
Introduction
Primary mediastinal large B-cell lymphoma (PMBL) is a rare form of non-Hodgkin lymphoma representing < 5% of mature B-cell NHL (B-NHL) [1, 2]. The immunophenotypic features and molecular characteristics distinguish PMBL from other types of aggressive B-NHL and large B cell lymphoma [3, 4]. PMBL shares one third of its genes with Hodgkin lymphoma [3]. PMBL is classified as midway in the biologic spectrum between diffuse large B-cell lymphoma (DLBCL) and classical Hodgkin lymphoma [3] with overexpression of genes in signaling pathways of NF-kB and Janus kinase (JAK)/signal transducer and transcription (STAT), high expression of CD20, downregulation of major histocompatibility class I and II molecules and overexpression of programmed death ligands [2, 5–9].
Obinutuzumab is a glycoengineered humanized monoclonal antibody (mAb) recognizing a unique CD20 type II epitope [10] with significant antibody dependent cellular cytotoxicity (ADCC) enhanced as compared to rituximab in-vitro and in-vivo [11–14]. The anti-tumor effects of obinutuzumab alone or in combination with other agents were further investigated in clinical trials. The safety and efficacy of obinutuzumab was compared with rituximab in relapsed indolent lymphoma in the randomized phase II trial (GAUSS) (NCT00576758) [15]. Among patients with follicular lymphoma (FL), obinutuzumab demonstrated a higher overall response rate than rituximab (44.6% v 33.3%; P = .08) but with no difference in progression-free survival (PFS) between the two arms [15]. The phase III GALLIUM trial (NCT01332968) and GADOLIN trial (NCT01059630) were conducted to treat previously untreated FL patients or patients with rituximab-refractory indolent non-Hodgkin lymphoma utilizing obinutuzumab combined with chemotherapy [16, 17]. Obinutuzumab-based therapy significantly reduced the risk of progression or death and prolonged overall survival (OS) as compared to rituximab-based therapy or chemotherapy [16, 17]. Obinutuzumab plus chlorambucil prolonged the OS or PFS and resulted in higher rates of complete response in patients with chronic lymphocytic leukemia (CLL) or coexisting conditions as compared to chlorambucil alone, or rituximab plus chlorambucil, respectively in the CLL11 clinical trial (NCT01010061b) [18]. Furthermore, the phase III iLLUMINATE trial (NCT02264574) demonstrated that obinutuzumab plus Ibrutinib is an efficacious combination therapy for previously untreated patients with CLL or small lymphocytic lymphoma [19]. Based on these exciting results, obinutuzumab in combination with chemotherapy has been approved for the treatment of untreated and rituximab refractory FL [16, 17] and CLL [19]. Unfortunately, the clinical results of obinutuzumab for patients with DLBCL were not promising. Obinutuzumab was not superior to rituximab when combined with chemotherapies in patients with DLBCL shown in the phase III GOYA trial (NCT01287741) and the GAINED trial (NCT01659099) [20–22]. Additionally, the pre-clinical and clinical efficacy of obinutuzumab compared to rituximab in patients with PMBL is currently unknown.
We hypothesize that obinutuzumab would be a superior anti-CD20 antibody in the treatment of PMBL by inducing targeted programmed cell death and enhancing immune cell mediated ADCC compared to rituximab. In this study, we report the in-vitro and in-vivo efficacy of obinutuzumab against PMBL cell lines and in human PMBL xenografted immunodeficient NOD scid gamma (NSG) mouse model compared to rituximab.
Results
Expression of CD20 mRNA and protein in obinutuzumab treated PMBL
CD20 mRNA and protein expression in Karpas-1106P were measured by real time quantitative reverse transcription polymerase chain reaction and immunoblotting prior to any anti-CD20 treatment. Karpas-1106P showed a significant increase in the expression of both CD20 mRNA and protein (Figure 1A and 1B) compared to Burkitt lymphoma (BL) (Raji) and Hodgkin lymphoma (HDLM-2) cell lines.
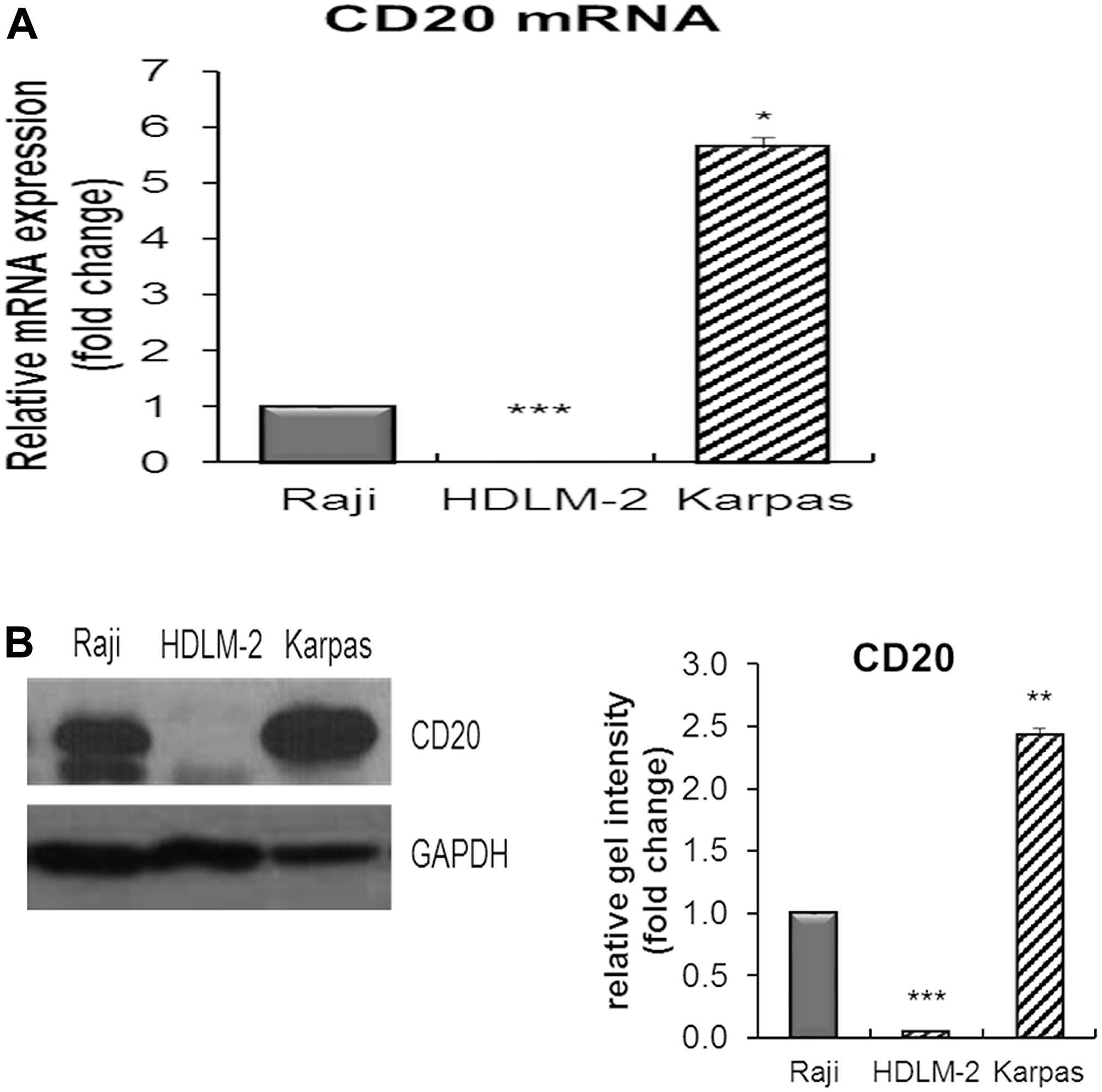
Figure 1: The expression of CD20 mRNA and protein in Karpas-1106 PMBL cells. (A) The CD20 mRNA, (B left) protein expression and (B right) its band intensity in Karpas-1106P PMBL cell line by qRT-PCR and immunoblotting compared to Raji (BL) and HDLM-2 (HL) cells as controls. Data are represented as the mean ± SD, *p < 0.01; **p < 0.001; ***p < 0.00005 (N = 3).
Significant decrease of cell viability in obinutuzumab treated PMBL
Karpas-1106P cells were treated with obinutuzumab, rituximab and IgG-isotype and viable cells were quantified by MTS assay. There was a significant decrease of viable cell number in the obinutuzumab-treated Karpas-1106P compared to control cells at each day regardless of the dose of obinutuzumab from 1–100 μg/ml (Figure 2A). Specifically, we observed significant reduction of viable cells with > 27% (p < 0.01), > 34% (p < 0.01), > 33% (p < 0.001) and > 35% (p < 0.01) in 1, 10, 20 and 100 μg/ml obinutuzumab treated Karpas-1106P cells for 24 hours and significantly reduction with > 27% (p < 0.001), > 31% (p < 0.01), > 37% (p < 0.01) and > 40% (p < 0.01) at 48 hours, respectively. In addition, we also observed significant reduction of viable Karpas-1106P cells by obinutuzumab at 1 (> 27%, p < 0.05), 10 (< 26%, p < 0.05), 20 (< 31%, p < 0.01), and 100 μg/ml (< 30%, 0.05) for 72 hours treatment compared to controls.
