
INTRODUCTION
Myeloid cells can differentiate into granulocytes, monocytes, and dendritic cells (DCs) under normal physiological conditions. However, in tumor microenvironments, the differentiation of myeloid lineage cells is aberrant because of the factors produced by tumor cells. Numerous clinical studies have shown that functionality of mature DCs decreased in patients with breast [1], non-small cell lung [2], pancreatic [3], cervical [4], hepatocellular [5], and prostate cancers as well as glioma [6] and oral squamous cell carcinoma (OSCC).
Myeloid-derived suppressor cells (MDSCs) are categorized into two main subpopulations with either a monocytic morphology (mo-MDSCs) or a polymorphonuclear morphology (PMN-MDSCs) [7, 8]. CD14+HLA-DRlow/- cells have been considered as mo-MDSCs in several kinds of tumors [9–11]. The number of MDSCs increases in the blood of patients with renal cell carcinoma [12], prostate cancer [11], none-small cell lung cancer [13] and other kinds of cancer [14, 15]. DCs and MDSCs are derived from common progenitor myeloid cells [16], and monocytes are the major precursor of DCs in humans [17]. It has been reported that CD14+ cells from cancer patients with an increased population of CD14+HLA-DRlow/- cells and have a deficiency to generate mature DCs [18].
Neoplastic and tumor-associated stromal cells release multiple soluble factors that perturb the myeloid compartment. Among these factors, vascular endothelial cell growth factor (VEGF) is one of the most important for angiogenesis. Our previous study demonstrated high expression of VEGF either in the serum or primary tumor site of OSCC patients, and its concentration was inversely correlated with the number of mature DCs [19], indicating an effect on the differentiation of DCs. Furthermore, some studies have shown an association between VEGF expression and signal transducer and activator of transcription 3 (STAT3) signaling, and inhibition of active STAT3 in tumor cells suppresses the expression of VEGF [20–22]. STAT3 is also constitutively activated in immune cells in the tumor microenvironment, which mediates cancer cell-initiated immune evasion signals in various immune cells [23, 24]. Inactivation of STAT3 signaling in hematopoietic cell-specific conditional knockout mice enhances antitumor immune responses through activation of various immune cells such as DCs [25]. However, it is unclear whether VEGF contributes to the recruitment and retention of monocytes in the primary tumor site or promotes differentiation of monocytes into immunosuppressive mo-MDSCs in OSCC. It is also unclear whether STAT3 participates in the dysfunction and dysdifferentiation of monocytes in OSCC.
In this study, frequencies of mo-MDSCs were higher in OSCC patients but decreased in postoperative patients. VEGF expression in the tumor nest (TN) was positively correlated with activation of STAT3 in the TN and tumor stroma (TS). Our data also indicated that VEGF affected the differentiation of monocytes and promoted the accumulation of mo-MDSC in OSCC via STAT3 activation. These findings suggest that inhibition of both VEGF expression and STAT3 activation may be an effective strategy for OSCC treatment.
RESULTS
Frequencies of mo-MDSCs were higher in OSCC patients but decreased in postoperative patients
To investigate the changes of monocytes in the peripheral blood of OSCC patients, we examined whole blood. The number of monocytes was increased slightly in OSCC patients (n = 330) compared with healthy controls (n = 60) but without a significant difference (P > 0.05; Figure 1A). However, upon removal of the OSCC tumor by resection, the numbers of monocytes were increased significantly in postoperative peripheral blood when compared with preoperative samples and controls (P < 0.001; Figure 1A). Moreover, we also investigated the number of leukocytes, the number of lymphocytes and neutrophils. There was a decrease in the number of leukocytes in OSCC patients, which increased significantly in postoperative peripheral blood compared healthy and preoperative peripheral blood samples (P < 0.01; Supplementary Figure 1A). The number of lymphocytes was more significantly decreased in OSCC patients when tumor was removed (P < 0.01; Supplementary Figure 1B). The number of neutrophils was increased significantly in both preoperative and postoperative peripheral blood of OSCC patients (P < 0.01; Supplementary Figure 1C). Furthermore, compared with preoperative peripheral blood, the number of neutrophils was elevated significantly in postoperative peripheral blood (P < 0.01; Supplementary Figure 1C). These results demonstrated that numbers of leukocytes, including monocytes, lymphocytes and neutrophils, were abnormal in OSCC patients.
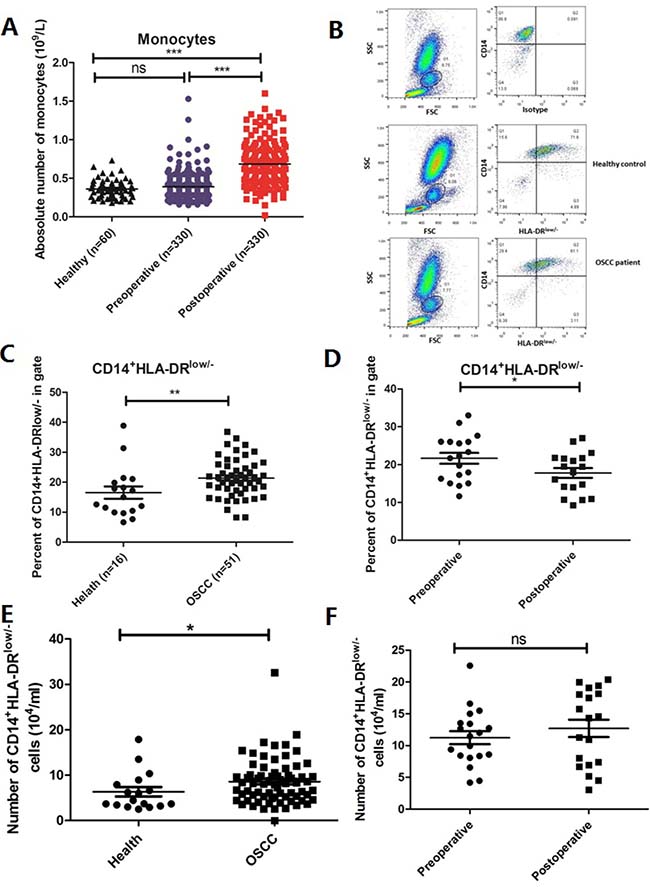
Figure 1: Frequencies of monocytes and CD14+HLA-DRlow/- mo-MDSCs are decreased in the postoperative peripheral blood of OSCC patients. (A) No significant difference was found in the frequency of monocytes between OSCC patients (n = 330) and healthy controls (n = 60), but it was significantly lower in postoperative peripheral blood than in preoperative and control samples (P < 0.001). (B) FCM was used to analyze mo-MDSCs (CD14+HLA-DRlow/-) in the peripheral blood of OSCC patients and healthy controls. (C) The frequency of CD14+HLA-DRlow/- mo-MDSCs was significantly higher in OSCC patients (n = 51) than in controls (n = 16) (P < 0.01). (D) The frequency of CD14+HLA-DRlow/- mo-MDSCs was significantly lower in postoperative OSCC patients than in preoperative OSCC patients (n = 18, P < 0.05). (E) The absolute number of mo-MDSCs was increased in OSCC patients (P < 0.05). (F) No significant difference was found in the absolute number of mo-MDSCs between preoperative and postoperative OSCC patients (P > 0.05). Data are shown as means ± SEM. Statistical significance was assessed by the Mann–Whitney U-test. *P < 0.05;**P < 0.01;***P < 0.001; ns: no significant difference.
