
Introduction
CD200 is a type I membrane-associated glycoprotein and a member of the immunoglobin superfamily [1–3]. While initially reported in the rat thymus in 1979 as the antigen of the MRC OX2 antibody [4], CD200 expression has since been detected on a variety of cells of hematopoietic, such as macrophages, dendritic cells, B cells, and activated T cells, and non-hematopoietic origin, including neurons, endothelial cells, trophoblasts, and epithelial keratinocytes [4–10]. The receptor for CD200, CD200R, is expressed mainly in myeloid cells, but is also detected on lymphoid lineage cells, such as natural killer (NK) and T cells [11–14] and contains a cytoplasmic tail capable of initiating downstream signaling cascades [3]. As CD200 ligand contains a short cytoplasmic tail of 19 amino acid residues that lacks a consensus signaling motif [1, 3, 15], CD200 function is primarily mediated via cell-cell interaction-dependent engagement of the CD200-CD200R axis.
The CD200-CD200R axis primarily functions as an immunoregulatory signaling pathway. Mice lacking Cd200 exhibit elevated numbers of activated Cd200r+ macrophages and granulocytes [11] but exhibit normal myelopoiesis [16]. Phenotypically, Cd200 loss and concomitant increased activated macrophage levels manifest in chronic nervous system inflammation, early onset of experimental autoimmune encephalomyelitis and susceptibility to experimental autoimmune uveoretinitis [11, 17]. These observations underscore an essential role of Cd200 in maintaining tissue homeostasis by tempering the level of activated myeloid cells. Under pathological conditions, Cd200 also plays a key role in directly suppressing Th1-mediated inflammation [18–20], thereby orchestrating a balance between effective pathogen clearance and preventing immunopathology. Collectively, these observations confer a primary immunosuppressive function for CD200 in maintaining tissue homeostasis that is facilitated via engagement with CD200R.
CD200 expression is reported across most cancer types including hematologic malignancies such as acute myeloid leukemia (AML) [21], multiple myeloma (MM) [22], chronic lymphocytic leukemia (CLL) [23], and B-cell lymphoma [24]; solid tumors such as rectal [25], breast [26–27], colon [28–29], lung [30–31], ovarian [32], head and neck [33–34], glioma [35], pancreatic [36–37], and bladder [38]; and a variety of skin cancers including squamous cell carcinoma [39–41], basal cell carcinoma [42], Merkel cell carcinoma [43] and melanoma [32, 44–45].
Similar to other immune checkpoint proteins, such as cytotoxic T lymphocyte antigen 4 (CTLA-4) and program death-1 (PD-1), CD200 is thought to play a pro-tumorigenic role, via engagement of CD200R, in many tumor types primarily by suppressing anti-tumor T-cell and natural killer cell responses [46–49] (Figure 1A). In melanoma cells, CD200 can inhibit Th1 cytokine production [32] and, in CLL, cytotoxic T cell proliferation [47]. CD200 overexpression in AML can directly suppress anti-tumor NK cell and memory T cell functions [48–49], and the suppression of pro-inflammatory cytokines by CD4+ Th1 cells and CD8+ memory T cell numbers in AML patients can be restored with anti-CD200 treatment [48–49]. Collectively, these studies illustrate a key pro-tumorigenic role for CD200 in directly suppressing T-cell and NK cell anti-tumor function and that blocking the engagement of the CD200-CD200R axis may provide therapeutic benefit to patients with CD200-expressing tumors. Further supporting this concept, in syngeneic and xenograft murine tumor models, treatment with CD200 neutralizing antibodies restored lymphocyte-mediated anti-tumor responses in vivo [26, 50].
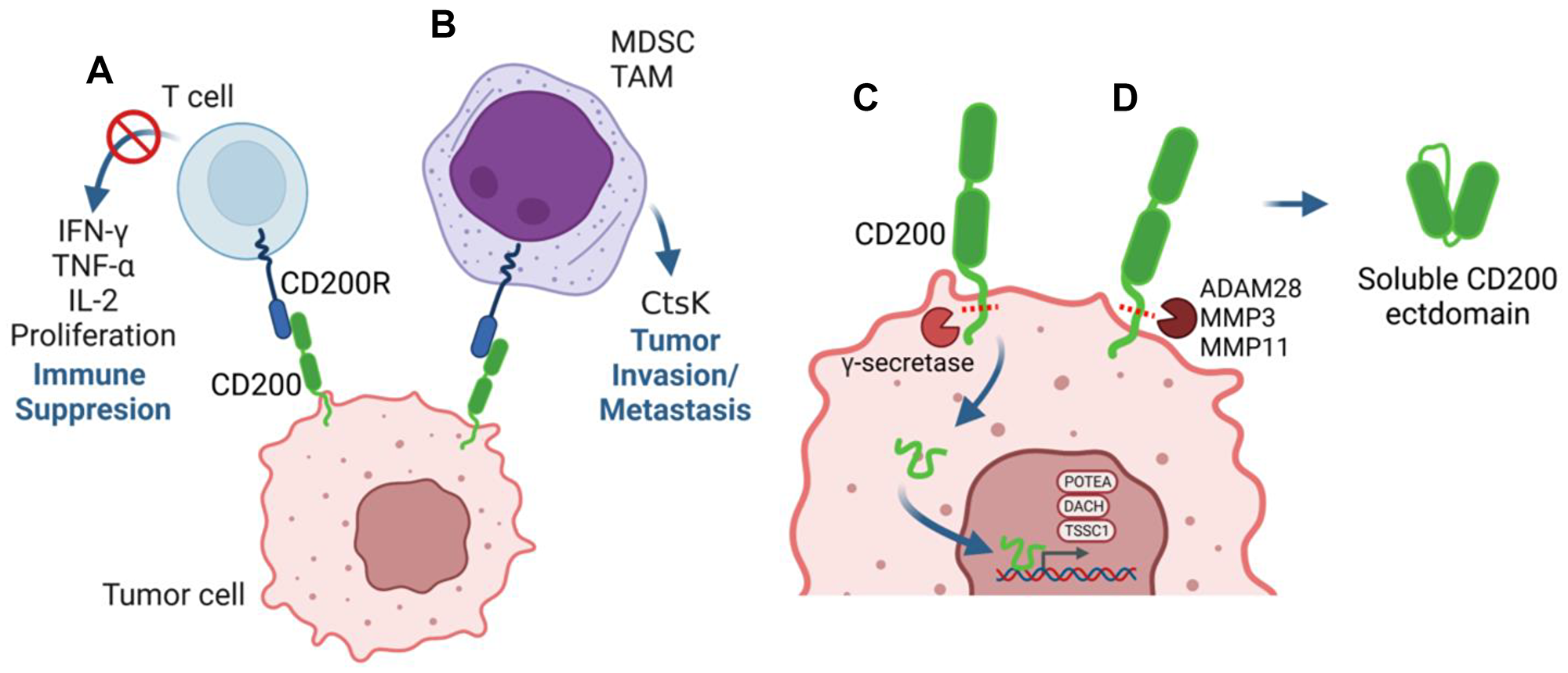
