
INTRODUCTION
Thyroid cancer is the most common malignant tumor of the endocrine system [1]. Most cancers derive from thyroid follicular cells and include well-differentiated forms, namely, papillary and follicular carcinomas and poorly differentiated forms such as anaplastic carcinoma [2]. About 15% of all thyroid cancers are follicular thyroid cancers (FTC) [3, 4]. Follicular carcinoma is considered more aggressive than papillary carcinoma. It occurs only rarely after radiation exposure, and generally in a slightly older age group than papillary thyroid cancer and is also less common in children.
The most frequent initial manifestation of thyroid cancer is the appearance of a nodule (more than 20% of the population has a palpable thyroid nodule and the percentage increases to 70% with ultrasound). In the great majority of cases (95%) it is simply an hyperplasia or a benign lesion, and only 5–30% of nodules are malignant and require surgical treatment. The most reliable diagnostic test for thyroid nodules is fine needle aspiration (FNA). Although this procedure clearly differentiates benign lesions from malignant lesions in about 60–70% of cases, cytology is “uncertain” in 20–30% of cases [5]. In these cases, a second FNA procedure or even surgery may be necessary, which results in additional morbidity and higher healthcare costs. Moreover, patients with malignant and indeterminate FNA cytology typically undergo limited surgery (i.e. lobectomy). If malignancy is diagnosed by pathological examination of the excised nodule, patients must undergo a second operation to complete the thyroidectomy, which, again, is associated with additional morbidity and costs. In addition, 1–3% of nodules diagnosed as benign on FNA are later found to be malignant on follow-up (false-negative FNA), and the delay in treatment before a definitive diagnosis results in high of risk of disease progression.
Molecular biology techniques are increasingly being used in routine cancer diagnostics [6–8]. Four molecular alterations, BRAF and RAS point mutations, and RET/PTC and PAX8/PPARγ rearrangements, are the most frequent causes of thyroid papillary and follicular carcinomas [9–11]. The most frequent genetic alterations in follicular carcinomas are point mutations of RAS and the PAX8/PPARγ rearrangement. These two alterations have been identified in approximately 80% of cases, and they seem to be mutually exclusive, which supports the hypothesis that they are distinct molecular subtypes. Moreover they are found in 18–52% of follicular carcinomas and in 24–53% of follicular adenomas. A much lower incidence has been reported in Hürthle cell tumors (15–25% of carcinomas and 0–4% of adenomas) [12].
The PAX8/PPARγ fusion oncogene [13] has been identified only in follicular carcinomas and in a small fraction (about 13%) of follicular adenomas [14]. Paired box 8 (PAX8), localized on chromosome 2q13, encodes the thyroid-specific paired domain transcription factor that is essential for the differentiation of follicular cells and the regulation of thyroid-specific genes. PAX8 is formed by 12 exons. Alternative splicing of exons 8–10 results in the production of multiple protein isoforms. The peroxisome proliferator–activated receptor gamma (PPARγ) gene, localized on chromosome 3p25 belongs to the nuclear receptor family of transcription factors that regulates cell differentiation and lipid metabolism. The two genes can fuse via translocation (2q:3p)(13:25) to form a new fusion gene that expresses a PAX8/PPARγ fusion protein, designated “PPFP”. Typically, the translocation fuses PAX8 intron 10 with the intron immediately preceding the first coding exon of PPARγ. Alternative splicing produces multiple RNA isoforms within the same neoplasia.
It is difficult, being a enough rare rearrangement, to have a PAX8/PPARγ fusion positive sample to use as positive control and for this reason we have made a sample suitable for this purpose.
Here we describe a rapid method to create a positive control and identify the rearrangement in one experiment in FNA thyroid samples by PCR.
RESULTS AND DISCUSSION
The PAX8 gene is formed by 12 exons but the first exon isn’t translated, alternative splicing of exon 8–10 results in different form that can be present simultaneously in the same tissue. The PAX8/PPARγ rearrangement is generally due to the translocation between PAX8 intron 10 and intron PPARγ immediately preceding exon 1. In Figure 1 we show a plain picture of the PAX8/PPARγ gene rearrangement and alternative splicing of PAX8. In Figure 1A are positioned, marked in red, the oligonucleotides we used, while in Figure 1B we presented the multiple rearrangement PAX8/PPARγ forms produced by alternative splicing all maintaining the open reading frame. Specifically, PAX8 exons 1–8 (exons 9 and 10 deleted), 1–9 (deleted exon 10 deleted) and 1–10 (exon 9 deleted) have been detected fused to the first coding exon of PPARγ, and all these situations maintain the PPARγ open reading frame [15].