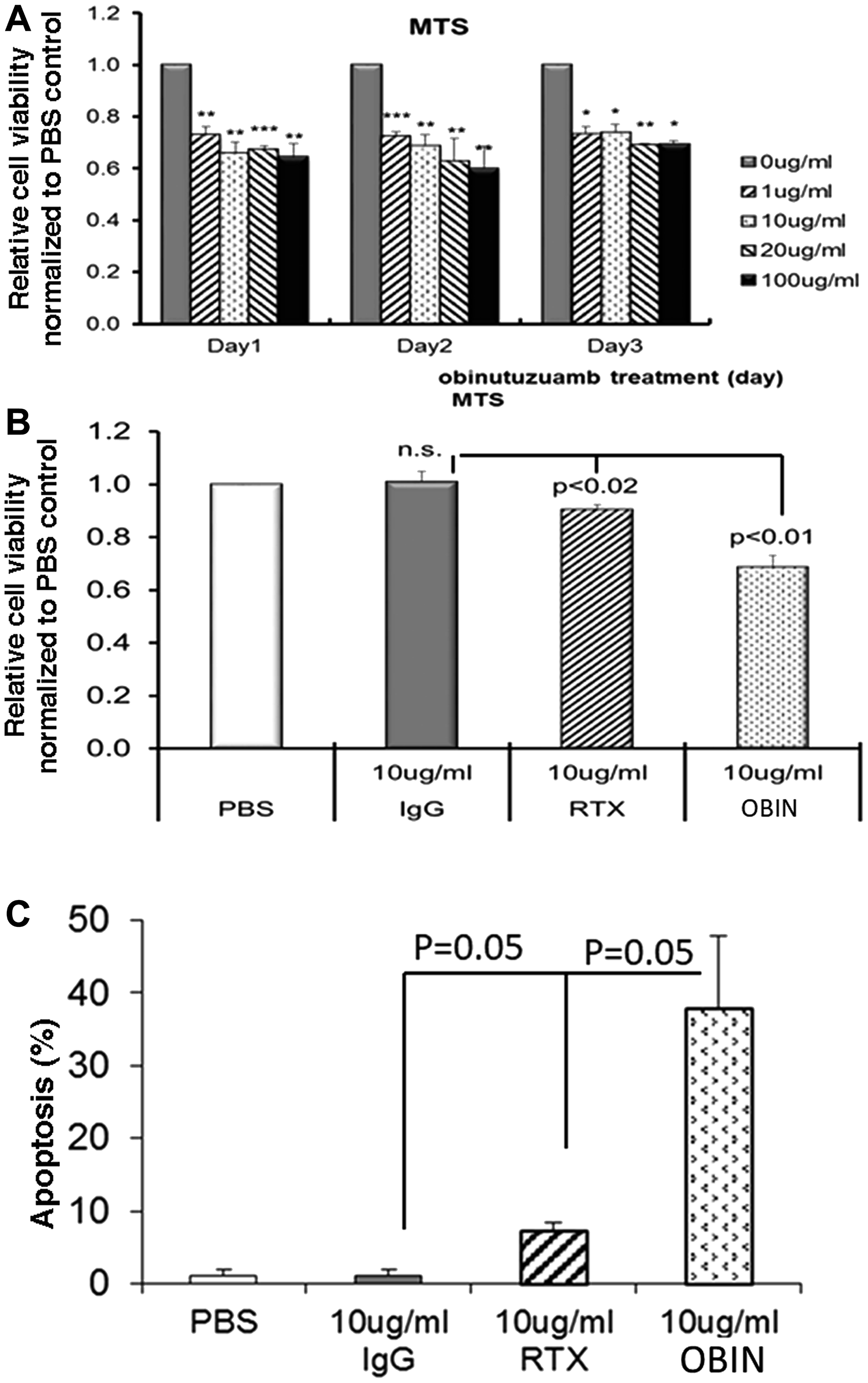
Figure 2: Cell proliferation and apoptosis activity in obinutuzumab treated Karpas-1106P PMBL cells. (A) Cells were plated (1 × 105) into 48 well plates and cell proliferation was measured every 48 hours via MTS assay following obinutuzumab treatment (0, 1, 10, 20 and 100 μg/ml) in Karpas-1106P PMBL cells. (B) Significant inhibition in cell proliferation in obinutuzumab (p < 0.01) and rituximab (p < 0.02) treated Karpas-1106P cells compared to PBS control. Data are represented as the mean ± SD, *p < 0.05; **p < 0.01; ***p < 0.001. (C) Apoptosis assays were performed by flow cytometric analysis using FITC-Annexin V/PI staining in obinutuzumab (10 ug/ml) treated Karpas-1106P PMBL cells compared to PBS control. Significant increase of apoptosis in obinutuzumab (p = 0.05) and rituximab (p = 0.05) treated Karpas-1106P cells compared to PBS control. Data are represented as the mean ± SD of triplicates (paired t test) (N = 3).
We further compared the cell viability of PMBL cells treated with obinutuzumab vs. rituximab. We found that the viability of Karpas-1106P cells was significantly reduced in the samples incubated with Obinutuzumab (10 μg/ml) compared to the samples incubated with rituximab (10 μg/ml) (viability reduction: 29.5% ± 4.60 vs. 4.4% ± 1.29, p < 0.01) at 48 hours (Figure 2B).
Significant increase in obinutuzumab-induced cell death in PMBL
Next, we compared the cell death in PMBL induced by 10 μg/ml obinutuzumab compared to 10 μg/ml rituximab at 48 hours treatment. Cell apoptosis was significantly enhanced after obinutuzumab treatment compared to rituximab at similar dose concentration (37.8% ± 10.096 vs. 7.16% ± 2.969 (p < 0.05) (Figure 2C).
Effect of ADCC by obinutuzumab vs. rituximab in combination with expanded natural killer (NK) cells against PMBL
We observed that obinutuzumab + expanded NK cells had significantly increased ADCC vs. rituximab + expanded NK with same antibody doses (10 μg/ml) against PMBL cells; 73.4 ± 3.4 vs 38.3 ± 9.2% (p = 0.0009) at 10:1 and 33.2 ± 11.1% vs. 11.6 ±.3.7% (p = 0.004) at E: T ratio 5:1 (n = 4) (Figure 3).