Figure 1: Multiple mechanisms underly the pro-tumorigenic role of CD200. (A, B) Tumor cell-extrinsic mechanisms for CD200 direct suppression of CD200R+ Th1 anti-tumor responses including pro-inflammatory cytokine production and proliferation (A) or CD200R-dependent activation of tumor-infiltrating myeloid lineages including MDSC and TAM (B). (C, D) Tumor cell-intrinsic mechanisms for CD200 include γ-secretase cleavage of the short CD200 cytoplasmic tail that translocates to the nucleus and regulates the expression of pro-tumorigenic target genes (C) or ADAM or MMP cleavage of CD200 to generate a soluble CD200 ectodomain (D), which has been shown to circulate systemically and in the tumor microenvironment. Abbreviations: MDSC: myeloid-derived suppressor cell; TAM: tumor-associated macrophage; ADAM: a disintegrin and metalloprotease domain; MMP: matrix metalloproteinase.
A recently completed Phase I trial assessed treatment responses of CLL and MM patients to samalizumab, a humanized anti-CD200 monoclonal antibody designed to block CD200-CD200R binding while minimizing cytotoxicity to CD200-expressing immune cell subsets (by using an IgG region with limited effector function) [51]. Decreases in overall tumor burden were observed in approximately 65% of CLL patients; however, a durable response was only observed in one out of twenty-three patients (4%). MM patients were refractory to samalizumab and a range of mild to moderate adverse outcomes were reported in the majority of CLL and MM patients, including skin rashes, joint stiffness/pain, headaches, and blood disorders. These adverse outcomes are consistent with the autoimmune phenotype reported in Cd200 null mice [11] and those observed in patients treated with PD-1 and/or CTLA-4 antibody therapy [52]. In 2017, a previous samalizumab Phase I trial was completed in patients with solid malignancies; however, to our knowledge the results of this study have yet to be disclosed. While the initial Phase I trial findings for samalizumab in CLL patients [51] certainly warrant further clinical investigation of different dosing regiments and its efficacy in additional blood cancers, shortcomings related to partial responses, lack of a durable response and the high incidence of mild to moderate toxicities raise concerns over the potential impact of CD200 antibody blockade therapy may hold for certain blood cancers other than CLL and solid malignancies.
A plausible explanation for the observed shortcomings in the samalizumab Phase I trial may be the alternative mechanisms for the CD200 pro-tumorigenic role that have recently emerged. Some of these alternative mechanisms extend beyond direct suppression of anti-tumor T cell responses and, as such, may not be susceptible to CD200 antibody blockade. First, in addition to targeting T-cell and NK cell numbers and/or activity, the recruitment and function of myeloid derived suppressor cells (MDSCs) and tumor-associated macrophages (TAMs) are known to be regulated by the CD200-CD200R axis in human cancer [37, 39, 43, 53] (Figure 1B). Initial work from our laboratory demonstrated MDSCs and TAMs constitute greater than 90% of the CD200R+ cells in the microenvironment of cutaneous squamous cell carcinoma (cSCC) and that production of critical microenvironment cues by these tumor-infiltrating myeloid lineages, including GM-CSF and G-CSF, was dependent on engagement of the Cd200-Cd200r axis [39]. More recently, using a Cd200 conditional null mouse model, we identified the collagen peptidase Cathepsin K (Ctsk) as a crucial target gene of the Cd200-Cd200r signaling axis in Cd200r+ tumor-infiltrating myeloid lineages, which was required for tumor cell invasion and metastasis [54] (Figure 1B). Interestingly, CTSK is a known biomarker for a variety of cancer types [55] and its expression is associated with metastasis of human solid malignancies [56–57] suggesting a broader functional role for CTSK in human tumor metastasis.
