
INTRODUCTION
Hepatocellular carcinoma (HCC) is the 5th most frequent malignancy disease and the 3rd cause of cancer-associated death in the world [1]. It accounts for most of primary liver cancers. There are over half a million new cases diagnosed each year, with an approximately equal mortality each year due to its high fatality. The incidence of HCC is on the rise [2]. It is well known that cirrhosis and viral hepatitis are the most frequent risk factors for HCC. However, the mechanism of hepatocarcinogenesis is still unclear.
HBx has been reported to be involved in the development of liver cancer, such as hepatocellular carcinoma (HCC). HBx protein plays multiple roles in cell proliferation and apoptosis inhibition, cell signaling transduction, and gene expression regulation. In addition, HBx acts as a major regulator for viral replication [3]. However, there is argument about the direct causal effect of HBx on development of HCC [4]. Many studies reported the HBx gene integrated into the host genome in tissues with hepatocarcinoma and the gene mutants in HBx that develops as a result of this integration process [5, 6].
MicroRNAs (miRNAs), a family of small non-coding RNAs (~18–22 nt), regulate the expression of target genes on transcriptional level by accelarating mRNA degradation. Tumorigenesis and neoplastic transformation could be caused by specific miRNAs which impacting the translation of a range of crucial cellular genes and hence pivotal biological processes. Recently, Jiang et al. reported that miRNAs are commonly reduced in HCC and highly related with its clinical pathological characteristics, including prognosis, recurrence, metastasis and cirrhosis [7]. MiRNAs regulate proliferation, metastasis and apoptosis of HCC cells in hepatocarcinogenesis [8]. MiRNAs are also correlated with HCC cells that were originated from chronic HBV and HCV carriers, indicating that miRNAs could trigger the infection with HBV and HCV causing disease progression to HCC [9]. Wang et al systematically confirmed that full-length HBx inhibits of miRNAs and studied the downstream mechanisms involving hepatocarcino-genesis [10]. Exogenous HBx in HepG2 cells significantly inhibited the expression of 7 miRNAs and increased the expression of 11 miRNAs than the parental cells [10]. Ct-HBx-induced transcriptional inhibition may be one of the factors causing the decreased productions of miR-26a and miR-29c in HCC cells because of over-expression of HBx in HBV-associated HCC and frequent integration of HBx into the host genome in truncated form [11, 12].
Previous studies reported that the expression of miR-143 as well as some other miRNAs was substantially altered in HCC patients infected with HBV compared with those HCC patients without HBV, which might be attributed to the presence of HBX coded by the genome of HBV. In addition, BAG3, an oncogene which was predicted to be a target gene of miR-143, has been shown to be involved in the pathogenesis of HCC by suppressing the apoptosis of tumor cells [13–15]. Here, we investigated the relationship between the HBx with miR-143-3p or BAG3, and explored the molecular mechanism underlying the development of HBV-induced HCC.
RESULTS
Demographic, clinicopathological and genotypic parameters of the participants recruited in this study
48 patients were enrolled in this study, including 24 HCC patients positive for HBV and 24 cases negative for HBV. The demographic and clinicopathological characteristics of the participants, such as sex, age, differentiation and stage were described in Table 1. And the Student t-test was used to estimate the difference between groups. No significant difference was observed with respect to age, sex, differentiation and stage.
