with the indicated expression vectors. Cells were treated with MG132 for 6 hours before they were harvested and lysed in denaturing buffer (6M guanidine-HCl, 0.1M Na2HPO4/NaH2PO4, 10mM imidazole). The cell lysates were then incubated with nickel beads for 3 hours, washed, and analyzed by IB with anti-p27. (F) The p27 (VP→AA) mutant showed slower turnover in the presence of COP1 than wild-type (wt) p27. 293T cells were co-transfected with GFP-COP1 and either wt Flag-p27 or Flag-p27 (VP→AA). Cells were treated with CHX (100 µg/ml) for the indicated times. Cell lysates were analyzed by IB with the indicated antibodies. (G) The p27 (VP→AA) mutant was resistant to COP1-mediated p27 polyubiquitination. 293T cells were co-transfected with the indicated plasmids. Cells were treated with 5 µg/ml MG132 (Sigma) for 6 hours before harvesting. Cells were lysed in guanidine–HCl containing buffer. The cell lysates then underwent PD with nickel beads and were analyzed by IB with anti-Flag. Equal amounts of cell lysates were analyzed by IB with the indicated antibodies.
COP1-mediated p27 downregulation has impact on cell cycle progression
We further showed that the steady–state expression of p27 (VP→AA) mutant is not affected by the COP1 (Figure 4A). In contrast, COP1 reduced the steady-state expression of wt p27. As expected, p27 (VP→AA) mutant is still capable of causing G1 arrest (Figure 4B). Importantly, the biological significance of the resistance of p27 (VP→AA) to COP1-mediated degradation is that p27 (VP→AA) can diminish COP1-mediated cell cycle progression better than wt p27 (Figure 4B). COP1 knockdown leads to reduction of p27 ubiquitination (Figure 2), suggesting that COP1 knockdown will increase the level of p27. Because p27 causes cell cycle arrest, the biological significance of p27 elevation caused by COP1 knockdown was a delay of cell cycle progression as demonstrated by fluorescence-activated cell sorting (Figure 5). These data indicate that COP1-p27 link is involved in cell cycle regulation.
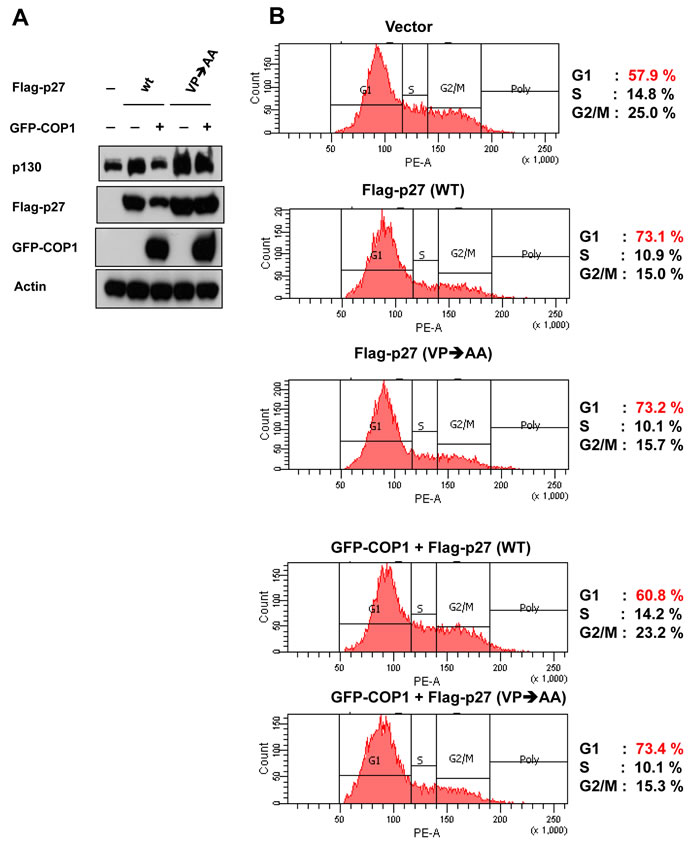
Figure 4: p27 (VP→AA) efficiently mitigates COP1-mediated cell cycle progression. (A) p27 (VP→AA) was resistant to COP1-meditated degradation. 293T cells were co-transfected with the indicated plasmids. Cell lysates were analyzed by immunoblotting with anti-p130, anti-Flag-p27, anti-GFP-COP1, or Actin antibodies. (B) p27 (VP→AA) diminished COP1-mediated cell cycle progression better than wt p27. 293T cells were co-transfected with the indicated plasmids as shown in A. Cells were analyzed for cell-cycle distribution using fluorescence-activated cell sorting.