MDSCs and DCs are commonly derived from myeloid cells, but these cell types have opposing functions. Therefore, we hypothesized that the frequency of mo-MDSCs was abnormal in OSCC patients and may change upon tumor removal because of decreases in the factors secreted by tumor cells. Therefore, the frequency and number of CD14+HLA-DRlow/-cells were assessed and the gating strategy are as follows: Firstly, monocytes were gated as G1 according to the FSC and SSC parameters, then cells in G1 were analyzed according to CD14 and HLA-DR expressions. Cells in upper left quadrant were analyzed for CD14+HLA-DRlow/− cells (Figure 1B). FCM analysis revealed that the frequency of CD14+HLA-DRlow/-mo-MDSCs was increased significantly in 51 OSCC patients compared with 16 healthy controls (Figure 1B and 1C, P < 0.001), which was decreased significantly in postoperative peripheral blood (n = 18) (Figure 1D, P < 0.05). We also calculated the absolute numbers of mo-MDSCs and found that it was increased in OSCC patients when compared with healthy controls (Figure 1E, P < 0.05). However, there was no significant difference in preoperative and postoperative patient samples (Figure 1F, P > 0.05). Importantly, the suppressive function of CD14+HLA-DRlow was determined by the co-culture with CD4+ T cells to analyze its proliferation. The data showed that CD14+HLA-DRlow/- cells could significantly inhibit the CD3/28-stimulated proliferation of T cells, indicating that CD14+HLA-DRlow/- cells are immunosuppressive cells (Supplementary Figure 2). These results suggested that certain factors secreted by OSCC tumor cells may contribute to the abnormal of myeloid cells.
Elevated VEGF expression and infiltrating CD14+ cells in tumor compartments including TS and TN
Our previous study demonstrated high concentration of VEGF in the serum of OSCC patients. However, we did not investigate the expression of VEGF in tumor compartments, including the tumor cells in TN and infiltrating leukocytes in TS including fibroblastic cells, and other cell types, as well as non-tumor (NT) compartments. Therefore, immunohistological analysis was carried out to detect VEGF expression in NT, TN and TS compartments. Three different areas in the same tumor sample were observed. Immunohistochemical staining patterns of VEGF in OSCC tumor tissues (n = 79) are shown in Figure 2A. The VEGF expression was lower in the NT compartment than that in the tumor compartments, and significantly higher in TN than that in TS compartments (P < 0.001, Figure 2B).These results suggested that OSCC cells secreted high amounts of VEGF. Correlations between VEGF expression in TN and TS and the tumor classification, lymph node metastasis, and tumor–node-metastasis (TNM) stage are shown in Supplementary Table 1. The VEGF-positive rate (32.2%) in TNM stages I and II was significantly lower than that in TNM stages III and IV (67.8%, P < 0.05).
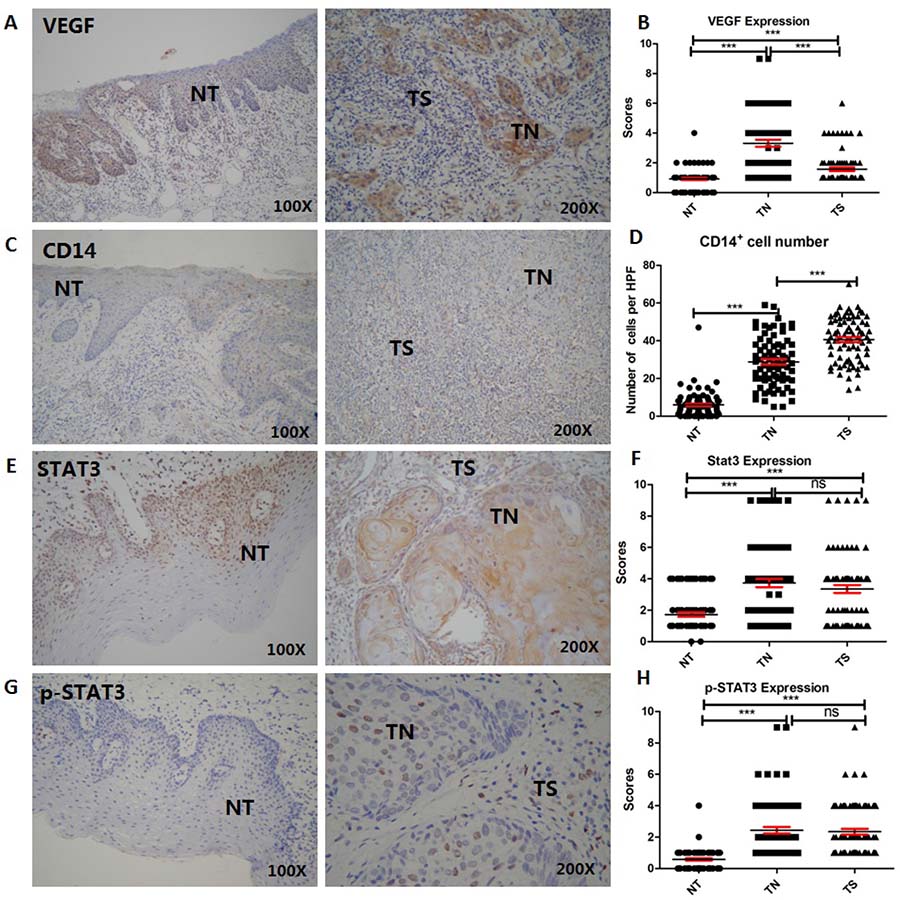
Figure 2: Expression of VEGF, STAT3, and p-STAT3 and the number of CD14+ mo-MDSCs are elevated in tumor compartments including TS and TN (n = 79). (A and B) VEGF expression was lower in the NT than tumor compartments and significantly higher in TN than TS compartments (P < 0.001). (C and D) The number of CD14+ cells was higher in tumor than non-tumor compartments (P < 0.001), indicating that the majority of CD14+ cells had infiltrated into tumor compartments, especially in the TS compartment. (E–H) Immunohistochemical staining patterns of STAT3 and p-STAT3 in NT and tumor compartments including TN and TS. STAT3 expression and activation were significantly higher in tumor than NT compartments, but there was no difference between TN and TS compartments. Data are shown as the means ± SEM. Statistical significance was assessed by the Mann–Whitney U-test. *P < 0.05;**P < 0.01;***P < 0.001; ns: no significant difference.