Table 1: Characteristics of hepatocellular carcinoma patients in the present study’
Characteristics |
Patient numbers n (%) |
P value |
|
|---|---|---|---|
HCC+HBV (n=24) |
HCC-HBV (n=24) |
||
Male gender (%) |
14 (58.3) |
15 (62.5) |
0.768 |
Age (yr, %) |
|||
<50 |
3 (12.5) |
5 (20.8) |
|
≥50 |
21 (87.5) |
19 (79.2) |
0.438 |
HCC differentiantion |
|||
Well |
2 (8.3) |
4 (16.7) |
|
Moderate |
15 (62.5) |
16 (66.7) |
|
Poor |
7 (19.2) |
4 (16.6) |
0.468 |
Pathological staging |
|||
Stage I |
1 (4.2) |
2 (8.3) |
|
Stage II |
15 (62.5) |
17 (70.8) |
|
Stage III |
8 (33.3) |
5 (20.9) |
0.562 |
Loss of miR-143-3p in human HBV (+) samples
Based on 24 HCC cases with HBV infection and 24 HCC patients without HBV infection, we evaluated the expression level of miR-143-3p using qRT-PCR. HCC patients with HBV have significant lower miR-143-3p level comparing with the control patients without HBV infections (Figure 1)
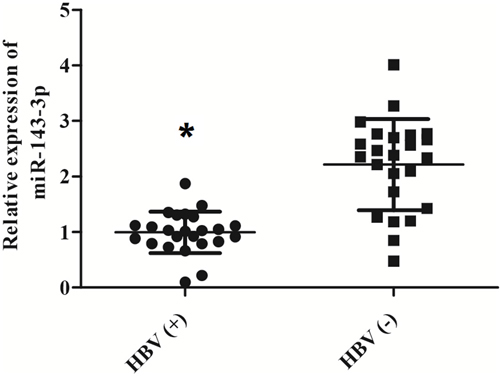
Figure 1: Loss of miR-143-3p is associated with HCC patients with HBV infaction. The level of miR-143-3p was measured among 48 participants consist of 24 HCC patients diagnosed with HBV and 24 HCC patients without HBV. (*, p<0.05).
BAG3 is a direct target gene of miR-143-3p
To investigate the target gene of miR-143-3p in the HBV-positive HCC samples, we searched miRBase and found that miR-143-3p directly recognized 375 candidate genes. We confirmed BAG3 is a target of miR-143-3p (Figure 2A). To further confirm that miR-143-3p directly inhibited expression of BAG3, we conducted luciferase reporter assay, Luc-BAG3-3’UTR-wt (wild-type) and its 3’UTR mut (mutant) plasmids were constructed, and transfected into HepG2 cells along with miR-143-3p mimic and anti-sense. The transfection of miR-143-3p significantly attenuated BAG3-3’UTR-wt reporter luciferase activity, but not the BAG3-3’UTR-mut. Meanwhile, the anti-sense of miR-143-3p significantly increase the expression of BAG3-3’UTR-wt, but not the-3’UTR-mut (Figure 2B). In words, BAG3 is a direct target gene of miR-143-3p.
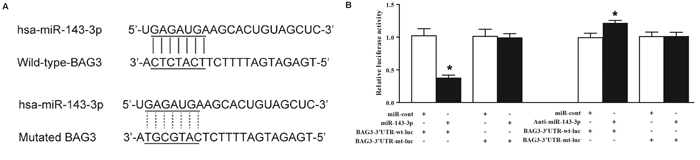
Figure 2: BAG3 is a direct target gene of miR-143-3p. (A) BAG3 was identified as the candidate target gene of miR-143-3p in cells with the “seed sequence” in the 3’UTR of BAG3. (B) In comparison with control, miR-143-3p mimics significantly inhibited the reporter luciferase activity of the BAG3 3-UTR-wt but not that of BAG3 3-UTR-mut. Meanwhile, miR-143-3p anti-sense increase the luciferase activity of BAG3-3UTR-wt, but not mutant reporter (*, p<0.05).
Gain of BAG3 in human HBV (+) samples
We evaluated the expression level of BAG3 in human samples, and confirmed that both mRNA level (Figure 3A) and protein level (Figure 3B) of BAG3 were significantly increased in HCC patients’ samples with HBV infection, which indicates that loss of miR-143-3p is highly correlated with the gain of BAG3.
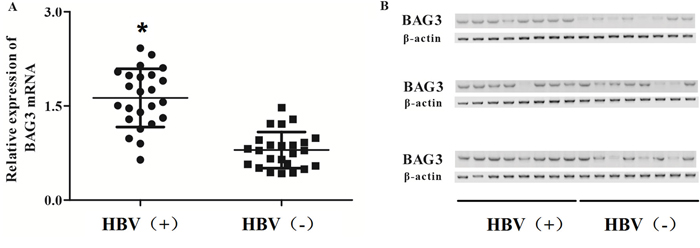
Figure 3: Gain of BAG3 is associated with HCC patients with HBV infraction. (A) The level of BAG3 mRNA was measured among 48 participants consist of 24 HCC patients diagnosed with HBV and 24 HCC patients without HBV, and BAG3 was highly expressed in HBV (+) group. (*, p<0.05). (B) The level of BAG3 protein was measured among 48 participants consist of 24 HCC patients diagnosed with HBV and 24 HCC patients without HBV, and BAG3 was highly expressed in HBV (+) group.
