
INTRODUCTION
According to the World Health Organization, breast cancer (BC) is the most common tumor in women worldwide [1]. It is known that 5–10% of BC cases have a hereditary component [2], being characterized by the presence of germline mutations in the BRCA1 [3] or BRCA2 [4] genes, which are associated with the hereditary breast and ovarian cancer predisposition syndrome (HBOC). HBOC patients have strong personal and family histories of cancer. Moreover, these patients are characterized by early age-at-diagnosis of cancer, increased frequency of bilateral tumors, and two or more generations affected by cancer [5, 6].
Recent studies have shown that alterations in other susceptibility genes, mainly involved in the homologous recombination and DNA repair pathways, can be causal factors of hereditary breast and ovarian cancers [7]. In spite of that, the predisposing genetic cause of about 50% of the families at-risk for hereditary breast and ovarian cancers remains unknown [8, 9]. These families are grouped in a category called BRCAX.
Evidences from the literature have shown that BRCAX tumors are rather heterogeneous, involving several different histopathological subgroups and genetic alterations [10, 11]. Several authors have shown the presence of new high penetrance genes associated with breast and ovarian cancers [11–17]. However, the opinion of the scientific community is controversial. There are authors who argue that the incidence of BRCAX tumors is associated with rare syndromes in which BC is only one component [12, 15, 16]. Other authors believe that this type of tumor results from mutations in several genes with low penetrance or population-specific [11, 13, 14, 18].
Studies using array-comparative genomic hybridization (aCGH) technique suggest that several chromosomal regions are associated with the development of hereditary BC, highlighting gains at chromosomes 1q, 8q, 17q and 20q and losses within chromosomes 8p, 11q, 13q and 17p [19–24]. Despite these findings, more studies are necessary to a better understanding of BRCAX molecular events in hereditary breast cancer. In this regard, the aim of this study was to identify chromosomal and subchromosomal copy number alterations in tumor samples from Brazilian women without BRCA1/BRCA2 germline mutations with family history strongly suggestive of HBOC syndrome.
RESULTS
In the present study, we analyzed 31 Brazilian women at-risk for hereditary breast/ovarian cancer (27 with personal history of BC and 4 with ovarian tumors) without BRCA1/BRCA2/TP53 germline mutations, by array-CGH. Clinicopathological characteristics and family history of the patients are specified in Table 1.
Table 1: Clinicopathological characteristics and family history of the patients at-risk for hereditary cancer
Family |
Cancer (age at diagnosis) |
Histological type |
Molecular subtype |
Breast/Ovarian cancer cases in the family (sex and age at diagnosis, if known) |
|---|---|---|---|---|
19 |
Breast (44) |
IDC |
ER: –; PR: +; HER2: – |
Sister: Breast (F,46; F,46) |
29 |
Ovarian (42), Breast (53) |
DCIS |
ER: +; PR: + |
Paternal side of the family: Breast (F,29); Ovarian (F,60; F,?; F, ?), Uterus (F,57; F,?; F,?); Gastric (M,42; M,?; M,?; M,?) |
65 |
Breast (35) |
IDC |
ER: –; PR: –; HER2: – |
Maternal side of the family: Breast (F,31; F,34; F,47; F,39; F,39; F,46) |
80 |
Breast (43) |
DCIS |
ER: +; PR: +; HER2: + |
Maternal side of the family: Breast (F,44; f,44; F,55; F,57; F,60), Prostate (M,?) |
85 |
Breast (51) |
IDC |
ER: +; PR:+; HER2: – |
Maternal side of the family: Breast (F,43; F,45; F,48), Stomach (F;45; M,56); Leukemia (M,69) |
179 |
Breast (47) |
IDC |
ER: +; PR: + |
Paternal side of the family: Breast (F,37; F,49; F,61), Throat (M,?; M,?) |
233 |
Breast (49) |
IDC |
ER: –; PR: –; HER2: – |
Maternal side of the family: Breast (F,?; F,50; F,33; F,70; F,60; F,60; F,46), Colorectal (M,65), Gastric (M,62), Pancreas (M,62), Lung (M,52; M,66; M,?) |
241 |
Breast (45) |
IDC |
ER: +; PR: +; HER2: – |
Paternal side of the family: Breast (F,48; F,49) |
275 |
Ovarian (60) |
Serous adenocarcinoma |
Not applicable |
Paternal side of the family: Breast (F,32; F,35), Prostate (M,80) |
289 |
Breast (48) |
IDC |
ER: –; PR: –; HER2: + |
Maternal side of the family: Breast (F,50; F,65; F,65) |
306 |
Melanoma (26), Breast (36) |
IDC |
ER: +; PR: +; HER2: – |
Maternal side of the family: Breast (F,43; F,?) |
320 |
Ovarian (53) |
Serous adenocarcinoma |
Not applicable |
Maternal side of the family: Breast (F,52), Ovarian (F,71), Uterus (F,60), Thyroid (M,29), Lung (M,83) |
426 |
Breast (38) |
IDC |
ER: +; PR: +; HER2: – |
Paternal side of the family: Breast (F,20); Ovarian (F,28), Leukemia (M,78), Esophagus (M,?) |
494 |
Breast (33) |
IDC |