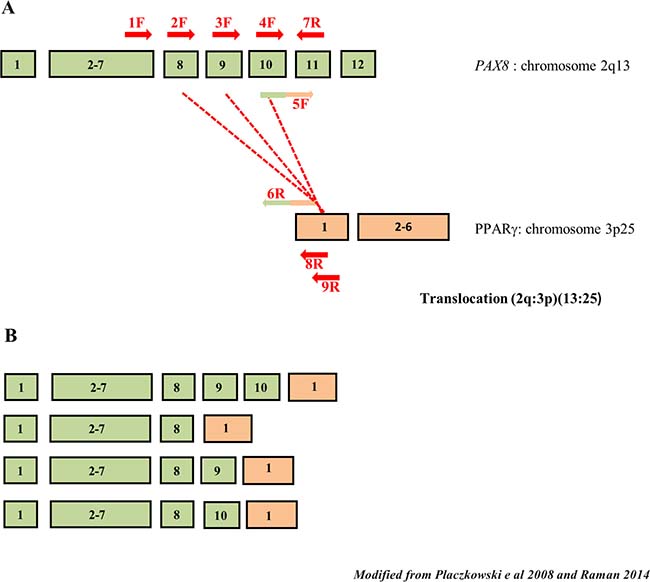
Figure 1: Schematic drawing of PAX8 splicing and PAX8/PPARγ rearrangements. (A) The PAX8 and PPARγ exons involved in the PAX8/PPARγ rearrangements. Exons 8, 9 and 10 of PAX8 are affected by alternative splicing. The primers used in our methodologies are reported in red (in different colors only the primers with the tail). (B) In rearrangement PAX8 alternative splicing can result in multiple forms in the same tumor. It is shown the more common rearrangements maintaining the reading frame.
Because the rarity of the PAX8/PPARγ rearrangement and the lack of cell lines carrying this mutation, we had to create a positive control to identify this chromosomic alteration. We approached this problem starting from wild-type thyroid RNA using the overlapping PCR technique, which is a simple, versatile technique for site-directed mutagenesis and gene splicing. We used two primers to join two DNA molecules: for each molecule, one primer is constructed such that it has a 5’ overhang complementary to the end of the other molecule. After the annealing, when the replication occurs, the DNA is extended to a new sequence that is complementary to the molecule to which it will be joined. Once both DNA molecules are extended, they are mixed and a PCR is carried out with only the primers for the far ends. The initial PCRs generate fragments with tails that are used as a template DNA to create a full-length product. The two stages of our overlapping PCR procedure are shown in Figure 2. As shown in Figure 2A we carried out two different amplifications: in step 1 the amplification of the PAX8 gene resulted in a 505 bp band, and in bands measuring 403 bp (no exon 10) and 316 bp (no exon 9) attributable to alternative splicing, whereas amplification of the PPARγ gene resulted in a 211 bp fragment (step 2). Then we pooled and amplified 10 μl of the products of steps 1 and 2 obtaining 5 fragments measuring 678 bp, 576 bp, 489 bp, 316 bp and 211bp, respectively (data no shown). In Figure 2B we show the longest purified amplification fragment (678 bp) that we used as positive control because it covers the whole rearrangement region. The fragment sizes from rearrangements are approximate because these processes can occur in different exon positions.