We further investigated the miRNA-mRNA regulatory relationship using Pearson correlation analysis. The negative correlation coefficient is -0.499 (r=-0.499) (Figure 4). Our clinical data support that miR-143-3p mediates HBx induced BAG3 upregulation.
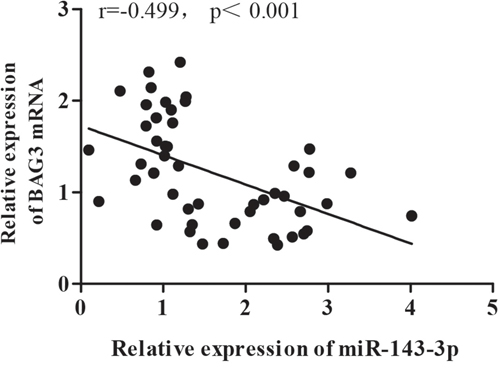
Figure 4: Gain of BAG3 is associated with loss of miR143-3p in human patients. The negative correlation coefficient between miR-143-3p and BAG3 was -0.499 (r=-0.499).
HBx regulates the expression level of MiR-143-3p and BAG3 in liver cells
To further confirm our finding based the clinical samples, we selected LO2, HepG2 and HepG2 2.2.15 cell lines for validation. LO2 is a normal liver cell line. HepG2 cell line is a liver cancer cell line without HBV infection. HepG2 2.2.15 is a liver cell line with HBV infection. We found that miR-143-3p was significantly inhibited the in HepG2 (P<0.05) and HepG2 2.2.15 (p<0.01) cells comparing with LO2 cells (Figure 5A). Meanwhile, the BAG3 was increased in in HepG2 (P<0.05) and HepG2 2.2.15 (p<0.01) cells comparing with LO2 cells (Figure 5B). The expression level of HBx was confirmed as what we expected (Figure 5C).
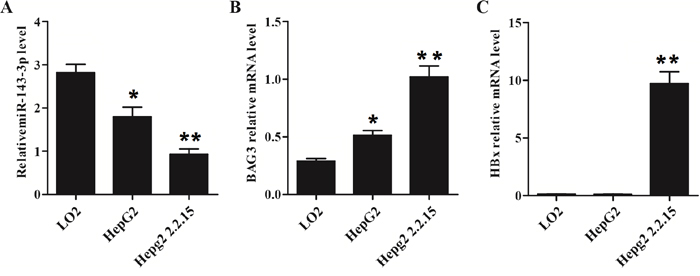
Figure 5: HBx is associated with lower miR-143-3p expression and higher BAG3 expression. MiR-143-3p, BAG3 and HBx expressions in two HCC cells line (HepG2 and HepG2 2.2.15) and normal liver cells (L02). (A) miR-143-3p was highly expressed in L02 cells than that in HepG2 and HepG2 2.2.15 cells. (*, p<0.05; **, p<0.01). (B) BAG3 mRNA was highly expressed in HepG2 2.2.15 cells than that in HepG2, and L02 cells. (*, p<0.05; **, p<0.01). (C) HBx was highly expressed in HepG2 2.2.15 cells comparing with HepG2 or L02 cells. (**, p<0.01).
To further investigate if the expression of BAG3 is regulated by miR-143-3p and HBx, we introduce miR-143-3p mimic or pHBx (HBx expression plasmid) into HepG2 cells. As shown in Figure 6, miR-143-3p expression level was significantly inhibited in cell introduction of miR-143-3p mimic or pHBx compared control group (Figure 6A). Meanwhile, the miR-143-3p expression promotion of pHBx was much stronger than miR-143-3p mimic. The protein level of BAG3 was evidently increased in pHBx group compared to that in the vector group (Figure 6B), suggested that HBX inhibited miR-143-3p expression and improved BAG3 expression.