Because the number of mo-MDSCs was increased in the peripheral blood of OSCC patients, we also examined the infiltration of monocyte s in tumor and NT compartments. The vast majority of CD14+ cells had infiltrated into tumor compartments, especially TS compartments (Figure 2C), and the CD14+ cell number in tumor compartments was higher than that in NT compartments (Figure 2D, P < 0.0001). The infiltrating CD14+ cell number in TS was higher in patients with moderately to poorly differentiated tumors than that in those with well-differentiated tumors (Table 1, P < 0.05).
Table 1: Correlations between tumor-infiltrating CD14+ MDSCs in TN and TS and the clinicopathologic features in OSCC patients
Patients and tumor characteristics |
N |
CD14+ in TN |
P |
CD14+ in TS |
P |
|---|---|---|---|---|---|
(mean ± s) |
(mean ± s) |
||||
Sex |
|||||
Male |
38 |
28.63 ± 2.06 |
0.93 |
39.94 ± 1.865 |
0.66 |
Female |
41 |
28.89 ± 2.006 |
41.13 ± 1.7793 |
||
Age |
|||||
<60 |
46 |
31.47 ± 1.894 |
0.02 |
39.53 ± 1.608 |
0.28 |
≥60 |
33 |
24.4 ± 2.018 |
42.47 ± 2.192 |
||
Smoking |
|||||
No |
54 |
28.86 ± 2.724 |
0.97 |
38.95 ± 2.264 |
0.44 |
Yes |
25 |
28.76 ± 1.721 |
41.26 ± 1.574 |
||
Stage |
|||||
I–II |
45 |
29.56 ± 1.893 |
0.51 |
39.71 ± 1.668 |
0.37 |
III–IV |
34 |
27.58 ± 1.721 |
42.1 ± 2.083 |
||
Tumor differentiation |
|||||
Well |
32 |
30.82 ± 2.158 |
0.18 |
37.29 ± 2.062 |
0.04 |
Medium to poor |
47 |
26.9 ± 1.922 |
43.95 ± 1.569 |
||
Lymph node metastasis |
|||||
No |
52 |
29.93 ± 1.693 |
0.16 |
40.57 ± 1.489 |
0.92 |
Yes |
27 |
25.16 ± 2.668 |
40.89 ± 2.743 |
Abbreviations: TN, tumor nest; TS, tumor stroma.
VEGF expression in tumor compartments is correlated with activation of STAT3 in TS and TN compartments
Next, we investigated the correlation between VEGF secreted by OSCC tumor cells and STAT3 activation in tumor compartments (n = 79). Expression and activation of STAT3 were evaluated in OSCC tumor cells and infiltrating leukocytes. STAT3 expression and activation were significantly higher in tumor than NT compartments (Figure 2E and 2G),which was, however, no difference between TN and TS compartments (Figure 2F and 2H). Moreover, there were no significant correlation between its expression and in terms of gender, age, tumor differentiation, clinical stage, smoking, inflammation, and lymphnode metastasis (P > 0.05, Supplementary Table 2). A higher level of p-STAT3 in TN was correlated with tumor differentiation (P = 0.05), and that in TS was correlated with leukocyte infiltration (P = 0.025, Supplementary Table 3).
To examine the associations between VEGF,STAT3, and p-STAT3 in TN or TS compartments(n = 79), unvariable analysis were performed on the IHC results. There was no significant difference between VEGF and STAT3 expression in TN and TS (P > 0.05) (Supplementary Table 3). However, there were significant correlation between VEGF expression in TN and p-STAT3 expression in TN and TS (P = 0.03 and P = 0.02, respectively), suggesting that VEGF was positively associated with STAT3 activation in stroma (Table 2).
Table 2: Pearson’s correlation coefficients among VEGF, STAT3 and p-STAT3 expression in TN, TS and NT
P/R2 |
STAT3 TN |
STAT3 TS |
STAT3 NT |
p-STAT3 TN |
p-STAT3 TS |
p-STAT3 NT |
|---|---|---|---|---|---|---|
VEGF TN |
0.38/0.228 |
0.83/0.103 |
0.45/0.234 |
0.02/0.398 |
0.03/0.421 |
0.91/0.089 |
VEGF TS |
0.37/0.277 |
0.54/0.165 |
0.76/0.194 |
0.35/0.253 |
0.35/0.307 |
0.61/0.106 |
VEGF NT |
0.270.221 |
0.38/0.289 |
0.25/0.357 |
0.93/0.046 |
0.16/0.321 |
0.2/0.205 |
For abberavate: TN: Tumorr nest, TS: Tumor stroma, NT: normal epthelial tissues.