ER: +; PR: +; HER2: + |
Maternal side of the family: Breast (F,38; F,?), Ovarian (F,38) |
558 |
Breast (37) |
IDC |
ER: –; PR: –; HER2: – |
Maternal side of the family: Breast (F,52; F,?; F,?; F,?, F,?), Ovarian (F,42), Skin (F,?) |
563 |
Breast (39) |
IDC |
ER: +; PR: –; HER2: – |
Paternal side of the family: Breast (F,30; F,40; F,45, F,50; F,51), Lung, (M,?), Colorectal (F,64), Skin (M,72) |
581 |
Breast (45) |
DCIS |
Not available |
Paternal side of the family: Breast (F,49; F,46; F,54), Prostate (M,60; M,70), head and neck (M,83) |
593 |
Breast (39) |
IDC |
ER: +; PR: +; HER2: – |
Paternal side of the family: Breast (F,50; F,?; F,?; F,?), Ovarian (F,?; F,?), Gastric (M,?; M,?), Colorectal (M,?; M,?) |
626 |
Breast (46) |
DCIS |
ER: +; PR: + |
Maternal side of the family: Breast (F,74; F,80; F,57; F,45), Ovarian (F,45), Thyroid (F,40), Skin (M,80), Pancreas (M,?), Myeloma (M,60), Lips (M,?) |
638 |
Breast (42) |
ILC |
ER: +; PR: + |
Maternal side of the family: Breast (F,49; F,50; F,?), Gastric (F,55), Thyroid (F,36), Lips (F,55) |
649 |
Breast (38) |
IDC |
ER: +; PR: +; HER2: – |
Maternal side of the family: Breast (F,64), Ovarian (F,61), Thyroid (F,61) |
695 |
Ovarian (21) |
Serous adenocarcinoma |
Not applicable |
Paternal side of the family: Breast (F,42), Ovarian (F,68), Colorectal (F,40; M,40), Gastric (F,50; F,70) |
960 |
Bilateral Breast (59,70) |
IDC |
ER: +; PR: +; HER2: – |
Maternal side of the family: Breast (F,34; F,59), Uterus (F,45), Lung (M,77; M,?) |
974 |
Breast (46) |
IDC |
ER: +; PR: +; HER2: – |
Maternal side of the family: Breast (F,55; F,45; F,60; F,60; F,55; F,45; F,60), Prostate (M,70; M,80) |
981 |
Breast (37) |
IDC |
ER: +; PR: +; HER2: – |
Maternal side of the family: Breast (F,32; F,70; F,60), Melanoma (F,30; F36), Leukemia (F,5), Bile ducts (M,49; F,55) |
1014 |
Breast (42) |
DCIS |
ER: +; PR: + |
Maternal side of the family: Breast (F,53; F,?); Melanoma (F,75), Lymphoma (M,19), Liver (F,?), Brain (F,?) |
1024 |
Breast (48) |
IDC |
ER: –; PR: –; HER2: – |
Paternal side of the family: Breast (F,70; F,72; F,44; F;44; F,49), Ovarian (F,56), Colorectal (M,20), Melanoma (M, ?), Prostate (M,50), Gastric (F,70; F,72; F,41) |
1055 |
Ovarian (57) |
Serous adenocarcinoma |
Not applicable |
Maternal side of the family: Breast (F,49; F,50), Pancreas (F,50), Lung (M,?) |
1095 |
Breast (43) |
IDC |
ER: –; PR: –; HER2: + |
Paternal side of the family: Breast (F,27; F,42), Uterus (F,98), Throat (M,72) |
1151 |
Breast (38) |
IDC |
ER: +; PR: +; HER2: – |
Maternal side of the family: Breast (F,35; F,60) |
1264 |
Breast (27) |
IDC |
ER: –; PR: +; HER2: – |
Maternal side of the family: Breast (F,50), Pancreas (M,75); Intestine (M,81) |
Briefly, the mean age at BC diagnosis was 42.9 years (SD = 7.9), ranging from 27–70 years. The majority of BC was invasive ductal carcinoma (77.8%), estrogen and progesterone positive (69.2% and 73.1%, respectively) and HER2 negative (64.0%). Regarding molecular classification, the majority of patients presented luminal type tumors (21 patients, 80.8%), four patients (15.4%) had triple negative tumors and only one patient (3.8%) was diagnosed with a HER2 subtype tumor.
All four ovarian cancer patients developed serous adenocarcinoma subtype tumors. The average age at diagnosis was 47.7 years (SD = 18.0), ranging from 21–60 years.
A detailed cancer family history can be found in Table 1. All patients reported at least one case of BC in the family, diagnosed at early age (<55 years for breast cancer cases). In addition, two women at-risk for hereditary BC (samples: 960 and 1024) had a family history with bilateral BC. Among patients diagnosed with BC, the majority reported more than three cases of BC in the family history (16 cases, 59.3%). Meanwhile, all patients diagnosed with ovarian cancer, reported three or less BC cases in their families (p = 0.043). Moreover, 12 patients reported the presence of ovarian cancer in the family history.
The molecular analysis revealed gained and lost regions across all chromosomes for both breast and ovarian tumors (Figure 1). We found 20 gained regions and 31 lost in BRCAX tumors. In addition, some variations, although not statistically significant, were found only in patients diagnosed with breast cancer, such as: gains of 7p22.1, 12p13.1, 14q13.3-q21.1, 17q11.2, 17q12 and 17q21.32-q21.33, and losses of 2p25.3, 6q25.3-q26 and 10q26.3. Moreover, the gain of Xq26 and loss of Xp22.32–22.31 were more frequent in ovarian cancer (100%), compared with breast cases (26% and 59%, respectively) (p = 0.01 for both regions). Loss of 22q13.31–13.32 was detected more often in ovarian than in breast cancer cases (p = 0.043). In addition, a significant number of copy number alterations involving chromosome 8 was observed.