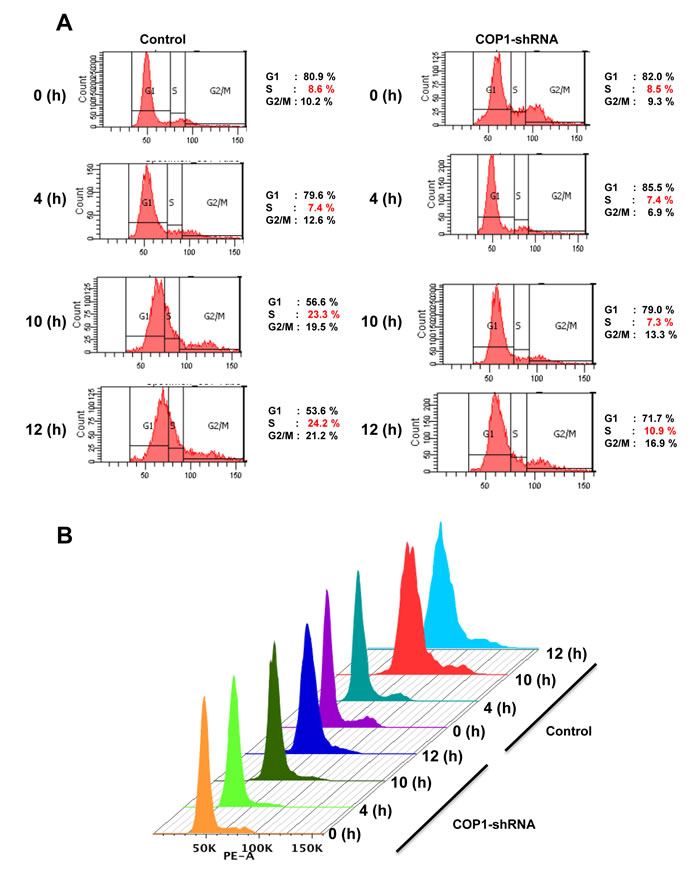
Figure 5: Cell cycle progression is delayed with COP1 deficiency. (A) COP1 knockdown led to a delay of cell cycle progression. MDA-MB231 cells infected with either lentiviral COP1 shRNA or control shRNA were synchronized in the G0/G1 phase after 72 hours of serum starvation before they were released into fresh medium containing serum. Cells were collected at the indicated time points. (B) DNA histograms from A are shown.
COP1-mediated p27 downregulation is independent of other known E3 ligases of p27
We showed that COP1 negatively regulated p27 expression, and we found that p27 levels were elevated when cells were treated with COP1-shRNA virus to perform COP1 knockdown (Figure 6A). Importantly, we also noted that expression levels of several known regulators involved in p27 degradation, including KPC1 [32], SKP2 [29], PirH2 [33], and Jab1 [34, 35], were not affected by the COP1 deficiency or overexpression in this process (Figure 6A). We noted that COP1-mediated p27 downregulation did not involve Akt-mediated phosphorylation (phosphorylation at T157), Cdk2-mediated phosphorylation (phosphorylation at T187 required for Skp2 binding) or Jab1 interaction (required Jab1-binding site on p27) (Figure 6B), as wt p27 and all these different mutants of p27 were downregulated by COP1. In assays using knockdown of KPC1, Jab1, PirH2, or SKP2, we found that COP1-mediated p27 downregulation did not require the participation of KPC1, Jab1, PirH2, or SKP2 (Figures 6C-6F). Together, these data indicate that COP1-mediated p27 downregulation is independent of these regulators.
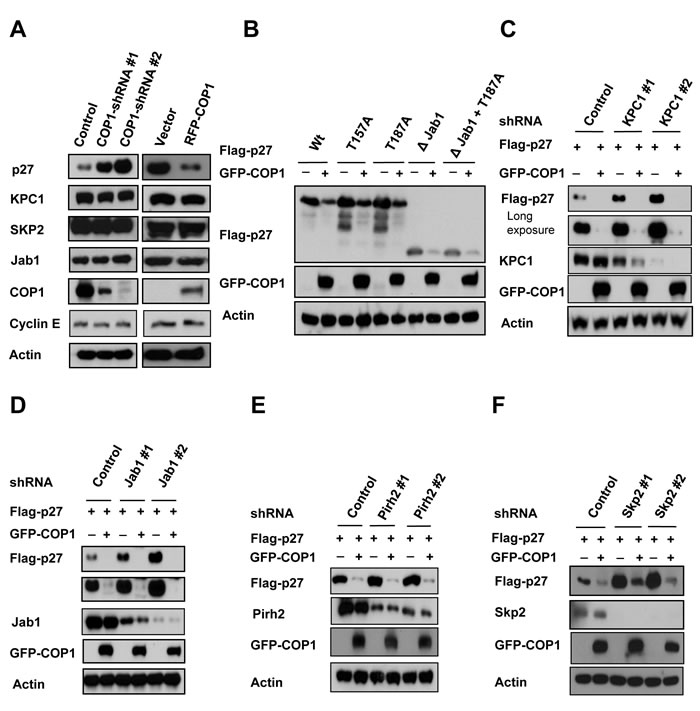
Figure 6: KPC1, Jab1, Pirh2, and SKP2 are not involved in COP1-mediated p27 downregulation. (A) Protein levels of KPC1, Jab1, Pirh2, and SKP2 were not changed when endogenous COP1 expression was inhibited with shRNA (left). Lysates of MDA-MB231 cells infected with either COP1-shRNA or control shRNA were analyzed by IB with the indicated antibodies. Lysates of COP1-overexpressing U2OS stable transfectants and vector control transfectants were analyzed by IB with the indicated antibodies (right). (B) 293T cells were co-transfected with the indicated plasmids. Lysates of cells transfected with indicated plasmids were analyzed by immunoblotting with the indicated antibodies. (C)-(F) 293T cells were co-transfected with the indicated plasmids. Lysates of cells infected with indicated shRNAs were analyzed by immunoblotting with the indicated antibodies.
