
INTRODUCTION
Staphylococcus aureus is the leading Gram-positive bacterium that can cause infections in humans worldwide, including mild skin infections, bacteremia, sepsis, and endocarditis [1–3]. Over the last century, infections with S. aureus have become increasingly difficult to treat owing to the emergence and rapid spread of multiple-drug resistant S. aureus [4–6].
Macrolides, which consist of a 14- to 16-membered lactone ring with different appended sugars and comprise a key group of inhibitors of bacterial translation, are ribosome-targeting antibiotics used to treat infections caused by Staphylococcus species [7, 8]. Erythromycin, azithromycin, and clarithromycin are members of the macrolide antibiotics, a large group of antibacterial agents that include natural or newer semi-synthetic compounds [9, 10]. Their inhibitory activity depends on binding to a site near the ribosomal nascent peptide exit tunnel, which starts at the peptidyl transferase center and spans the body of the large ribosomal subunit, thereby halting translation of a particular subset of nascent peptides [11–13].
Resistance to macrolide may be mediated by three primary mechanisms: a) modification of ribosomes, such as dimethylation of a unique adenine residue in the 23S ribosomal RNA (rRNA), A2085 in S. aureus (corresponding to E. coli A2058), which is located in the macrolide-binding site in the nascent peptide exit tunnel, by the erythromycin resistance methyltransferase encoded by the erm genes [14–16]; b) activated efflux systems, involving a member of the ATP-binding cassette (ABC) family of transporters encoded by the macrolide-streptogramins resistance A (msrA) gene, keeping intracellular antibiotic concentration at a subtoxic level and conferring inducible resistance to erythromycin and type B streptogramins in staphylococci [17, 18]; and c) production of antibiotic-inactivating enzymes, such as phosphorylase, a macrolide phosphotransferase C (encoded by mphC in staphylococci) that inactivates antibiotics [19, 20]. According to other studies, mutations in Escherichia coli rplV and rplD genes coding for ribosomal proteins L22 and L4, respectively, can also confer resistance to macrolide antibiotics [21, 22]. A mutant change in rplV was also observed in antibiotic-resistant S. aureus [23].
Here, we screened a resistant strain obtained by culturing the sensitive S. aureus strain 8325-4 in the presence of erythromycin. A 27-nt repeat sequence insertion in the rplV (rplVindel) gene was identified in this erythromycin-resistant strain, which induced specific resistance to macrolides.
RESULTS
An erythromycin-resistant strain of S. aureus 8325-4 is screened in vitro
To explore the mechanism underlying the resistance to macrolide in S. aureus, we cultured wild-type S. aureus 8325-4 in BHI medium while continuously doubling the concentration of erythromycin (Figure 1A). An isolate with acquired resistance to erythromycin was screened and named 8325ER+. It was able to grow in a medium containing 80 μg/mL of erythromycin, and the survival rates of 8325ER+ strain in different concentration of erythromycin were significantly higher than the parent strain (Figure 1B). Besides, the minimal inhibitory concentration (MIC) of erythromycin of 8325ER+ was 160 μg/mL in a drug susceptibility test, which was interpreted as erythromycin resistant according to Clinical and Laboratory Standards Institute (CLSI) criteria [24].
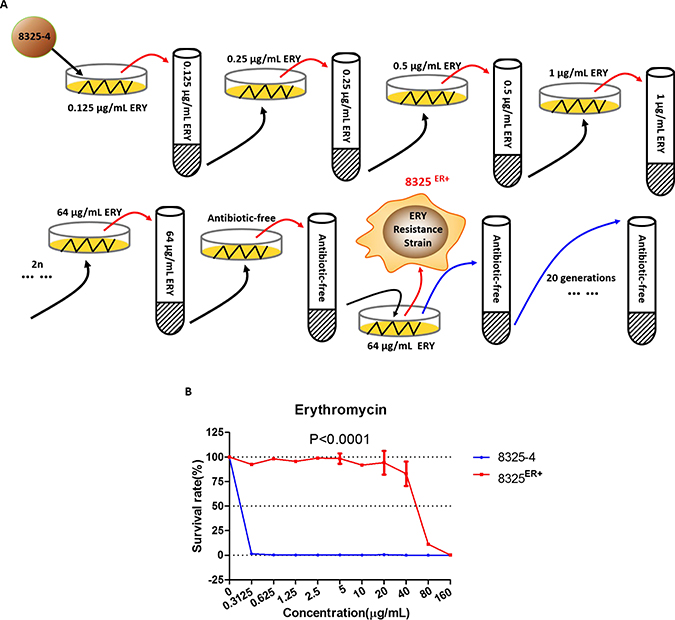
Figure 1: Erythromycin-resistant strain 8325ER+. (A) Schematic diagram of stepwise screening for resistance to erythromycin (ERY) of wild-type S. aureus. 8325–4 was cultured and passaged in BHI medium supplemented with various concentrations of erythromycin (initially 0.125 μg/mL, followed by two-fold increased until the concentration of erythromycin reached 64 μg/mL). Each screening step included solid and liquid BHI medium for screening. Solid medium was used for picking an isolate of S. aureus at 37° C incubator for 12 h, and liquid medium was used for enrichment of the isolate at 37° C with shaking at 220 rpm. When screening was completed, the erythromycin-resistant isolate was inoculated in BHI medium without antibiotics for 20 generations. (B) Survival rates of 8325–4 and 8325ER+ in different concentration of erythromycin. The survival curve of wild-type 8325–4 is shown in blue and the 8325ER+ strain in red. Values are the means of triplicate wells; error bars indicate SD.