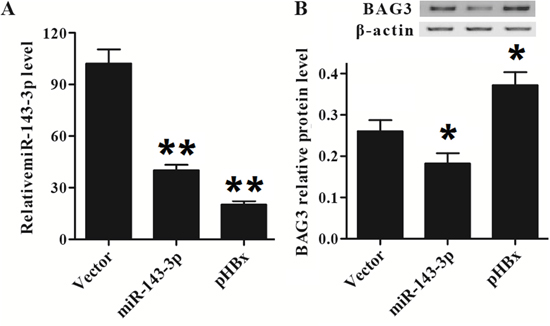
Figure 6: HBx regulates miR143-3p and BAG3 level in HepG2 cells. (A) The expression of miR-143-3p in cells introduction with pHBx was decreased. (B) The expression of BAG3 in cells introduction with pHBx was increased.
Next, we establish a stable HepG2 cells with HBx expression. The gain of HBx in HepG2 cells significantly inhibited miR-143-3p and increased BAG3 in HepG2 cells (Figure 7A and 7B). We further confirmed the BAG3 protein was significantly increased in the HepG2 cells (Figure 7C and 7D). This results further confirm that miR-143-3p mediates the HBx induced BAG3 upregulation.
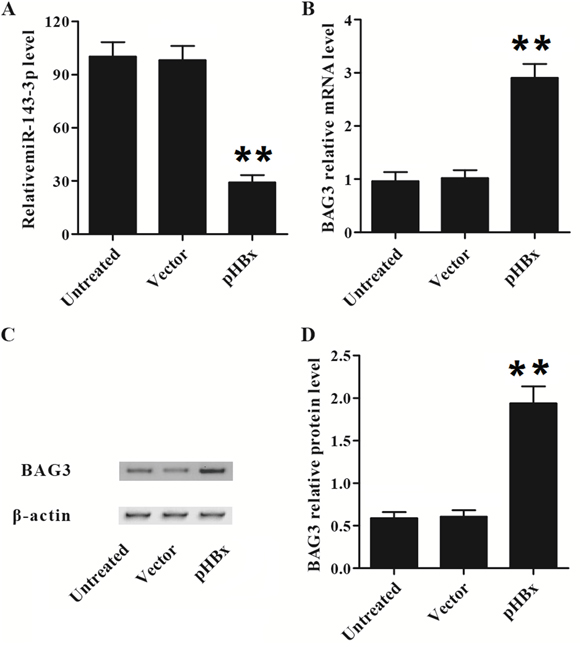
Figure 7: HBx inhibited the expression of miR143-3p and increased BAG3 expression in HepG2 cells. (A) The relative expression level of miR-146-3p was suppressed by pHBx. (**, p<0.01). (B) BAG3 mRNA level was over-expressed by pHBx. (**, p<0.01). (C and D) BAG3 protein level was over-expressed by pHBx. (**, p<0.01).
MiR-143-3p and HBx affected HepG2 cells proliferation and apoptosis
To investigate if the miR-143-3p and HBx regulates the proliferation and apoptosis, we establish stable HepG2 cells with miR-143-3p or HBx. The over-expression of miR-143-3p significantly inhibited proliferation and promoted apoptosis. On the contrary, HBx expression in Hepg2 cells significantly promoted the proliferation and inhibited apoptosis (Figure 8). The results suggested that BAG3 may mediates HBx induced proliferation and apoptosis inhibition
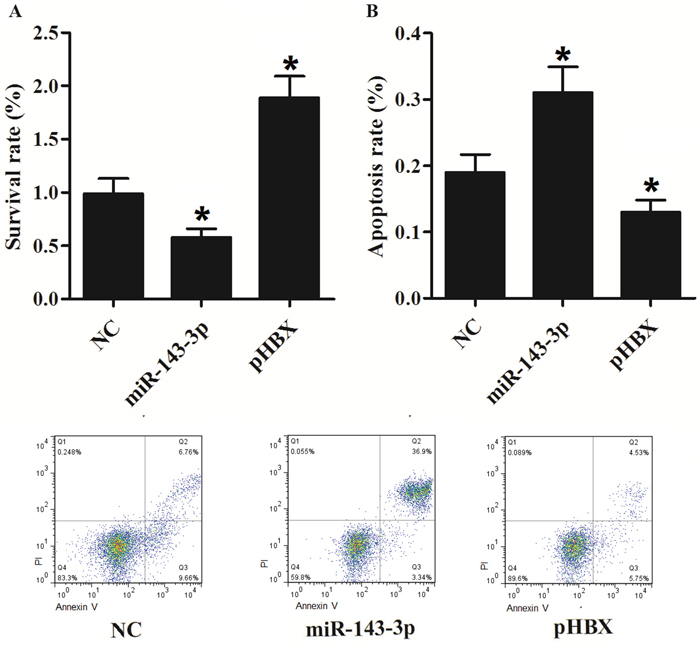
Figure 8: HBx promotes cell proliferation and inhibits apoptosis. (A) MiR-143-3p inhibited cell survival, while HBx improved cells growth. (*, p<0.05). (B) MiR-143-3p promoted cell proliferation, while HBx suppressed cells apoptosis. (*, p<0.05).
