
Introduction
Cancer stem cells (CSCs), or tumor-initiating cells (TICs), are thought to be resistant to conventional anti-cancer therapies, and have been implicated in treatment failure, tumor recurrence and distant metastasis [1, 2]. Thus, residual treatment-resistant cancer stem cells are believed to be responsible for poor clinical outcomes in most cancer types [2-4]. Since CSCs are relatively rare and elusive, very little is known about them, especially regarding their physiology and metabolic phenotype.
Consistent with the idea that CSCs are resistant to cellular stress, they are able to undergo anchorage-independent growth in low-attachment plates, allowing the formation of 3D spheroids with the properties of cancer stem cells and/or progenitor cells [5]. Under these suspension conditions, most epithelial cancer cells undergo a specialized form of cell death/apoptosis, termed anoikis. Importantly, each of these 3D spheroids is formed from the anchorage-independent clonal expansion of a single cancer stem cell, and not from the self-aggregation of existing cancer cells [5]. Therefore, the preparation of 3D spheroid cultures provides a functional assay to enrich for a population of cells with an epithelial stem cell-like phenotype. In this regard, the behavior of 3D spheroids (also known as mammosphere cultures) prepared from primary breast cancer cells or breast cancer epithelial cell lines are the most well-characterized.
Here, we isolated large numbers of mammospheres from two independent ER-positive breast cancer cell lines, namely MCF7 and T47D cells, in an attempt to better understand their phenotypic behavior at a molecular level. The large-scale preparation of mammospheres allowed us to then perform unbiased label-free proteomics analysis, in an attempt to understand the proteome that is characteristic of cancer stem cells. Interestingly, based on this initial analysis, we noticed that mammospheres dramatically overexpress mitochondrial-related proteins. Thus, we focused on the mitochondrial proteins that were upregulated, relative to cells cultured as epithelial monolayers, in parallel. Based on this analysis, we speculate that CSCs become resistant to stress by fortifying their capacity to produce ATP by oxidative mitochondrial metabolism. Treatment with the MCT1/2 inhibitor (AR-C155858) validated that mammosphere formation is dependent on the uptake of specific mitochondrial fuels, such as L-lactate and ketone bodies.
Importantly, over the last several decades, significant progress has been made in understanding the critical role of cellular metabolism in tumor initiation, progression and metastasis, including studies related to ROS production, oxidative stress, glycolysis, glutamine metabolism and oxidative mitochondrial metabolism [6-13]. However, most of these studies have focused on “bulk” cancer cells, but not on cancer stem cells or the progenitor cell population. Thus, there is a great need to understand tumor metabolism in the specific context of “stemness”, to identify a metabolic “Achilles’ Heel” to eradicate cancer stem cells. As such, our current findings provide an unbiased systematic approach for identifying new metabolic targets in cancer stem cells, using quantitative proteomics analysis, and a strong rationale for therapeutically targeting L-lactate, ketone bodies and mitochondrial metabolism in the cancer stem cell population.
Results
Proteomic analysis of MCF7 mammospheres
Monolayer and mammosphere cultures of MCF7 cells, an ER-positive cell line, were subjected to quantitative label-free proteomics analysis. Greater than 500 proteins were found to be upregulated or downregulated. For simplicity, we focused on the proteins that were significantly upregulated in mammospheres, relative to mononlayer cell cultures (p < 0.05). Immediately, we noticed that several mitochondrial proteins were highly upregulated in mammospheres, so we restricted our analysis to mitochondrial proteins and key related metabolic enzymes.
Table 1 shows a non-redundant list of 62 mitochondrial-related proteins that were selectively upregulated in MCF7 mammospheres. Only proteins with a fold increase of ~2 or greater were selected for this analysis. Note that 9 mitochondrial proteins were infinitely upregulated, as compared with monolayer cultures. A functional analysis of this list revealed that 12 proteins were specifically related to beta-oxidation and ketone metabolism/re-utilization (HSD17B10, BDH1, ACAT1, ACADVL, ACACA, ACLY, HADHB, SUCLG2, ACAD9, HADHA, ECHS1, ACADSB). Also, 8 proteins involved in mitochondrial biogenesis were dramatically upregulated (HSPA9, TIMM8A, GFM1, MRPL45, MRPL17, HSPD1(HSP60), TSFM, TUFM). In addition, many proteins related to electron transport (NDUFB10, COX6B1, PMPCA, COX5B, SDHA, UQCRC1), ATP synthesis (ATP5B, ATPIF1, ATP5A1, ATP5F1, ATP5H, ATP5O), ADP/ATP exchange/transport (SLC25A5), CoQ synthesis (COQ9), or ROS production (GPD2) were also increased. Finally, two proteins involved in the suppression of glycolysis, autophagy and mitophagy were also significantly increased (SOGA1, LRPPRC). Thus, increased mitochondrial biogenesis and decreased mitochondrial degradation could provide a novel mechanism for the overall accumulation of mitochondrial mass in cancer stem cells.