In the further investigation, 8325ER+ was cultured in BHI broth without erythromycin for 20 generations, the susceptibility to erythromycin of bacteria from different generations was determined individually. We found that the MIC of S. aureus from different generations was not altered, which suggested that resistance to erythromycin in 8325ER+ was inheritable.
Whole-genome sequencing identifies gene mutations in 8325ER+
To test whether high expression of known erythromycin resistance genes in 8325ER+ contributed to the resistance to erythromycin, we extracted total RNA from the erythromycin-sensitive strain 8325-4 and the erythromycin-resistant strain 8325ER+ (Supplementary Figure 1A). Reverse transcription-polymerase chain reaction (RT-PCR) showed that erythromycin resistance-related genes, including 23 S rRNA adenine-specific N-methyltransferases (encoded by ermA/ermB/ermC), mphC, and msrA were not detected in 8325ER+ (Supplementary Figure 1B). These results suggest that another mechanism is responsible for the resistance to erythromycin of 8325ER+.
To investigate the potential genes involved in the resistance occurrence to erythromycin, we extracted total genomic DNA and compared the genome sequence of 8325ER+ with that of 8325-4.
Sequence analysis showed that ten genes mutated, and six of them encoded different proteins (Table 1). Considering that five of the ten mutated genes were involved in the translation process, we decided to compare cell growth between 8325ER+ and 8325-4. We did not see a significant difference between both strains (Supplementary Figure 1C).
Table 1: Comparison of whole-genome sequence of 8325ER+ with 8325–4
Type of mutation |
Nucleotide |
Amino acid |
Locus tag |
Product |
Position in chromosome |
|---|---|---|---|---|---|
Point mutation |
T to A |
— |
— |
Noncoding |
75324 of CP000253.1 |
T 102 G |
G 34 G |
SAOUHSC_01078 |
ribosomal protein L32 |
1042000 of CP000253.1 |
|
C 1988 T |
S 663 L |
SAOUHSC_01583 |
conserved hypothetical phage protein |
1508580 of CP000253.1 |
|
C 530 G |
A 177 G |
SAOUHSC_01748 |
queuine tRNA-ribosyltransferase |
1653225 of CP000253.1 |
|
A 184 G |
R 62 G |
SAOUHSC_02163 |
conserved hypothetical phage protein |
2031924 of CP000253.1 |
|
G to A |
— |
SAOUHSC_R0005 |
16S ribosomal RNA |
2243146 of CP000253.1 |
|
A 208 C |
T 70 P |
SAOUHSC_02511 |
ribosomal protein L4 |
2316907 of CP000253.1 |
|
G 206 C |
G 69 A |
SAOUHSC_02511 |
ribosomal protein L4 |
2316909 of CP000253.1 |
|
A to G |
— |
— |
Noncoding |
2350008 of CP000253.1 |
|
Fragment insertion |
C 291 or 318 to |
KRTSHITIV |
SAOUHSC_02507 |
ribosomal protein L22 |
2314658 of CP000253.1 |
Further analysis showed that seven genes were identified with one or two nucleotide mutations, and only one gene, rplV, encoding ribosomal protein L22 exhibited an insertion of a 27-nt fragment (Table 1). In further investigation, we found that sequence of the insertion segment correspond to a duplication of the region 292–318 of rplV (Figure 2A). Moreover, the insertion of the 27-nt fragment happen at nucleotide 292C or 318C in rplV gene in 8325ER+ (Figure 2B). Furthermore, we found that the 27-nt fragment led to a 9-amino acids insertion but did not induce a frame-shifting mutation (Figure 2C).