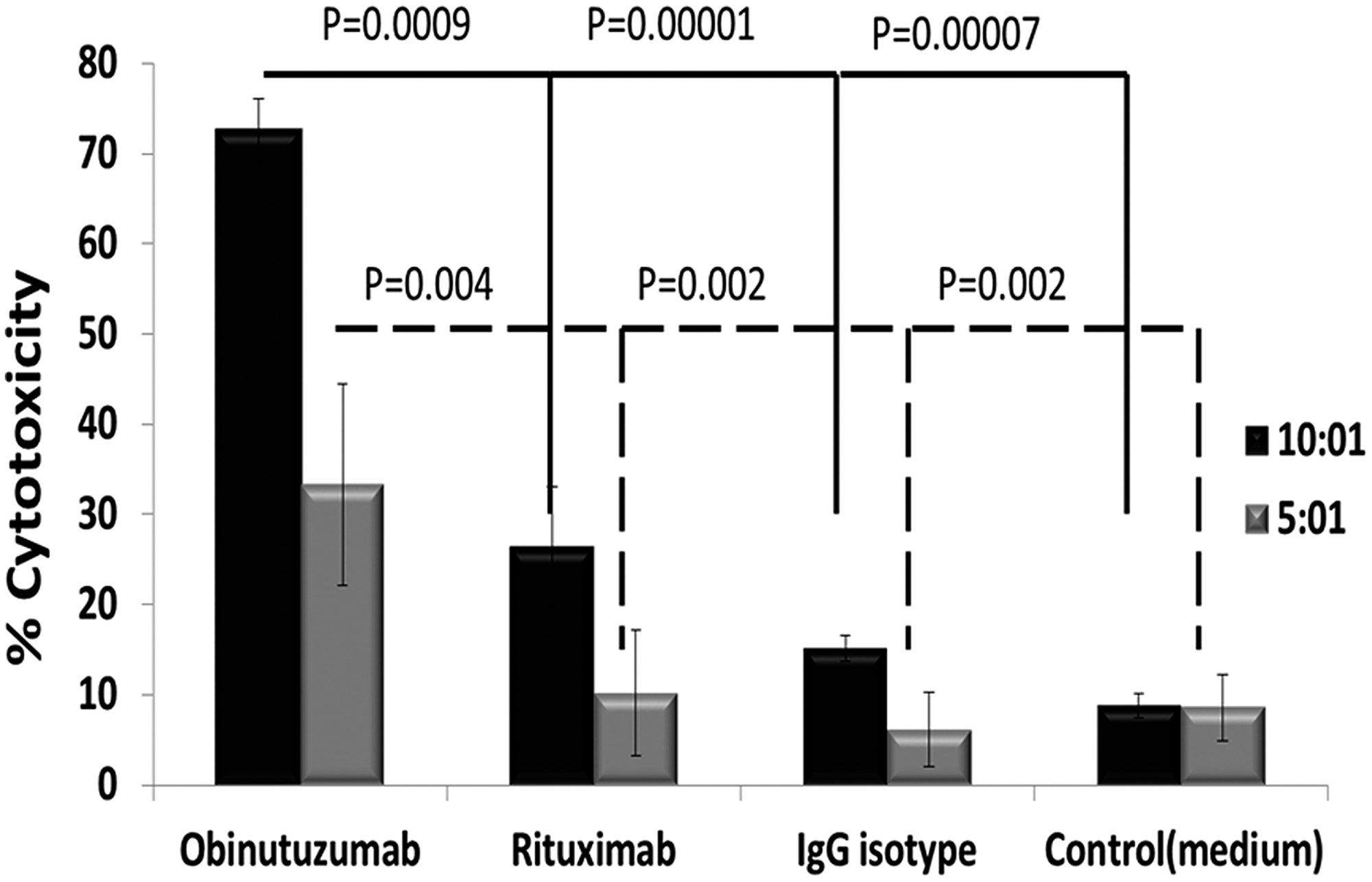
Figure 3: Obinutuzumab is significantly more effective in inducing NK cell mediated ADCC activity compared to rituximab. Karpas-1106P cells were treated with obinutuzumab, rituximab and IgG isotype control (10 μg/ml) and incubated with expanded and activated NK cells for additional 4 hours at 37°C at a 5:1 & 10:1 effector: targets ratio. Cell lysis was determined by manufacturer’s protocol using DELFIA cell cytotoxicity assays. Data shown are mean ± SD (n = 4) obinutuzumab vs rituximab, (p = 0.0009 and p = 0.004).
Obinutuzumab effects on the STAT6/NF-kB/PI3K/AKT pathway
Constitutive STAT6 activation is identified as a key feature of PMBL from other lymphoma and the NF-κB signaling pathway and the PI3K/AKT pathway are also reported to be activated in PMBL [2, 23]. Based on this known information, we firstly examined if obinutuzumab could potentially inhibit these pathways by decreasing the phosphorylations of some key members of these pathways. Karpas-1106P cells were treated with obinutuzumab (10 μg/ml) for 48 hours and the phosphorylation levels of AKT, STAT6 and IκBα were measured by Western blotting. There was a significant decrease of p-STAT6 (treated vs control: 0.725 ± 0.0065, p < 0.05) in the JAK/STAT pathway, p-AKT (treated vs control: 0.69 ± 0.0028, p < 0.005) in the PI3K/AKT pathway and p-IκBα (treated vs control: 0.706 ± 0.009, p < 0.005) in the NF-κB pathway in obinutuzumab treated Karpas 1106P cells compared to PBS controls (Figure 4A and 4B). There were no significant differences of total STAT6 and AKT proteins between the treated and the controls (treated vs control: 0.978 ± 0.0071, p > 0.05; 0.976 ± 0.011, p > 0.05; respectively) (Figure 4B). Total IκBα protein level was significantly reduced in obinutuzumab treated Karpas-1106P cells compared to PBS controls (treated vs control: 0.682 ± 0.0066, p < 0.005) (Figure 4B).
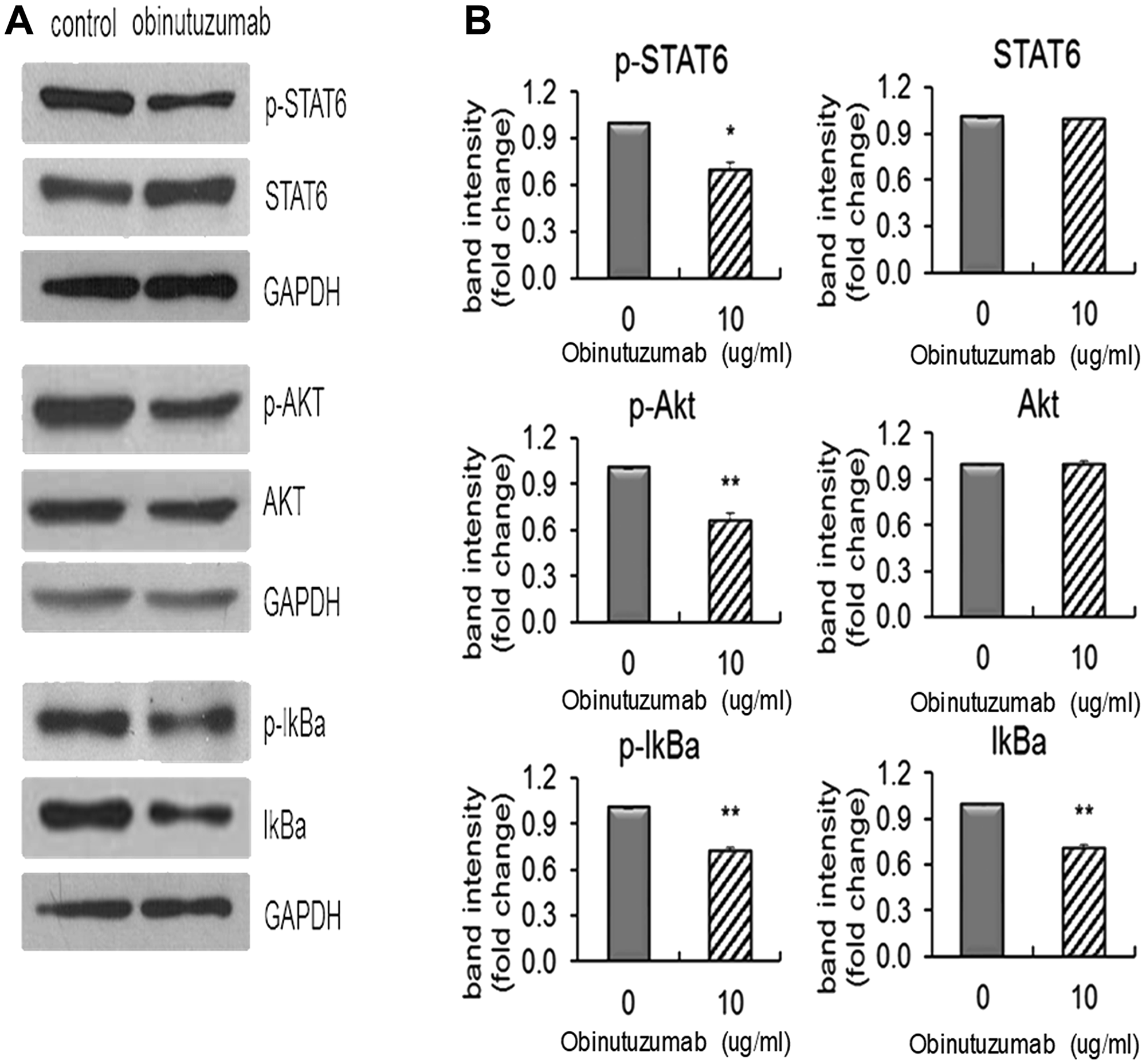
Figure 4: Obinutuzumab effect on the STAT6 and NFkB/PI3K pathways in PMBL cells. Karpas-1106P PMBL cells were treated with or without obinutuzumab (10 μg/ml) for 48 hours and analyzed by (A) western blotting, and (B) the intensity of the bands was quantified by Image J. Data are represented as the mean ± SD. *p < 0.05; **p < 0.005 (N = 3).
Gene expression changes in obinutuzumab treated PMBL cells
To identify other biological processes affected by obinutuzumab, we investigated gene expression changes in obinutuzumab treated PMBL cells. Genomics employing Illumina microarray technology in obinutuzumab treated Karpas-1106P PMBL cells were compared to PBS or IgG isotype control. A total of 65 differentially expressed genes (0.3% of all genes) were identified (>1.5-fold) from 21,381 human genes (Figure 5). Among the 65 filtered genes, 38 genes (58.5%) were up-regulated and 27 (41.5%) genes including ID3 and RAB6B were down-regulated. When we performed gene set enrichment analysis (GSEA) to identify the biological processes affected by the obinutuzumab treatment, we observed 19 gene sets were enriched to the obinutuzumab treatment while 8 were enriched to the IgG treatment (Supplementary Table 1) and 9 were enriched after PBS treatment (Supplementary Table 2), respectively. Gene sets enriched in the obinutuzumab treatment group included IL6-JAK-STAT6 signaling, Hedgehog signaling, interferon gamma signaling, apoptosis and MTORc1 signaling pathways (Supplementary Tables 1 and 2).