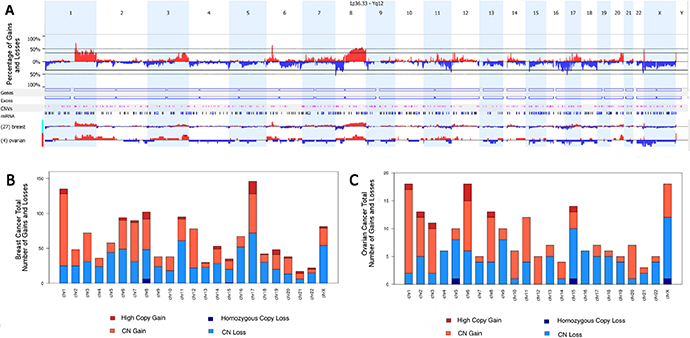
Figure 1: Overview of gained and lost regions across all chromosomes. (A) Overall and specific breast and ovarian copy number aberration frequencies. Regions presenting copy gains are shown in red and with copy loss in blue. (B) Overview of gained and lost regions across all chromosomes in breast tumors. (C) Overview of gained and lost regions across all chromosomes in ovarian tumors.
When family history was taken into consideration for copy number variation analyses, we observed that loss of 22q13.31–13.32 region was significantly associated with the presence of ovarian family history (p = 0.03). This region includes MIR3201, LOC284933, FAM19A5, MIR4535, LINC01310 genes. Other significant association found included gains in the 6p22.1 region (including 13 histone family genes) in 100% of metastatic cases (p = 0.03). Finally, we found loss of 6q25.1 in 71% of patients with metastasis (p = 0.01). This region includes RAET1E, RAET1E-AS1, RAET1G, ULBP2, ULBP1, RAET1K, RAET1L, ULBP3, PPP1R14C, IYD, PLEKHG1, MTHFD1L genes (Supplementary Table 1).
In addition, when comparing our findings with those of the literature of BRCAX tumors, we observed that our results corroborate some findings reported by Didraga et al. (2011), Alvarez et al. (2016) and Mangia et al. (2008), showing 50%, 21% and 12% of common regions, respectively (Figure 2).
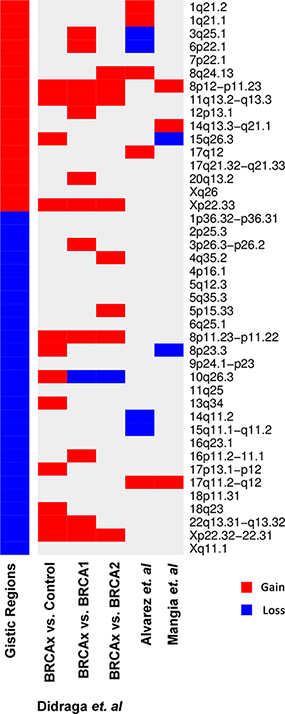
Figure 2: Heatmap representing the gains (in red) and losses (in blue) through aCGH found by GISTIC algorighm in common with previous studies by Didraga et al. (2011), Alvarez et al. (2016) and Mangia et al. (2008).
Finally, we found that 22 genes present in gained regions also present overexpression in the Oncomine database, whereas 21 genes present in lost regions show loss of expression in the same database (p < 0.01, Table 2).
Table 2: Genes in gained regions that presented in silico overexpression and genes in lost regions that presented in silico loss of expression
Event1 |
Cytoband |
Genes |
|---|---|---|
Gain/Overexp |
1q21.1-q21.2 |
PEX11B, PDE4DIP, ECM1, TARS2, RPRD2 |
Gain/Overexp |
6p22.1 |
HIST1H3H, HIST1H4J, HIST1H4K |
Gain/Overexp |
7p21.1 |
HDAC9 |
Gain/Overexp |
8p11.23-p11.22 |
TM2D2, LETM2, RNF5P1, FGFR1 |
Gain/Overexp |
8q24.13 |
NSMCE2, KIAA0196 |
Gain/Overexp |
11q13.3 |
CTTN |
Gain/Overexp |
17q12 |
ERBB2, STARD3, GRB7 |
Gain/Overexp |
17q21.32-q21.33 |
PHB, ABI3 |
Gain/Overexp |
Xp22.33 |
CRLF2 |
Loss/LOExp |
1p36.32 |
TPRG1L, AJAP1 |
Loss/LOExp |
2p25.3 |
FAM150B, TMEM18, TPO |
Loss/LOExp |
3p26.3-p26.2 |
CRBN, CNTN4 |
Loss/LOExp |
5q35.3 |
ADAMTS2, ZNF879, COL23A1 |
Loss/LOExp |
8p23.3 |
ERICH1, RPL23AP53, OR4F21, ZNF596 |
Loss/LOExp |
11q25 |
JAM3, LOC283177, THYN1 |
Loss/LOExp |
14q11.2 |
OR4K5 |
Loss/LOExp |
16q23.1 |
CNTNAP4, SYCE1L |
Loss/LOExp |
Xp22.32-p22.31 |
NLGN4X |
1Events represent Gain or Loss in our samples with concurrent Overexpression (Overexp) or Loss of Expression (LOExp) on Oncomine samples.
DISCUSSION
In the present study, a BRCAX tumor characterization of FFPE samples has been performed by array comparative genomic hybridization. Among the altered loci, we can highlight the identification of several alterations in chromosome 8, including losses on 8p12-p11.23 and gains on 8p12-p11.23 and 8q24.13, in concordance with previous studies of BRCAX tumors [14, 25, 26]. Besides, the chromosomal region 8p12-p11 has been reported to be amplified in 10–23% of BC cases [27–29], and some studies have shown that amplification on this region is associated with poor clinical outcome [27, 30]. We found by in silico analysis that 4 genes present in this region (including FGFR1 and NSMCE2) are overexpressed.