DISCUSSION
More and more studies support that HBx regulates miRNAs. Especially, HBx medicated extensive regulation of miRNAs in liver cells. The HBx mRNA acts in synergism with the HBx protein to inhibit miR-15a/16 productions via c-Myc [16]. Up-regulation of miR-29a by HBx protein was discovered, which in turn promotes cell migration by impacting PTEN in hepatoma cell lines [17]. The HBx protein inhibits miR-101 mediated DNA methylation by impacting DNA methyl transferase 3A [18]. It has been reported that HBx inhibited miR-145 mediated MAP3K upregulation, which promotes cell proliferation [19]. HBx expression also leads the loss of miR-21, the oncogenic miRNA, was observed in HepG2 cells. [19]. In this study, we recruited 48 patients: 24 HCC patients with HBV infection and 24 HCC patients without HBV infection to evaluate the relationship of HBx with miR143-3p and/or BAG3. Our study confirmed that miR-143-3p is inhibited in HBV positive patients.
Earlier studies have demonstrated substantial loss of miR-143 in gastric cancer, bladder, lung and colorectal cancers [20]. While gain of miR-143 is reported in esophageal cancer and pancreatic stellate cells cancer, there are still few studies investigated the correlation between the diagnosis of HCC and miR-143 expression [21]. To date, the correlation between HCC and miR-143 was rarely reported. Zhang et al. suggested that the expressions of miR-143 are significantly elevated in metastatic HBV-HCC of both HCC patients and p21-HBx transgenic mice. Gain of miR-143 enhances cancer cell migration/invasion and metastasis [22]. It also has been found that increased expressions of miR-143 and miR-215 in the serum samples derived from patients with HCC and chronic hepatitis when compared to the controls [23], both of which are different from our results of this study. The previous reports about the role of miR-143 in oncogenesis is controversial, while the majority of the references reported that miR-143 functions as a tumor suppressor in the tumorigenesis including bladder cancer, lung cancer, esophageal cancer, prostate cancer, colon cancer, ovarian cancer, breast cancer, osteosarcoma, and pancreatic cancer, which is in line with the data of our study [24–33]. In liver, the reports about miR-143 in oncogenesis is conflicting, and our results are consistent with the data from the following two studies [34, 35].
HBV infection is a primary cause to hepatocellular carcinoma (HCC) and human cirrhosis [36]. 7 types of viral proteins are encoded by 4 open reading frames of HBV. HBx plays a critical role among these proteins for replication of virus and pathogenesis of HCC [37, 38]. HBx is a 17-kDa multifunctional modulator that regulates gene expression via cytoplasmic signaling pathways and nuclear transcription factors [39]. HBx acts as an oncogene in the occurrence of hepatocellular carcinoma (HCC) [40]. It has reported that frequent integration of HBx into the host genome in truncated form which did not have its carboxyl-terminus and over-expression of HBx in HCC tissues related to HBV [11]. It is demonstrated that these variants of carboxyl-terminal truncated HBx (Ct-HBx) cancel the apoptotic and growth-inhibitory effects of full-length HBx [41]. Moreover, HBx is associated with epigenetic modifications during hepatocarcinogenesis [42]. The mechanism of HBx-induced hepatocarcinogenesis is still unclear.