![]()
Proteomic analysis of T47D mammospheres
For comparison purposes, we also performed unbiased label-free proteomic analysis on a second independent ER-positive breast cancer cell line, namely T47D cells.
Our results are summarized in Table 2. Note that 49 mitochondrial-related proteins were specifically over-expressed in T47D mammospheres, as compared with T47D monolayer cultures processed in parallel. Remarkably, 41 of these proteins overlapped with the proteins that were upregulated in MCF7 mammospheres (41/49 = 84% overlap). See the Venn diagram presented in Figure 1. Therefore, many of the same biological processes would be expected to be activated or enhanced. Thus, beta-oxidation, ketone re-utilization, mitochondrial biogenesis, and ROS production, with decreased mitochondrial degradation and reduced autophagy, are likely to be key biological features of both MCF7 and T47D mammospheres. Interestingly, CHCHD2 and CPOX were infinitely upregulated in both MCF7 and T47D data sets.


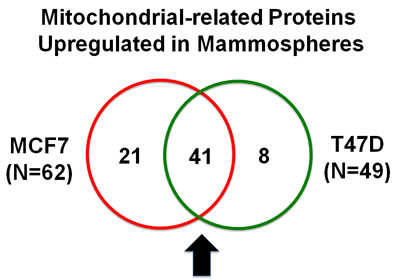
Figure 1: Venn diagram highlighting the conserved upregulation of mitochondrial related proteins in both MCF7 and T47D mammospheres. Note that >40 mitochondrial-related proteins were commonly upregulated in both data sets.
Functional effects of MCT1/2 inhibition on mammosphere formation
Next, to functionally validate the hypothesis that mammosphere formation may require ketone re-utilization and oxidative mitochondrial metabolism, we used a highly specific inhibitor (AR-C155858) of the relevant monocarboxylate transporters, namely MCT1/2 [14, 15]. MCT1/2 normally function as specific transporters for the uptake of ketone and L-lactate [16]. AR-C155858 effectively inhibits MCT1/2 function, and blocks the cellular uptake of both ketone bodies and L-lactate [14, 15].
Figure 2 shows the effects of increasing concentrations of AR-C155858 on mammosphere formation, using an ER-positive cell line (MCF7). Importantly, the MCT1/2 inhibitor AR-C155858 significantly reduces mammosphere formation, with an IC-50 of ~ 1µM.
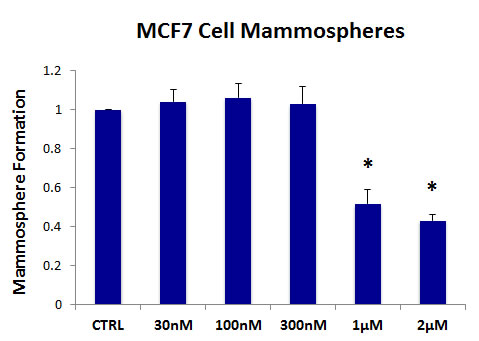
Figure 2: The MCT1/2 inhibitor AR-C155858 significantly reduces mammosphere formation in MCF7 cells. Increasing concentrations of AR-C155858 inhibit mammosphere formation, using an ER-positive cell line (MCF7). Importantly, AR-C155858 significantly reduces mammosphere formation, with an IC-50 of ~ 1 µM. The vehicle-alone control was normalized to one. (*)p <6.0E-06.
As breast cancer stem cells are thought to be ER-negative, we also evaluated the effects of AR-C155858 on an ER-negative cell line, namely MDA-MB-231 cells. Figure 3 shows that the MCT1/2 inhibitor AR-C155858 also effectively reduces mammosphere formation in this cellular context, with an IC-50 of ~ 1-2 µM. Therefore, MCT1/2 inhibition may be a new general therapeutic strategy that could be utilized to treat several different epithelial subtypes of human breast cancers.