COP1-p27 axis regulates cell proliferation and tumor growth
We further confirmed that COP1-p27 axis could affect cell proliferation. COP1-expression facilitates cell growth (Figure 7A). Since COP1 can mediate p27 inhibition, we sought to examine the growth effect of replenishing p27 in terms of cell proliferation, foci formation, and anchorage-independence in COP1-overexpressing cells. We found that p27 replenishment (through Adenoviral delivery) antagonized COP1-mediated cell proliferation, foci formation, and anchorage-independent growth (Figures 7A-7D). On the other hand, the COP1 knockdown cells have elevated p27 and thus a slower rate of cell proliferation, colony formation, and tumorigenicity (Figures 8A-8C) compared with control cells. Additional p27 knockdown in COP1 knockdown cells reverses COP1 shRNA-mediated reduction of cell proliferation, colony formation, and tumorigenicity (Figures 8A-8C). Significantly, COP1 is overexpressed in many types of cancer (Figure 8D), suggesting that tumors with COP1 overexpression may have growth advantage as shown in our cell and mouse model studies.
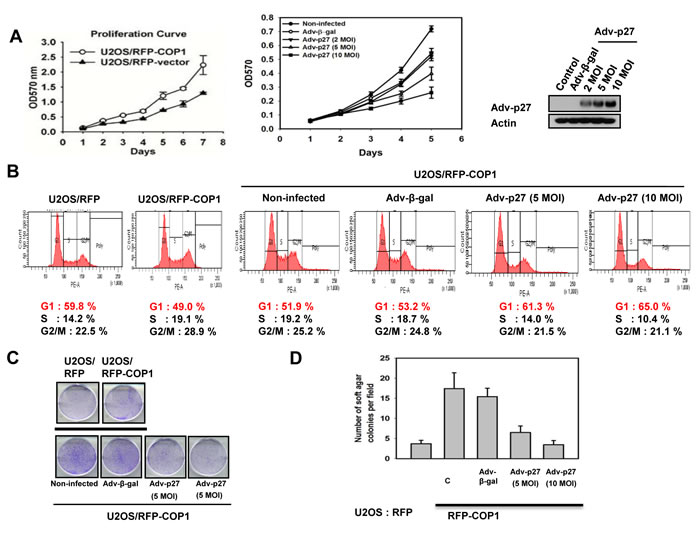
Figure 7: COP1-p27 axis in cell proliferation and transformation. (A) p27 inhibited COP1-mediated cell proliferation. RFP-COP1 overexpressing U2OS stable transfectants and vector control transfectants were estimated by MTT assay. Also, RFP-COP1 overexpressing cells were infected with Ad-β-gal or Ad-p27 and assayed by MTT assay. Error bars represent 95% confidence intervals. Expression of p27 was shown by Western blot. MOI (multiplicity of infection). (B) p27 increased G1 population in RFP-COP1 overexpressing cells. Cells were infected with Ad-β-gal or Ad-p27. Cell cycle distribution was determined using propidium iodide (PI). Cells were stained with PI and analyzed with a FACScalibur flow cytometer. (C) –(D) p27 replenishment antagonized COP1-mediated foci and soft agar colony formation. Indicated cells were infected with Ad-β-gal or Ad-p27 and analyzed for foci formation or soft agar colony formation. Error bars represent 95% confidence intervals.