Adding to this complexity, non-canonical mechanisms for CD200 in tumorigenesis have also recently emerged. The intracellular tail of CD200, previously thought to be signaling inert, is a target for cleavage by γ-secretase, resulting in release of a CD200 tail fragment that is capable of nuclear translocation and DNA binding [58]. DNA binding by the CD200 cytoplasmic tail leads to increased expression of transcription factors associated with leukemic cell growth [58] (Figure 1C). In the extracellular space, ADAM28-mediated CD200 ectodomain shedding leads to increased serum levels of a biologically-active, soluble CD200 ectodomain fragment in B-cell CLL patients [59] (Figure 1D). Recently, MMP-mediated CD200 ectodomain shedding in basal cell carcinoma was shown to regulate NK cell dysfunction and apoptosis in the microenvironment [60] (Figure 1D). It remains an open question as to whether antibody therapies designed to block membrane-tethered CD200-CD200R binding may be effective against soluble CD200 ectodomain fragments of variable sizes that may be generated by different protease families. Collectively, these observations underscore i) a broader pro-tumorigenic role for CD200 in the tumor microenvironment and ii) potential pleiotropic mechanisms adopted by CD200 to mediate tumor cell survival, invasion and metastasis that may be difficult to block via a monoclonal antibody treatment modality targeting a specific ectodomain region.
Overall, the inconsistent response rates across different tumor types, low incidence of durable responses and observed undesirable cytotoxicity outcomes to samalizumab therapy together with newly emerging non-canonical roles for CD200 in cancer provides rationale for alternative strategies designed to efficaciously target this protein. An underexplored aspect of the role of CD200 in cancer is the identification of tumor-specific mechanisms for the regulation of CD200 expression. A better understanding of CD200 regulatory mechanisms may be relevant for multiple CD200 pro-tumorigenic functions including engagement of the CD200-CD200R axis, transcriptional mechanisms related to the cleaved cytoplasmic tail and ectodomain shedding. Clouding our understanding of this issue is that, in normal and neoplastic cells, the induction or suppression of CD200 expression is reported to be associated with a wide variety of signaling pathways and CD200 expression can be regulated by both constitutive and inducible pathways. Genomic analysis of the human CD200 promoter identified C/EBPβ as a key regulator of constitutive CD200 expression [61]; while upstream enhancer regions harbor binding sites for IFNγ- and TNFα-induced signaling pathway effectors that are proposed to be necessary for inducible CD200 expression [62].
In the central nervous system (CNS), the CD200-CD200R axis is a fundamental facilitator of neuron-microglia cell-cell interactions that maintain physiological levels of inflammation. Multiple pathways are reported to regulate CD200 expression in the CNS. In both astrocytes [63] and neurons [64], CD200 expression is regulated by FGFR1 activation, which is critical for suppression of neuroinflammation. In addition to FGFR signaling, treatment with PPARγ ligands can suppress CD200 induction in activated glial cells [65], which is thought to play a functional role in PPARγ-mediated neuroprotection. Under pathological conditions, CD200 expression in microglia can be inhibited by IFNγ derived from a leaky blood brain barrier [66]. In other tissues, the molecules involved in the regulation of CD200 expression appear to be unique to each cell lineage. CD200 is a downstream target of p53 during caspase-dependent dendritic cell apoptosis [67]. In a mouse model of meningococcal infection, CD200 induction in macrophages is dependent on TLR4 and downstream NF-κB signaling [68]. In bone marrow mesenchymal stem cells, CD200 expression can be induced by osteogenic and pro-inflammatory cytokines also in a NF-κB-dependent manner [69]. In the skeletal system, CD200 expression in osteoblasts is dependent on IL15RA signaling [70]. In the lung, both airway epithelial cell and capillary endothelial cell expression of CD200 is shown to be regulated by corticosteroids [71–72]. In hair follicle epithelial progenitors, CD200 expression is reported to be dependent on β1 integrin activation [73].
In human cancers, deeper knowledge gaps exist in our understanding of the regulation of CD200 expression. First, relative to normal tissue, there is a paucity of information regarding CD200 expression regulation in human cancers. Second, CD200 expression may be present at early stages of development for certain tumors, whereas, in other lesions, CD200 expression is induced during late-stage progression and only in a sub-set of tumor cells. Third, there appears to be little to no overlap in the putative molecular regulators of CD200 expression across different tumor types. Currently, these bottlenecks preclude our ability to effectively pinpoint potential regulatory targets to block CD200 expression. In metastatic melanoma, CD200 is regulated by ERK activation downstream of N-RAS or B-RAF mutations [44] suggesting that CD200 induction is an early event in melanoma pathogenesis. In human and murine cSCC lesions, we previously observed little to no CD200 expression in early, well-differentiated tumors [39]. However, CD200 expression was induced in poorly-differentiated primary and metastatic cSCC with enrichment of CD200 localized to leading edge tumor cells [39, 54]. In endometriosis patients, CD200 expression is upregulated in lesional stromal cells and in the blood and 17β-estradiol treatment of stromal cells in culture increased expression of CD200 [74]. In colorectal carcinoma cells, CD200 expression is dependent on the activity of the Rho GTPase effector protein FMNL2 [75]. CD200 is a target of miR-499a and a polymorphism in miR-499a is a poor prognostic factor for non-small cell lung cancer cases who also exhibit elevated CD200 expression [76].