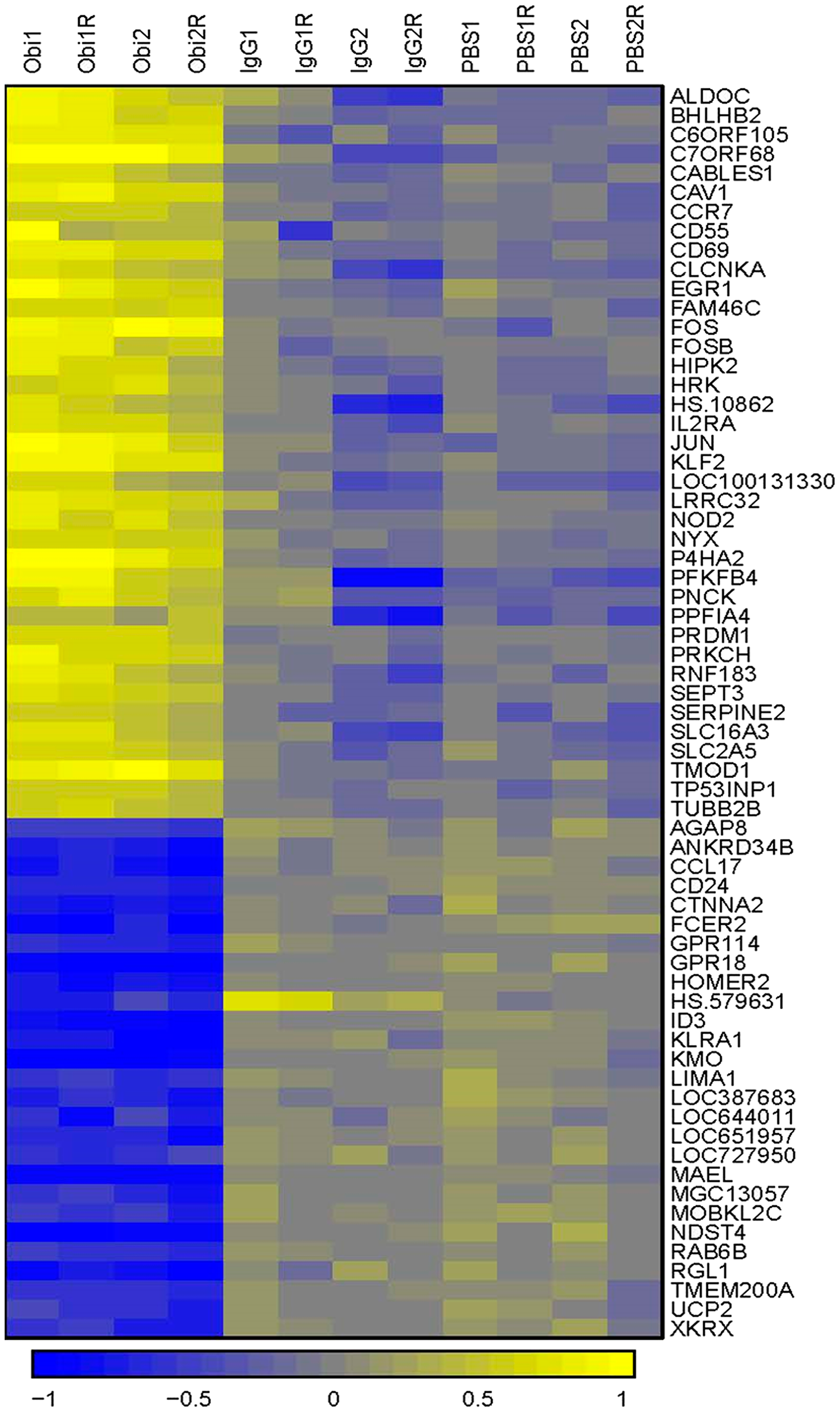
Figure 5: Hierarchical clustering comparing gene expression patterns between obinutuzumab and human IgG or PBS treated cells. A total of 65 differentially expressed genes were identified by gene expression profiling (> 1.5-fold) in obinutuzumab treated Karpas-1106P PMBL cells compared to PBS and IgG controls. Yellow represents increase expression and blue decreased expression, according to the scale. (N = 3).
Effect of obinutuzumab on survival in human PMBL xenografted NSG mice
We observed that obinutuzumab (30 mg/kg) treated Karpas-1106P PMBL cells xenografted NSG mice had significantly increased prolonged survival time with a median of 200 days compared to rituximab (143.5 days, p = 0.05), control (80 days, p = 0.003) and hu-IgG (91 days) (Figure 6A). The obinutuzumab treated group significantly reduced tumor burden compared to rituximab treated group (day 63, p = 0.03) (Figure 6B).
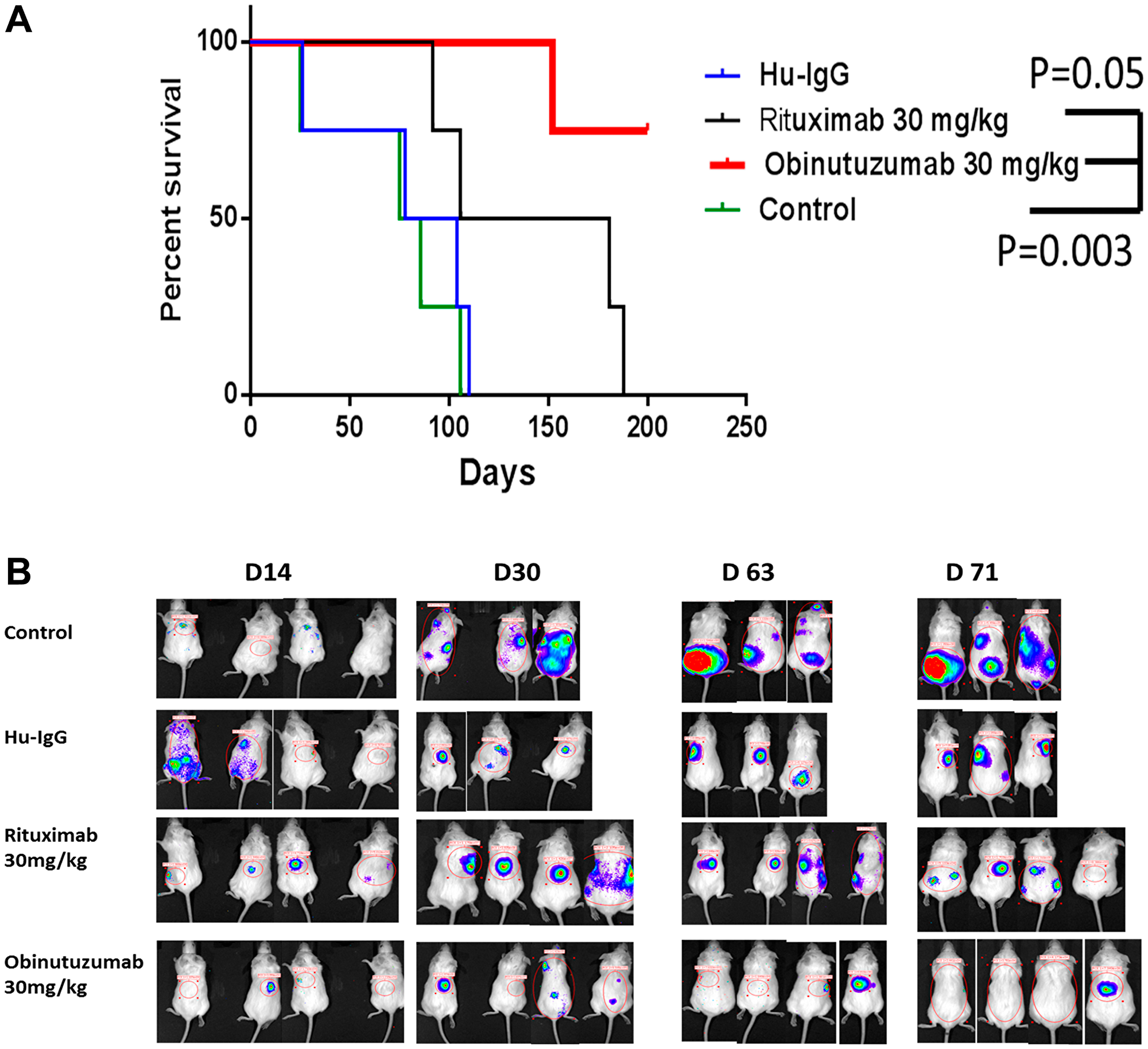
Figure 6: Effect of Obinutuzumab vs. Rituximab in PMBL NSG xenograft in-vivo. NSG mice were injected intravenously with human Karpas-1106P Luc+. Survival rates were analyzed following PBS, IgG, obinutuzumab, or rituximab treatment (30 mg/kg) once a week till 8 weeks. (A) The Karpas1106p-Luc xenografted NSG mice were observed until day 250. The Kaplan–Meier survival curves were generated following therapy initiation until mice death or mice sacrifice. Comparison of survival between groups is shown. Obinutuzumab significantly extended the survival of Karpas1106p-Luc mice compared to the group treated with rituximab (p < 0.05) or with PBS/IgG (p < 0.003). (B) Representative bioluminescence imaging of Karpas1106p-Luc xenografted NSG mice are shown at day 14, day 30, day 63 and day 71 after implantation of Karpas1106p-Luc cells in different treatment groups.