VEGF inhibits the differentiation of monocytes into DCs and increases the proportion of mo-MDSCs in OSCC by activating STAT3
Because of the high number of infiltrating CD14+mo-MDSCsand high expression of VEGF in blood and tumor compartments, we hypothesized that VEGF may participate in the differentiation of monocytes in OSCC. Therefore, we examined the influence and mechanism of VEGF in OSCC on the dysdifferentiation of monocytes into DCs or mo-MDSCs.
To clarify the effects of VEGF in OSCC on the differentiation of monocytes, surface markers of DCs and mo-MDSCs were detected at day 5. We found that the proportions of CD11a+,CD40+, CD80+, CD86+ and HLA-DR+ cells were decreased significantly among cells treated with TSN but the proportion of CD14+HLA-DR low/- mo-MDSCs was increased significantly, suggesting that TSN promoted monocyte differentiation into mo-MDSCs but not DCs (Figure 3A). However, inhibition of VEGF by Avastin significantly increased the proportions of CD11a+,CD40+, CD80+, CD86+ and HLA-DR+ cells, while that of CD14+HLA-DRlow/-mo-MDSCs was decreased significantly, suggesting that VEGF inhibition could rescue the dysdifferentiation of monocytes (Figure 3A). To further clarify the mechanism of VEGF in TSN on the differentiation of monocytes, AG490 was added in the TSN to block activation of the JAK pathway. Addition of AG490 to TSN resulted in significantly increased expression of DC surface markers, such as CD1a, CD40, CD80, CD86 and HLA-DR, but the proportions of CD14+HLA-DRlow/-mo-MDSCs were decreased significantly (Figure3A. Western blotting also demonstrated that TSN promoted the activation of JAK and STAT3,whereasAvastin and AG490 effectively inhibited activation of JAK and STAT3 (Figure 3B).
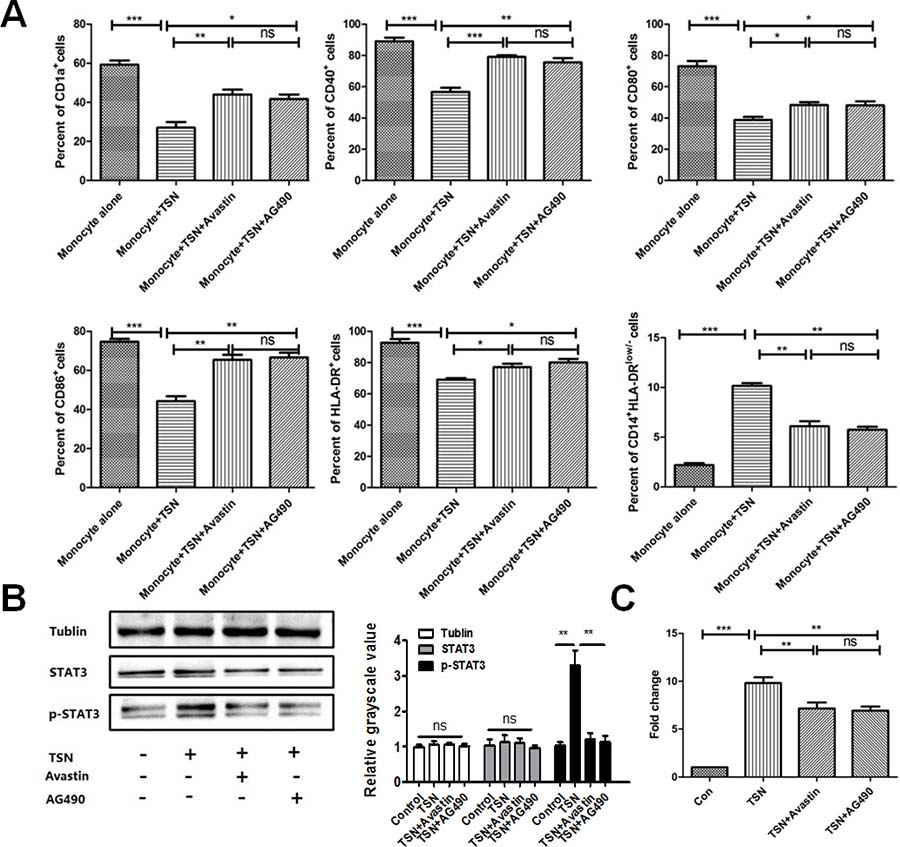
Figure 3: EGF in TSN inhibits the differentiation of monocytes into DCs and increases the proportion of mo-MDSCs in OSCC. (A) Expression of CD1a, CD40, CD80, CD86, and HLA-DR was decreased significantly and the proportion of CD14+HLA-DRlow/– mo-MDSCs was increased. (B) Western blotting indicated that TSN promoted the activation of JAK and STAT3, whereas Avastin and AG490 effectively inhibited activation of JAK and STAT3. Quantification of the western blot was performed. (C) The migratory ability of cells treated with TSN was significantly higher than that of the control. However, blocking VEGF with Avastin or STAT3 activation with AG490 significantly decreased the migratory ability of these cells. Data are shown as the means ± SEM. Statistical significance was assessed by the Mann–Whitney U-test. *P < 0.05; **P < 0.01; ns: no significant difference.
VEGF secreted by OSCC promotes the migration of monocytes in OSCC
Next, we determined whether VEGF promoted the migration of monocytes and differentiation of mo-MDSC in the OSCC microenvironment by performing chemotaxis assays. The level of VEGF in TSN was confirmed to be higher than that in normal epithelial cell HaCaT (Supplementary Figure 3).To confirm that VEGF was a major factor contributing to DC migration, cells were treated with TSN and Avastin. After 3 h, the number of cells treated with TSN in the lower chambers was significantly higher compared with the control (Figure 3C). However, inhibition of VEGF by Avastin significantly decreased the number of cells in the lower chambers, suggesting that VEGF promotes the migration of DCs in OSCC (Figure 3C). Of note, blocking STAT3 activation by AG490 significantly decreased the number of cells, indicating that VEGF promotes DC migration in OSCC by STAT3 activation.