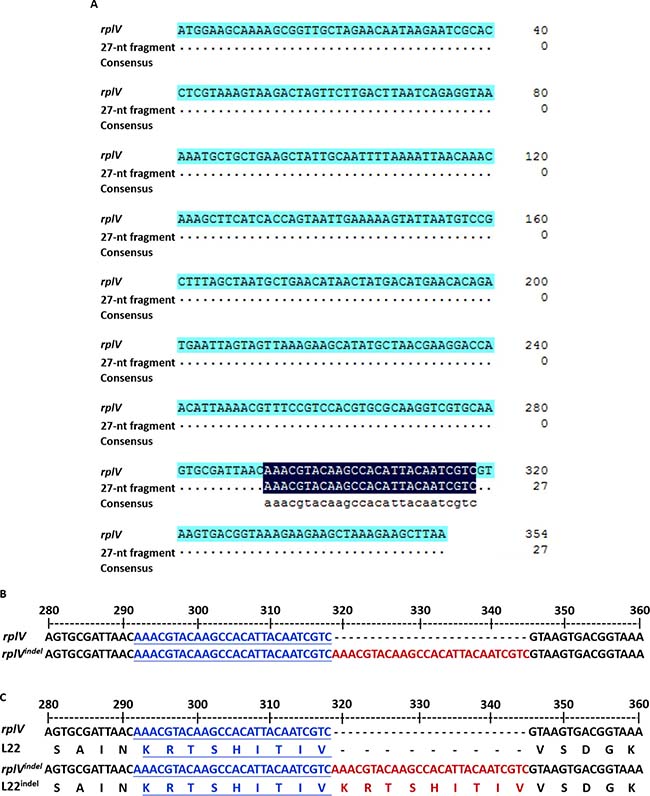
Figure 2: Insertion of a 27 nucleotide-repeat fragment in the rplV gene. (A) BLAST analysis of 27-nt fragment sequence. The top line shows the rplV gene, and the middle line shows 27-nt fragment sequences. BLAST results are shown for the total rplV nucleotides. (B) Nucleotides alignment of partial sequences of wild-type rplV and rplVindel to display the region 27-nt fragment insertion. The letters in blue and underlined on the middle line and bottom line are coincide with the inserted 27-nt fragment shown in red on the bottom line, and the 27-nt fragment is exactly adjacent to the region 292–318 of rplV. (C) Protein alignment of partial sequences of wild-type L22 and L22indel to display the inserted 27-nt fragment that led to a 9-amino acid insertion without frame-shifting mutation. The letters in blue and underlined are coincide with the inserted fragment shown in red on the line.
Twenty-seven-nucleotide insertion in the rplV (rplVindel) gene induces resistance to erythromycin in S. aureus
The rplV gene encodes the ribosomal 50S subunit protein L22, which is important for ribosomal 50S subunit assembly at the early stage. It is essential for the formation of the nascent peptide exit tunnel of the mature ribosome [22]. Given that mutation in the rplV gene was reported to be involved in resistance to antibiotics in E. coli and S. aureus [11, 21–23], we focused on investigating whether the rplVindel gene induced resistance to erythromycin. Firstly, the rplV genes were amplified by PCR from the genomes of 8325ER+ and 8325-4. We found the band of PCR product from 8325ER+ was bigger than that from 8325-4 (Figure 3A). Following analysis showed that the sequence of rplV gene in 8325ER+ containing the 27-nt insertion fragment, which was consistent with the genome sequence results (Figure 3B).
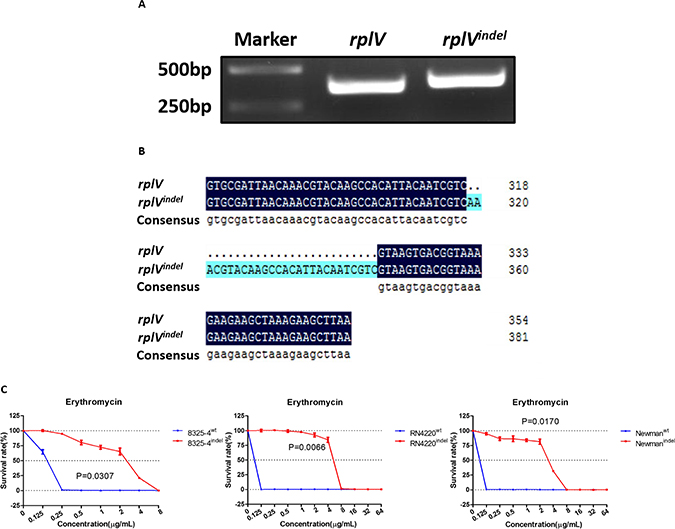
Figure 3: rplVindel gene contributes to resistance to erythromycin in drug-susceptible S. aureus. (A) rplV genes were amplified by PCR from the genomes of 8325–4 and 8325ER+, and PCR products were resolved on a 2% agarose gel and visualized by ultraviolet imaging. (B) The nucleotides were sequenced by Sangon Biotech, and BLAST analysis was performed using DNAMAN. The top line shows wild-type rplV, and the middle line shows rplVindel. BLAST results are shown for part of the total rplV nucleotides. (C) Survival rates of recombinant 8325–4, RN4220, and Newman cells in different concentration of erythromycin. Drug-susceptible S. aureus cells transformed with the rplVindel gene exhibit decreased sensitivity to erythromycin. The survival curve of cells harboring wild-type rplV gene is shown in blue and the rplVindel gene is shown in red. Values are the means of triplicate wells; error bars indicate SD.