BAG3 is identified as a binding partner of Bcl-2 [43]. The expression of BAG3 was robust in a lot of solid tumors, including colorectal carcinomas, non-small cell lung cancer and HCC [13, 14, 44]. The upregulation of BAG3 is triggered by a lot of stressful stimuli, including heavy metal exposure and high temperature, even though its expression is restricted to the striated muscle cells [45, 46]. BAG3 is associated with a board range of biological functions including cell invasion, cell survival and cell adhesion [47]. Loss of BAG3 promotes apoptosis of renal cancer cells. Meanwhile suppression of JNK signal pathway regulates BAG3 [48]. Our recent study in unilateral urinary obstruction (UUO) rat models have shown that BAG3 is implicated in fibroblast growth factor-2 (FGF2) mediated epithelial-mesenchymal transition (EMT) of HK2 cells, and elevated expression is found in tubular epithelium [49]. Although BAG3 is described as an oncogene, there are several studies with opposite findings. Recently, De-Hui Kong et al. determined that BAG3 prevented of anti-apoptotic effect [50]. Li et al. found that BAG3 could ensure stability of JunD mRNA to enhance growth suppression by serum starvation [51]. We investigated the role of BAG3 in HCC and found that BAG3 facilitated epithelial–mesenchymal transition, angiogenesis, invasion and tumor growth of HCC [13]. It has also been identified that the signaling pathway of HIF-1α might be part of the mechanisms of BAG3 in regulation of the angiogenesis and metastasis of HCC cells [52]. In this study, we found that the expression of BAG3 was apparently enhanced in HCC patients with HBV infection. We also found that miR-143-3p level in HepG2 was higher than HepG2 2.2.15 cells but lower than L02 cells. Meanwhile BAG3 in HepG2 was lower than HepG2 2.2.15 cells but higher than L02 cells. In addition, the gain of HBx inhibited the expression of miR-143-3p and promoted expression of BAG3 in HepG2 cells. Gain of HBx promotes the proliferation of HepG2 cells and inhibits apoptosis.
MATERIALS AND METHODS
Subjects
24 HCC patients with chronic HBV infection were enrolled from Department of Hepatobiliary Surgery, Affiliated Hospital of Guilin Medical University (Guilin, China) from April 2013 to November 2014. 24 HCC patients free of HBV infection were also enrolled during the same time. The participants with other liver diseases (such as drug-induced liver injury, Wilson’s disease, alcoholic hepatitis, steatohepatitis, autoimmune hepatitis, viral hepatitis A, hepatitis C, hepatitis E; metabolic), endocrine diseases (such as human immunodeficiency virus infection, hyperthyroidism, diabetes mellitus), and other comorbidities (such as renal dysfunction, respiratory system and cardiovascular) were excluded from our research. Participants or their first-degree relatives had already signed the informed consents before participating in the study. The Ethical Committee of Guilin Medical University (Guilin, china) approved the protocol of this study. The study was conducted according to the Declaration of Helsinki.
RNA isolation and quantitative real-time PCR
TRIzol reagent (Invitrogen, CA, USA) was used to extract the total RNA from tissue samples and cultured cells in accordance with the manufacturer’s protocol. Prime Script II 1st Strand cDNA Synthesis Kit (Takara Bio Inc., Japan) was used to reverse transcript the RNA samples based on the manufacturer’s recommendation. ABI PRISM 7900 Sequence Detection System (Applied Biosystems, Foster City, CA) with SYBR Green PCR mixture was used to perform the qRT-PCR (Quantitative real-time PCR) to quantify the expression of BAG3 and GAPDH, and GAPDH was used as the internal control to normalize the relative expression of BAG3 mRNA. ABIPRISM 7900 Sequence Detection System (Applied Biosystems, Foster City, CA) was used to perform the PCR in accordance with the manufacturer’s mannual. 2−ΔΔCt was used to analyze the relative quantification of the BAG3 mRNA and miR-143-3p. Three independent experiments were performed.