The FGFR1 gene encodes a transmembrane protein that interacts with fibroblast growth factors and directly influence mitogenesis and cell differentiation. In fact, there are several studies showing different treatment outcomes of breast cancer women depending on the FGFR1 status [31–33]. Similarly, NSMCE2 plays an important role in cell cycle, since its depletion in MCF-7 breast cancer cells affected cell cycle and G1-S transition [34]. Moreover, the overexpression of cortactin (CTTN), present in 11q13.3, was linked to CCND1 amplification in premenopausal breast cancer [35], although it failed to demonstrate a strong prognostic value in patients with breast cancer [36]. Conversely, its upregulation promoted colon cancer progression through ERK pathway [37]. Therefore, other studies have shown that amplification on chromosomal region 8p12-p11 in combination with amplification on 11q13 have more impact on patient outcome than amplification on only one of the two loci [27, 38].
In addition to gains and losses on chromosome 8 and alterations on chromosome 11, alterations in chromosome X seem to be characteristic of BRCAX tumors. In our study, a great number of samples showed gains on regions 11q13.2-q13.3 and Xp22.33, which were also identified by Didraga and collaborators [25]. Although it is not extensively studied in breast cancer, the overexpression of CRLF2, present in Xp22.33, has been demonstrated to be a marker of poor outcome of pediatric and adult B-precursor acute lymphoblastic leucemia (ALL) (as reviewed in [39]).
Study performed by Gronwald et al. [19] compared BRCAX with sporadic breast cancers and identified several altered regions (114 gains and 36 losses) in 18 patients. Their findings showed concordances with our results, presenting more often gains in 1q, 6p, 17q and frequent loss of 8p. Beside the well known effects of ERBB2 amplification in breast cancer development, the overexpression of STARD3 (located in the same locus) seems to be important, since it may contribute to increased proliferation, migration and invasion of breast cancer cells (as reviewed in [40]). Finally, considering our findings of altered regions found in BRCAX associated with metastasis (gain of 6p22.1 and loss of 6q25.1), gain of 6p was previously associated with BRCAX, and loss of 6q with BRCA1 tumors [41]. In fact, there are several members of RAET and ULBP family present in this locus. These members are ligands of C-type lectin-like receptor NKG2D, present in NK and T cells subsets, highly involved in tumor immunosurveillance [42]. Therefore, the loss of this region may have led to lower expression of these ligands, leading to less immunogenicity of the tumor cells. In fact, there are reports on colorectal cancer that have demonstrated this same pattern, and several authors discuss the potential therapeutic utility of NKG2D ligands in the treatment of this disease [42–44]. Therefore, these alterations on chromosome 6 seem to be highly associated with breast cancer tumors and may be of interest for further studies.
We also found that loss of 22q13.31–13.32 was significantly associated with presence of ovarian tumors (in the proband or in the family). The loss of heterozygosis (LOH) of chromosome 22q has been reported in a variety of cancers, including ovarian cancers, where the LOH rates reached 70% of cases [45, 46]. Study published by Zweemer el al. (2001) reported a significant loss of 22q, identified through aCGH in familial ovarian tumors [47]. Interestingly, MIR3201 was significantly downregulated in recurrent epithelial ovarian cancer (EOC), when compared to primary EOC [48]. To the best of our knowledge, there are no studies pointing to the functional relevance of MIR3201 in ovarian cancer, however, further studies may be performed to evaluate its possible role as a biomarker of EOC recurrence.
In summary, our findings support previous data of BRCAX related alterations and point to new regions potentially associated with personal and family history of ovarian cancer. In the present study, we could identify by aCGH analysis a potentially BRCAX-associated ovarian region on chromosome 22. Given our limited sample size, further work should be performed in order to validate our findings, to identify the driver genes associated with the BRCAX tumor development, as well as to uncover the role of those altered regions in cancer formation and progression.
MATERIALS AND METHODS
Ethics statement
All participants gave their consent to use tumor samples for academic genetic research. In addition, the ethics committee of the Barretos Cancer Hospital (BCH) approved this study (approval number: 916/2015).
Patients
This study included 31 unrelated Brazilian women at-risk for hereditary breast and ovarian cancer from the Oncogenetics Department of BCH. Those women were referred from the Oncogenetics Department of BCH for BRCA1, BRCA2 and TP53 genetic testing due to the presence of clinical criteria for HBOC, but no genetic alterations in these genes were found. For the purpose of the present study, were included only families fulfilling the following criteria: patients diagnosed with breast/ovarian cancer at an early age (<55 years), with at least two relatives with breast and/or ovarian cancer, two or more generations affected by cancer and absence of male BC.
Clinical information was obtained through detailed review of the patient´s clinical chart. For family history data, all pedigrees were revised.
Sequencing of BRCA1, BRCA2 and TP53
Analysis of the presence of germline mutations in BRCA1/BRCA2/TP53 genes was conducted at the Center of Molecular Diagnosis of BCH as part of routine care through NGS sequencing followed by rearrangement analysis through MLPA (Multiplex Ligation-dependent Probe Amplification Analysis), as described elsewhere by Fernandes et al. [49].
Tumor samples
For aCGH analysis, a representative section of FFPE tumor tissue from the breast or the ovarian tumor was stained by hematoxylin and eosin (H&E) and evaluated by a pathologist to verify tumor content (>70% tumor) and further microdissection.