Thus, 3D spheroid cultures appear to require oxidative mitochondrial metabolism, related to the re-use of monocarboxylic acids (ketones or L-lactate), for the proliferative anchorage-independent expansion of cancer stem cells.
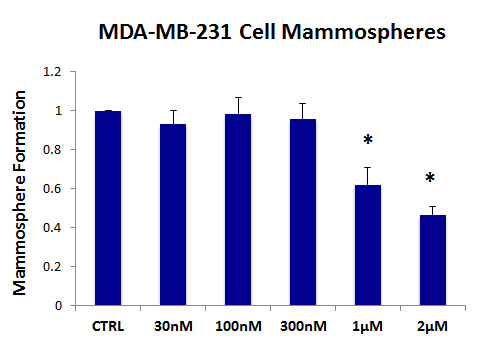
Figure 3: The MCT1/2 inhibitor AR-C155858 significantly reduces mammosphere formation in MDA-MB-231 cells. Note that AR-C155858 also effectively reduces mammosphere formation in this cellular context, with an IC-50 of ~ 1-2 µM. The vehicle-alone control was normalized to one. (*)p <0.0005.
Functional effects of inhibition of the mitochondrial ATP synthase (complex V) on mammosphere formation
Finally, to further validate that mammosphere formation is strictly dependent on oxidative mitochondrial metabolism, we used a well-established investigational compound that potently inhibits the mitochondrial ATP synthase (complex V), namely oligomycin A. Importantly, five protein components of the mitochondrial ATP synthesis were highly up-regulated in MCF7 mammospheres (ATP5B, ATP5A1, ATP5F1, ATP5H, ATP5O).
Figure 4A,B shows the effects of increasing concentrations of oligomycin A on mammosphere formation. Note that oligomycin A significantly reduces mammosphere formation in MCF7 cells, with an IC-50 of ~ 100 nM. Oligomycin A also significantly inhibited mammosphere formation in MDA-MB-231 cells, but with less potency, with an IC-50 between 5-10 µM. As such, oligomycin A was 50-100 times less potent in MDA-MB-231 cells, as compared with MCF7 mammospheres.
Therefore, MCT1/2 inhibition may be a more effective strategy for eradicating cancer stem cells in multiple breast cancer types, rather than targeting the mitochondrial ATP synthase.
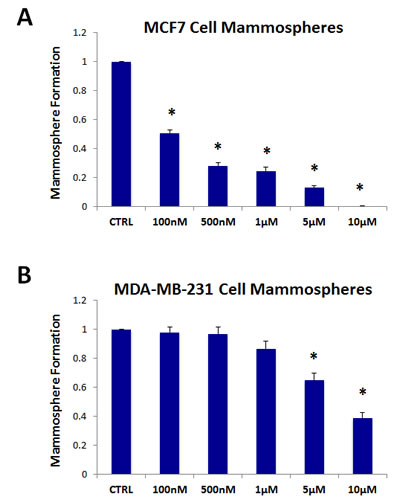
Figure 4: The mitochondrial ATP synthase inhibitor oligomycin A significantly reduces mammosphere formation in both MCF7 and MDA-MB-231 cells. Note that oligomycin A effectively reduces mammosphere formation, with an IC-50 of ~100 nM in MCF7 cells (panel A) and ~5-10 µM in MDA-MB-231 cells (panel B). Thus, oligomycin A was nearly 50-100 times more potent in MCF7 cells. The vehicle-alone control was normalized to one. (*)p <3.2E-06.
Clinical relevance of mitochondrial targets in human breast cancers
To determine the potential clinical relevance of our findings, we next assessed whether the “metabolic targets” that we identified in mammospheres were also transcriptionally upregulated in human breast cancer cells in vivo.
For this purpose, we employed a published clinical data set of N=28 breast cancer patients in which their tumor samples were subjected to laser-capture micro-dissection, to physically separate epithelial cancer cells from their adjacent tumor stroma [17]. Table 3 presents a summary of these findings. Overall, 39 of the “metabolic targets” that we identified in mammospheres (Tables 1 & 2) were also transcriptionally elevated in human breast cancer cells in vivo (Table 3) and the majority of these targets were upregulated in both MCF7 and T47D mammospheres (21 out of 39, ~54% ).
In light of these findings, the new “metabolic targets” that we identified in mammospheres are especially clinically relevant, for improving both the diagnosis and treatment of human breast cancers.