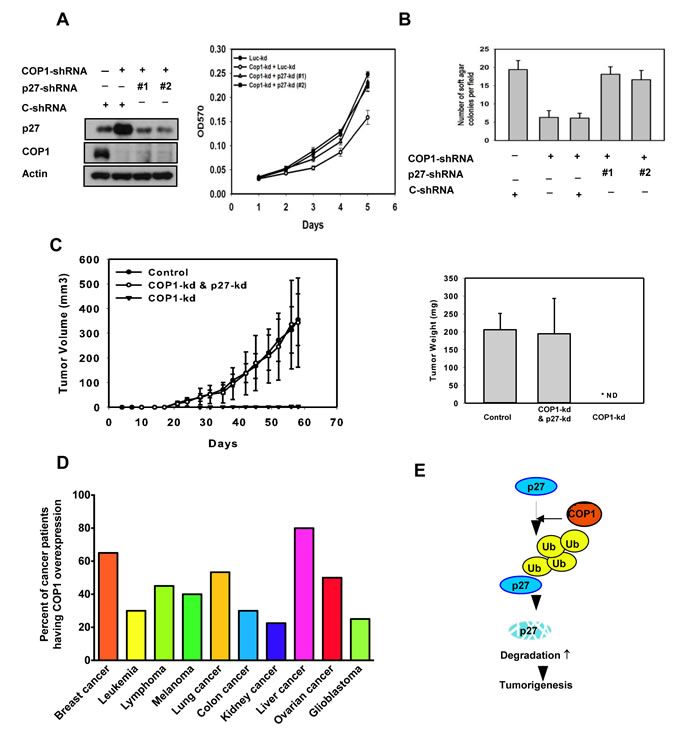
Figure 8: COP1-p27 axis in tumorigenesis. (A-B) COP1-knockdown reduced cell growth and soft agar colony formation. Indicated MDA-MB-231 cells with COP1 or p27 knockdown were analyzed for cell growth by MTT assay or assessed for transformation by soft agar colony formation. Error bars represent 95% confidence intervals. (C) COP1-knockdown suppressed tumorigenesis. COP1-knockdown MDA-MB-231 cells were further knocked down with shp27. Cells were harvested and s.c. injected into the flank of nude mice. Tumor volumes were monitored. Tumor growth curves are shown; error bars represent 95% confidence intervals. Tumor weights from each group were measured. Error bars represent 95% confidence intervals. (D) COP1 is frequently overexpressed in many common types of cancer. Human cancer patient datasets were obtained from Oncomine and Gene Expression Omnibus. Data were analyzed using Oncomine analysis tools and Nexus Expression 3.0. Patients with more than a 60% increase in COP1 mRNA levels compared with normal tissues were counted as “COP1 overexpression.” The number of patients analyzed: breast cancer (910), leukemia (127), lymphoma (94), melanoma (83), lung cancer (260), colon cancer (237), kidney cancer (114), liver cancer (197), ovarian cancer (50), and glioblastoma (84). (E) A model of the relationship between COP1 regulation, p27 stability, and tumor growth is shown.
Discussion
COP1 can suppress p53 activity [9] and thus acts as an oncoprotein, but COP1 knockout mouse model data suggest that COP1 may also function as a tumor suppressor via antagonizing oncogenic activity of c-Jun and ETV1 [2, 7, 8] in some tissues; therefore, its role in cancer requires further studies. Importantly, its many substrates remain to be characterized. Further, its role in cell cycle regulation is not clear. A better understanding of the COP1 regulatory complexity is critical for managing cancer therapy. In this study, we found that COP1 could reduce the steady-state expression of p27, a critical CDK inhibitor involved in cell cycle regulation, suggesting that a new layer of regulation on p27 exists in cell cycle progression. The fact that COP1 functions as an E3 ligase of p53 as well as p27, and causes their degradation, suggests an oncogenic role. The COP1 KO mouse model presents a surprising discovery about its tumor suppressor role. Our human cancer sample studies indicate that COP1 is highly elevated in many types of cancer, which is in conflict with tumor suppressive role, suggesting that COP1 knockout mouse cancer studies may have some discrepancies that currently cannot be explained.
The fact that COP1 participates in p27 ubiquitination/degradation is unprecedented as the ubiquitin ligase component F-box protein Skp2 is the major regulator for polyubiquitination of p27 and mediates its degradation [29, 36-39]. However, in the absence of Skp2, p27 is still degraded, suggesting that other E3 ubiquitin ligases may regulate p27 turnover [40]. PirH2, a RING containing protein [41], is the recently identified E3 ligases for p27 [40]. Nonetheless, we still characterized that COP1 is an E3 ligase regulating p27 without the involvement of PirH2, Jab1, KPC1 or Skp2. Further, we showed that COP1 enhances p27 ubiquitination, and that p27 (VP→AA) construct is resistant to COP1-mediated ubiquitination and subsequent degradation, providing important insight into the structure/functional relationship between COP1 and p27.
In our cancer model study, COP1 knockdown inhibited tumor growth in xenograft breast cancer model. Further p27 knockdown in these COP1 knockdown cells reverses COP1 shRNA-mediated reduction of tumorigenicity. Thus, these studies recapitulated the relationship between COP1 and p27 in vivo. We also found that COP1 overexpression is quite common in different types of cancer. p27 downregulation is frequently observed in human cancer samples [26]. Our findings demonstrate that COP1 overexpression can at least partially account for p27 downregulation in many cancers. Together, our results demonstrated a link between COP1 regulation, p27 stability, and cell cycle growth (Figure 8E). The role of COP1 in attenuating p27 offers a new bridge for knowledge gap regarding the role of COP1 in cell cycle and tumorigenicity. Further identifying the structure and functional relationship between COP1 and p27 can provide the basis for developing inhibitors that will block COP1-mediated p27 degradation and functions for rational cancer therapy.