DNA isolation and quality control
Following microdissection, DNA extraction steps were carried out using DNeasy Blood and Tissue kit (Qiagen), following the manufacturer’s instructions. The quality and integrity of the extracted DNA was assessed by multiplex PCR reaction using four primer pairs for the GAPDH gene (amplifying 100, 200, 300 and 400 bp, respectively), as described by Van Beers et al. [50]. The PCR reaction carried out contained (in a final volume of 30 μL) 1.5 mM MgCl2; 0.2 mM dNTP (Invitrogen); 0.133 μM of each primer; 1 U Taq DNA polymerase (Invitrogen) and 60 ng of tumor DNA. Reactions were performed in a Veriti thermocycler (Thermo Fisher Scientific) using the following amplification parameters: 94° C for 1 minute, 35 cycles of 94° C for 1 minute, 56° C for 1 minute, and 72° C for 3 minutes. Finally, a final extension at 72° C for 7 minutes. Amplification of DNA was verified by agarose gel electrophoresis.
Array comparative genomic hybridization
aCGH was performed on oligonucleotide-based SurePrint G3 Unrestricted CGH 8 × 60 K microarray slides, according the protocol provided by the manufacturer. In brief, 1 μg in final volume of 13 μL of normal female control DNA – reference DNA (DNA universal control-Promega Madison WI USA- Woman Reference: G152A) and patient’s DNA were differentially labeled with Cy3 (cyanine 3-deoxyuridine triphosphate) and Cy5 (cyanine 5-deoxyuridina triphosphate), respectively, using Agilent SureTag Complete DNA Labeling Kit (Agilent Technologies). Labeled DNA was then cleaned with purification columns (Agilent Technologies) and hybridized on array at 65° C for 24 hours, according to manufacturer’s recommendations. Microarrays were washed using Agilent Oligo aCGH Wash Buffers and scanning was performed using Agilent SureScan Microarray Scanner according to manufacturer’s instructions (Agilent Technologies).
Data analysis
Data quantification of aCGH was performed with Feature Extraction software (Agilent Technologies) and the txt output files were imported into Nexus Copy Number v8.0 (BioDiscovery Inc) for visualization and downstream analysis. BioDiscovery’s FASST2 Segmentation Algorithm, a Hidden Markov Model (HMM) based approach, was used to make copy number calls. The FASST2 algorithm, unlike other common HMM methods for copy number estimation, does not aim to estimate the copy number state at each probe but uses many states to cover more possibilities, such as mosaic events. These state values are then used to make calls based on a log-ratio threshold. The significance threshold for segmentation was set at 5.0E-6 also requiring a minimum of 3 probes per segment and a maximum probe spacing of 1,000 kb between adjacent probes before breaking a segment. The log ratio thresholds for single copy gain (or amplification) and single copy loss (or deletion) were set at 0.2 and -0.23, respectively. The log ratio thresholds for two or more copy gain (or high copy gain) and homozygous loss (or high copy loss) were set at 1.14 and -1.1 respectively. A 3:1 sex chromosome gain threshold was set to 1.2 and a 4:1 sex chromosome gain threshold was set to 1.7. Male sex chromosome big loss threshold was set to -1.1. GISTIC (Genomic Identification of Significant Targets in Cancer) algorithm was used within Nexus 8.0 to identify regions that are significantly amplified or deleted across a set of samples. It was considered the default parameters of Q-bound ≤ 0.05 with False Discovery Rate (FDR) correction and G-score cut-off ≤ 1.0. The identification of genes and CNVs were also performed within Nexus 8.0, being CNVs filtered according to 1000 genomes project. It was calculated the frequency of the gained and lost remaining CNVs and further separated according to <1% (rare CNVs) and ≥1% (common CNVs). The peaks identified by GISTIC algorithm were associated to breast and ovarian cancer family history and clinical characteristics, i.e. clinical staging, age at diagnosis (≤30, 31–45 and ≥45 years), molecular subtype, histological subtype, presence of metastasis and recurrence. These analyses were done by Fisher’s exact test (within SPSS v.21.0 software for Windows (Chicago, IL) considering the significance level of 5%.
Besides, the genomic regions found to be significant in GISTIC were considered for further analysis using the professional version of the compendium of cancer transcriptome profiles, Oncomine™ (Compendia Bioscience, Ann Arbor, MI). There were selected 13 breast and 5 ovarian cancer datasets (totalizing over 4000 samples). For each cancer type (breast or ovary), we selected the genes that presented gain or loss in our aCGH, and considered relevant those that presented gain in our aCGH and overexpression in Oncomine (P < 0.01), or those that presented loss in our aCGH and loss of expression in Oncomine (P < 0.01).
Abbreviations
aCGH: Array comparative genomic hybridization; BC: breast cancer; BCH: Barretos Cancer Hospital; ER: estrogen receptor; HBOC: Hereditary Breast and Ovarian Cancer Predisposition Syndrome; HER2: Human Epidermal growth factor Receptor 2; PR: progesterone receptor.
ACKNOWLEDGMENTS
The authors thank the patients, the Pathology Department and the Center of Molecular Diagnosis of Barretos Cancer Hospital.
CONFLICTS OF INTEREST
The authors declare that they have no competing interests.
FUNDING
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP - 2013/24633-2). EIP is recipient of a National Council of Technological and Scientific Development (CNPq) scholarship.
REFERENCES
1. WHO. World Health Organization - Breast Cancer. 2016. available from: http://www.who.int/cancer/detection/breastcancer/en/.