To evaluate the role of rplVindel in raising resistance of S. aureus to erythromycin, we generated several erythromycin-susceptible S. aureus strains (8325-4indel, RN4220indel, and Newmanindel) with ectopic expression of rplVindel. Meanwhile, these strains transferred with the wild-type rplV gene were used as control (Supplementary Figure 2). We found that the survival rates of S. aureus strains with ectopic expression of rplVindel in different concentration of erythromycin were significantly higher than control strains (Figure 3C). And erythromycin MICs in 8325-4indel, RN4220indel, and Newmanindel were 8 μg/mL respectively (Table 2), which suggests that ectopic expression of rplVindel in susceptible strains induced resistance to erythromycin.
Table 2: Antimicrobial agent susceptibility of Staphylococcus strains
Antimicrobial Agents |
MIC (μg/mL)* |
|||||||
|---|---|---|---|---|---|---|---|---|
8325–4 |
RN4220 |
Newman |
||||||
8325–4 |
8325ER+ |
8325–4wt |
8325–4indel |
RN4220wt |
RN4220indel |
Newmanwt |
Newmanindel |
|
Erythromycin |
0.3125S |
160R |
0.25S |
8R |
0.125S |
8R |
0.125S |
8R |
Azithromycin |
0.78125S |
200R |
0.5S |
16R |
0.5S |
32R |
0.5S |
32R |
Clarithromycin |
0.1953125S |
100R |
0.125S |
8R |
0.125S |
8R |
0.125S |
8R |
Chloramphenicol |
5 |
5 |
− |
− |
− |
− |
− |
− |
Linezolid |
1.25 |
1.25 |
0.625 |
0.625 |
1 |
1 |
1 |
1 |
Tobramycin |
1.25 |
1.25 |
2.5 |
2.5 |
2 |
2 |
2 |
2 |
Kanamycin |
5 |
5 |
5 |
5 |
8 |
8 |
8 |
8 |
Vancomycin |
1.25 |
1.25 |
1 |
1 |
1 |
1 |
2 |
2 |
* The MICs of corresponding antibiotic shown with letter “R” represent “Resistant”, those with letter “S” represent “Susceptible” according to CLSI criteria.
rplVindel contributes to specific resistance to macrolides in S. aureus
Next, we asked whether the rplVindel gene could induce resistance to other macrolides, including azithromycin and clarithromycin. First, we also found the same results with erythromycin susceptibility test. The survival rates of 8325ER+ strain in different concentration of azithromycin and clarithromycin were significantly higher than parent strain (Figure 4A). We then determined the MIC of azithromycin and clarithromycin in 8325ER+ as 200 μg/mL and 100 μg/mL, respectively (Table 2). In line with expectation, the ectopic expression of rplVindel in susceptible strains also induced resistance to these two antibiotics (Figure 4B–4D, Table 2).
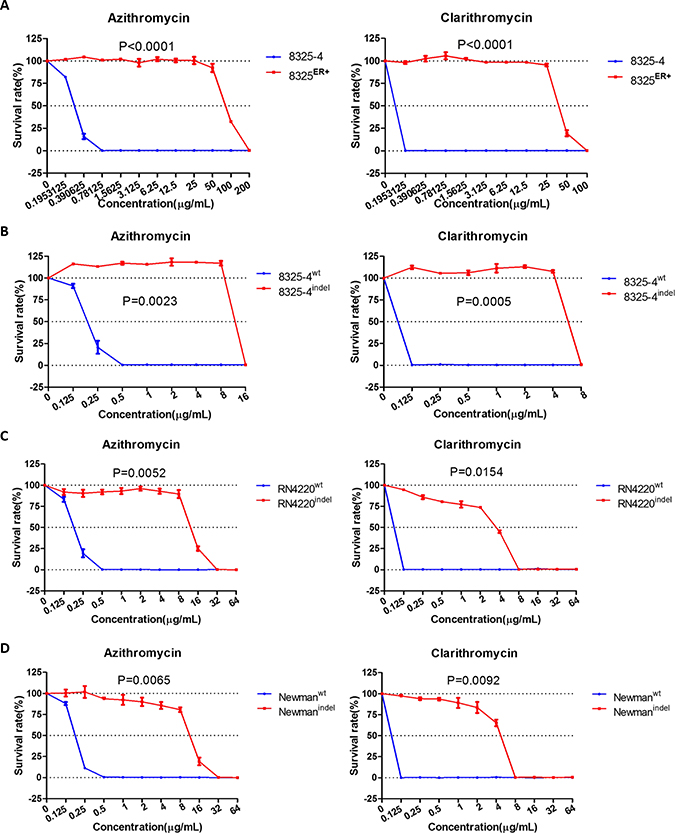
Figure 4: rplVindel gene contributes to resistance to macrolides in drug-susceptible S. aureus. (A) Survival rates of 8325 and 8325ER+ in different concentration of azithromycin (left) and clarithromycin (right). (B) Survival rates of wild-type S. aureus 8325–4 cell transformed with the rplVindel gene exhibit decreased sensitivity to azithromycin (left) and clarithromycin (right). (C) Survival rates of drug-susceptible RN4220 cell transformed with the rplVindel gene exhibit decreased sensitivity to azithromycin (left) and clarithromycin (right). (D) Survival rates of drug-susceptible Newman cell transformed with the rplVindel gene exhibit decreased sensitivity to azithromycin (left) and clarithromycin (right). The survival curve of cells harboring wild-type rplV gene is shown in blue and the rplVindel gene is shown in red. Values are the means of triplicate wells; error bars indicate SD.