DISCUSSION
We have demonstrated significant in-vivo and in-vitro effects of obinutuzumab against PMBL. Specifically, we describe superior efficacy of obinutuzumab vs. rituximab (10 μg/ml) against Karpas 1106-P PMBL cells, including significantly decreased cell viability, increased cell death and enhanced ADCC. In addition, PMBL xenografted NSG mice treated with obinutuzumab showed a significant increase in survival compared with rituximab.
Obinutuzumab has been reported to have increased cytotoxicity causing B-cell depletion with greater activity against CD20+ tumor cells resulting in improved overall survival [11, 24]. Treatment in adults with relapsed indolent CD20+ B-NHL has demonstrated a higher overall response rate compared to rituximab [15]. As mentioned earlier, studies of obinutuzumab vs rituximab in patients with newly diagnosed or aggressive DLBCL in the upfront setting have not shown the same survival advantage [21, 22]. The reasons for the lack of advantage with obinutuzumab in DLBCL were unclear. This could be the result of differences in biologic and clinical profiles as well as differences in the immune microenvironment [22].
In contrast to the Type I anti-CD20 antibody rituximab, the tumor-killing mechanisms of obinutuzumab are reported to associate with enhanced direct cell death and ADCC, but reduced CDC [25]. Treatment of Karpas-1106P PMBL cells with obinutuzumab showed significant decreased cell viability irrespective of administered dose (Figure 2). This report is also consistent with increased cell death by increased ADCC in BL and pre B-cell acute lymphoblastic leukemia cell lines [11], and by increased activation of polymorphonuclear neutrophils resulting in increase in phagocytosis in chronic lymphocytic lymphoma cell lines [26]. Consistently with previous reports that obinutuzumab induced superior ADCC with effector cells using peripheral blood mononuclear cells [24], we found that obinutuzumab significantly enhanced ADCC mediated by ex vivo expanded NK cells (Figure 3). Constitutive activation of STAT6 pathway in PMBL has been previously reported by other investigators [27]. Differential regulation of AKT phosphorylation by obinutuzumab has also been reported in a 3D NHL model [28, 29]. STAT6, NF-κB and PI3K/AKT pathways were reported to be activated in PMBL [2, 23]. Our group recently demonstrated the difference in differential phosphorylation of 978 unique phosphoproteins between rituximab sensitive and resistant BL cell lines after obinutuzumab vs. rituximab treatments [12]. In this study, we found that obinutuzumab significantly reduces the phosphorylations of STAT6, AKT and IκBα in PMBL compared to controls (Figure 4). The treatment of a PMBL cell line with obinutuzumab differentially phosphorylated the PI3K/AKT pathway. This, in turn, affected phospho- IκBα and phospho-STAT6 signaling and NF-kB, JAK/STAT signaling pathways. Our results indicate that obinutuzumab inhibits PMBL proliferation partially by reducing the activation of the PI3K/AKT, NF-kB, JAK/STAT pathways.
Obinutuzumab enhances ADCC through several mechanisms. The prominent one is involved in enhancing the binding affinity to the FcγRIII receptor on immune effector cells such as NK cells through the post-translational glycoengineering [25, 30]. The second mechanism is that obinutuzumab overcomes the inhibition from inhibitory killer cell Ig-like receptors (KIRs) on NK cells [31]. Another mechanism is related to CD32B mediated internalization. CD32B is the inhibitor Fcγ receptor IIB (FcγRIIB) with low-affinity for IgG [32]. It promotes rituximab internalization from B cells and abrogates ADCC, which contributes to rituximab resistance and is correlated with poor clinical responsiveness [33, 34]. Obinutuzumab was found to be less sensitive to the internalization, partially due to their ineffectiveness to redistribute CD20 into lipid rafts [35] and less ability to interact with and activate CD32B [36]. Obinutuzumab significantly enhanced expanded NK mediated ADCC than rituximab (Figure 3) possibly through all of the mechanisms.
Gene expression analysis uncovered very small number of genes (0.3%) exhibiting differential expression between obinutuzumab treated and control treated cell lines. GSEA clearly revealed the upregulation of genes related to apoptosis and mitogen-activated protein kinase signaling pathway and protracted downregulation of genes related to cell cycle and cell proliferation including ID3 and RAB6B though the reason for this is not well understood [28, 37, 38].
We demonstrated that obinutuzumab showed improved survival and inhibition of tumor progression in PMBL xenografted NSG mice. This data is consistent with superior activity of obinutuzumab in other B-cell lymphoma xenografts when used alone or in combination with other agents [1, 39–43].
In conclusion, this study reveals that using obinutuzumab as a single agent is significantly more effective in PMBL cells both in-vitro and in-vivo utilizing PMBL xenografted NSG mice compared to equal doses of rituximab. This novel anti-CD20 type II antibody should be considered as an alternative treatment for PMBL utilizing chemoimmunotherapy regimens.
Materials and Methods
Reagents and antibodies
PMBL Karpas-1106P cells (DSMZ, Germany) were cultured in RPMI 1640 with 20% FBS [24]. Obinutuzumab was generously provided by Hoffman LaRoche (Basel, Switzerland). Antibodies specific for phospho-protein kinase B, phospho-IκBα, phosphor-Stat6, AKT, IκBα and Stat6, were purchased from Cell Signaling Technology (Cambridge, MA & Santa Cruz, CA, USA).
MTS assay
Karpas-1106P cells (5.0 × 105) were treated with obinutuzumab at increasing dose concentrations (0, 1, 10, 20 and 100 μg/ml) for three days. Cell viability was measured by CellTiter 96 AQueous one solution cell proliferation assay (Pormega, Madison, WI, USA) [44] and measured by a multifilter plate reader (Molecular Device, CA, USA) at OD490. The obinutuzumab dose of 10 μg/ml was then compared to equal dosing of rituximab (10 μg/ml).
Immunoblotting
Whole-cell extracts were resolved on sodium dodecyl sulfate polyacrylamide gel electrophoresis, transferred to nitrocellulose membrane, probed with appropriate antibodies and blocked in 5% milk for 1 hour, subsequently incubated with primary antibodies at 4°C for overnight and developed using Enhanced Chemiluminescence Reagent (GE Healthcare Bio-Sciences, Piscataway, NJ, USA) [44]. Band intensities on sodium dodecyl sulfate polyacrylamide gel electrophoresis were measured using ImageJ software.
Quantitative reverse-transcription polymerase chain reaction
Total RNA was prepared using Trizol reagent (Invitrogen) according to the manufacturer’s manual. The cDNA was then synthesized using qScript™ cDNA Synthesis Kit (Quantas). Relative quantification (ddCt) of mRNA expression was determined by normalizing to the housekeeping gene (GAPDH).
Cell apoptosis assays
Cell death was assayed by flow cytometric analysis (Beckman Coulter, Brea, CA, USA) of Annexin V, and propidium iodide (BD Biosciences, San Jose, CA, USA). 3 × 105 cells were either untreated (control) or treated with 10 μg/ml of obinutuzumab, rituximab or human-immunoglobulin G for 24 to 72 hours. Annexin V/PI staining was performed according to the manufacturer instructions.
NK cell isolation and expansion
Peripheral blood was obtained from normal healthy adult donors after informed consent from the New York Blood Center and mononuclear cells were isolated by Ficoll-Paque (Amersham Biosciences, Piscataway, NJ, USA) density gradient separation as we have previously described [45].
NK cells were expanded in tissue culture plates with K562-mbIL15-41BBL in RPMI 10 IU/ml of IL-2 (R&D Systems, Inc., Minneapolis, MN, USA) and 10% FCS at 37°C with 5% CO2 as we have previously described [11, 46].