DISCUSSION
Monocytes are the major precursor of DCs in humans, which are dysfunctional in cancer patients. VEGF is over-expressed in tumor cells, but the effects of VEGF on the dysdifferentiation of monocytes have been poorly known in OSCC patients. In this study, we found that the numbers of monocytes were decreased significantly in postoperative peripheral blood compared with preoperative blood. Notably, the frequency of CD14+HLA-DRlow/- MDSCs was increased significantly in OSCC patients, which was decreased postoperatively. In tumor sites, VEGF expression in TN was associated with activation of STAT3 in TN and TS compartments. In vitro, wealso showed that VEGF inhibited the differentiation of monocytes into functional DCs but promoted the differentiation to immunosuppressive mo-MDSCs by activation of STAT3. We also found that VEGF promoted the migration of monocytes by activating STAT3.
Under physiological conditions, monocytes could differentiate into immatureDCs [26]. Furthermore, tumor cells can affect distant sites, such as the bone marrow and spleen, by releasing soluble factors that drive accumulation of myeloid cells. Therefore, we inferred that monocytes must be ‘educated’ in tumors and its differentiation also must be ‘educated’. The number and distribution of monocytes in peripheral blood and tumor sites have not been characterized in OSCC patients. In this study, we found that the numbers of monocytes in OSCC patients was elevated slightly, but was decreased significantly in postoperative peripheral blood when compared with preoperative samples and controls. Recently, it has been shown that monocyte prevalence in peripheral blood inversely correlates with survival [27].
MDSCs are immature myeloid cells and contribute to negative regulation of immune responses in cancer [28, 29]. Tumor cells secrete numerous factors, such as VEGF, transforming growth factor-beta, and prostaglandin E2 (PGE2), which perturb the myeloid compartment and function. Our previous study showed that a decrease in the proportion of CD83+ mature DCs in OSCC [19]. In this study, we found that the frequency of CD14+HLA-DRlow/-mo-MDSCs was increased significantly in OSCC patients and was decreased significantly in postoperative peripheral blood, suggesting that certain factors secreted by OSCC cells may contribute to the dysdifferentiation of myeloid cells. Lechner et al. showed that GM-CSF, IL-6, PGE2, tumor necrosis factor-α,and VEGF induce human CD33+ monocytes with a potent suppressive capacity in vitro and suppressive CD33+ cells from PBMCs, which are consistent with human MDSCs [30]. Hoechst et al. demonstrated that mo-MDSCs induce CD4+CD25+Foxp3+ regulatory T cells (Tregs) in vitro. Thus, it is possible that the increased number of MDSCs in part contribute to the increased number of Tregs in OSCC patients.
Tumor-infiltrating DCs have an immature phenotype, while DCs in surrounding stromal tissue adopt a more mature phenotype [31], indicating that certain factors secreted by tumor cells influence the maturation of DCs. Therefore, it is important to identify the micro-localization of certain factors in OSCC tumors, including in TN and TS compartments. We found that VEGF expression was much stronger in TN than in TS, suggesting that VEGF may participate in the abnormal differentiation of myeloid cells in OSCC. There is an inverse correlation between VEGF expression and DCs in peripheral blood [19], and VEGF inhibits the maturation of DCs in vitro [32]. In this study, we found that VEGF expression in TN, but not in TS, was correlated with the TNM stage, suggesting that VEGF promoted the proliferation of OSCC cells and CD14+monocytes in TS, but not in TN, which was correlated with the TNM stage of OSCC. These results suggested that VEGF expression and the numbers of CD14+monocytes and CD14+HLA-DR-/low mo-MDSCs in OSCC were abnormal and VEGF may influence the migration and differentiation of monocytes.
In our previous study, we demonstrated that VEGF inhibits the maturation of DCs in OSCC in vitro [32]. However, the influence and mechanism of VEGF in the differentiation of DCs were unknown. Recently, the monoclonal antibody bevacizumab has been associated with favorable outcomes of patients with head and neck squamous cell carcinoma [33–35]. Blockade of STAT3 signaling in DCs inhibits functional maturation and decreases the number of immature CD11c+ MHC class IIlow CD86low DCs [36]. These findings were confirmed in a study that blocking STAT3 signaling in DCs neutralized the inhibitory effects of the tumor cells on DC maturation [37]. Stat3 activation in tumor cells has been associated with cytokine-induced proliferation, anti-apoptosis, and transformation and constitutively activated Stat3 has been found in many human tumors including HNSCC [38]. It exhibits its pro-transcription effects in response to signals from upstream receptors including the IL-6 cytokine receptor family, growth factor receptors. Aberrant regulation of STAT3 in HNSCC underlies malignant behaviors, contributing to growth, survival [39]. Aberrant STAT3 signaling in tumor is also immunosuppressive, protecting HNSCC cells from recognition and lysis by cytotoxic T lymphocytes [23]. In OSCC patients we found the expression of STAT3 was also increased in TN and TS as compared to NT indicating some factors derived from tumor cells and other cell types in the tumor microenvironment activate the transduction and activation of STAT3.
Another study showed that tumor-derived factors induce STAT3 signaling in immature myeloid cells, thereby preventing them from differentiating into mature DCs. We found that VEGF and TSN decreased the proportions of DCs but significantly increased the proportion of mo-MDSCs, suggesting that VEGF inhibited monocyte differentiation into mo-MDSC but not DCs. However, blocking VEGF by Avastin or STAT3 activation by AG490 significantly increased the proportion of DCs and decreased the proportion of mo-MDSCs. These data suggested that inhibition of both VEGF expression and STAT3 activation rescued the dysdifferentiation of monocytes. Functional DCs must migrate into the tumor site to exert their function. We uncovered that the high numbers of monocytes and mo-MDSCs found in these tumors were due to increased recruitment of progenitors by certain factors secreted by tumors.In addition, the migratory ability of monocytes was inhibited by Avastin or AG490, suggesting that VEGF promotes the migration of DCs in OSCC by activating STAT3. The accumulation of myeloid cells, an important immunosuppressive population in the tumor microenvironment, is partly due to STAT3 activation.
In conclusion, our study revealed a high prevalence of mo-MDSC in the peripheral blood and tumor sites of OSCC patients and VEGF in OSCC inhibited the differentiation of monocytes to promote the accumulation of MDSCs by activation of STAT3.