2. Lynch HT, Snyder C, Lynch J. Hereditary breast cancer: practical pursuit for clinical translation. Ann Surg Oncol. 2012; 19:1723–31. https://doi.org/10.1245/s10434-012-2256-z.
3. Miki Y, Swensen J, Shattuck-Eidens D, Futreal PA, Harshman K, Tavtigian S, Liu Q, Cochran C, Bennett LM, Ding W, Bell R, Rosenthal J, Hussey C, et al. A strong candidate for the breast and ovarian cancer susceptibility gene BRCA1. Science. 1994; 266:66–71.
4. Wooster R, Neuhausen SL, Mangion J, Quirk Y, Ford D, Collins N, Nguyen K, Seal S, Tran T, Averill D. Localization of a breast cancer susceptibility gene, BRCA2, to chromosome 13q12-13. Science. 1994; 265:2088–90.
5. Gronwald J, Byrski T, Huzarski T, Oszurek O, Janicka A, Szymanska-Pasternak J, Gorski B, Menkiszak J, RzepkaGorska I, Lubinski J. Hereditary breast and ovarian cancer. Hered Cancer Clin Pract. 2008; 6:88–98. https://doi.org/10.1186/1897-4287-6-2-88.
6. Petrucelli N, Daly MB, Pal T. BRCA1- and BRCA2-Associated Hereditary Breast and Ovarian Cancer. In: Pagon RA, Adam MP, Ardinger HH, Wallace SE, Amemiya A, Bean LJH, Bird TD, Ledbetter N, Mefford HC, Smith RJH and Stephens K, eds. GeneReviews(R). (Seattle (WA). 1993. Available from: https://www.ncbi.nlm.nih.gov/books/NBK1247/.
7. Friedenson B. BRCA1 and BRCA2 pathways and the risk of cancers other than breast or ovarian. MedGenMed. 2005; 7:60.
8. Couch FJ, Nathanson KL, Offit K. Two decades after BRCA: setting paradigms in personalized cancer care and prevention. Science. 2014; 343:1466–70. https://doi.org/10.1126/science.1251827.
9. Mavaddat N, Antoniou AC, Easton DF, Garcia-Closas M. Genetic susceptibility to breast cancer. Mol Oncol. 2010; 4:174–91. https://doi.org/10.1016/j.molonc.2010.04.011.
10. Tanic M, Andres E, Rodriguez-Pinilla SM, Marquez-Rodas I, Cebollero-Presmanes M, Fernandez V, Osorio A, Benitez J, Martinez-Delgado B. MicroRNA-based molecular classification of non-BRCA1/2 hereditary breast tumours. Br J Cancer. 2013; 109:2724–34. https://doi.org/10.1038/bjc.2013.612.
11. Honrado E, Osorio A, Milne RL, Paz MF, Melchor L, Cascon A, Urioste M, Cazorla A, Diez O, Lerma E, Esteller M, Palacios J, Benitez J. Immunohistochemical classification of non-BRCA1/2 tumors identifies different groups that demonstrate the heterogeneity of BRCAX families. Mod Pathol. 2007; 20:1298–306. https://doi.org/10.1038/modpathol.3800969.
12. Hopper JL. More breast cancer genes? Breast Cancer Res. 2001; 3:154–7.
13. Oldenburg RA, Meijers-Heijboer H, Cornelisse CJ, Devilee P. Genetic susceptibility for breast cancer: how many more genes to be found? Crit Rev Oncol Hematol. 2007; 63:125–49. https://doi.org/10.1016/j.critrevonc.2006.12.004.
14. Mangia A, Chiarappa P, Tommasi S, Chiriatti A, Petroni S, Schittulli F, Paradiso A. Genetic heterogeneity by comparative genomic hybridization in BRCAx breast cancers. Cancer Genet Cytogenet. 2008; 182:75–83. https://doi.org/10.1016/j.cancergencyto.2008.01.002.
15. Lacroix M, Leclercq G. The “portrait” of hereditary breast cancer. Breast Cancer Res Treat. 2005; 89:297–304. https://doi.org/10.1007/s10549-004-2172-4.
16. Rahman N, Teare MD, Seal S, Renard H, Mangion J, Cour C, Thompson D, Shugart Y, Eccles D, Devilee P, Meijers H, Nathanson KL, Neuhausen SL, et al. Absence of evidence for a familial breast cancer susceptibility gene at chromosome 8p12-p22. Oncogene. 2000; 19:4170–3. https://doi.org/10.1038/sj.onc.1203735.
17. Imbert A, Chaffanet M, Essioux L, Noguchi T, Adelaide J, Kerangueven F, Le Paslier D, Bonaiti-Pellie C, Sobol H, Birnbaum D, Pebusque MJ. Integrated map of the chromosome 8p12-p21 region, a region involved in human cancers and Werner syndrome. Genomics. 1996; 32:29–38.
18. Nathanson KL, Wooster R, Weber BL. Breast cancer genetics: what we know and what we need. Nat Med. 2001; 7:552–6. https://doi.org/10.1038/87876.
19. Gronwald J, Jauch A, Cybulski C, Schoell B, Bohm-Steuer B, Lener M, Grabowska E, Gorski B, Jakubowska A, Domagala W, Chosia M, Scott RJ, Lubinski J. Comparison of genomic abnormalities between BRCAX and sporadic breast cancers studied by comparative genomic hybridization. Int J Cancer. 2005; 114:230–6. https://doi.org/10.1002/ijc.20723.