Discussion
Previous immuno-histochemical studies have shown that markers of cell proliferation (Ki67) and mitochondrial mass/function (TOMM20 and Complex IV activity) specifically co-localize to the basal stem cell layer in human oral mucosal tissue [18]. Interestingly, in this context, MCT1 was the most specific marker of the basal stem cell layer, suggesting that normal stem cells may use L-lactate and ketone bodies to fuel oxidative mitochondrial metabolism and stem cell proliferation [18]. Similarly, Ki67, mitochondrial markers and MCT1 also co-localized in aggressive head and neck tumor cells, consistent with the idea that amplification of mitochondrial metabolism may contribute to human tumor formation and cancer progression [18].
Numerous studies have also implicated ketone bodies and L-lactate metabolism in cancer biology and/or “stemness” in cancer cells. For example, treatment with mitochondrial fuels (such as L-lactate and 3-hydroxy-butyrate) is sufficient to stimulate mitochondrial biogenesis in MCF7 cells and dramatically increases the levels of gene transcripts normally expressed in embryonic, neuronal and hematopoietic stem cells [19]. Importantly, the transcriptional profiles of lactate- or ketone-treated MCF7 cells effectively predicted poor clinical outcome (tumor recurrence and metastasis) in ER-positive breast cancer patients [19]. Consistent with these findings, Cuyas et al. recently showed that cancer stem cells created by silencing of E-cadherin expression prefer to use L-lactate and ketone bodies as mitochondrial fuels [20].
Here, using unbiased label-free proteomics analysis, we show that mammospheres (a population of cells enriched in cancer stem cells and progenitor cells) functionally overexpress numerous mitochondrial proteins, related to mitochondrial biogenesis, electron transport, OXPHOS, ATP synthesis, as well as beta-oxidation and ketone re-utilization. The potential clinical relevance of these targets was further validated using a previously published data set of human breast cancer samples (N=28 patients), that were subjected to laser-capture microdissection, to separate the epithelial tumor cells from the adjacent tumor stroma [17]. Thus, these novel mitochondrial-based targets may reveal a metabolic “Achilles’ Heel” to allow the eradication of cancer stem cells.
In accordance with this idea, we demonstrate that therapeutic targeting of MCT1/2 in cancer stem cells may be a viable strategy, via inhibiting the uptake of necessary key mitochondrial fuels (ketone bodies and L-lactate), that may be required for anchorage-independent growth, as well as cancer stem cell proliferative expansion and survival. Further validation was also provided by experiments with oligomycin A, a well-established inhibitor of the mitochondrial ATP synthase (complex V).
Interestingly, here we observed that the mitochondrial protein CHCHD2 was infinitely upregulated in both MCF7 and T47D mammospheres (Tables 1 and 2), and was also the most highly transcriptionally upregulated protein in the context of human breast cancer cells in vivo (Table 3). Thus, future studies may be warranted on the specific role of CHCHD2 in cancer stem cell metabolism. Currently, very little is known about CHCHD2. However, CHCHD2 has been previously implicated functionally in the response to hypoxia and in the transcriptional upregulation of members of the OXPHOS complexes, as well as a positive regulator of cell migration [21-23].
In conclusion, based on our quantitative proteomics analysis and functional validation studies using mammosphere cultures, we propose that mitochondria are new therapeutic targets for eradicating cancer stem cells, to prevent tumor recurrence, metastasis and poor clinical outcome in breast cancer patients.
Materials and Methods
Materials
Breast cancer cell lines (MCF7, T47D and MDA-MB-231 cells) were purchased from the ATCC. AR-C155858 was obtained commercially from MedChem Express (UK). Gibco-brand cell culture media (DMEM/F12) was purchased from Life Technologies. Oligomycin A was obtained from Sigma-Aldrich.
Mammosphere Culture
A single cell suspension was prepared using enzymatic (1x Trypsin-EDTA, Sigma Aldrich, #T3924), and manual disaggregation (25 gauge needle) to create a single cell suspension [5]. Cells were plated at a density of 500 cells/cm2 in mammosphere medium (DMEM-F12/B27/20ng/ml EGF/PenStrep) in non-adherent conditions, in culture dishes coated with (2-hydroxyethylmethacrylate) (poly-HEMA, Sigma, #P3932). Cells were grown for 5 days and maintained in a humidified incubator at 37°C at an atmospheric pressure in 5% (v/v) carbon dioxide/air. After 5 days for culture, spheres >50 µm were counted using an eye piece graticule, and the percentage of cells plated which formed spheres was calculated and is referred to as percentage mammosphere formation, and was normalized to one (1 = 100 %MSF). For proteomic analysis, mammospheres were collected by centrifugation at 800 rpm for 10 minutes.