In the future, unbiased genomic- and proteomic-based approaches may help to clarify these issues by identifying tumor-specific mechanisms of CD200 expression regulation across a variety of human cancers that may be leveraged for broader therapeutic benefit.
CONFLICTS OF INTEREST
Authors have no conflicts of interest to declare.
FUNDING
A. Shao and D.M. Owens. were supported by NIH R01AG073874.
References
1. Barclay AN, Wright GJ, Brooke G, Brown MH. CD200 and membrane protein interactions in the control of myeloid cells. Trends Immunol. 2002; 23:285–90. https://doi.org/10.1016/s1471-4906(02)02223-8. [PubMed].
2. Barclay AN, Clark MJ, McCaughan GW. Neuronal/lymphoid membrane glycoprotein MRC OX-2 is a member of the immunoglobulin superfamily with a light-chain-like structure. Biochem Soc Symp. 1986; 51:149–57. [PubMed].
3. Hatherley D, Lea SM, Johnson S, Barclay AN. Structures of CD200/CD200 receptor family and implications for topology, regulation, and evolution. Structure. 2013; 21:820–32. https://doi.org/10.1016/j.str.2013.03.008. [PubMed].
4. McMaster WR, Williams AF. Monoclonal antibodies to Ia antigens from rat thymus: cross reactions with mouse and human and use in purification of rat Ia glycoproteins. Immunol Rev. 1979; 47:117–37. https://doi.org/10.1111/j.1600-065x.1979.tb00291.x. [PubMed].
5. Webb M, Barclay AN. Localisation of the MRC OX-2 glycoprotein on the surfaces of neurones. J Neurochem. 1984; 43:1061–67. https://doi.org/10.1111/j.1471-4159.1984.tb12844.x. [PubMed].
6. Bukovský A, Presl J, Zidovský J. Association of some cell surface antigens of lymphoid cells and cell surface differentiation antigens with early rat pregnancy. Immunology. 1984; 52:631–40. [PubMed].
7. Dick AD, Broderick C, Forrester JV, Wright GJ. Distribution of OX2 antigen and OX2 receptor within retina. Invest Ophthalmol Vis Sci. 2001; 42:170–76. [PubMed].
8. Wright GJ, Jones M, Puklavec MJ, Brown MH, Barclay AN. The unusual distribution of the neuronal/lymphoid cell surface CD200 (OX2) glycoprotein is conserved in humans. Immunology. 2001; 102:173–79. https://doi.org/10.1046/j.1365-2567.2001.01163.x. [PubMed].
9. Rosenblum MD, Olasz EB, Yancey KB, Woodliff JE, Lazarova Z, Gerber KA, Truitt RL. Expression of CD200 on epithelial cells of the murine hair follicle: a role in tissue-specific immune tolerance? J Invest Dermatol. 2004; 123:880–87. https://doi.org/10.1111/j.0022-202X.2004.23461.x. [PubMed].
10. Meyer KC, Klatte JE, Dinh HV, Harries MJ, Reithmayer K, Meyer W, Sinclair R, Paus R. Evidence that the bulge region is a site of relative immune privilege in human hair follicles. Br J Dermatol. 2008; 159:1077–85. https://doi.org/10.1111/j.1365-2133.2008.08818.x. [PubMed].
11. Hoek RM, Ruuls SR, Murphy CA, Wright GJ, Goddard R, Zurawski SM, Blom B, Homola ME, Streit WJ, Brown MH, Barclay AN, Sedgwick JD. Down-regulation of the macrophage lineage through interaction with OX2 (CD200). Science. 2000; 290:1768–71. https://doi.org/10.1126/science.290.5497.1768. [PubMed].
12. Jenmalm MC, Cherwinski H, Bowman EP, Phillips JH, Sedgwick JD. Regulation of myeloid cell function through the CD200 receptor. J Immunol. 2006; 176:191–99. https://doi.org/10.4049/jimmunol.176.1.191. [PubMed].
13. Stack G, Jones E, Marsden M, Stacey MA, Snelgrove RJ, Lacaze P, Jacques LC, Cuff SM, Stanton RJ, Gallimore AM, Hussell T, Wilkinson GW, Ghazal P, et al. CD200 receptor restriction of myeloid cell responses antagonizes antiviral immunity and facilitates cytomegalovirus persistence within mucosal tissue. PLoS Pathog. 2015; 11:e1004641. https://doi.org/10.1371/journal.ppat.1004641. [PubMed].
14. Rijkers ES, de Ruiter T, Baridi A, Veninga H, Hoek RM, Meyaard L. The inhibitory CD200R is differentially expressed on human and mouse T and B lymphocytes. Mol Immunol. 2008; 45:1126–35. https://doi.org/10.1016/j.molimm.2007.07.013. [PubMed].
15. Moreaux J, Veyrune JL, Reme T, De Vos J, Klein B. CD200: a putative therapeutic target in cancer. Biochem Biophys Res Commun. 2008; 366:117–22. https://doi.org/10.1016/j.bbrc.2007.11.103. [PubMed].
16. Rijkers ES, de Ruiter T, Buitenhuis M, Veninga H, Hoek RM, Meyaard L. Ligation of CD200R by CD200 is not required for normal murine myelopoiesis. Eur J Haematol. 2007; 79:410–16. https://doi.org/10.1111/j.1600-0609.2007.00920.x. [PubMed].