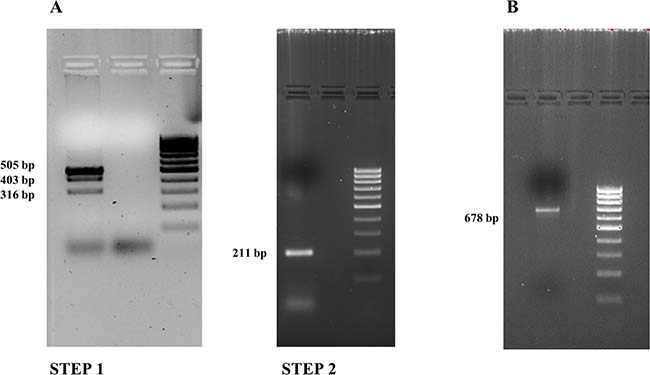
Figure 2: Overlapping PCR results (2% agarose gel). (A) Step 1: PAX8 amplification bands: 505 bp, 403 bp (no exon 10) and 316 bp (no exon 9) fragments. Step 2: PPARγ amplification band: 211 bp. (B) The PAX8/PPARγ fusion gene.
To set-up our multiplex method we have performed some steps: in the first step we have amplified Cwt in three different reactions using forward primers positioned respectively on exons 8, 9 and 10 of the PAX8 gene and reverse primer on PAX8 exon 11 and, as shown in Figure 3A, the amplification with the forward primer on PAX8 exon 8 resulted in multiple products (384 bp, 282 bp and 195 bp) originating, as mentioned above, from alternative splicing, while a 228 bp band appeared when we use a forward primer on PAX8 exon 9 and a 107 bp band appeared when we used an oligonucleotide as forward primer on PAX8 exon 10. Then we have amplified Cr using forward primers respectively on PAX8 exon 8, exon 9 and exon 10 and reverse primer on PPARγ exon 1. In Figure 3B are shown the amplification results: with the forward primer on PAX8 exon 8 we got multiple products (423 bp, 321 bp and 234 bp), with the forward primer on PAX8 exon 9 we obtained a 267 bp band and with the forward primer on PAX8 exon 10 a 146 bp band. Lastly we carried out a multiplex amplification in which we mixed three forward primers and two different reverse primers. The results obtained on positive controls are reported in Figure 3C.
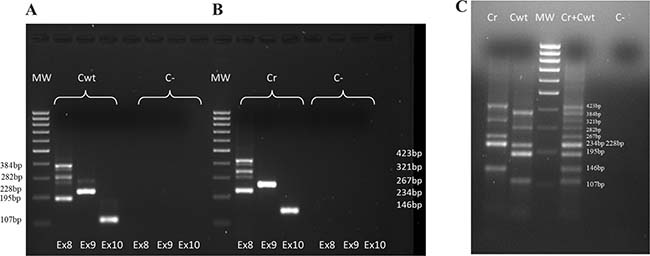
Figure 3: Positive controls amplifications. (A) Wild type PAX8 positive control (Cwt): PAX8 forms amplifications (forward primers respectively on PAX8 exon 8, exon 9 and exon 10 and reverse primer on PAX8 exon 11). (B) PAX8/PPARγ rearrangement positive control (Cr): PAX8/PPARγ rearrangement forms amplifications (forward primers respectively on PAX8 exon 8, exon 9 and exon 10 and reverse primer on PPARγ exon 1). (C) Cwt, Cr and (Cwt + Cr) amplifications in multiplex (simultaneously forward primers on PAX8 exon 8, exon 9 and exon 10 and reverse primers on PAX8 exon 11 and PPARγ exon 1)
In Figure 4 we show the last procedure with patients samples and we study with a single experiment both wild type PAX8 and for PAX8/PPARγ rearrangement. In fact it is possible to distinguish in the figure that the sample 121 is wild-type PAX8 positive and the sample 99 is affected by the PAX8/PPARγ rearrangement.
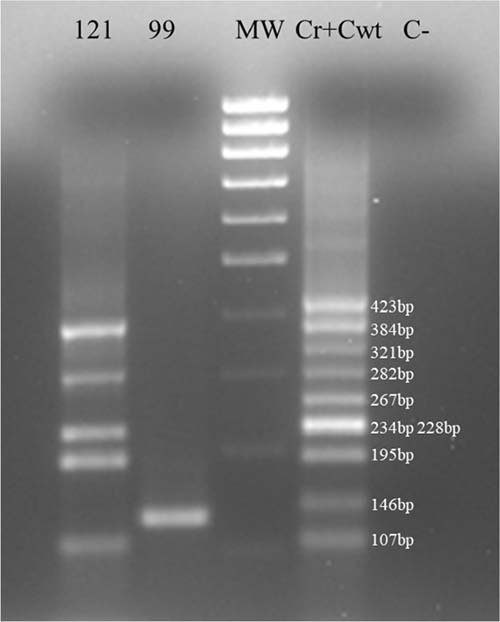
Figure 4: Multiplex samples amplification. Multiplex amplifications of sample 121, sample 99 and (Cwt + Cr). Sample 99 was negative for PAX8 wild-type amplification and positive for PAX8/PPARγ rearrangement.