As the ribosomal protein L22 is essential in formation of the ribosomal polypeptide exit tunnel [22], we then determined whether rplVindel was involved in resistance to antibiotics targeting the ribosome. It was revealed that neither 8325ER+ nor susceptible strains with ectopic expression of rplVindel were resistant to chloramphenicol and linezolid (Table 2), which target the 50 S ribosomal subunit. Similar results were also obtained in a drug susceptibility test of antibiotics that target the 30 S subunit or cell wall (Table 2). Consistent with the MICs of non-macrolide antibiotics of 8325ER+ strain or susceptible strains with ectopic expression of rplVindel, there were no significant differences observed among the survival rates of those S. aureus strains comparing with their control strains (Supplementary Figures 3A–3B, 4A–4C).
DISCUSSION
Macrolides are usually used in clinical therapy for skin infections caused by S. aureus. Several mechanisms involved in S. aureus resistance to macrolides have been revealed. In this study, we revealed that a 27-nt insertion in the rplV gene induced a specific resistance to macrolides.
The 8325ER+ resistance to macrolides was not due to the occurrence of identified erythromycin-resistant genes (ermA/ermB/ermC/mphC/msrA) but the rplVindel gene. Interestingly, the 27-nt insertion sequence is a repeat sequence of the rplV gene, but it did not induce frame-shifting mutation. Sequence analysis showed that this fragment might be inserted behind 291C or 318C. The ectopic expression of the rplVindel gene in several susceptible strains specifically conferred resistance to macrolide antibiotics. As shown in Table 2, the MIC of macrolides was higher in 8325ER+ than in 8325indel. We think this may be majorly due to the coexistence of rplV and rplVindel. The ribosome consisting of ribosomal protein L22 (rplV) is still sensitive to macrolides. Mutations in other genes may also contribute the resistance to macrolides in 8325ER+. These assumptions will be investigated in the future.
In 1967, bacterial resistance to macrolides, caused by mutations in ribosomal protein, was reported [25]. In E. coli, it has been found that the deletion of M82K83R84 increases expression of the AcrAB-TolC efflux system and results in resistance to macrolides [21, 26]. Here, we reveal that a 27-nt insertion in the rplV gene confers specific resistance to macrolides in S. aureus. However, the level of msrA, a well-identified gene of the efflux system involved in resistance to macrolides in staphylococci [17, 18], was not altered in 8325ER+ compared with that of 8325-4. The resistance to macrolides induced by rplVindel may be due to the conformational changes of L22 protein induced by the 27-nt insertion, which will be investigated in the future.
Ribosome protein L4 forms part of the lining of the peptide exit tunnel with L22. Mutations in ribosome protein L4 also induce macrolides resistance in a variety of pathogenic and non-pathogenic bacteria [21, 27–29]. There are two amino acids mutations (G69A, T70P) identified in the L4 protein from 8325ER+, which may also contribute to the macrolides resistance. Although the ectopic expression of the rplVindel gene in several susceptible strains specifically conferred resistance to macrolide antibiotics, combination with the mutant L4 protein may further elevate the resistance.
In this study, we tried to detect rplVindel in 84 clinical S. aureus isolates resistant to macrolides. Most of these strains harbor ermA/B/C genes (Supplementary Table 1). There was no strain identified that contained the rplVindel genes (data not shown). These results indicate that the occurrence of rplVindel in clinical isolates is rare compared with that of erm genes.