Antibody dependent cell mediated cytotoxicity assay
ADCC was determined using DELFIA EuTDA cytotoxicity assay as we have previously described [45]. 1 × 106 target cells were labeled with BATDA labeling ligand for 30 minutes. Tumor target cells were washed and incubated with 10 μg/ml obinutuzumab, rituximab or polyclonal IgG at 37°C and incubated with expanded NK effector cells, at 10:1 and 5:1 E: T target ratio for 4 hours at 37°C. Plates were centrifuged and the supernatant was collected into a flat-bottom plate and Europium solution was added and incubated for 15 minutes at room temperature. Fluorescence was measured with a time-resolved fluorometer (Perkin Elmer). The percentage of cytotoxicity was calculated using following equation: % specific release = (E-S/(M-S) × 100) E = experimental release, S = spontaneous release (target cells without effector cells), M = maximum release (lysed target cells).
Gene expression microarray
Gene expression experiments were performed using Illumina Human-HT12 v4.0 array at the Rockefeller University Genomics Facility (New York, NY, USA). Biotin-labeled RNA was prepared using 200 μg of total RNA by MessageAmp™ Premier RNA Amplification Kit (Applied Biosystems, Foster City, CA, USA). Antisense RNA (750 ng) was mixed with hybridisation reagents and heated at 65°C for 5 minutes. After it was cooled to room temperature, hybridization solution was applied to Illumina HumanHT-12 v4 array. The array was scanned using Illumina BeadArray Reader. The scanning was done using standard DirectHyb Gene Expression protocol. The raw data was extracted using Illumina BeadStudio software without normalization. Data has been deposited in the Gene Expression Omnibus database (accession number GSE75605).
GSEA with MSigMB v5.2 software was used to investigate the biological processes and pathways differentially implicated between obinutuzumab treated and PBS or IgG treated PBML cells [47, 48]. Genes with > 1.5 fold or < 1.5 fold expression ratio were considered to be significantly differentially expressed genes as we and others have previously discussed [48, 49].
In-vivo human PMBL xenograft NSG mice studies
The experimental animal protocols, procedures and care were approved by the Institutional Animal Care and Use Committee, at New York Medical College (NYMC, 33-2-0615H), Valhalla, NY, USA on January 20, 2015. Karpas-1106P PMBL cell line was stably transfected with a firefly luciferase expression plasmid (ffluc-zeo) to generate Karpas-1106P Luc+ cells [11]. Mice were γ- irradiated (2.5 Gy) 1 day before tumor cells were injected. Six- to eight- week-old female NSG (NOD. Cg-Prkdcscid Il2rgtm1Wjl/SzJ) mice from Jackson laboratory (Bar Harbor, ME, USA) were intravenously injected with 5 × 106 Karpas-1106P Luc+ cell as we have previously described [45].
The in-vivo anti-tumor function of obinutuzumab, (30 mg/kg) and rituximab (30 mg/kg) was determined in 6–8 week old mice. NSG mice were divided into 4 groups: 1) PBS only (control), 2) isotype control (IgG), 3) obinutuzumab (30 mg/kg), and 4) rituximab (30 mg/kg). One week after tumor cell injection, mice were injected every 7 days with the respective therapy for 8 weeks. Tumor regression and/or progression of xenografted mice were monitored weekly by in-vivo bioluminescence imaging and survival was monitored for 250 days.
Statistical analysis
Data obtained from 3 independent experiments are represented as means ± SD and statistical significance of the difference between two groups was determined by using two-tailed Student’s t-test. In in-vivo study, survival rates were analyzed by the Kaplan-Meier method and their statistical significance was evaluated by log-rank test using the Prism Version 6.0 software. P values < 0.05 were considered statistically significant for all in-vitro and in-vivo studies.
Abbreviations
PMBL: primary mediastinal large B-cell lymphoma; B-NHL: B-non Hodgkin lymphoma; BL: Burkitt lymphoma; JAK: Janus kinase; STAT: signal transducer and transcription; ADCC: antibody dependent cellular cytoxicity; FL: follicular lymphoma; NK: natural killer; PBS: phosphate buffer saline; GSEA: gene set enrichment analysis; DLBCL: diffuse large B-Cell lymphoma.
ACKNOWLEDGMENTS AND FUNDING
This work is supported by grant from Pediatric Cancer Research Foundation (PCRF), St. Baldrick’s Foundation and New York Medical College Child Health Translational Science Institute (CHTSI). Authors would like to thank Dean Lee, MD, PhD for generously providing artificial antigen-presenting cells (APC) (K562-mb-IL-15-41-BLL). The authors also want to thank Erin Morris, BSN and Virginia Davenport, RN for their expert assistance in the preparation of the manuscript.
CONFLICTS OF INTEREST
Dr. Klein declares employment, ownership interest and patents with Roche. All other authors declare that they have no competing interest.
References
1. Dunleavy K, Pittaluga S, Maeda LS, Advani R, Chen CC, Hessler J, Steinberg SM, Grant C, Wright G, Varma G, Staudt LM, Jaffe ES, Wilson WH. Dose-adjusted EPOCH-rituximab therapy in primary mediastinal B-cell lymphoma. N Engl J Med. 2013; 368:1408–1416. https://doi.org/10.1056/NEJMoa1214561. [PubMed].
2. Steidl C, Gascoyne RD. The molecular pathogenesis of primary mediastinal large B-cell lymphoma. Blood. 2011; 118:2659–2669. https://doi.org/10.1182/blood-2011-05-326538. [PubMed].
3. Rosenwald A, Wright G, Leroy K, Yu X, Gaulard P, Gascoyne RD, Chan WC, Zhao T, Haioun C, Greiner TC, Weisenburger DD, Lynch JC, Vose J, et al. Molecular diagnosis of primary mediastinal B cell lymphoma identifies a clinically favorable subgroup of diffuse large B cell lymphoma related to Hodgkin lymphoma. J Exp Med. 2003; 198:851–862. https://doi.org/10.1084/jem.20031074. [PubMed].
4. Savage KJ, Monti S, Kutok JL, Cattoretti G, Neuberg D, De Leval L, Kurtin P, Dal Cin P, Ladd C, Feuerhake F, Aguiar RC, Li S, Salles G, et al. The molecular signature of mediastinal large B-cell lymphoma differs from that of other diffuse large B-cell lymphomas and shares features with classical Hodgkin lymphoma. Blood. 2003; 102:3871–3879. https://doi.org/10.1182/blood-2003-06-1841. [PubMed].
5. Lim MS. PMBCL: a molecular diagnosis? Blood. 2018; 132:2319–2320. https://doi.org/10.1182/blood-2018-10-878660. [PubMed].
6. Mottok A, Wright G, Rosenwald A, Ott G, Ramsower C, Campo E, Braziel RM, Delabie J, Weisenburger DD, Song JY, Chan WC, Cook JR, Fu K, et al. Molecular classification of primary mediastinal large B-cell lymphoma using routinely available tissue specimens. Blood. 2018; 132:2401–2405. https://doi.org/10.1182/blood-2018-05-851154. [PubMed].
7. Roberts RA, Wright G, Rosenwald AR, Jaramillo MA, Grogan TM, Miller TP, Frutiger Y, Chan WC, Gascoyne RD, Ott G, Muller-Hermelink HK, Staudt LM, Rimsza LM. Loss of major histocompatibility class II gene and protein expression in primary mediastinal large B-cell lymphoma is highly coordinated and related to poor patient survival. Blood. 2006; 108:311–318. https://doi.org/10.1182/blood-2005-11-4742. [PubMed].
8. Rigaud G, Moore PS, Taruscio D, Scardoni M, Montresor M, Menestrina F, Scarpa A. Alteration of chromosome arm 6p is characteristic of primary mediastinal B-cell lymphoma, as identified by genome-wide allelotyping. Genes Chromosomes Cancer. 2001; 31:191–195. https://doi.org/10.1002/gcc.1133. [PubMed].
9. Chen BJ, Chapuy B, Ouyang J, Sun HH, Roemer MG, Xu ML, Yu H, Fletcher CD, Freeman GJ, Shipp MA, Rodig SJ. PD-L1 expression is characteristic of a subset of aggressive B-cell lymphomas and virus-associated malignancies. Clin Cancer Res. 2013; 19:3462–3473. https://doi.org/10.1158/1078-0432.CCR-13-0855. [PubMed].
10. Salles G, Morschhauser F, Lamy T, Milpied N, Thieblemont C, Tilly H, Bieska G, Asikanius E, Carlile D, Birkett J, Pisa P, Cartron G. Phase 1 study results of the type II glycoengineered humanized anti-CD20 monoclonal antibody obinutuzumab (GA101) in B-cell lymphoma patients. Blood. 2012; 119:5126–5132. https://doi.org/10.1182/blood-2012-01-404368. [PubMed].