MATERIALS AND METHODS
Patients
Tumor samples from patients were obtained from Nanjing Stomatological Hospital, Nanjing University. A total of 330 OSCC patients and 60 controls sex-matched were included to analyze preoperative blood cell counts including lymphocytes, neutrophilic granulocytes, and monocytes. All of the patients diagnosed with primary OSCC were confirmed by hematoxylin and eosin staining by experienced pathologists from the Department of Pathology at Nanjing Stomatology Hospital. The ethical approval for this study was obtained from the Research Ethics Committee of Nanjing Stomatology Hospital.51 OSCC patients and 16 healthy controls were included to assess the frequency of CD14+HLA-DRlow/- mo-MDSCs. Furthermore, we analyzed the frequency of mo-MDSCs in preoperative and postoperative peripheral blood of 18 OSCC patients. Another 79 OSCC samples were collected for analysis of VEGF, CD14, STAT3 and p-STAT3 expressions by immunohistochemistry. All tumor patients had histologically confirmed OSCC and had not undergone chemotherapy, radiotherapy, or biotherapy. The patients did not have unrelated diseases such as diabetes, hematological disease, autoimmune diseases, or other malignant tumors at the time of diagnosis.
Flow cytometry (FCM)
To analyze the frequency of CD14+HLA-DRlow/- mo-MDSCs in the peripheral blood of OSCC patients, we used anti-human CD14-APC and HLA-DR-FITC monoclonal antibodies and FCM. For analysis of the effect of VEGF on monocytes, we used monoclonal antibodies against the following antigens: CD40-PE, CD80-FITC, CD86-FITC, HLA-DR-PE, and CD1a-PE. All antibodies were purchased from EBioscience.The Fc-R block (Human BD Fc Block™) was used. A total of 10,000 events were collected using a FACSCalibur (BD Biosciences, USA).Data were analyzed with Cell Quest software (BD Biosciences).
Cell culture
The human tumor cell line HSC-3 was cultured in Dulbecco’s modified Eagle’s medium with 10% fetal bovine serum (FBS) at 37° C in a humidified atmosphere with 5% CO2. The tumor cell culture supernatant (TSN) was harvested and stored at −20° C.
Immunohistochemical analysis
Formalin-fixed, paraffin-embedded sections of OSCC samples were deparaffinized in xylene and then rehydrated in graded ethanol concentrations. Endogenous peroxidase was blocked with 0.3% hydrogen peroxide/methanol for 10 min at room. Antigen retrieval was performed by immersing the sections in citrate-buffered solution (pH 6.0) and heating in a microwave for 20 min. The sections (n = 79)were allowed to cool for 40 min and then incubated with diluted anti-VEGF (1:200, Zhongshanjinqiao) -CD14 (1:200, Nowa), -STAT3 (1:200, Cell Signaling Technology) or -p-STAT3 (1:200; Cell Signaling Technology) antibodies overnight in a humidified box at 4° C. Nuclei were counterstained with hematoxylin. Two independent observers quantitatively evaluated VEGF, CD14, STAT3 and p-STAT3 expression by analyzing the cells of each specimen in high-power fields (40× objective). The observers examined each sample in duplicate in different areas of the same tumor in a blinded fashion. The scores of VEGF and STAT3 expression were calculated by the product of the percentage of positive cells (0, 0% positive cells; 1, <25% positive cells; 2, 26–50% positive cells; 3, 51–75% positive cells; 4, >75%) and the staining intensity (0, negative; 1, weak; 2, moderate; 3, high). To quantitatively evaluate CD14- and p-STAT3-positive cell infiltration, the number of positive cells was counted by NIS-Elements software (Nikon).
Isolation of human monocytes
Peripheral blood mononuclear cells (PBMCs) were isolated from human blood using Ficoll/Hypaque (density 1.077 g/ml, Shanghai Shisheng Cytobiotechnological Corp, China) according to the manufacturer’s instructions. After the last wash, PBMCs were resuspended in RPMI 1640 medium with 10% FBS and then seeded in a 24-well culture plate. After 3 h of culture, the monocytes identified by adherent cells were isolated by removing the non-adherent cells in the culture supernatant. To obtain immature DCs, monocytes were treated with 100 ng/ml recombinant human granulocyte-macrophage colony-stimulating factor (GM-CSF) and 40 ng/ml recombinant human interleukin (IL)-4 (Peprotech UK) for 5 days.
Cell co-culture
Cells were treated with TSN, TSN plus 5 μg/ml bevacizumab (recombinant human anti-VEGF antibody, Avastin), or TSN plus 2.7 μg/ml beyotime (STAT3 inhibitor, AG490) at 37° C in a humidified atmosphere with 5% CO2. Untreated monocytes considered as the control. Half of the culture medium was changed every 2 days. After 5 days of culture, non-adherent and loosely adherent cells were harvested for western blotting, FCM analysis, and chemotaxis assays.
Western blotting
Treated cells were collected, adjusted to a concentration of 1 × 106/ml, and then lysed in ice-cold lysis buffer. For immunoblotting, samples of 40–80 μg proteins were subjected to 12% SDS-polyacrylamide gel electrophoresis and then transferred to a polyvinylidene difluoride membrane (Roche, Germany) by electroblotting. The membrane was blocked for 1 h with 5% bovine serum albumin in 0.1% Tween 20/ TBS-T and then probed with primary antibodies at a dilution of 1:2000 overnight at 4° C. After three washes with TBS-T, the membrane was incubated with a horseradish peroxidase-conjugated secondary antibody (1:3000) for 2 h at room temperature. After washing, immunoreactive protein bands were detected by enhanced chemiluminescence. Images were acquired by the FluorChem FC2 System (Alpha InnotechStatistical Analysis, USA).
Chemotaxis assay
Chemotaxis assays were performed in 24-well Transwell plates with 8-μm pore polycarbonate filters. A total of 1 × 106 cells were transferred to the upper chambers, while the lower chambers contained PBS with 1% FBS. After 3 h of incubation at 37° C, the cells that migrated through the filters into the lower chamber were collected, fixed, and counted by FCM.
Statistical analysis
Statistical package for social sciences version 16.0 (SPSS Inc., Chicago, IL, USA) and the Prism statistical software package (version 5.0, Graphpad Software Inc.) were used for statistical analyses. Kolmogorov-Smirnov and Shapiro-Wilk tests were used to assess the data distribution. The Mann–Whitney U-test was used to compare two groups. Differences between more than two groupswere analyzed by the Kruskal–Wallis test. Statistical analysis was performed using Fisher’s exact test for 2 × 2 tables. Correlations of STAT3 and p-STAT3 expression in different compartments were assessed by univariable analysis. Differences were considered statistically significant at P < 0.05.