20. Jonsson G, Naylor TL, Vallon-Christersson J, Staaf J, Huang J, Ward MR, Greshock JD, Luts L, Olsson H, Rahman N, Stratton M, Ringner M, Borg A, et al. Distinct genomic profiles in hereditary breast tumors identified by array-based comparative genomic hybridization. Cancer Res. 2005; 65:7612–21. https://doi.org/10.1158/0008-5472.CAN-05-0570.
21. Naylor TL, Greshock J, Wang Y, Colligon T, Yu QC, Clemmer V, Zaks TZ, Weber BL. High resolution genomic analysis of sporadic breast cancer using array-based comparative genomic hybridization. Breast Cancer Res. 2005; 7:R1186-98. https://doi.org/10.1186/bcr1356.
22. Nessling M, Richter K, Schwaenen C, Roerig P, Wrobel G, Wessendorf S, Fritz B, Bentz M, Sinn HP, Radlwimmer B, Lichter P. Candidate genes in breast cancer revealed by microarray-based comparative genomic hybridization of archived tissue. Cancer Res. 2005; 65:439–47.
23. Fridlyand J, Snijders AM, Ylstra B, Li H, Olshen A, Segraves R, Dairkee S, Tokuyasu T, Ljung BM, Jain AN, McLennan J, Ziegler J, Chin K, et al. Breast tumor copy number aberration phenotypes and genomic instability. BMC Cancer. 2006; 6:96. https://doi.org/10.1186/1471-2407-6-96.
24. van Beers EH, van Welsem T, Wessels LF, Li Y, Oldenburg RA, Devilee P, Cornelisse CJ, Verhoef S, Hogervorst FB, van’t Veer LJ, Nederlof PM. Comparative genomic hybridization profiles in human BRCA1 and BRCA2 breast tumors highlight differential sets of genomic aberrations. Cancer Res. 2005; 65:822–7.
25. Didraga MA, van Beers EH, Joosse SA, Brandwijk KI, Oldenburg RA, Wessels LF, Hogervorst FB, Ligtenberg MJ, Hoogerbrugge N, Verhoef S, Devilee P, Nederlof PM. A non-BRCA1/2 hereditary breast cancer sub-group defined by aCGH profiling of genetically related patients. Breast Cancer Res Treat. 2011; 130:425–36. https://doi.org/10.1007/s10549-011-1357-x.
26. Alvarez C, Aravena A, Tapia T, Rozenblum E, Solis L, Corvalan A, Camus M, Alvarez M, Munroe D, Maass A, Carvallo P. Different Array CGH profiles within hereditary breast cancer tumors associated to BRCA1 expression and overall survival. BMC Cancer. 2016; 16:219. https://doi.org/10.1186/s12885-016-2261-x.
27. Letessier A, Sircoulomb F, Ginestier C, Cervera N, Monville F, Gelsi-Boyer V, Esterni B, Geneix J, Finetti P, Zemmour C, Viens P, Charafe-Jauffret E, Jacquemier J, et al. Frequency, prognostic impact, and subtype association of 8p12, 8q24, 11q13, 12p13, 17q12, and 20q13 amplifications in breast cancers. BMC Cancer. 2006; 6:245. https://doi.org/10.1186/1471-2407-6-245.
28. Yang ZQ, Streicher KL, Ray ME, Abrams J, Ethier SP. Multiple interacting oncogenes on the 8p11-p12 amplicon in human breast cancer. Cancer Res. 2006; 66:11632–43. https://doi.org/10.1158/0008-5472.CAN-06-2946.
29. Bernard-Pierrot I, Gruel N, Stransky N, Vincent-Salomon A, Reyal F, Raynal V, Vallot C, Pierron G, Radvanyi F, Delattre O. Characterization of the recurrent 8p11-12 amplicon identifies PPAPDC1B, a phosphatase protein, as a new therapeutic target in breast cancer. Cancer Res. 2008; 68:7165–75. https://doi.org/10.1158/0008-5472.CAN-08-1360.
30. Gelsi-Boyer V, Orsetti B, Cervera N, Finetti P, Sircoulomb F, Rouge C, Lasorsa L, Letessier A, Ginestier C, Monville F, Esteyries S, Adelaide J, Esterni B, et al. Comprehensive profiling of 8p11-12 amplification in breast cancer. Mol Cancer Res. 2005; 3:655–67. https://doi.org/10.1158/1541-7786.MCR-05-0128.
31. Chen H, Singh RR, Lu X, Huo L, Yao H, Aldape K, Abraham R, Virani S, Mehrotra M, Mishra BM, Bousamra A, Albarracin C, Wu Y, et al. Genome-wide copy number aberrations and HER2 and FGFR1 alterations in primary breast cancer by molecular inversion probe microarray. Oncotarget. 2017; 8:10845–57. https://doi.org/10.18632/oncotarget.14802.
32. Luo J, Liu S, Leung S, Gru AA, Tao Y, Hoog J, Ho J, Davies SR, Allred DC, Salavaggione AL, Snider J, Mardis ER, Nielsen TO, et al. An mRNA Gene Expression-Based Signature to Identify FGFR1-Amplified Estrogen Receptor-Positive Breast Tumors. J Mol Diagn. 2017; 19:147–61. https://doi.org/10.1016/j.jmoldx.2016.09.007.
33. Shi YJ, Tsang JY, Ni YB, Chan SK, Chan KF, Tse GM. FGFR1 is an adverse outcome indicator for luminal A breast cancers. Oncotarget. 2016; 7:5063–73. https://doi.org/10.18632/oncotarget.6563.