This result was confirmed by Sanger sequencing (Figure 5, reverse sequencing). The fusion sample shows a translocation between PAX8 exon 8 and PPARγ exon 1 with a deletion of PAX8 exons 9 and 10 by alternative splicing and for this reason we did not obtain any result for sequences with the forward primer on exon 9 or on exon 10. Using this strategy we analyzed 60 RNA from FNA thyroid samples but only 4 were positive for the chromosomal rearrangement.
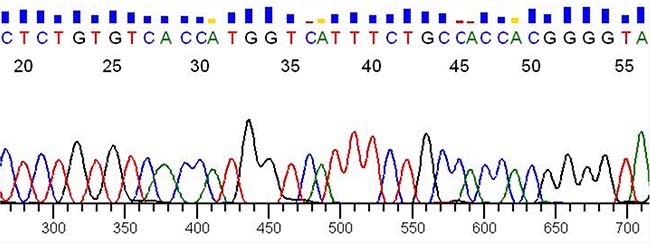
Figure 5: Sanger sequencing. Verification by Sanger sequencing of the reverse sequence of sample 99 shown in Figure 4.
The PAX8/PPARγ fusion protein has been implicated in thyroid follicular oncogenesis because it abrogates normal PPARγ function [16], increases cell cycle transition, reduces apoptosis and induces loss of both anchorage-dependence and contact inhibition [17].
The protein product of PAX8/PPARγ acts, at least in part, by inhibiting wild-type PPARγ signaling, which suggests that FTCs that present this rearrangement could be sensitive to PPARγ agonist therapy. Chromosomal alterations of PPARγ that result in the expression of the fusion protein PPFP may be an early event in the development or progression of FTC, and perhaps, the PAX8/PPARγ rearrangement could not be sufficient for the development of a full malignant phenotype. Indeed, additional genetic or epigenetic events may be required to enable the full phenotypic expression of FTCs [18].
Here we describe rapid inexpensive methods to create a positive control and to identify the wild type PAX8 and PAX8/PPARγ rearrangements in one experiment in FNA thyroid samples by PCR. Both methods are feasible even in laboratories that do not have sophisticated equipment or highly experienced staff. The described procedures could help to determine diagnosis. In addition, the identification of PAX8 gene alteration could help to develop selective and personalized therapy.
MATERIALS AND METHODS
Routine sample collection
We performed routine ultrasound-guided FNA using a 23 gauge needle. The aspirated sample was used for cytology and the residual material and the needle wash were directly collected into a tube containing 1 ml of RLT buffer (Qiagen, Germantown, MD, USA) and 10 μL 2-mercaptoethanol. The tube was stored frozen at –80° C until extraction of nucleic acid. Nucleic acid extraction was performed according to the manufacturer’s instruction (Qiagen RNeasy mini kit). Quantity and quality were assessed with a NanoDrop 1000 spectrophotometer (Thermo Scientific, Waltham, MA, USA), 1 μg of RNA was reverse transcribed into cDNA using the “QuantiTect Reverse Transcription kit” (Qiagen). The cDNA was stored at –20° C. GAPDH a housekeeping gene was used to verify that all the RNA samples were suitable for molecular analysis.