Author contributions
YH.N., L.D. QG.H. and YY.H. designed experiments with valuable help from XF.H. ZY.W.. YH.N. and L.D. performed and analyzed data; YH.N. wrote the manuscript. XF.H and S.C. collected surgical specimens with the help of YC.D.; YY.H. and YH.N. oversaw the overall project. ZX.H. contributed to the manuscript writing.
ACKNOWLEDGMENTS
We thank Hospital of Stomatology, Nanjing for helpful and essentially clinical samples.
CONFLICTS OF INTEREST
The authors declare no commercial or financial conflicts of interest.
FUNDING
This work was supported by a grant from the National Natural Science Foundation of China (No. 81402238, 81072213 and 81271698), the Nanjing Medical Science & Research Project (No. YKK13145), China Postdoctoral Science Foundation Funded Project (No.:2015M571740), Nanjing Medical Young engineer (QRX113311), Nanjing Municipal Key Medical Laboratory Constructional Project Funding (Since 2012), Center of Nanjing Clinical Medicine of tumor project (Since 2014).
REFERENCES
1. Pinzon-Charry A, Ho CS, Maxwell T, McGuckin MA, Schmidt C, Furnival C, Pyke CM, Lopez JA. Numerical and functional defects of blood dendritic cells in early- and late-stage breast cancer. Br J Cancer. 2007; 97:1251–1259.
2. Perrot I, Blanchard D, Freymond N, Isaac S, Guibert B, Pacheco Y, Lebecque S. Dendritic cells infiltrating human non-small cell lung cancer are blocked at immature stage. J Immunol. 2007; 178:2763–2769.
3. Bellone G, Carbone A, Smirne C, Scirelli T, Buffolino A, Novarino A, Stacchini A, Bertetto O, Palestro G, Sorio C, Scarpa A, Emanuelli G, Rodeck U. Cooperative induction of a tolerogenic dendritic cell phenotype by cytokines secreted by pancreatic carcinoma cells. J Immunol. 2006; 177:3448–3460.
4. Lee BN, Follen M, Rodriquez G, Shen DY, Malpica A, Shearer WT, Reuben JM. Deficiencies in myeloid antigen-presenting cells in women with cervical squamous intraepithelial lesions. Cancer. 2006; 107:999–1007.
5. Ormandy LA, Farber A, Cantz T, Petrykowska S, Wedemeyer H, Horning M, Lehner F, Manns MP, Korangy F, Greten TF. Direct ex vivo analysis of dendritic cells in patients with hepatocellular carcinoma. World J Gastroenterol. 2006; 12:3275–3282.
6. Pinzon-Charry A, Maxwell T, Lopez JA. Dendritic cell dysfunction in cancer: a mechanism for immunosuppression. Immunol Cell Biol. 2005; 83:451–461.
7. Gabrilovich DI, Ostrand-Rosenberg S, Bronte V. Coordinated regulation of myeloid cells by tumours. Nat Rev Immunol. 2012; 12:253–268.
8. Bronte V, Brandau S, Chen SH, Colombo MP, Frey AB, Greten TF, Mandruzzato S, Murray PJ, Ochoa A, Ostrand-Rosenberg S, Rodriguez PC, Sica A, Umansky V, et al. Recommendations for myeloid-derived suppressor cell nomenclature and characterization standards. Nat Commun. 2016; 7:12150.
9. Panni RZ, Sanford DE, Belt BA, Mitchem JB, Worley LA, Goetz BD, Mukherjee P, Wang-Gillam A, Link DC, Denardo DG, Goedegebuure SP, Linehan DC. Tumor-induced STAT3 activation in monocytic myeloid-derived suppressor cells enhances stemness and mesenchymal properties in human pancreatic cancer. Cancer Immunol Immunother. 2014; 63:513–528.
10. Poschke I, Mougiakakos D, Hansson J, Masucci GV, Kiessling R. Immature immunosuppressive CD14+HLA-DR-/low cells in melanoma patients are Stat3hi and overexpress CD80, CD83, and DC-sign. Cancer Res. 2010; 70:4335–4345.
11. Vuk-Pavlovic S, Bulur PA, Lin Y, Qin R, Szumlanski CL, Zhao X, Dietz AB. Immunosuppressive CD14+HLA-DRlow/- monocytes in prostate cancer. Prostate. 2010; 70:443–455.
12. Ochoa AC, Zea AH, Hernandez C, Rodriguez PC. Arginase, prostaglandins, and myeloid-derived suppressor cells in renal cell carcinoma. Clin Cancer Res. 2007; 13:721s–726s.
13. Huang A, Zhang B, Wang B, Zhang F, Fan KX, Guo YJ. Increased CD14HLA-DR myeloid-derived suppressor cells correlate with extrathoracic metastasis and poor response to chemotherapy in non-small cell lung cancer patients. Cancer Immunol Immunother. 2013; 62:1439–51.
14. Almand B, Clark JI, Nikitina E, van Beynen J, English NR, Knight SC, Carbone DP, Gabrilovich DI. Increased production of immature myeloid cells in cancer patients: a mechanism of immunosuppression in cancer. J Immunol. 2001; 166:678–689.
15. Diaz-Montero CM, Salem ML, Nishimura MI, Garrett-Mayer E, Cole DJ, Montero AJ. Increased circulating myeloid-derived suppressor cells correlate with clinical cancer stage, metastatic tumor burden, and doxorubicin-cyclophosphamide chemotherapy. Cancer Immunol Immunother. 2009; 58:49–59.
16. Liu K, Nussenzweig MC. Origin and development of dendritic cells. Immunol Rev. 2010; 234:45–54.
17. Idoyaga J, Steinman RM. SnapShot: Dendritic Cells. Cell. 2011; 146:660–660 e662.
18. Laborde RR, Lin Y, Gustafson MP, Bulur PA, Dietz AB. Cancer Vaccines in the World of Immune Suppressive Monocytes (CD14(+)HLA-DR(lo/neg) Cells): The Gateway to Improved Responses. Front Immunol. 2014; 5:147.