In conclusion, our findings present a 27-nt insertion in rplV that induces the specific resistance to macrolides in S. aureus.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions
The strains and plasmids used in this study are listed in Supplementary Table 2. 84 Clinical macrolides-resistant S. aureus obtained from Department of Clinical Laboratory, Peking University People’s Hospital. Distributions of the 84 clinical samples of S. aureus by origin of recovery were 22 strains from blood (26.2%), 11 strains from pus (13.1%), 14 strains from secretions (16.7%), 27 strains from sputum (32.1%), 9 strains from wound (10.7%) and one strain from abdominal fluid (1.2%). Strains were cultured using brain heart infusion (BHI) medium (BD) at 37° C for 12 h with shaking at 220 rpm. Clinical isolates and wild-type strains including 8325-4, RN4220 and Newman were cultured in antibiotic-free BHI broth, while 8325ER+ strain was cultured in BHI broth supplemented with 50 μg/mL erythromycin, those wild-type strains transformed with the shuttle plasmid pOS1 supplemented with 25 μg/mL chloramphenicol, E. coli strain transformed with a cloning plasmid pMD-19T supplemented with 100 μg/mL ampicillin.
Erythromycin screen in vitro
Schematic diagram of stepwise screening for resistance to erythromycin of wild-type S. aureus was showed in Figure 1A. 8325-4 was cultured and passaged in BHI medium supplemented with various concentrations of erythromycin (initially 0.125 μg/mL, followed by two-fold increased until the concentration of erythromycin reached 64 μg/mL). Each screening step included solid and liquid BHI medium for screening. Solid medium was used for picking an isolate of S. aureus at 37° C incubator for 12 h, and liquid medium was used for enrichment of the isolate at 37° C with shaking at 220 rpm. When screening was completed, the erythromycin-resistant isolate was inoculated in BHI medium without antibiotics for 20 generations.
Measurement of bacterial growth curve
Bacteria were incubated in BHI broth at 37° C with shaking at 220 rpm overnight. The concentration of bacteria was adjusted to 1 × 107 cfu/mL, then 1:100 inoculated in BHI broth without antibiotics at 37° C with shaking at 220 rpm for 12 hours. Growth curves of bacteria were constructed by measuring of the cell density at A600 nm at one-hour intervals for 12 hours.
Antibiotic susceptibility assay
Antibiotics were purchased from Selleck. Susceptibility to antibiotics was tested by using broth microdilution method according to the Clinical and Laboratory Standards Institute (CLSI) [24]. Briefly, antibiotics were prepared by serial two-fold dilutions in BHI broth, then various concentration of antibiotics were made in triplicate in 96-well culture dishes containing 1 × 105 cfu/well bacteria and incubated for 18–24 h at 37° C. Control wells were free of antibiotic. Bacteria growth was determined by reading the optical density (OD) at 630 nm. The survival rates of bacteria were calculated by the rates of OD 630 nm measurement at each concentration of antibiotic versus control wells. The MIC was determined to be the dose of antibiotic that inhibited bacteria growth by >95%.
RNA isolation and RT-PCR
For detecting erythromycin resistance genes in S. aureus. Total bacterial RNA was extracted from S. aureus, which were grown with shaking at 37° C using Trizol (Invitrogen) as previously described [30]. Briefly, DNase digestion of 80 μL of total RNA was performed with 10U of RNase-free DNase I (Promega) and 10 μL of the 10 × reaction buffers in a total reaction volume of 100 μL for 30 min at 37° C. For cDNA synthesis, 6 μL total RNA (≈250 ng) was incubated at 65° C for 5 min, then add 2 μL of 4 × DNA remove buffer and incubate at 37° C for 5 min, finally add 2 μL of 5 × RT Master MixII (TOYOBO) and incubate at 37° C for 15 min, 50° C for 5 min, 98° C for 5 min.
Detection of macrolides-resistance genes
Macrolides resistance genes ermA, ermB, ermC, msrA and mphC were examined in the erythromycin-sensitive strain 8325-4 and the erythromycin-resistant strain 8325ER+ with primers listed in Supplementary Table 3. The PCR reaction mixture contained 2.5 μL of 10 × PCR reaction buffer, 0.25 μL enzyme, 0.5 μL dNTP mix, 0.3 mM of gene-specific forward and reverse primers, and 2 μL of template, made up to a final volume of 25 μL with distilled water. Cycling parameters were set as follows: initial activation step at 95° C for 5 min, denaturation at 95° C for 30 s, annealing at 55° C for 30 s, and extension at 72° C for 30 s. gyrB was used as the endogenous reference gene. The PCR products were resolved in 2% agarose gel and visualized by ultraviolet imaging.
Whole-genome sequencing of S. aureus
Bacteria were grown in BHI broth at 37° C for 12 h with shaking at 220 rpm and harvested by centrifuge at 12000 rpm for 1 min. Genomic DNA was extracted by using EasyPure® Bacteria Genomic DNA Kit (TransGen Biotech) according to manufacturer’s instruction. Sequencing with constructed shotgun libraries of 8325-4 and 8325ER+ was performed by Illumina Hiseq 2000. Fragmentation, library construction, and sequencing were carried out by oebiotech company.