17. Broderick C, Hoek RM, Forrester JV, Liversidge J, Sedgwick JD, Dick AD. Constitutive retinal CD200 expression regulates resident microglia and activation state of inflammatory cells during experimental autoimmune uveoretinitis. Am J Pathol. 2002; 161:1669–77. https://doi.org/10.1016/S0002-9440(10)64444-6. [PubMed].
18. Gorczynski RM, Chen Z, Lee L, Yu K, Hu J. Anti-CD200R ameliorates collagen-induced arthritis in mice. Clin Immunol. 2002; 104:256–64. https://doi.org/10.1006/clim.2002.5232. [PubMed].
19. Rygiel TP, Rijkers ES, de Ruiter T, Stolte EH, van der Valk M, Rimmelzwaan GF, Boon L, van Loon AM, Coenjaerts FE, Hoek RM, Tesselaar K, Meyaard L. Lack of CD200 enhances pathological T cell responses during influenza infection. J Immunol. 2009; 183:1990–96. https://doi.org/10.4049/jimmunol.0900252. [PubMed].
20. Snelgrove RJ, Goulding J, Didierlaurent AM, Lyonga D, Vekaria S, Edwards L, Gwyer E, Sedgwick JD, Barclay AN, Hussell T. A critical function for CD200 in lung immune homeostasis and the severity of influenza infection. Nat Immunol. 2008; 9:1074–83. https://doi.org/10.1038/ni.1637. [PubMed].
21. Tonks A, Hills R, White P, Rosie B, Mills KI, Burnett AK, Darley RL. CD200 as a prognostic factor in acute myeloid leukaemia. Leukemia. 2007; 21:566–68. https://doi.org/10.1038/sj.leu.2404559. [PubMed].
22. Moreaux J, Hose D, Reme T, Jourdan E, Hundemer M, Legouffe E, Moine P, Bourin P, Moos M, Corre J, Möhler T, De Vos J, Rossi JF, et al. CD200 is a new prognostic factor in multiple myeloma. Blood. 2006; 108:4194–97. https://doi.org/10.1182/blood-2006-06-029355. [PubMed].
23. McWhirter JR, Kretz-Rommel A, Saven A, Maruyama T, Potter KN, Mockridge CI, Ravey EP, Qin F, Bowdish KS. Antibodies selected from combinatorial libraries block a tumor antigen that plays a key role in immunomodulation. Proc Natl Acad Sci U S A. 2006; 103:1041–46. https://doi.org/10.1073/pnas.0510081103. [PubMed].
24. Miao Y, Fan L, Wu YJ, Xia Y, Qiao C, Wang Y, Wang L, Hong M, Zhu HY, Xu W, Li JY. Low expression of CD200 predicts shorter time-to-treatment in chronic lymphocytic leukemia. Oncotarget. 2016; 7:13551–62. https://doi.org/10.18632/oncotarget.6948. [PubMed].
25. Bisgin A, Meng WJ, Adell G, Sun XF. Interaction of CD200 Overexpression on Tumor Cells with CD200R1 Overexpression on Stromal Cells: An Escape from the Host Immune Response in Rectal Cancer Patients. J Oncol. 2019; 2019:5689464. https://doi.org/10.1155/2019/5689464. [PubMed].
26. Gorczynski RM, Chen Z, Diao J, Khatri I, Wong K, Yu K, Behnke J. Breast cancer cell CD200 expression regulates immune response to EMT6 tumor cells in mice. Breast Cancer Res Treat. 2010; 123:405–15. https://doi.org/10.1007/s10549-009-0667-8. [PubMed].
27. Curry A, Khatri I, Kos O, Zhu F, Gorczynski R. Importance of CD200 expression by tumor or host cells to regulation of immunotherapy in a mouse breast cancer model. PLoS One. 2017; 12:e0171586. https://doi.org/10.1371/journal.pone.0171586. [PubMed].
28. Kawasaki BT, Mistree T, Hurt EM, Kalathur M, Farrar WL. Co-expression of the toleragenic glycoprotein, CD200, with markers for cancer stem cells. Biochem Biophys Res Commun. 2007; 364:778–82. https://doi.org/10.1016/j.bbrc.2007.10.067. [PubMed].
29. Gemei M, Di Noto R, Mirabelli P, Del Vecchio L. Cytometric profiling of CD133+ cells in human colon carcinoma cell lines identifies a common core phenotype and cell type-specific mosaics. Int J Biol Markers. 2013; 28:267–73. https://doi.org/10.5301/JBM.5000020. [PubMed].
30. Bohling SD, Davis E, Thompson K, Kussick SJ, Love J. Flow cytometric analysis of CD200 expression by pulmonary small cell carcinoma. Cytometry B Clin Cytom. 2016; 90:493–98. https://doi.org/10.1002/cyto.b.21340. [PubMed].
31. Love JE, Thompson K, Kilgore MR, Westerhoff M, Murphy CE, Papanicolau-Sengos A, McCormick KA, Shankaran V, Vandeven N, Miller F, Blom A, Nghiem PT, Kussick SJ. CD200 Expression in Neuroendocrine Neoplasms. Am J Clin Pathol. 2017; 148:236–42. https://doi.org/10.1093/ajcp/aqx071. [PubMed].