19. Wang ZY, Shi PH, Huang XF, Hua ZC, Hou YY, Han W, Hu QG. Peripheral blood dendritic cells and vascular endothelial growth factor in oral squamous cell carcinoma: correlation analysis and in vitro study. Int J Oral Maxillofac Surg. 2010; 39:713–720.
20. Carbajo-Pescador S, Ordonez R, Benet M, Jover R, Garcia-Palomo A, Mauriz JL, Gonzalez-Gallego J. Inhibition of VEGF expression through blockade of Hif1alpha and STAT3 signalling mediates the anti-angiogenic effect of melatonin in HepG2 liver cancer cells. Br J Cancer. 2013; 109:83–91.
21. Zhang X, Yue P, Page BD, Li T, Zhao W, Namanja AT, Paladino D, Zhao J, Chen Y, Gunning PT, Turkson J. Orally bioavailable small-molecule inhibitor of transcription factor Stat3 regresses human breast and lung cancer xenografts. Proc Natl Acad Sci U S A. 2012; 109:9623–9628.
22. Yokogami K, Yamashita S, Takeshima H. Hypoxia-induced decreases in SOCS3 increase STAT3 activation and upregulate VEGF gene expression. Brain Tumor Pathol. 2013; 30:135–143.
23. Wang T, Niu G, Kortylewski M, Burdelya L, Shain K, Zhang S, Bhattacharya R, Gabrilovich D, Heller R, Coppola D, Dalton W, Jove R, Pardoll D, Yu H. Regulation of the innate and adaptive immune responses by Stat-3 signaling in tumor cells. Nat Med. 2004; 10:48–54.
24. Yu H, Kortylewski M, Pardoll D. Crosstalk between cancer and immune cells: role of STAT3 in the tumour microenvironment. Nat Rev Immunol. 2007; 7:41–51.
25. Kortylewski M, Kujawski M, Wang T, Wei S, Zhang S, Pilon-Thomas S, Niu G, Kay H, Mule J, Kerr WG, Jove R, Pardoll D, Yu H. Inhibiting Stat3 signaling in the hematopoietic system elicits multicomponent antitumor immunity. Nat Med. 2005; 11:1314–1321.
26. Ho CS, Lopez JA, Vuckovic S, Pyke CM, Hockey RL, Hart DN. Surgical and physical stress increases circulating blood dendritic cell counts independently of monocyte counts. Blood. 2001; 98:140–145.
27. Sanford DE, Belt BA, Panni RZ, Mayer A, Deshpande AD, Carpenter D, Mitchem JB, Plambeck-Suess SM, Worley LA, Goetz BD, Wang-Gillam A, Eberlein TJ, Denardo DG, et al. Inflammatory monocyte mobilization decreases patient survival in pancreatic cancer: a role for targeting the CCL2/CCR2 axis. Clin Cancer Res. 2013; 19:3404–3415.
28. Gabrilovich DI, Nagaraj S. Myeloid-derived suppressor cells as regulators of the immune system. Nat Rev Immunol. 2009; 9:162–174.
29. Ding L, Ren J, Zhang D, Li Y, Huang X, Ji J, Hu Q, Wang H, Ni Y, Hou Y. The TLR3 Agonist Inhibit Drug efflux and Sequentially Consolidates Low-dose Cisplatin-based Chemoimmunotherapy while Reducing Side effects. Mol Cancer Ther. 2017; 16:1068–79.
30. Lechner MG, Liebertz DJ, Epstein AL. Characterization of cytokine-induced myeloid-derived suppressor cells from normal human peripheral blood mononuclear cells. J Immunol. 2010; 185:2273–2284.
31. Bell D, Chomarat P, Broyles D, Netto G, Harb GM, Lebecque S, Valladeau J, Davoust J, Palucka KA, Banchereau J. In breast carcinoma tissue, immature dendritic cells reside within the tumor, whereas mature dendritic cells are located in peritumoral areas. J Exp Med. 1999; 190:1417–1426.
32. Ni YH, Wang ZY, Huang XF, Shi PH, Han W, Hou YY, Hua ZC, Hu AQ. Effect of siRNA-mediated downregulation of VEGF in Tca8113 cells on the activity of monocyte-derived dendritic cells. Oncol Lett. 2012; 3:885–892.
33. Fury MG, Xiao H, Sherman EJ, Baxi S, Smith-Marrone S, Schupak K, Gewanter R, Gelblum D, Haque S, Schoder H, Shah JP, Katabi N, Kurtzman R, et al. Phase II trial of bevacizumab + cetuximab + cisplatin with concurrent intensity-modulated radiation therapy for patients with stage III/IVB head and neck squamous cell carcinoma. Head Neck. 2016; 38:E566–70.
34. Yao M, Galanopoulos N, Lavertu P, Fu P, Gibson M, Argiris A, Rezaee R, Zender C, Wasman J, Machtay M, Savvides P. Phase II study of bevacizumab in combination with docetaxel and radiation in locally advanced squamous cell carcinoma of the head and neck. Head Neck. 2015; 37:1665–71.
35. Bhatt VR, Ganti AK. Combination therapy with bevacizumab and temsirolimus in squamous cell carcinoma of the head and neck. Oral Oncol. 2013; 49:e25.
36. Shellenberger M. Nebraska’s first certified nurse midwife. Interview by Sue Bristol. Nebr Nurse. 1992; 25:12.
37. Nabarro S, Himoudi N, Papanastasiou A, Gilmour K, Gibson S, Sebire N, Thrasher A, Blundell MP, Hubank M, Canderan G, Anderson J. Coordinated oncogenic transformation and inhibition of host immune responses by the PAX3-FKHR fusion oncoprotein. J Exp Med. 2005; 202:1399–1410.
38. Geiger JL, Grandis JR, Bauman JE. The STAT3 pathway as a therapeutic target in head and neck cancer: Barriers and innovations. Oral Oncol. 2016; 56:84–92.
39. Sen M, Joyce S, Panahandeh M, Li C, Thomas SM, Maxwell J, Wang L, Gooding WE, Johnson DE, Grandis JR. Targeting Stat3 abrogates EGFR inhibitor resistance in cancer. Clin Cancer Res. 2012; 18:4986–4996.