PAX8/PPARγ fusion positive control
To obtain a PAX8/PPARγ fusion positive control, we carried out a two-step overlapping PCR procedure (Figure 6A). In the first step we amplified the PAX8 and PPARγ genes. The primers for PAX8 amplification were located on exons 7 and 10 indicated respectively 1F and 6R in Figures 1 and 6. The primer on exon 10 was built with a tail corresponding to a region of exon 1 of PPARγ, whereas the primers for PPARγ amplification were located on PPARγ exon 1 but the forward primer was linked to a region of PAX8 exon 10 (5F and 9R Figures 1 and 6). Thus, the amplification products each bear nucleotide tails that correspond to the other studied gene. In Figure 6B we list overlapping PCR used primers. We performed PCR in a 25 μl final volume using as template 250 ng of cDNA product obtained by RT-PCR starting from 1 μg of total RNA. The amplification mixture consisted of 1× PCR buffer (Roche, Pleasanton, CA, USA), 200 μmol of dNTP and 1U of Taq DNA polymerase (Roche) and 0.2 pmol/μl of forward and reverse primers. The PCR thermal profile consisted of initial denaturation at 94° C for 10 min, followed by 40 cycles: denaturation at 95° C for 30 s, annealing at 48° C for 30 s and extension at 72° C for 1 min. Final extension was at 72° C for 10 min.
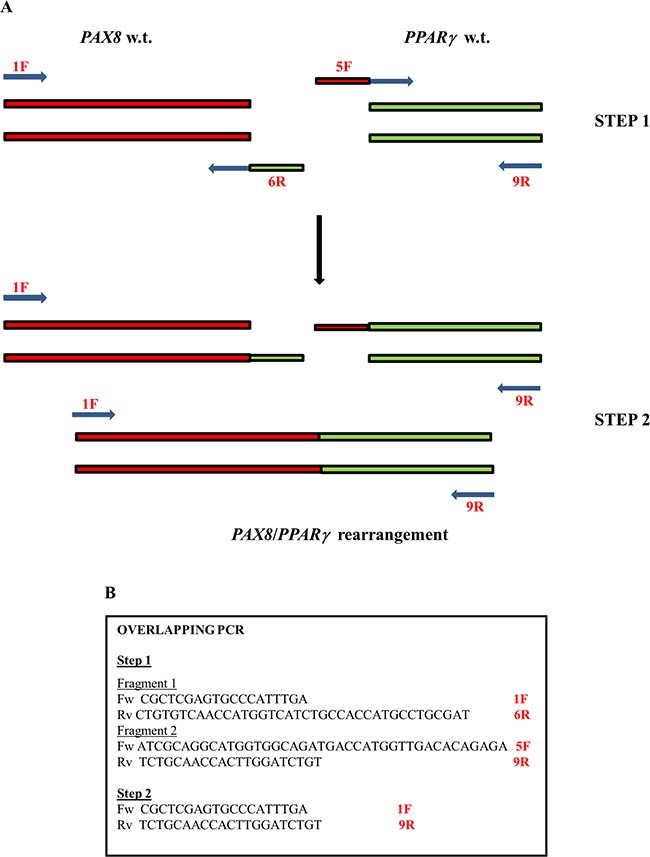
Figure 6: The plan of overlapping PCR. (A) The two-step overlapping PCR procedure used to obtain a PAX8/PPARγ fusion positive control. (B) The panel shows the primers used.
In the second step we used the same PAX8 forward primer and PPARγ reverse primer used in the first amplification. We also used the same mixture as in the first step but added, as template, 10 μl of product obtained from PAX8 amplification and 10 μl of product obtained from PPARγ amplification. The PCR thermal profile was designed to foster the formation of a hybrid PAX8 and PPARγ product. Initial denaturation, at 94° C for 10 min, was followed by 10 cycles consisting of denaturation at 95° C for 30 s, annealing at 48° C for 30 s and extension at 72° C for 1 min, and then by 30 cycles consisting of denaturation at 95° C for 30 s, annealing at 52° C for 30 s and extension at 72° C for 1 min. Final extension was at 72° C for 10 min. The amplification products were controlled on a 2% agarose gel. We next cloned the fusion gene using the Stratagene StrataClone PCR cloning kit (La Jolla, CA, USA), and extracted the plasmid DNA using the GenElute HP plasmid miniprep kit by Sigma-Aldrich (St Louis, MO, USA) according to the supplier’s instructions.