Materials and Methods
Cell culture and reagents
Human 293T, HeLa, and MDA-MB231 cells were cultured in Dulbecco’s modified Eagle medium/F12 medium supplemented with 10% fetal bovine serum, 100 units/ml penicillin G, 100 µg/ml streptomycin, and 0.25 µg/ml amphotericin. U2OS cells were maintained as previously described [42, 43]. For transient transfection, cells were transfected with DNA using either Lipofectamine 2000 (Invitrogen) or FuGENE HD (Roche) reagents according to protocols of the manufacturers. Antibodies to the following epitopes and proteins were purchased from the indicated vendors: Jab1 (GeneTex), Lamin B1 (Abcam), HA (12CA5, Roche), Pirh2 (Bethyl Laboratories), ubiquitin (Zymed Laboratories), p27 (BD Transduction Laboratories and Santa Cruz Biotechnology), and COP1 (Bethyl Laboratories and Santa Cruz Biotechnology). Flag (M2 monoclonal antibody), Tubulin, and Actin were purchased from Sigma. GFP, KPC1, SKP2, Myc (mouse monoclonal 9E10), Cyclin E, and CDK4 were purchased from Santa Cruz Biotechnology.
Plasmids
pCMV5-Flag-COP1 was kindly provided by E. Bianchi. pcDNA6-Myc-COP1 was constructed by our lab. The COP1 C136S/CS39S mutant was generated using PCR-directed mutagenesis (Stratagene) and verified by DNA sequencing. Wild-type (wt) GFP-COP1, GFP-COP1 NES mutant, and GFP-COP1 S387A were constructed by PCR cloning. The pET15b plasmids expressing Flag-tagged COP1 (aa1-334) and COP1 (aa 401-731) were generated using PCR. pET-p27 wt 1-86, 87-198, Flag-p27 wt, mutants (T157A, T187A, ΔJab1, ΔJab1+T187A) [36], and RFP-p27 Flag-p27 (VP→AA) mutant [44] were generated using PCR-directed mutagenesis.
Immunoprecipitation and immunoblotting
Total cell lysates were solubilized in lysis buffer (50mM Tris pH 7.5, 150mM NaCl, 1mM EDTA, 0.5% Nonidet P-40, 0.5% Triton X-100, 1mM phenylmethylsulfonyl fluoride, 1mM sodium fluoride, 5mM sodium orthovanadate, and 1 µg/ml each of aprotinin, leupeptin, and pepstatin) and processed as previously described [45, 46]. Lysates were immunoprecipitated with indicated antibodies. Proteins were resolved by SDS-PAGE gels and proteins were transferred to polyvinylidene difluoride membranes (Millipore). The membranes were blocked with 5% nonfat milk for 1 hour at room temperature prior to incubation with indicated primary antibodies. Subsequently, membranes were washed and incubated for 1 hour at room temperature with peroxidase-conjugated secondary antibodies (Thermo Scientific). Following several washes, chemiluminescent images of immunodetected bands on the membranes were recorded on X-ray films using the enhanced chemiluminescence system (Millipore).
In vitro binding assay
For the in vitro binding assay, both Myc-COP1 and PET-p27 were prepared by in vitro transcription and translation using the TNT system as previously described [45]. TNT protein was mixed and immunoprecipitated with anti-p27 followed by immunoblotting with anti-Myc.
In vivo ubiquitination assay
U2OS, and MDA-MB231 cells were used to detect endogenous p27 ubiquitination as previously described [44]. 293T cells were transiently co-transfected with indicated plasmids to detect exogenous p27 ubiquitination. Forty-eight hours later, cells were treated with 5 µg/ml MG132 (Sigma) for 6 hours before being harvested. Cell lysates were immunoprecipitated with anti-p27 and examined for the levels of p27 ubiquitination. In other cases, cells were lysed in denaturing buffer (6M guanidine-HCl, 0.1M Na2HPO4/NaH2PO4, 10mM imidazole). Cell lysates were then incubated with nickel beads for 3 hours, washed, and immunoblotted with anti-p27.
In vitro ubiquitination assay
Experiments were performed as previously described [42, 43]. Both Myc-COP1 and PET-p27 were prepared by in vitro transcription and translation using the TNT system.
For detection of ubiquitinated p27 in vitro, TNT p27 proteins were incubated with various combinations of ubiquitin (200 pmol), E1 (2 pmol), E2-UbcH5a/5b (10 pmol), in vitro translated COP1, and ATP (2mM) in a total volume of 50 μl for 1 hour at 37°C. Reaction products were resolved by 10% SDS-polyacrylamide gel and probed with anti-p27. His-Ubiquitin (UW 8610), E1 (UW 9410), and E2 (UW 9050) were purchased from BioMol International.
Cell lysates fractionated by gel filtration
Cell lysates were fractionated through a Superose 6 column (GEHealthcare) equilibrated with lysis buffer at a flow rate of 0.3 ml/minute as previously described [6]. Fractions of 300 µl were collected.