32. Siva A, Xin H, Qin F, Oltean D, Bowdish KS, Kretz-Rommel A. Immune modulation by melanoma and ovarian tumor cells through expression of the immunosuppressive molecule CD200. Cancer Immunol Immunother. 2008; 57:987–96. https://doi.org/10.1007/s00262-007-0429-6. [PubMed].
33. Clasper S, Royston D, Baban D, Cao Y, Ewers S, Butz S, Vestweber D, Jackson DG. A novel gene expression profile in lymphatics associated with tumor growth and nodal metastasis. Cancer Res. 2008; 68:7293–303. https://doi.org/10.1158/0008-5472.CAN-07-6506. [PubMed].
34. Jung YS, Vermeer PD, Vermeer DW, Lee SJ, Goh AR, Ahn HJ, Lee JH. CD200: association with cancer stem cell features and response to chemoradiation in head and neck squamous cell carcinoma. Head Neck. 2015; 37:327–35. https://doi.org/10.1002/hed.23608. [PubMed].
35. Moertel CL, Xia J, LaRue R, Waldron NN, Andersen BM, Prins RM, Okada H, Donson AM, Foreman NK, Hunt MA, Pennell CA, Olin MR. CD200 in CNS tumor-induced immunosuppression: the role for CD200 pathway blockade in targeted immunotherapy. J Immunother Cancer. 2014; 2:46. https://doi.org/10.1186/s40425-014-0046-9. [PubMed].
36. Lawlor RT, Daprà V, Girolami I, Pea A, Pilati C, Nottegar A, Piccoli P, Parolini C, Sperandio N, Capelli P, Scarpa A, Luchini C. CD200 expression is a feature of solid pseudopapillary neoplasms of the pancreas. Virchows Arch. 2019; 474:105–9. https://doi.org/10.1007/s00428-018-2437-7. [PubMed].
37. Choueiry F, Torok M, Shakya R, Agrawal K, Deems A, Benner B, Hinton A, Shaffer J, Blaser BW, Noonan AM, Williams TM, Dillhoff M, Conwell DL, et al. CD200 promotes immunosuppression in the pancreatic tumor microenvironment. J Immunother Cancer. 2020; 8:e000189. https://doi.org/10.1136/jitc-2019-000189. [PubMed].
38. Rexin P, Tauchert A, Hänze J, Heers H, Schmidt A, Hofmann R, Hegele A. The Immune Checkpoint Molecule CD200 Is Associated with Tumor Grading and Metastasis in Bladder Cancer. Anticancer Res. 2018; 38:2749–54. https://doi.org/10.21873/anticanres.12517. [PubMed].
39. Stumpfova M, Ratner D, Desciak EB, Eliezri YD, Owens DM. The immunosuppressive surface ligand CD200 augments the metastatic capacity of squamous cell carcinoma. Cancer Res. 2010; 70:2962–72. https://doi.org/10.1158/0008-5472.CAN-09-4380. [PubMed].
40. Belkin DA, Mitsui H, Wang CQ, Gonzalez J, Zhang S, Shah KR, Coats I, Suàrez-Fariñas M, Krueger JG, Felsen D, Carucci JA. CD200 upregulation in vascular endothelium surrounding cutaneous squamous cell carcinoma. JAMA Dermatol. 2013; 149:178–86. https://doi.org/10.1001/jamadermatol.2013.1609. [PubMed].
41. Li L, Tian Y, Shi C, Zhang H, Zhou Z. Over-Expression of CD200 Predicts Poor Prognosis in Cutaneous Squamous Cell Carcinoma. Med Sci Monit. 2016; 22:1079–84. https://doi.org/10.12659/msm.895245. [PubMed].
42. Colmont CS, Benketah A, Reed SH, Hawk NV, Telford WG, Ohyama M, Udey MC, Yee CL, Vogel JC, Patel GK. CD200-expressing human basal cell carcinoma cells initiate tumor growth. Proc Natl Acad Sci U S A. 2013; 110:1434–39. https://doi.org/10.1073/pnas.1211655110. [PubMed].
43. Gaiser MR, Weis CA, Gaiser T, Jiang H, Buder-Bakhaya K, Herpel E, Warth A, Xiao Y, Miao L, Brownell I. Merkel cell carcinoma expresses the immunoregulatory ligand CD200 and induces immunosuppressive macrophages and regulatory T cells. Oncoimmunology. 2018; 7:e1426517. https://doi.org/10.1080/2162402X.2018.1426517. [PubMed].
44. Petermann KB, Rozenberg GI, Zedek D, Groben P, McKinnon K, Buehler C, Kim WY, Shields JM, Penland S, Bear JE, Thomas NE, Serody JS, Sharpless NE. CD200 is induced by ERK and is a potential therapeutic target in melanoma. J Clin Invest. 2007; 117:3922–29. https://doi.org/10.1172/JCI32163. [PubMed].
45. Talebian F, Liu JQ, Liu Z, Khattabi M, He Y, Ganju R, Bai XF. Melanoma cell expression of CD200 inhibits tumor formation and lung metastasis via inhibition of myeloid cell functions. PLoS One. 2012; 7:e31442. https://doi.org/10.1371/journal.pone.0031442. [PubMed].