Wilde type PAX8 and PAX8/PPARγ fusion gene analysis
Samples, previously extracted and evaluated with the ubiquitous GAPDH gene, and controls, one positive wild type PAX8 sample (Cwt) and for the PAX8/PPARγ rearrangement, the fragment obtained by the overlapping PCR (Cr), are amplified for the wild-type PAX8 gene using the forward primers positioned respectively on exons 8, exon 9, exon 10 and reverse primer on exon 11 of the PAX8 gene, while for the PAX8/PPARγ amplification the forward primer is the same of previous reaction but the reverse primer is on exon 1 of PPARγ. The PCR conditions are the same described in overlapping PCR procedure while the PCR thermal profile consisted of initial denaturation at 94° C for 10 min, followed by 30 cycles: denaturation at 95° C for 30 s, annealing at 60° C for 30 s and extension at 72° C for 1 min. Final extension was at 72° C for 10 min. All PCR amplifications are performed with the C-1000 Thermal Cycler (Bio-Rad, CA, USA). In the same conditions we set up the simultaneous amplification of the three different PAX8 isoform both in Cwt than in Cr adding in mix forward primers on PAX8 exon 8, exon 9 and exon 10. Then we make a multiplex amplification of wild type PAX8 and PAX8/PPARγ rearrangement. The amplification conditions are the same as previously described but the annealing time is 1 min 30 s and reverse primers concentration, exon 11 of the PAX8 gene and on exon 1 of PPARγ, is modified to 0.3 pmol/μl. The amplifications are gel controlled (2% agarose gel in 1X TBE) and rearrangement-positive PCR products are purified (High Pure PCR Product Purification Kit Roche) and sequenced on both strands (forward and reverse) according to the manual of the Applied Biosystem (Waltham, MA, USA) “Big dye Terminator v3.1 Cycle Sequencing kit”. All sequencing amplification reactions of the individual PAX8/PPARγ rearrangement regions were performed using forward primers positioned respectively on exons 8, 9 and 10 of the PAX8 gene, while the reverse primer is the same for all the three amplifications positioned on exon 1 of PPARγ. The primers used, for both amplification and sequencing reactions, modified from Algeciras-Schimnich et al. [19], are listed in Figure 7. Subsequently, the sequence reactions were purified by Agencourt CleanSeq Beckman Coulter system (Brea, CA, USA) and analyzed by capillary electrophoresis on the Applied Biosystems “3730 DNA Analyzer”.

Figure 7: Primer’s list. Primers used for multiplex amplification of wild type PAX8 and PAX8/PPARγ rearrangement.
ACKNOWLEDGMENTS
We thank Jean Ann Gilder (Scientific Communication srl., Naples, Italy) for revising and editing the manuscript and Elvira Mauriello for technical support.
CONFLICTS OF INTEREST
None.
GRANT SUPPORT
This work was partially supported by grant from the Associazione Italiana per la Ricerca sul Cancro (AIRC) IG 12962 awarded to G.C.
REFERENCES
1. Gandhi M, Evdokimova V, Nikiforov YE. Mechanisms of chromosomal rearrangements in solid tumors: the model of papillary thyroid carcinoma. Mol Cell Endocrinol. 2010; 321:36–43.
2. Sherman SI. Thyroid carcinoma. Lancet. 2003; 361:501–511.
3. Proietti A, Sartori C, Borrelli N, Giannini R, Materazzi G, Leocata P, Elisei R, Vitti P, Miccoli P, Basolo F. Follicular-derived neoplasms: morphometric and genetic differences. J Endocrinol Invest. 2013; 36:1055–1061.
4. Cipriani NA, Nagar S, Kaplan SP, White MG, Antic T, Sadow PM, Aschebrook-Kilfoy B, Angelos P, Kaplan EL, Grogan RH. Follicular Thyroid Carcinoma: how have histologic diagnoses changed in the last half-century and what are the prognostic implications? Thyroid. 2015; 25:1209–16.
5. Dalquen P, Rashed B, Hinsch A, Issa R, Clauditz T, Luebke A, Lüttges J, Saeger W, Bohuslavizki KH. [Fine-needle aspiration (FNA) of the thyroid gland: analysis of discrepancies between cytological and histological diagnoses]. [Article in German]. Pathologe. 2016; 37:465–72.
6. Nikiforov YE, Yip L, Nikiforova MN. New strategies in diagnosing cancer in thyroid nodules: impact of molecular markers. Clin Cancer Res. 2013; 19:2283–2288.