Generation of stable transfectants
To generate stable COP1 knockdown cell lines, MDA-MB231 cells were infected with lentiviral shRNA transduction particles (Sigma; NM_001001740 ring finger and WD repeat domain 2 MISSION shRNA lentiviral transduction particles) containing either control shRNA or COP1 shRNA. After infection, cells were selected with 2 µg/ml puromycin for 2 weeks. For generation of COP1 overexpression stable transfectants, U2OS cells were transfected with indicated plasmids for the generation of stable overexpression transfectants.
Thymidine-nocodazole block
Experiments were performed as previously described [47].
FACS cell-cycle analysis
Samples were analyzed using a BD Facscanto II flow cytometer (BD Biosciences) to determine the distribution in different cell-cycle phases (FACS). These experiments were carried out as previously described [47].
Soft agar colony formation, Foci formation, MTT assay
The experiments were performed as previously described [48].
Xenograft experiment
Athymic (nu/nu) mice were housed in AAALAC-approved barrier facilities. COP1-expressing cells infected with Ad-β-gal (MOI =100) or Ad-p27 (MOI =100) were harvested and injected into the flank of each mouse. Tumor volumes were measured and recorded. At the end of the experiment, the tumors were removed and weighed.
Acknowledgments and funding
This work was supported in part by grants from the National Institutes of Health (R01CA089266 to M.H.L.), Directed Medical Research Programs (DOD SIDA BC062166 to S.J.Y. and M.H.L.), Susan G. Komen Breast Cancer Foundation (KG081048 to S.J.Y. and M.H.L.), and the Fidelity foundation to M.H.L. The University of Texas MD Anderson Cancer Center is supported by NIH core grant CA16672.
conflict of interest
The authors declare that there is no conflict of interest.
References
1. Hardtke CS, Gohda K, Osterlund MT, Oyama T, Okada K and Deng XW. HY5 stability and activity in arabidopsis is regulated by phosphorylation in its COP1 binding domain. The EMBO journal. 2000; 19:4997-5006.
2. Wei W and Kaelin WG, Jr. Good COP1 or bad COP1? In vivo veritas. J Clin Invest. 2011; 121:1263-1265.
3. Marine JC. Spotlight on the role of COP1 in tumorigenesis. Nat Rev Cancer. 2012; 12:455-464.
4. Choi HH, Gully C, Su CH, Velazquez-Torres G, Chou PC, Tseng C, Zhao R, Phan L, Shaiken T, Chen J, Yeung SC and Lee MH. COP9 signalosome subunit 6 stabilizes COP1, which functions as an E3 ubiquitin ligase for 14-3-3sigma. Oncogene. 2011; 30:4791-4801.
5. Zhao R, Yeung SC, Chen J, Iwakuma T, Su CH, Chen B, Qu C, Zhang F, Chen YT, Lin YL, Lee DF, Jin F, Zhu R, Shaikenov T, Sarbassov D, Sahin A, et al. Subunit 6 of the COP9 signalosome promotes tumorigenesis in mice through stabilization of MDM2 and is upregulated in human cancers. J Clin Invest. 2011; 121:851-865.
6. Chen J, Shin JH, Zhao R, Phan L, Wang H, Xue Y, Post SM, Ho Choi H, Chen JS, Wang E, Zhou Z, Tseng C, Gully C, Velazquez-Torres G, Fuentes-Mattei E, Yeung G, et al. CSN6 drives carcinogenesis by positively regulating Myc stability. Nat Commun. 2014; 5:5384.
7. Migliorini D, Bogaerts S, Defever D, Vyas R, Denecker G, Radaelli E, Zwolinska A, Depaepe V, Hochepied T, Skarnes WC and Marine JC. Cop1 constitutively regulates c-Jun protein stability and functions as a tumor suppressor in mice. J Clin Invest. 2011; 121:1329-1343.
8. Vitari AC, Leong KG, Newton K, Yee C, O’Rourke K, Liu J, Phu L, Vij R, Ferrando R, Couto SS, Mohan S, Pandita A, Hongo JA, Arnott D, Wertz IE, Gao WQ, et al. COP1 is a tumour suppressor that causes degradation of ETS transcription factors. Nature. 2011; 474:403-406.
9. Dornan D, Wertz I, Shimizu H, Arnott D, Frantz GD, Dowd P, O’Rourke K, Koeppen H and Dixit VM. The ubiquitin ligase COP1 is a critical negative regulator of p53. Nature. 2004; 429:86-92. Epub 2004 Apr 2021.
10. Wei N and Deng XW. Making sense of the COP9 signalosome. A regulatory protein complex conserved from Arabidopsis to human. Trends Genet. 1999; 15:98-103.
11. Seeger M, Kraft R, Ferrell K, Bech-Otschir D, Dumdey R, Schade R, Gordon C, Naumann M and Dubiel W. A novel protein complex involved in signal transduction possessing similarities to 26S proteasome subunits. FASEB J. 1998; 12:469-478.