Label-free Quantitative Proteomics analysis
Cell lysates were prepared for trypsin digestion by sequential reduction of disulphide bonds with TCEP and alkylation with MMTS [24]. Then, the peptides were extracted and prepared for LC-MS/MS. All LC-MS/MS analyses were performed on an LTQ Orbitrap XL mass spectrometer (Thermo Scientific, San Jose, CA) coupled to an Ultimate 3000 RSLCnano system (Thermo Scientific, formerly Dionex, The Netherlands). Xcalibur raw data files acquired on the LTQ-Orbitrap XL were directly imported into Progenesis LCMS software (Waters Corp., Milford, MA, formerly Non-linear dynamics, Newcastle upon Tyne, UK) for peak detection and alignment. Data were analyzed using the Mascot search engine. Five replicates were analyzed for each sample type (N = 5). Statistical analyses were performed using ANOVA and only fold-changes in proteins with a p-value less than 0.05 were considered significant.
A more detailed proteomics protocol is provided as Supplementary Information.
Data Mining
To firmly establish the clinical relevance of our results from the quantitative proteomics analysis of mammosheres, we re-analyzed the transcriptional profiles of epithelial breast cancer cells and adjacent tumor stromal cells that were physically separated by laser-capture microdissection (from N=28 human breast cancer patients) [17].
Acknowledgements
We thank the University of Manchester for providing start-up funds that contributed to the success of this study. RL, HH, MPL, and FS were supported, in part, by funding from the European Union (ERC Advanced Grant), Breakthrough Breast Cancer, and the Manchester Cancer Research Centre (MCRC). DLS was core-funded by CRUK.
References
1. Zhang M and Rosen JM. Stem cells in the etiology and treatment of cancer. Curr Opin Genet Dev. 2006; 16(1):60-64.
2. Chandler JM and Lagasse E. Cancerous stem cells: deviant stem cells with cancer-causing misbehavior. Stem Cell Res Ther. 2010; 1(2):13.
3. Scopelliti A, Cammareri P, Catalano V, Saladino V, Todaro M and Stassi G. Therapeutic implications of Cancer Initiating Cells. Expert Opin Biol Ther. 2009; 9(8):1005-1016.
4. Duggal R, Minev B, Geissinger U, Wang H, Chen NG, Koka PS and Szalay AA. Biotherapeutic approaches to target cancer stem cells. J Stem Cells. 2013; 8(3-4):135-149.
5. Shaw FL, Harrison H, Spence K, Ablett MP, Simoes BM, Farnie G and Clarke RB. A detailed mammosphere assay protocol for the quantification of breast stem cell activity. J Mammary Gland Biol Neoplasia. 2012; 17(2):111-117.
6. Borodkina A, Shatrova A, Abushik P, Nikolsky N and Burova E. Interaction between ROS dependent DNA damage, mitochondria and p38 MAPK underlies senescence of human adult stem cells. Aging (Albany NY). 2014; 6(6):481-495.
7. Marcinek DJ and Siegel MP. Targeting redox biology to reverse mitochondrial dysfunction. Aging (Albany NY). 2013; 5(8):588-589.
8. Yoon MJ, Lee AR, Jeong SA, Kim YS, Kim JY, Kwon YJ and Choi KS. Release of Ca2+ from the endoplasmic reticulum and its subsequent influx into mitochondria trigger celastrol-induced paraptosis in cancer cells. Oncotarget. 2014; 5(16):6816-6831.
9. Corazao-Rozas P, Guerreschi P, Jendoubi M, Andre F, Jonneaux A, Scalbert C, Garcon G, Malet-Martino M, Balayssac S, Rocchi S, Savina A, Formstecher P, Mortier L, Kluza J and Marchetti P. Mitochondrial oxidative stress is the Achille’s heel of melanoma cells resistant to Braf-mutant inhibitor. Oncotarget. 2013; 4(11):1986-1998.
10. Hall A, Meyle KD, Lange MK, Klima M, Sanderhoff M, Dahl C, Abildgaard C, Thorup K, Moghimi SM, Jensen PB, Bartek J, Guldberg P and Christensen C. Dysfunctional oxidative phosphorylation makes malignant melanoma cells addicted to glycolysis driven by the (V600E)BRAF oncogene. Oncotarget. 2013; 4(4):584-599.