7. Mehta V, Nikiforov YE, Ferris RL. Use of molecular biomarkers in FNA specimens to personalize treatment for thyroid surgery. Head Neck. 2013; 35:1499–1506.
8. Haugen BR, Alexander EK, Bible KC, Doherty GM, Mandel SJ, Nikiforov YE, Pacini F, Randolph GW, Sawka AM, Schlumberger M, Schuff KG, Sherman SI, Sosa JA, et al. 2015 American Thyroid Association Management Guidelines for Adult Patients with Thyroid Nodules and Differentiated Thyroid Cancer: The American Thyroid Association Guidelines Task Force on Thyroid Nodules and Differentiated Thyroid Cancer. Thyroid. 2016; 26:1–133.
9. Musholt TJ, Fottner C, Weber MM, Eichhorn W, Pohlenz J, Musholt PB, Springer E, Schad A. Detection of papillary thyroid carcinoma by analysis of BRAF and RET/PTC1 mutations in fine-needle aspiration biopsies of thyroid nodules. World J Surg. 2010; 34:2595–2603.
10. Ciampi R, Nikiforov YE. RET/PTC rearrangements and BRAF mutations in thyroid tumorigenesis. Endocrinology. 2007; 148:936–941.
11. Giordano TJ, Kuick R, Thomas DG, Misek DE, Vinco M, Sanders D, Zhu Z, Ciampi R, Roh M, Shedden K, Gauger P, Doherty G, Thompson NW, et al. Molecular classification of papillary thyroid carcinoma: distinct BRAF, RAS, and RET/PTC mutation-specific gene expression profiles discovered by DNA microarray analysis. Oncogene. 2005; 24:6646–6656.
12. Nikiforova MN, Lynch RA, Biddinger PW, Alexander EK, Dorn GW 2nd, Tallini G, Kroll TG, Nikiforov YE. RAS point mutations and PAX8-PPAR gamma rearrangement in thyroid tumors: evidence for distinct molecular pathways in thyroid follicular carcinoma. J Clin Endocrinol Metab. 2003; 88:2318–2326.
13. Kroll TG, Sarraf P, Pecciarini L, Chen CJ, Mueller E, Spiegelman BM, Fletcher JA. PAX8-PPARgamma1 fusion oncogene in human thyroid carcinoma [corrected]. Science. 2000; 289:1357–1360.
14. Marques AR, Espadinha C, Catarino AL, Moniz S, Pereira T, Sobrinho LG, Leite V. Expression of PAX8-PPAR gamma 1 rearrangements in both follicular thyroid carcinomas and adenomas. J Clin Endocrinol Metab. 2002; 87:3947–3952.
15. Raman P, Koenig RJ. Pax-8-PPAR-gamma fusion protein in thyroid carcinoma. Nat Rev Endocrinol. 2014; 10:616–623.
16. Placzkowski KA, Reddi HV, Grebe SK, Eberhardt NL, McIver B. The Role of the PAX8/PPARgamma Fusion Oncogene in Thyroid Cancer. PPAR Res. 2008; 2008:672829.
17. Gregory Powell J, Wang X, Allard BL, Sahin M, Wang XL, Hay ID, Hiddinga HJ, Deshpande SS, Kroll TG, Grebe SK, Eberhardt NL, McIver B. The PAX8/PPARgamma fusion oncoprotein transforms immortalized human thyrocytes through a mechanism probably involving wild-type PPARgamma inhibition. Oncogene. 2004; 23:3634–3641.
18. Sahpaz A, Onal B, Yesilyurt A, Han U, Delibasi T. BRAF(V600E) Mutation, RET/PTC1 and PAX8-PPAR Gamma Rearrangements in Follicular Epithelium Derived Thyroid Lesions - Institutional Experience and Literature Review. Balkan Med J. 2015; 32:156–166.
19. Algeciras-Schimnich A, Milosevic D, McIver B, Flynn H, Reddi HV, Eberhardt NL, Grebe SK. Evaluation of the PAX8/PPARG translocation in follicular thyroid cancer with a 4-color reverse-transcription PCR assay and automated high-resolution fragment analysis. Clin Chem. 2010; 56:391–398.