11. Awasthi A, Ayello J, Van de Ven C, Elmacken M, Sabulski A, Barth MJ, Czuczman MS, Islam H, Klein C, Cairo MS. Obinutuzumab (GA101) compared to rituximab significantly enhances cell death and antibody-dependent cytotoxicity and improves overall survival against CD20(+) rituximab-sensitive/-resistant Burkitt lymphoma (BL) and precursor B-acute lymphoblastic leukaemia (pre-B-ALL): potential targeted therapy in patients with poor risk CD20(+) BL and pre-B-ALL. Br J Haematol. 2015; 171:763–775. https://doi.org/10.1111/bjh.13764. [PubMed].
12. Awasthi A, Rolland DCM, Ayello J, van de Ven C, Basrur V, Conlon K, Fermin D, Barth MJ, Klein C, Elenitoba-Johnson KSJ, Lim MS, Cairo MS. A comparative global phosphoproteomics analysis of obinutuzumab (GA101) versus rituximab (RTX) against RTX sensitive and resistant Burkitt lymphoma (BL) demonstrates differential phosphorylation of signaling pathway proteins after treatment. Oncotarget. 2017; 8:113895–113909. https://doi.org/10.18632/oncotarget.23040. [PubMed].
13. Bologna L, Gotti E, Manganini M, Rambaldi A, Intermesoli T, Introna M, Golay J. Mechanism of action of type II, glycoengineered, anti-CD20 monoclonal antibody GA101 in B-chronic lymphocytic leukemia whole blood assays in comparison with rituximab and alemtuzumab. J Immunol. 2011; 186:3762–3769. https://doi.org/10.4049/jimmunol.1000303. [PubMed].
14. Loisel S, Andre PA, Golay J, Buchegger F, Kadouche J, Cerutti M, Bologna L, Kosinski M, Viertl D, Delaloye AB, Berthou C, Mach JP, Boumsell L. Antitumour effects of single or combined monoclonal antibodies directed against membrane antigens expressed by human B cells leukaemia. Mol Cancer. 2011; 10:42. https://doi.org/10.1186/1476-4598-10-42. [PubMed].
15. Sehn LH, Goy A, Offner FC, Martinelli G, Caballero MD, Gadeberg O, Baetz T, Zelenetz AD, Gaidano G, Fayad LE, Buckstein R, Friedberg JW, Crump M, et al. Randomized Phase II Trial Comparing Obinutuzumab (GA101) With Rituximab in Patients With Relapsed CD20+ Indolent B-Cell Non-Hodgkin Lymphoma: Final Analysis of the GAUSS Study. J Clin Oncol. 2015; 33:3467–3474. https://doi.org/10.1200/JCO.2014.59.2139. [PubMed].
16. Cheson BD, Chua N, Mayer J, Dueck G, Trneny M, Bouabdallah K, Fowler N, Delwail V, Press O, Salles G, Gribben JG, Lennard A, Lugtenburg PJ, et al. Overall Survival Benefit in Patients With Rituximab-Refractory Indolent Non-Hodgkin Lymphoma Who Received Obinutuzumab Plus Bendamustine Induction and Obinutuzumab Maintenance in the GADOLIN Study. J Clin Oncol. 2018; 36:2259–2266. https://doi.org/10.1200/JCO.2017.76.3656. [PubMed].
17. Marcus R, Davies A, Ando K, Klapper W, Opat S, Owen C, Phillips E, Sangha R, Schlag R, Seymour JF, Townsend W, Trneny M, Wenger M, et al. Obinutuzumab for the First-Line Treatment of Follicular Lymphoma. N Engl J Med. 2017; 377:1331–1344. https://doi.org/10.1056/NEJMoa1614598. [PubMed].
18. Goede V, Fischer K, Busch R, Engelke A, Eichhorst B, Wendtner CM, Chagorova T, de la Serna J, Dilhuydy MS, Illmer T, Opat S, Owen CJ, Samoylova O, et al. Obinutuzumab plus chlorambucil in patients with CLL and coexisting conditions. N Engl J Med. 2014; 370:1101–1110. https://doi.org/10.1056/NEJMoa1313984. [PubMed].
19. Moreno C, Greil R, Demirkan F, Tedeschi A, Anz B, Larratt L, Simkovic M, Samoilova O, Novak J, Ben-Yehuda D, Strugov V, Gill D, Gribben JG, et al. Ibrutinib plus obinutuzumab versus chlorambucil plus obinutuzumab in first-line treatment of chronic lymphocytic leukaemia (iLLUMINATE): a multicentre, randomised, open-label, phase 3 trial. Lancet Oncol. 2019; 20:43–56. https://doi.org/10.1016/S1470-2045(18)30788-5. [PubMed].
20. Sehn LH, Martelli M, Trněný M, Liu W, Bolen CR, Knapp A, Sahin D, Sellam GU, Vitolo U. Final Analysis of GOYA: A Randomized, Open-Label, Phase III Study of Obinutuzumab or Rituximab Plus CHOP in Patients with Previously Untreated Diffuse Large B-Cell Lymphoma. Blood. 2019; 134:S4088. https://doi.org/10.1182/blood-2019-123433.
21. Casasnovas RO, Ysebaert L, Thieblemont C, Bachy E, Feugier P, Delmer A, Tricot S, Gabarre J, Andre M, Fruchart C, Mounier N, Delarue R, Meignan M, et al. FDG-PET-driven consolidation strategy in diffuse large B-cell lymphoma: final results of a randomized phase 2 study. Blood. 2017; 130:1315–1326. https://doi.org/10.1182/blood-2017-02-766691. [PubMed].
22. Vitolo U, Trneny M, Belada D, Burke JM, Carella AM, Chua N, Abrisqueta P, Demeter J, Flinn I, Hong X, Kim WS, Pinto A, Shi YK, et al. Obinutuzumab or Rituximab Plus Cyclophosphamide, Doxorubicin, Vincristine, and Prednisone in Previously Untreated Diffuse Large B-Cell Lymphoma. J Clin Oncol. 2017; 35:3529–3537. https://doi.org/10.1200/JCO.2017.73.3402. [PubMed].
23. Renné C, Willenbrock K, Martin-Subero JI, Hinsch N, Döring C, Tiacci E, Klapper W, Möller P, Küppers R, Hansmann ML, Siebert R, Bräuninger A. High expression of several tyrosine kinases and activation of the PI3K/AKT pathway in mediastinal large B cell lymphoma reveals further similarities to Hodgkin lymphoma. Leukemia. 2007; 21:780–787. https://doi.org/10.1038/sj.leu.2404594. [PubMed].
24. Mossner E, Brunker P, Moser S, Puntener U, Schmidt C, Herter S, Grau R, Gerdes C, Nopora A, van Puijenbroek E, Ferrara C, Sondermann P, Jager C, et al. Increasing the efficacy of CD20 antibody therapy through the engineering of a new type II anti-CD20 antibody with enhanced direct and immune effector cell-mediated B-cell cytotoxicity. Blood. 2010; 115:4393–4402. https://doi.org/10.1182/blood-2009-06-225979. [PubMed].
25. Tobinai K, Klein C, Oya N, Fingerle-Rowson G. A Review of Obinutuzumab (GA101), a Novel Type II Anti-CD20 Monoclonal Antibody, for the Treatment of Patients with B-Cell Malignancies. Adv Ther. 2017; 34:324–356. https://doi.org/10.1007/s12325-016-0451-1. [PubMed].
26. Herter S, Birk MC, Klein C, Gerdes C, Umana P, Bacac M. Glycoengineering of therapeutic antibodies enhances monocyte/macrophage-mediated phagocytosis and cytotoxicity. J Immunol. 2014; 192:2252–2260. https://doi.org/10.4049/jimmunol.1301249. [PubMed].
27. Miloudi H, Leroy K, Jardin F, Sola B. STAT6 is a cargo of exportin 1: Biological relevance in primary mediastinal B-cell lymphoma. Cell Signal. 2018; 46:76–82. https://doi.org/10.1016/j.cellsig.2018.02.016. [PubMed].
28. Decaup E, Jean C, Laurent C, Gravelle P, Fruchon S, Capilla F, Marrot A, Al Saati T, Frenois FX, Laurent G, Klein C, Varoqueaux N, Savina A, et al. Anti-tumor activity of obinutuzumab and rituximab in a follicular lymphoma 3D model. Blood Cancer J. 2013; 3:e131. https://doi.org/10.1038/bcj.2013.32. [PubMed].
29. Yasuhiro T, Sawada W, Klein C, Kozaki R, Hotta S, Yoshizawa T. Anti-tumor efficacy study of the Bruton’s tyrosine kinase (BTK) inhibitor, ONO/GS-4059, in combination with the glycoengineered type II anti-CD20 monoclonal antibody obinutuzumab (GA101) demonstrates superior in vivo efficacy compared to ONO/GS-4059 in combination with rituximab. Leuk Lymphoma. 2017; 58:699–707. https://doi.org/10.1080/10428194.2016.1201567. [PubMed].