12. Richardson KS and Zundel W. The emerging role of the COP9 signalosome in cancer. Mol Cancer Res. 2005; 3:645-653.
13. Qi L, Heredia JE, Altarejos JY, Screaton R, Goebel N, Niessen S, Macleod IX, Liew CW, Kulkarni RN, Bain J, Newgard C, Nelson M, Evans RM, Yates J and Montminy M. TRB3 links the E3 ubiquitin ligase COP1 to lipid metabolism. Science. 2006; 312:1763-1766.
14. Dentin R, Liu Y, Koo SH, Hedrick S, Vargas T, Heredia J, Yates J, 3rd and Montminy M. Insulin modulates gluconeogenesis by inhibition of the coactivator TORC2. Nature. 2007; 449:366-369.
15. Li DQ, Ohshiro K, Reddy SD, Pakala SB, Lee MH, Zhang Y, Rayala SK and Kumar R. E3 ubiquitin ligase COP1 regulates the stability and functions of MTA1. Proc Natl Acad Sci U S A. 2009; 106:17493-17498.
16. Su CH, Zhao R, Velazquez-Torres G, Chen J, Gully C, Yeung SC and Lee MH. Nuclear export regulation of COP1 by 14-3-3sigma in response to DNA damage. Mol Cancer. 2010; 9:243.
17. Su CH, Zhao R, Zhang F, Qu C, Chen B, Feng YH, Phan L, Chen J, Wang H, Yeung SC and Lee MH. 14-3-3sigma exerts tumor-suppressor activity mediated by regulation of COP1 stability. Cancer Res. 2011; 71:884-894.
18. Kato S, Ding J, Pisck E, Jhala US and Du K. COP1 functions as a FoxO1 ubiquitin E3 ligase to regulate FoxO1-mediated gene expression. J Biol Chem. 2008; 283:35464-35473.
19. Polyak K, Lee MH, Erdjument-Bromage H, Koff A, Roberts JM, Tempst P and Massague J. Cloning of p27Kip1, a cyclin-dependent kinase inhibitor and a potential mediator of extracellular antimitogenic signals. Cell. 1994; 78:59-66.
20. Yang HY, Zhou BP, Hung MC and Lee MH. Oncogenic signals of HER-2/neu in regulating the stability of the cyclin-dependent kinase inhibitor p27. J Biol Chem. 2000; 275:24735-24739.
21. Newman L, Xia W, Yang HY, Sahin A, Bondy M, Lukmanji F, Hung MC and Lee MH. Correlation of p27 protein expression with HER-2/neu expression in breast cancer. Mol Carcinog. 2001; 30:169-175.
22. Yang HY, Shao R, Hung MC and Lee MH. p27 Kip1 inhibits HER2/neu-mediated cell growth and tumorigenesis. Oncogene. 2001; 20:3695-3702.
23. Zhang Y, Yang HY, Zhang XC, Yang H, Tsai M and Lee MH. Tumor suppressor ARF inhibits HER-2/neu-mediated oncogenic growth. Oncogene. 2004; 23:7132-7143.
24. Yang H, Zhao R, Yang HY and Lee MH. Constitutively active FOXO4 inhibits Akt activity, regulates p27 Kip1 stability, and suppresses HER2-mediated tumorigenicity. Oncogene. 2005; 24:1924-1935.
25. Liang J, Zubovitz J, Petrocelli T, Kotchetkov R, Connor MK, Han K, Lee JH, Ciarallo S, Catzavelos C, Beniston R, Franssen E and Slingerland JM. PKB/Akt phosphorylates p27, impairs nuclear import of p27 and opposes p27-mediated G1 arrest. Nat Med. 2002; 8:1153-1160. Epub 2002 Sep 1116.
26. Lee MH and Yang HY. Negative regulators of cyclin-dependent kinases and their roles in cancers. Cell Mol Life Sci. 2001; 58:1907-1922.
27. Nourse J, Firpo E, Flanagan WM, Coats S, Polyak K, Lee MH, Massague J, Crabtree GR and Roberts JM. Interleukin-2-mediated elimination of the p27Kip1 cyclin-dependent kinase inhibitor prevented by rapamycin. Nature. 1994; 372:570-573.
28. Yang H, Zhang Y, Zhao R, Wen YY, Fournier K, Wu HB, Yang HY, Diaz J, Laronga C and Lee MH. Negative cell cycle regulator 14-3-3sigma stabilizes p27 Kip1 by inhibiting the activity of PKB/Akt. Oncogene. 2006; 25:4585-4594.
29. Carrano AC, Eytan E, Hershko A and Pagano M. SKP2 is required for ubiquitin-mediated degradation of the CDK inhibitor p27. Nature cell biology. 1999; 1:193-199.
30. Gully CP, Velazquez-Torres G, Shin JH, Fuentes-Mattei E, Wang E, Carlock C, Chen J, Rothenberg D, Adams HP, Choi HH, Guma S, Phan L, Chou PC, Su CH, Zhang F, Chen JS, et al. Aurora B kinase phosphorylates and instigates degradation of p53. Proc Natl Acad Sci U S A. 2012; 109:E1513-1522.