Analysis of the rplV gene in clinical isolates
For detecting 27-nt fragment insertion in rplV gene in clinical macrolides resistant S. aureus isolates, the rplVindel gene was detected by PCR amplification. Primers used for the rplV gene were rplV-F and rplV-R listed in Supplementary Table 3. Clinical isolates were grown on blood agar plates and incubated overnight at 37° C, bacteria DNA was prepared by suspending a fresh colony in 400 μL of sterile distilled water and heating at 100° C for 10 min and then centrifuged at 12000 rpm for 5 min. PCR amplification was carried out under the following conditions: 95° C for 5 min, followed by 34 cycles of 95° C for 30 s, 56° C for 30 s, 72° C for 30 s, and 72° C for 5 min. PCR products were resolved on a 2% agarose gel and visualized by ultraviolet imaging. The nucleotides were sequenced by Sangon Biotech, and BLAST analysis was performed using DNAMAN.
Ectopic expression of rplVindel in S. aureus strains
rplV genes were amplified by PCR from the genomes of wild-type S. aureus 8325-4 and the erythromycin-resistant strain 8325ER+ with primers rplV-F-EcoRI and rplV-R-BamHI (Supplementary Table 3). The PCR products were ligated into pMD-19T vector, the recombinant plasmids were transformed into DH5ɑ. The recombinant pMD-19T plasmid was eliminated by cutting the plasmid with the EcoRI and BamHI restriction enzymes, then digested fragments were ligated into EcoRI and BamHI-digested pOS1 vector. The recombinant plasmids were transformed into DH5ɑ, then electro-transformation into recipient strains S. aureus RN4220. The plasmid was isolated from RN4220, then electro-transformation into S. aureus 8325-4 and Newman.
Statistical analyses
Statistical tests were performed using GraphPad Prism v.5.0 (GraphPad Software Inc., San Diego, CA, United States). The Differences between survival curves were evaluated for statistical significance using the unpaired t test. All P-values of ≤ 0.05 was considered significant.
Abbreviations
rplV: ribosomal protein L22; rplVindel: 27-nucleotide repeat sequence insertion in rplV gene; rRNA: ribosomal RNA; erm: erythromycin resistance methyltransferase; ABC: ATP-binding cassette; msrA: macrolide-streptogramins resistance A; mphC: macrolide phosphotransferase C; RT-PCR: reverse transcription-polymerase chain reaction; ERY: erythromycin; MIC: minimal inhibitory concentration; CLSI: Clinical and Laboratory Standards Institute.
Author contributions
G.Y. study concept and design, obtained funding; D.H. acquisition of data, analysis and interpretation of data; D.H. and Y.L. drafting of the manuscript; Y.L., J.L., C.L., Y.G., J.F. and H.L. administrative, technical, and material support; G.Y. and H.L. study supervision.
ACKNOWLEDGMENTS
We thank Dr. Hui Wang of Peking university people’s hospital for providing clinical macrolides-resistant S. aureus.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
FUNDING
This work was supported by grant from National Natural Science Foundation of China (http://www.nsfc.gov.cn) [31370170].
REFERENCES
1. Day NPJ, Moore CE, Enright MC, Berendt AR, Smith JM, Murphy MF, Peacock SJ, Spratt BG, Feil EJ. A link between virulence and ecological abundance in natural populations of Staphylococcus aureus (Retraction of vol 292, 114, 2001). Science. 2002; 295:971–971.
2. Archer GL. Staphylococcus aureus: a well-armed pathogen. Clin Infect Dis. 1998; 26:1179–1181.
3. Beceiro A, Tomas M, Bou G. Antimicrobial resistance and virulence: a successful or deleterious association in the bacterial world? Clin Microbiol Rev. 2013; 26:185–230.
4. Klevens RM, Morrison MA, Nadle J, Petit S, Gershman K, Ray S, Harrison LH, Lynfield R, Dumyati G, Townes JM, Craig AS, Zell ER, Fosheim GE, et al. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA. 2007; 298:1763–1771.
5. Boucher HW, Sakoulas G. Perspectives on Daptomycin resistance, with emphasis on resistance in Staphylococcus aureus. Clin Infect Dis. 2007; 45:601–608.
6. Sanchez Garcia M, De la Torre MA, Morales G, Pelaez B, Tolon MJ, Domingo S, Candel FJ, Andrade R, Arribi A, Garcia N, Martinez Sagasti F, Fereres J, Picazo J. Clinical outbreak of linezolid-resistant Staphylococcus aureus in an intensive care unit. JAMA. 2010; 303:2260–2264.
7. Brisson-Noel A, Trieu-Cuot P, Courvalin P. Mechanism of action of spiramycin and other macrolides. J Antimicrob Chemother. 1988; 22:13–23.
8. Walsh C. Molecular mechanisms that confer antibacterial drug resistance. Nature. 2000; 406:775–781.
9. Watanabe Y, Morimoto S, Adachi T, Kashimura M, Asaka T. Chemical modification of erythromycins. IX. Selective methylation at the C-6 hydroxyl group of erythromycin A oxime derivatives and preparation of clarithromycin. J Antibiot (Tokyo). 1993; 46:647–660.