Cell culture and transfection
DMEM/F12 (Thermo Scientific, South Logan, Utah) containing 20 ng/mL EGF (Peprotech, New Jersey, USA), 100 ng/mL cholera toxin (Sigma, St. Louis, MO), 5% horse serum (Invitrogen, CA, USA), 10 μg/mL insulin (Sigma, St. Louis, MO), 0.5 μg/mL hydrocortisone (Sigma, St. Louis, MO), 100 μg/mL streptomycin and 100 units/mL penicillin was used to maintain the LO2, HepG2 and HepG2.1.1.15 cells at 37°C under a humidified atmosphere of 5% CO2/95% air. Lipofectamine™ plus Reagent (Invitrogen, CA, USA) was used to perform transient transfect, and then X-treme GENE siRNA transfection reagent (Roche, Basel, Switzerland) was used to transfect the HepG2 and HepG2.1.1.15 cells with miR-143 mimics or inhibitors and BAG3 siRNA in accordance with the manufacturer’s recommendation. Each test was repeated three times.
Proliferation assay
Cell proliferation reagent kit I (MTT) (Roche Applied Science, Indianapolis, IN, USA) and FACSCalibur flow cytometer (BD Biosciences, New Jersey, USA) were used to assess the proliferation of HepG2 cells in accordance with the manufacturer’s guidelines.
Luciferase assay
MiRBase (www.mirbase.org) and MircroRNA (www.microrna.org) were used to predict that the BAG3 is a target gene of miR-143-3p. High-Capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, CA, USA) was used to perform the reverse transcription (RT-PCR) to synthesize cDNA (BAG3) including binding sites for miR-143 based on the described from the manufacture. Agarose gel was used to separate and extract the PCR products following standard protocol. The PCR products were sub-cloned into the downstream of the luciferase gene in a pGL3-promoter plasmid using TA cloning Kit (Invitrogen, CA, USA), and the sequencing was used to confirm the efficiency. The mutagenesis was inserted into the same site of a control vector (Ambion, Cambridgeshire, UK) at the same time. Lipofectamine™ transfection Reagent (Invitrogen, CA, USA) was used to co-transfect the HepG2 cells with wild-type pGL3-BAG3 3’UTR or mutant pGL3-BAG3 3’UTR and negative controls (GenePharma, Shanghai, China) in accordance with the manufacturer’s instruction. Two days later, Dual-Luciferase Reporter Assay System (Promega, WI, USA) was used to detect the renilla luciferase activity/firefly luciferase activity following the supplier’s instruction. Each test was carried out three times.
Western blot analysis
To analyze the expression of BAG3 protein, Ice-cold RIPA lysis buffer including 0.01% phosphatase inhibitor cocktail (Sigma, St. Louis, MO) and phosphatase and protease inhibitors (Protease Inhibitor Cocktail Tablet, Roche, Indianapolis, IN) was used to prepare the whole cell extracts to lyse the cultured cells in accordance with manufacturer’s instruction. 12% SDS-PAGE was used to separate the whole cell proteins and the protein were transfer onto a nitrocellulose membrane (Millipore, Bedford, MA, USA). 5% defatted milk (Merck, Darmstadt, Germany) was used to block the membranes to avoid unspecific binding. GAPDH (glyceraldehyde-3-phosphate dehydrogenase) (Cell Signaling, Danvers, MA) was used to as the internal control. The primary antibodies (1:1000, Cell Signaling, Danvers, MA) and anti-β-actin (1:8000, Cell Signaling, Danvers, MA) was used to treat the membrane at 4°C overnight, and the secondary antibodies (1:15000, Cell Signaling, Danvers, MA) was used to incubate the membrane for 2 hours. And Super Signal West Femto chemoluminescent substrate (Thermo Scientific, IL, USA) was used to develop the blots, and digital acquisition system (ChemiDoc-It; UVP, Upland, CA) was used to visualize the bands, and ImageJ1.42 software (NIH, Bethesda, MD) was used to quantify the protein in accordance with the manufacturer’s recommendation. GAPDH was used as the internal control to normalize the expression of BAG3 protein. Each test was performed three times.
Apoptosis analysis
FACS Calibur flow cytometer (BD Biosciences, New Jersey, USA) was used to evaluate the apoptosis of HepG2 cells in accordance with the manufacturer’s protocol.
Statistical analysis
Data are shown as the means ± SD (standard deviation). One-way analysis was used to perform the comparisons, followed by and Mann-Whitney U test and Bonferroni post hoc test. SPSS version 16.0 (SPSS, Inc., Chicago, IL, USA) was used to perform the statistical analysis. The value of P less than 0.05 was considered significant.