34. Ni HJ, Chang YN, Kao PH, Chai SP, Hsieh YH, Wang DH, Fong JC. Depletion of SUMO ligase hMMS21 impairs G1 to S transition in MCF-7 breast cancer cells. Biochim Biophys Acta. 2012; 1820:1893–900. https://doi.org/10.1016/j.bbagen.2012.08.002.
35. Lundgren K, Holm K, Nordenskjold B, Borg A, Landberg G. Gene products of chromosome 11q and their association with CCND1 gene amplification and tamoxifen resistance in premenopausal breast cancer. Breast Cancer Res. 2008; 10:R81. https://doi.org/10.1186/bcr2150.
36. Sheen-Chen SM, Huang CY, Liu YY, Huang CC, Tang RP. Cortactin in breast cancer: analysis with tissue microarray. Anticancer Res. 2011; 31:293–7.
37. Ni QF, Yu JW, Qian F, Sun NZ, Xiao JJ, Zhu JW. Cortactin promotes colon cancer progression by regulating ERK pathway. Int J Oncol. 2015; 47:1034–42. https://doi.org/10.3892/ijo.2015.3072.
38. Karlsson E, Waltersson MA, Bostner J, Perez-Tenorio G, Olsson B, Hallbeck AL, Stal O. High-resolution genomic analysis of the 11q13 amplicon in breast cancers identifies synergy with 8p12 amplification, involving the mTOR targets S6K2 and 4EBP1. Genes Chromosomes Cancer. 2011; 50:775–87. https://doi.org/10.1002/gcc.20900.
39. Tasian SK, Loh ML. Understanding the biology of CRLF2-overexpressing acute lymphoblastic leukemia. Crit Rev Oncog. 2011; 16:13–24.
40. Jacot W, Fiche M, Zaman K, Wolfer A, Lamy PJ. The HER2 amplicon in breast cancer: Topoisomerase IIA and beyond. Biochim Biophys Acta. 2013; 1836:146–57. https://doi.org/10.1016/j.bbcan.2013.04.004.
41. Kamieniak MM, Munoz-Repeto I, Rico D, Osorio A, Urioste M, Garcia-Donas J, Hernando S, Robles-Diaz L, Ramon YCT, Cazorla A, Saez R, Garcia-Bueno JM, Domingo S, et al. DNA copy number profiling reveals extensive genomic loss in hereditary BRCA1 and BRCA2 ovarian carcinomas. Br J Cancer. 2013; 108:1732–42. https://doi.org/10.1038/bjc.2013.141.
42. Rothe A, Jachimowicz RD, Borchmann S, Madlener M, Kessler J, Reiners KS, Sauer M, Hansen HP, Ullrich RT, Chatterjee S, Borchmann P, Yazaki P, Koslowsky TC, et al. The bispecific immunoligand ULBP2-aCEA redirects natural killer cells to tumor cells and reveals potent anti-tumor activity against colon carcinoma. Int J Cancer. 2014; 134:2829–40. https://doi.org/10.1002/ijc.28609.
43. Ramutton T, Buccheri S, Dieli F, Todaro M, Stassi G, Meraviglia S. γδ T cells as a potential tool in colon cancer immunotherapy. Immunotherapy. 2014; 6:989–99. https://doi.org/10.2217/imt.14.59.
44. Xia Y, Chen B, Shao X, Xiao W, Qian L, Ding Y, Ji M, Gong W. Treatment with a fusion protein of the extracellular domains of NKG2D to IL-15 retards colon cancer growth in mice. J Immunother. 2014; 37:257–66. https://doi.org/10.1097/CJI.0000000000000033.
45. Castells A, Ino Y, Louis DN, Ramesh V, Gusella JF, Rustgi AK. Mapping of a target region of allelic loss to a 0.5-cM interval on chromosome 22q13 in human colorectal cancer. Gastroenterology. 1999; 117:831–7.
46. Cliby W, Ritland S, Hartmann L, Dodson M, Halling KC, Keeney G, Podratz KC, Jenkins RB. Human epithelial ovarian cancer allelotype. Cancer Res. 1993; 53:2393–8.
47. Zweemer RP, Ryan A, Snijders AM, Hermsen MA, Meijer GA, Beller U, Menko FH, Jacobs IJ, Baak JP, Verheijen RH, Kenemans P, van Diest PJ. Comparative genomic hybridization of microdissected familial ovarian carcinoma: two deleted regions on chromosome 15q not previously identified in sporadic ovarian carcinoma. Lab Invest. 2001; 81:1363–70.
48. Chong GO, Jeon HS, Han HS, Son JW, Lee YH, Hong DG, Lee YS, Cho YL. Differential MicroRNA Expression Profiles in Primary and Recurrent Epithelial Ovarian Cancer. Anticancer Res. 2015; 35:2611–7.
49. Palmero EI, Galvao HC, Fernandes GC, Paula AE, Oliveira JC, Souza CP, Andrade CE, Romagnolo LG, Volc S, Neto MC, Sabato C, Grasel R, Mauad E, et al. Oncogenetics service and the Brazilian public health system: the experience of a reference Cancer Hospital. Genet Mol Biol. 2016; 39:168–77. https://doi.org/10.1590/1678-4685-GMB-2014-0364.
50. van Beers EH, Joosse SA, Ligtenberg MJ, Fles R, Hogervorst FB, Verhoef S, Nederlof PM. A multiplex PCR predictor for aCGH success of FFPE samples. Br J Cancer. 2006; 94:333–7. https://doi.org/10.1038/sj.bjc.6602889.