11. Muppani N, Nyman U and Joseph B. TAp73alpha protects small cell lung carcinoma cells from caspase-2 induced mitochondrial mediated apoptotic cell death. Oncotarget. 2011; 2(12):1145-1154.
12. Francipane MG and Lagasse E. Selective targeting of human colon cancer stem-like cells by the mTOR inhibitor Torin-1. Oncotarget. 2013; 4(11):1948-1962.
13. Cufi S, Corominas-Faja B, Vazquez-Martin A, Oliveras-Ferraros C, Dorca J, Bosch-Barrera J, Martin-Castillo B and Menendez JA. Metformin-induced preferential killing of breast cancer initiating CD44+CD24-/low cells is sufficient to overcome primary resistance to trastuzumab in HER2+ human breast cancer xenografts. Oncotarget. 2012; 3(4):395-398.
14. Ovens MJ, Manoharan C, Wilson MC, Murray CM and Halestrap AP. The inhibition of monocarboxylate transporter 2 (MCT2) by AR-C155858 is modulated by the associated ancillary protein. Biochem J. 2010; 431(2):217-225.
15. Ovens MJ, Davies AJ, Wilson MC, Murray CM and Halestrap AP. AR-C155858 is a potent inhibitor of monocarboxylate transporters MCT1 and MCT2 that binds to an intracellular site involving transmembrane helices 7-10. Biochem J. 2010; 425(3):523-530.
16. Draoui N, Schicke O, Seront E, Bouzin C, Sonveaux P, Riant O and Feron O. Antitumor activity of 7-aminocarboxycoumarin derivatives, a new class of potent inhibitors of lactate influx but not efflux. Mol Cancer Ther. 2014; 13(6):1410-1418.
17. Casey T, Bond J, Tighe S, Hunter T, Lintault L, Patel O, Eneman J, Crocker A, White J, Tessitore J, Stanley M, Harlow S, Weaver D, Muss H and Plaut K. Molecular signatures suggest a major role for stromal cells in development of invasive breast cancer. Breast Cancer Res Treat. 2009; 114(1):47-62.
18. Curry JM, Tuluc M, Whitaker-Menezes D, Ames JA, Anantharaman A, Butera A, Leiby B, Cognetti DM, Sotgia F, Lisanti MP and Martinez-Outschoorn UE. Cancer metabolism, stemness and tumor recurrence: MCT1 and MCT4 are functional biomarkers of metabolic symbiosis in head and neck cancer. Cell Cycle. 2013; 12(9):1371-1384.
19. Martinez-Outschoorn UE, Prisco M, Ertel A, Tsirigos A, Lin Z, Pavlides S, Wang C, Flomenberg N, Knudsen ES, Howell A, Pestell RG, Sotgia F and Lisanti MP. Ketones and lactate increase cancer cell “stemness,” driving recurrence, metastasis and poor clinical outcome in breast cancer: achieving personalized medicine via Metabolo-Genomics. Cell Cycle. 2011; 10(8):1271-1286.
20. Cuyas E, Corominas-Faja B and Menendez JA. The nutritional phenome of EMT-induced cancer stem-like cells. Oncotarget. 2014; 5(12):3970-3982.
21. Aras S, Bai M, Lee I, Springett R, Huttemann M and Grossman LI. MNRR1 (formerly CHCHD2) is a bi-organellar regulator of mitochondrial metabolism. Mitochondrion. 2014.
22. Aras S, Pak O, Sommer N, Finley R, Jr., Huttemann M, Weissmann N and Grossman LI. Oxygen-dependent expression of cytochrome c oxidase subunit 4-2 gene expression is mediated by transcription factors RBPJ, CXXC5 and CHCHD2. Nucleic Acids Res. 2013; 41(4):2255-2266.
23. Seo M, Lee WH and Suk K. Identification of novel cell migration-promoting genes by a functional genetic screen. Faseb J. 2010; 24(2):464-478.
24. Holland M, Castro FV, Alexander S, Smith D, Liu J, Walker M, Bitton D, Mulryan K, Ashton G, Blaylock M, Bagley S, Connolly Y, Bridgeman J, Miller C, Krishnan S, Dempsey C, et al. RAC2, AEP, and ICAM1 expression are associated with CNS disease in a mouse model of pre-B childhood acute lymphoblastic leukemia. Blood. 2011; 118(3):638-649.