31. Bianchi E, Denti S, Catena R, Rossetti G, Polo S, Gasparian S, Putignano S, Rogge L and Pardi R. Characterization of human constitutive photomorphogenesis protein 1, a RING finger ubiquitin ligase that interacts with Jun transcription factors and modulates their transcriptional activity. J Biol Chem. 2003; 278:19682-19690.
32. Kamura T, Hara T, Matsumoto M, Ishida N, Okumura F, Hatakeyama S, Yoshida M, Nakayama K and Nakayama KI. Cytoplasmic ubiquitin ligase KPC regulates proteolysis of p27(Kip1) at G1 phase. Nat Cell Biol. 2004; 6:1229-1235.
33. Halaby MJ, Hakem R and Hakem A. Pirh2: an E3 ligase with central roles in the regulation of cell cycle, DNA damage response, and differentiation. Cell Cycle. 2013; 12:2733-2737.
34. Tomoda K, Kubota Y and Kato J. Degradation of the cyclin-dependent-kinase inhibitor p27Kip1 is instigated by Jab1. Nature. 1999; 398:160-165.
35. Tomoda K, Kubota Y, Arata Y, Mori S, Maeda M, Tanaka T, Yoshida M, Yoneda-Kato N and Kato JY. The cytoplasmic shuttling and subsequent degradation of p27Kip1 mediated by Jab1/CSN5 and the COP9 signalosome complex. J Biol Chem. 2002; 277:2302-2310.
36. Yang HY, Yang H, Zhao R and Lee MH. Modified p27 Kip1 is efficient in suppressing HER2-mediated tumorigenicity. J Cell Biochem. 2006; 98:128-138.
37. Tsvetkov LM, Yeh KH, Lee SJ, Sun H and Zhang H. p27(Kip1) ubiquitination and degradation is regulated by the SCF(Skp2) complex through phosphorylated Thr187 in p27. Current biology : CB. 1999; 9:661-664.
38. Hao B, Zheng N, Schulman BA, Wu G, Miller JJ, Pagano M and Pavletich NP. Structural basis of the Cks1-dependent recognition of p27(Kip1) by the SCF(Skp2) ubiquitin ligase. Mol Cell. 2005; 20:9-19.
39. Sutterluty H, Chatelain E, Marti A, Wirbelauer C, Senften M, Muller U and Krek W. p45SKP2 promotes p27Kip1 degradation and induces S phase in quiescent cells. Nat Cell Biol. 1999; 1:207-214.
40. Hattori T, Isobe T, Abe K, Kikuchi H, Kitagawa K, Oda T, Uchida C and Kitagawa M. Pirh2 promotes ubiquitin-dependent degradation of the cyclin-dependent kinase inhibitor p27Kip1. Cancer Res. 2007; 67:10789-10795.
41. Leng RP, Lin Y, Ma W, Wu H, Lemmers B, Chung S, Parant JM, Lozano G, Hakem R and Benchimol S. Pirh2, a p53-induced ubiquitin-protein ligase, promotes p53 degradation. Cell. 2003; 112:779-791.
42. Wen YY, Chou PC, Pham L, Su CH, Chen J, Hsieh YC, Xue Y-W, Qu C-J, Gully C, Parreno K, Teng C, Hsu SL, Yeung SC, Wang HM and Lee MH. DNA damage-mediated c-Myc degradation requires 14-3-3 sigma. Cancer Hallmarks. 2013; 1:3-17.
43. Zhao R, Phan L, Chen B, Yang HY, Chen J, Che TF, Qiao Y, Zhang J, Yeung SC and Lee MH. Ubiquitination-mediated p57Kip2 Degradation by CSN5 Confers Cancer Cell Proliferation. Cancer Hallmarks. 2014; in press.
44. Choi HH, Su CH, Fang LK, Zhang J, Yeung SC and Lee MH. CSN6 deregulation impairs genome integrity in a COP1-dependent pathway. Oncotarget. 2015; in press.
45. Laronga C, Yang HY, Neal C and Lee MH. Association of the cyclin-dependent kinases and 14-3-3 sigma negatively regulates cell cycle progression. J Biol Chem. 2000; 275:23106-23112.
46. Fuentes-Mattei E, Velazquez-Torres G, Phan L, Zhang F, Chou PC, Shin JH, Choi HH, Chen JS, Zhao R, Chen J, Gully C, Carlock C, Qi Y, Zhang Y, Wu Y, Esteva FJ, et al. Effects of obesity on transcriptomic changes and cancer hallmarks in estrogen receptor-positive breast cancer. J Natl Cancer Inst. 2014; 106.
47. Gully CP, Zhang F, Chen J, Yeung JA, Velazquez-Torres G, Wang E, Yeung SC and Lee MH. Antineoplastic effects of an Aurora B kinase inhibitor in breast cancer. Mol Cancer. 2010; 9:42.
48. Yang HY, Wen YY, Chen CH, Lozano G and Lee MH. 14-3-3 sigma positively regulates p53 and suppresses tumor growth. Mol Cell Biol. 2003; 23:7096-7107.