46. Kretz-Rommel A, Qin F, Dakappagari N, Ravey EP, McWhirter J, Oltean D, Frederickson S, Maruyama T, Wild MA, Nolan MJ, Wu D, Springhorn J, Bowdish KS. CD200 expression on tumor cells suppresses antitumor immunity: new approaches to cancer immunotherapy. J Immunol. 2007; 178:5595–605. https://doi.org/10.4049/jimmunol.178.9.5595. [PubMed].
47. Pallasch CP, Ulbrich S, Brinker R, Hallek M, Uger RA, Wendtner CM. Disruption of T cell suppression in chronic lymphocytic leukemia by CD200 blockade. Leuk Res. 2009; 33:460–64. https://doi.org/10.1016/j.leukres.2008.08.021. [PubMed].
48. Coles SJ, Wang EC, Man S, Hills RK, Burnett AK, Tonks A, Darley RL. CD200 expression suppresses natural killer cell function and directly inhibits patient anti-tumor response in acute myeloid leukemia. Leukemia. 2011; 25:792–99. https://doi.org/10.1038/leu.2011.1. [PubMed].
49. Coles SJ, Hills RK, Wang EC, Burnett AK, Man S, Darley RL, Tonks A. Expression of CD200 on AML blasts directly suppresses memory T-cell function. Leukemia. 2012; 26:2148–51. https://doi.org/10.1038/leu.2012.77. [PubMed].
50. Gorczynski RM, Chen Z, Hu J, Kai Y, Lei J. Evidence of a role for CD200 in regulation of immune rejection of leukaemic tumour cells in C57BL/6 mice. Clin Exp Immunol. 2001; 126:220–29. https://doi.org/10.1046/j.1365-2249.2001.01689.x. [PubMed].
51. Mahadevan D, Lanasa MC, Farber C, Pandey M, Whelden M, Faas SJ, Ulery T, Kukreja A, Li L, Bedrosian CL, Zhang X, Heffner LT. Phase I study of samalizumab in chronic lymphocytic leukemia and multiple myeloma: blockade of the immune checkpoint CD200. J Immunother Cancer. 2019; 7:227. https://doi.org/10.1186/s40425-019-0710-1. [PubMed].
52. Teply BA, Lipson EJ. Identification and management of toxicities from immune checkpoint-blocking drugs. Oncology (Williston Park). 2014 (Suppl 3); 28:30–38. [PubMed].
53. Takahashi H, Sakakura K, Kudo T, Toyoda M, Kaira K, Oyama T, Chikamatsu K. Cancer-associated fibroblasts promote an immunosuppressive microenvironment through the induction and accumulation of protumoral macrophages. Oncotarget. 2017; 8:8633–47. https://doi.org/10.18632/oncotarget.14374. [PubMed].
54. Khan IZ, Del Guzzo CA, Shao A, Cho J, Du R, Cohen AO, Owens DM. The CD200-CD200R Axis Promotes Squamous Cell Carcinoma Metastasis via Regulation of Cathepsin K. Cancer Res. 2021; 81:5021–32. https://doi.org/10.1158/0008-5472.CAN-20-3251. [PubMed].
55. Qian D, He L, Zhang Q, Li W, Tang D, Wu C, Yang F, Li K, Zhang H. Cathepsin K: A Versatile Potential Biomarker and Therapeutic Target for Various Cancers. Curr Oncol. 2022; 29:5963–87. https://doi.org/10.3390/curroncol29080471. [PubMed].
56. Xu H, Ma Y, Zhang Y, Pan Z, Lu Y, Liu P, Lu B. Identification of Cathepsin K in the Peritoneal Metastasis of Ovarian Carcinoma Using In-silico, Gene Expression Analysis. J Cancer. 2016; 7:722–29. https://doi.org/10.7150/jca.14277. [PubMed].
57. Wu N, Wang Y, Wang K, Zhong B, Liao Y, Liang J, Jiang N. Cathepsin K regulates the tumor growth and metastasis by IL-17/CTSK/EMT axis and mediates M2 macrophage polarization in castration-resistant prostate cancer. Cell Death Dis. 2022; 13:813. https://doi.org/10.1038/s41419-022-05215-8. [PubMed].
58. Chen Z, Kapus A, Khatri I, Kos O, Zhu F, Gorczynski RM. Cell membrane-bound CD200 signals both via an extracellular domain and following nuclear translocation of a cytoplasmic fragment. Leuk Res. 2018; 69:72–80. https://doi.org/10.1016/j.leukres.2018.04.007. [PubMed].
59. Twito T, Chen Z, Khatri I, Wong K, Spaner D, Gorczynski R. Ectodomain shedding of CD200 from the B-CLL cell surface is regulated by ADAM28 expression. Leuk Res. 2013; 37:816–21. https://doi.org/10.1016/j.leukres.2013.04.014. [PubMed].
60. Morgan HJ, Rees E, Lanfredini S, Powell KA, Gore J, Gibbs A, Lovatt C, Davies GE, Olivero C, Shorning BY, Tornillo G, Tonks A, Darley R, et al. CD200 ectodomain shedding into the tumor microenvironment leads to NK cell dysfunction and apoptosis. J Clin Invest. 2022; 132:e150750. https://doi.org/10.1172/JCI150750. [PubMed].
61. Chen Z, Marsden PA, Gorczynski RM. Cloning and characterization of the human CD200 promoter region. Mol Immunol. 2006; 43:579–87. https://doi.org/10.1016/j.molimm.2005.04.014. [PubMed].