10. Klein JO. History of macrolide use in pediatrics. Pediatr Infect Dis J. 1997; 16:427–431.
11. Peterson JH, Woolhead CA, Bernstein HD. The conformation of a nascent polypeptide inside the ribosome tunnel affects protein targeting and protein folding. Mol Microbiol. 2010; 78:203–217.
12. Menninger JR, Otto DP. Erythromycin, carbomycin, and spiramycin inhibit protein synthesis by stimulating the dissociation of peptidyl-tRNA from ribosomes. Antimicrob Agents Chemother. 1982; 21:811–818.
13. Gaynor M, Mankin AS. Macrolide antibiotics: binding site, mechanism of action, resistance. Curr Top Med Chem. 2003; 3:949–961.
14. Jenssen WD, Thakker-Varia S, Dubin DT, Weinstein MP. Prevalence of macrolides-lincosamides-streptogramin B resistance and erm gene classes among clinical strains of staphylococci and streptococci. Antimicrob Agents Chemother. 1987; 31:883–888.
15. Weisblum B. Erythromycin resistance by ribosome modification. Antimicrob Agents Chemother. 1995; 39:577–585.
16. Oldenburg M, Kruger A, Ferstl R, Kaufmann A, Nees G, Sigmund A, Bathke B, Lauterbach H, Suter M, Dreher S, Koedel U, Akira S, Kawai T, et al. TLR13 recognizes bacterial 23S rRNA devoid of erythromycin resistance-forming modification. Science. 2012; 337:1111–1115.
17. Reynolds E, Ross JI, Cove JH. Msr(A) and related macrolide/streptogramin resistance determinants: incomplete transporters? Int J Antimicrob Agents. 2003; 22:228–236.
18. Otto M, Gotz F. ABC transporters of staphylococci. Res Microbiol. 2001; 152:351–356.
19. Wright GD. Bacterial resistance to antibiotics: enzymatic degradation and modification. Adv Drug Deliv Rev. 2005; 57:1451–1470.
20. Matsuoka M, Inoue M, Endo Y, Nakajima Y. Characteristic expression of three genes, msr(A), mph(C) and erm(Y), that confer resistance to macrolide antibiotics on Staphylococcus aureus. FEMS Microbiol Lett. 2003; 220:287–293.
21. Chittum HS, Champney WS. Ribosomal protein gene sequence changes in erythromycin-resistant mutants of Escherichia coli. J Bacteriol. 1994; 176:6192–6198.
22. Gregory ST, Dahlberg AE. Erythromycin resistance mutations in ribosomal proteins L22 and L4 perturb the higher order structure of 23 S ribosomal RNA. J Mol Biol. 1999; 289:827–834.
23. Malbruny B, Canu A, Bozdogan B, Fantin B, Zarrouk V, Dutka-Malen S, Feger C, Leclercq R. Resistance to quinupristin-dalfopristin due to mutation of L22 ribosomal protein in Staphylococcus aureus. Antimicrob Agents Chemother. 2002; 46:2200–2207.
24. CLSI. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically, 9th ed. Approved standard. Clinical and Laboratory Standards Institute, Wayne, PA. 2012.
25. Wittmann HG, Stoffler G, Apirion D, Rosen L, Tanaka K, Tamaki M, Takata R, Dekio S, Otaka E. Biochemical and genetic studies on two different types of erythromycin resistant mutants of Escherichia coli with altered ribosomal proteins. Mol Gen Genet. 1973; 127:175–189.
26. Moore SD, Sauer RT. Revisiting the mechanism of macrolide-antibiotic resistance mediated by ribosomal protein L22. Proc Natl Acad Sci U S A. 2008; 105:18261–18266.
27. Hao H, Yuan Z, Shen Z, Han J, Sahin O, Liu P, Zhang Q. Mutational and transcriptomic changes involved in the development of macrolide resistance in Campylobacter jejuni. Antimicrob Agents Chemother. 2013; 57:1369–1378.
28. Descours G, Ginevra C, Jacotin N, Forey F, Chastang J, Kay E, Etienne J, Lina G, Doublet P, Jarraud S. Ribosomal Mutations Conferring Macrolide Resistance in Legionella pneumophila. Antimicrob Agents Chemother. 2017; 61:e02188–16.
29. Prunier AL, Trong HN, Tande D, Segond C, Leclercq R. Mutation of L4 ribosomal protein conferring unusual macrolide resistance in two independent clinical isolates of Staphylococcus aureus. Microb Drug Resist. 2005; 11:18–20.
30. Yan J, Han D, Liu C, Gao Y, Li D, Liu Y, Yang G. Staphylococcus aureus VraX specifically inhibits the classical pathway of complement by binding to C1q. Mol Immunol. 2017; 88:38–44.