30. Herter S, Herting F, Mundigl O, Waldhauer I, Weinzierl T, Fauti T, Muth G, Ziegler-Landesberger D, Van Puijenbroek E, Lang S, Duong MN, Reslan L, Gerdes CA, et al. Preclinical activity of the type II CD20 antibody GA101 (obinutuzumab) compared with rituximab and ofatumumab in vitro and in xenograft models. Mol Cancer Ther. 2013; 12:2031–2042. https://doi.org/10.1158/1535-7163.MCT-12-1182. [PubMed].
31. Terszowski G, Klein C, Stern M. KIR/HLA interactions negatively affect rituximab- but not GA101 (obinutuzumab)-induced antibody-dependent cellular cytotoxicity. J Immunol. 2014; 192:5618–5624. https://doi.org/10.4049/jimmunol.1400288. [PubMed].
32. Rankin CT, Veri MC, Gorlatov S, Tuaillon N, Burke S, Huang L, Inzunza HD, Li H, Thomas S, Johnson S, Stavenhagen J, Koenig S, Bonvini E. CD32B, the human inhibitory Fc-gamma receptor IIB, as a target for monoclonal antibody therapy of B-cell lymphoma. Blood. 2006; 108:2384–2391. https://doi.org/10.1182/blood-2006-05-020602. [PubMed].
33. Lim SH, Vaughan AT, Ashton-Key M, Williams EL, Dixon SV, Chan HT, Beers SA, French RR, Cox KL, Davies AJ, Potter KN, Mockridge CI, Oscier DG, et al. Fc gamma receptor IIb on target B cells promotes rituximab internalization and reduces clinical efficacy. Blood. 2011; 118:2530–2540. https://doi.org/10.1182/blood-2011-01-330357. [PubMed].
34. Roghanian A, Teige I, Martensson L, Cox KL, Kovacek M, Ljungars A, Mattson J, Sundberg A, Vaughan AT, Shah V, Smyth NR, Sheth B, Chan HT, et al. Antagonistic human FcgammaRIIB (CD32B) antibodies have anti-tumor activity and overcome resistance to antibody therapy in vivo. Cancer Cell. 2015; 27:473–488. https://doi.org/10.1016/j.ccell.2015.03.005. [PubMed].
35. Cragg MS, Morgan SM, Chan HT, Morgan BP, Filatov AV, Johnson PW, French RR, Glennie MJ. Complement-mediated lysis by anti-CD20 mAb correlates with segregation into lipid rafts. Blood. 2003; 101:1045–1052. https://doi.org/10.1182/blood-2002-06-1761. [PubMed].
36. Vaughan AT, Iriyama C, Beers SA, Chan CH, Lim SH, Williams EL, Shah V, Roghanian A, Frendeus B, Glennie MJ, Cragg MS. Inhibitory FcgammaRIIb (CD32b) becomes activated by therapeutic mAb in both cis and trans and drives internalization according to antibody specificity. Blood. 2014; 123:669–677. https://doi.org/10.1182/blood-2013-04-490821. [PubMed].
37. Heinrich DA, Weinkauf M, Hutter G, Zimmermann Y, Jurinovic V, Hiddemann W, Dreyling M. Differential regulation patterns of the anti-CD20 antibodies obinutuzumab and rituximab in mantle cell lymphoma. Br J Haematol. 2015; 168:606–610. https://doi.org/10.1111/bjh.13132. [PubMed].
38. Miyazaki M, Miyazaki K, Chen S, Chandra V, Wagatsuma K, Agata Y, Rodewald HR, Saito R, Chang AN, Varki N, Kawamoto H, Murre C. The E-Id protein axis modulates the activities of the PI3K-AKT-mTORC1-Hif1a and c-myc/p19Arf pathways to suppress innate variant TFH cell development, thymocyte expansion, and lymphomagenesis. Genes Dev. 2015; 29:409–425. https://doi.org/10.1101/gad.255331.114. [PubMed].
39. Frazer JK, Li KJ, Galardy PJ, Perkins SL, Auperin A, Anderson JR, Pinkerton R, Buxton A, Gross TG, Michon J, Leverger G, Weinstein HJ, Harrison L, et al. Excellent outcomes in children and adolescents with CNS(+) Burkitt lymphoma or other mature B-NHL using only intrathecal and systemic chemoimmunotherapy: results from FAB/LMB96 and COG ANHL01P1. Br J Haematol. 2019; 185:374–377. https://doi.org/10.1111/bjh.15520. [PubMed].
40. Gerrard M, Waxman IM, Sposto R, Auperin A, Perkins SL, Goldman S, Harrison L, Pinkerton R, McCarthy K, Raphael M, Patte C, Cairo MS, and French-American-British/Lymphome Malins de Burkitt 96 (FAB/LMB 96) International Study Committee. Outcome and pathologic classification of children and adolescents with mediastinal large B-cell lymphoma treated with FAB/LMB96 mature B-NHL therapy. Blood. 2013; 121:278–285. https://doi.org/10.1182/blood-2012-04-422709. [PubMed].
41. Giulino-Roth L, O’Donohue T, Chen Z, Bartlett NL, LaCasce A, Martin-Doyle W, Barth MJ, Davies K, Blum KA, Christian B, Casulo C, Smith SM, Godfrey J, et al. Outcomes of adults and children with primary mediastinal B-cell lymphoma treated with dose-adjusted EPOCH-R. Br J Haematol. 2017; 179:739–747. https://doi.org/10.1111/bjh.14951. [PubMed].
42. Goldman S, Smith L, Anderson JR, Perkins S, Harrison L, Geyer MB, Gross TG, Weinstein H, Bergeron S, Shiramizu B, Sanger W, Barth M, Zhi J, Cairo MS. Rituximab and FAB/LMB 96 chemotherapy in children with Stage III/IV B-cell non-Hodgkin lymphoma: a Children’s Oncology Group report. Leukemia. 2013; 27:1174–1177. https://doi.org/10.1038/leu.2012.255. [PubMed].
43. Vassilakopoulos TP, Pangalis GA, Chatziioannou S, Papageorgiou S, Angelopoulou MK, Galani Z, Kourti G, Prassopoulos V, Leonidopoulou T, Terpos E, Dimopoulou MN, Sachanas S, Kalpadakis C, et al. PET/CT in primary mediastinal large B-cell lymphoma responding to rituximab-CHOP: An analysis of 106 patients regarding prognostic significance and implications for subsequent radiotherapy. Leukemia. 2016; 30:238–242. https://doi.org/10.1038/leu.2015.120. [PubMed].
44. Chu Y, Yahr A, Huang B, Ayello J, Barth M, Cairo MS. Romidepsin alone or in combination with anti-CD20 chimeric antigen receptor expanded natural killer cells targeting Burkitt lymphoma in vitro and in immunodeficient mice. Oncoimmunology. 2017; 6:e1341031. https://doi.org/10.1080/2162402X.2017.1341031. [PubMed].
45. Ayello J, Hochberg J, Flower A, Chu Y, Baxi LV, Quish W, van de Ven C, Cairo MS. Genetically re-engineered K562 cells significantly expand and functionally activate cord blood natural killer cells: Potential for adoptive cellular immunotherapy. Exp Hematol. 2017; 46:38–47. https://doi.org/10.1016/j.exphem.2016.10.003. [PubMed].
46. Chu Y, Hochberg J, Yahr A, Ayello J, van de Ven C, Barth M, Czuczman M, Cairo MS. Targeting CD20+ Aggressive B-cell Non-Hodgkin Lymphoma by Anti-CD20 CAR mRNA-Modified Expanded Natural Killer Cells In Vitro and in NSG Mice. Cancer Immunol Res. 2015; 3:333–44. https://doi.org/10.1158/2326-6066.CIR-14-0114. [PubMed].
47. Lee S, Day NS, Miles RR, Perkins SL, Lim MS, Ayello J, van de Ven C, Harrison L, El-Mallawany NK, Goldman S, Cairo MS. Comparative genomic expression signatures of signal transduction pathways and targets in paediatric Burkitt lymphoma: a Children’s Oncology Group report. Br J Haematol. 2017; 177:601–611. https://doi.org/10.1111/bjh.14604. [PubMed].
48. Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub TR, Lander ES, Mesirov JP. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A. 2005; 102:15545–15550. https://doi.org/10.1073/pnas.0506580102. [PubMed].
49. Lee S, Luo W, Shah T, Yin C, O’Connell T, Chung TH, Perkins SL, Miles RR, Ayello J, Morris E, Harrison L, van de Ven C, Cairo MS. The effects of DLEU1 gene expression in Burkitt lymphoma (BL): potential mechanism of chemoimmunotherapy resistance in BL. Oncotarget. 2017; 8:27839–27853. https://doi.org/10.18632/oncotarget.15711. [PubMed].