62. Chen Z, Marsden PA, Gorczynski RM. Role of a distal enhancer in the transcriptional responsiveness of the human CD200 gene to interferon-gamma and tumor necrosis factor-alpha. Mol Immunol. 2009; 46:1951–63. https://doi.org/10.1016/j.molimm.2009.03.015. [PubMed].
63. Downer EJ, Cowley TR, Cox F, Maher FO, Berezin V, Bock E, Lynch MA. A synthetic NCAM-derived mimetic peptide, FGL, exerts anti-inflammatory properties via IGF-1 and interferon-gamma modulation. J Neurochem. 2009; 109:1516–25. https://doi.org/10.1111/j.1471-4159.2009.06076.x. [PubMed].
64. Cox FF, Berezin V, Bock E, Lynch MA. The neural cell adhesion molecule-derived peptide, FGL, attenuates lipopolysaccharide-induced changes in glia in a CD200-dependent manner. Neuroscience. 2013; 235:141–48. https://doi.org/10.1016/j.neuroscience.2012.12.030. [PubMed].
65. Dentesano G, Serratosa J, Tusell JM, Ramón P, Valente T, Saura J, Solà C. CD200R1 and CD200 expression are regulated by PPAR-γ in activated glial cells. Glia. 2014; 62:982–98. https://doi.org/10.1002/glia.22656. [PubMed].
66. Singh V, Kushwaha S, Gera R, Ansari JA, Mishra J, Dewangan J, Patnaik S, Ghosh D. Sneaky Entry of IFNγ Through Arsenic-Induced Leaky Blood-Brain Barrier Reduces CD200 Expression by Microglial pro-Inflammatory Cytokine. Mol Neurobiol. 2019; 56:1488–99. https://doi.org/10.1007/s12035-018-1155-0. [PubMed].
67. Rosenblum MD, Olasz E, Woodliff JE, Johnson BD, Konkol MC, Gerber KA, Orentas RJ, Sandford G, Truitt RL. CD200 is a novel p53-target gene involved in apoptosis-associated immune tolerance. Blood. 2004; 103:2691–98. https://doi.org/10.1182/blood-2003-09-3184. [PubMed].
68. Mukhopadhyay S, Plüddemann A, Hoe JC, Williams KJ, Varin A, Makepeace K, Aknin ML, Bowdish DM, Smale ST, Barclay AN, Gordon S. Immune inhibitory ligand CD200 induction by TLRs and NLRs limits macrophage activation to protect the host from meningococcal septicemia. Cell Host Microbe. 2010; 8:236–47. https://doi.org/10.1016/j.chom.2010.08.005. [PubMed].
69. Pontikoglou C, Langonné A, Ba MA, Varin A, Rosset P, Charbord P, Sensébé L, Deschaseaux F. CD200 expression in human cultured bone marrow mesenchymal stem cells is induced by pro-osteogenic and pro-inflammatory cues. J Cell Mol Med. 2016; 20:655–65. https://doi.org/10.1111/jcmm.12752. [PubMed].
70. Loro E, Ramaswamy G, Chandra A, Tseng WJ, Mishra MK, Shore EM, Khurana TS. IL15RA is required for osteoblast function and bone mineralization. Bone. 2017; 103:20–30. https://doi.org/10.1016/j.bone.2017.06.003. [PubMed].
71. BinMahfouz H, Borthakur B, Yan D, George T, Giembycz MA, Newton R. Superiority of combined phosphodiesterase PDE3/PDE4 inhibition over PDE4 inhibition alone on glucocorticoid- and long-acting β2-adrenoceptor agonist-induced gene expression in human airway epithelial cells. Mol Pharmacol. 2015; 87:64–76. https://doi.org/10.1124/mol.114.093393. [PubMed].
72. Tsai MH, Chu CC, Wei TS, Chiu MM, Chang CY, Wei IH, Chien HF, Chen HM, Wu CH, Jiang-Shieh YF. CD200 in growing rat lungs: developmental expression and control by dexamethasone. Cell Tissue Res. 2015; 359:729–42. https://doi.org/10.1007/s00441-014-2065-8. [PubMed].
73. Ernst N, Yay A, Bíró T, Tiede S, Humphries M, Paus R, Kloepper JE. β1 integrin signaling maintains human epithelial progenitor cell survival in situ and controls proliferation, apoptosis and migration of their progeny. PLoS One. 2013; 8:e84356. https://doi.org/10.1371/journal.pone.0084356. [PubMed].
74. Weng LC, Hou SH, Lei ST, Peng HY, Li MQ, Zhao D. Estrogen-regulated CD200 inhibits macrophage phagocytosis in endometriosis. J Reprod Immunol. 2020; 138:103090. https://doi.org/10.1016/j.jri.2020.103090. [PubMed].
75. Zhu XL, Zeng YF, Guan J, Li YF, Deng YJ, Bian XW, Ding YQ, Liang L. FMNL2 is a positive regulator of cell motility and metastasis in colorectal carcinoma. J Pathol. 2011; 224:377–88. https://doi.org/10.1002/path.2871. [PubMed].
76. Ge N, Mao C, Yang Q, Han B, Wang Y, Xu L, Yang X, Jiao W, Li C. Single nucleotide polymorphism rs3746444 in miR-499a affects susceptibility to non-small cell lung carcinoma by regulating the expression of CD200. Int J Mol Med. 2019; 43:2221–29. https://doi.org/10.3892/ijmm.2019.4124. [PubMed].