CONCLUSION
Gain of HBx caused by HBV infection inhibits the expression of miR-143-3p, which leads to the upregulation of BAG3, a well-documented oncogene. HBx promotes the proliferation and inhibits the apoptosis of HepG2 cells. The HBV-HBX-miR-143-BAG3 signaling pathway is involved in the pathogenesis of HBV induced HCC.
Author contributions
Bo Tang designed bioinformatics algorithms, designed and performed experiments, analyzed data, and wrote the manuscript; Guangying Qi, Xingsi Liang, Fang Tang performed bioinformatics analysis; Shuiping Yu and Shengguang Yuan analyzed data; Xingyuan Jiao designed experiments and provided data; and Songqing He designed and performed experiments, analyzed data, and wrote the manuscript.
ACKNOWLEDGMENTS
This research was supported in part by The National Natural Science Foundation of China (No. 81360367, No. 81160066 and No. 30870719), Scientific Research Foundation for Returned Scholars, Ministry of Education of China (jyb2010-01), Key project of scientific research in colleges and universities in Guangxi (2013ZD046). Guangxi Natural Science Fund project (2014GXNSFBA118162), Health Department of Guangxi Chinese medicine science and technology special projects (GZPT13-45), Guangxi Distinguished Experts Special Fund, Project supported by the Guangxi culture of new century academic and technical leader of special funds.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
1. El-Serag, H.B., K.L. Rudolph, Hepatocellular carcinoma: epidemiology and molecular carcinogenesis. Gastroenterology. 2007; 132:2557-76.
2. Rahib, L., et al., Projecting cancer incidence and deaths to 2030: the unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014; 74:2913-21.
3. Murakami, S., Hepatitis B virus X protein: a multifunctional viral regulator. J Gastroenterol. 2001; 36:651-60.
4. Zhu, H., et al., Transgenic mice expressing hepatitis B virus X protein are more susceptible to carcinogen induced hepatocarcinogenesis. Exp Mol Pathol. 2004; 76:44-50.
5. Minemura, M., et al., Functional analysis of transactivation by mutants of hepatitis B virus X gene in human hepatocellular carcinoma. Oncol Rep. 2005; 14:495-9.
6. Yeh, C.T., et al., Identification and characterization of a prevalent hepatitis B virus X protein mutant in Taiwanese patients with hepatocellular carcinoma. Oncogene. 2000; 19:5213-20.
7. Jiang, J., et al., Association of MicroRNA expression in hepatocellular carcinomas with hepatitis infection, cirrhosis, and patient survival. Clin Cancer Res. 2008; 14:419-27.
8. Wong, Q.W., et al., MiR-222 overexpression confers cell migratory advantages in hepatocellular carcinoma through enhancing AKT signaling. Clin Cancer Res. 2010; 16:867-75.
9. Wong, Q.W., et al., MicroRNA-223 is commonly repressed in hepatocellular carcinoma and potentiates expression of Stathmin1. Gastroenterology. 2008; 135:257-69.
10. Wang, Y., et al., Lethal-7 is down-regulated by the hepatitis B virus x protein and targets signal transducer and activator of transcription 3. J Hepatol. 2010; 53:57-66.
11. Sirma, H., et al., Hepatitis B virus X mutants, present in hepatocellular carcinoma tissue abrogate both the antiproliferative and transactivation effects of HBx. Oncogene. 1999; 18:4848-59.
12. Yip, W.K., et al., Carboxyl-terminal truncated HBx regulates a distinct microRNA transcription program in hepatocellular carcinoma development. PLoS One. 2011; 6:e22888.
13. Xiao, H., et al., BAG3 regulates epithelial-mesenchymal transition and angiogenesis in human hepatocellular carcinoma. Lab Invest. 2014; 94:252-61.
14. Yang, X., et al., Bag-3 expression is involved in pathogenesis and progression of colorectal carcinomas. Histol Histopathol. 2013; 28:1147-56.
15. Peng, F., et al., HBx down-regulated Gld2 plays a critical role in HBV-related dysregulation of miR-122. PLoS One. 2014; 9:e92998.
16. Wu, G., et al., Hepatitis B virus X protein downregulates expression of the miR-16 family in malignant hepatocytes in vitro. Br J Cancer. 2011; 105:146-53.
17. Kong, G., et al., Upregulated microRNA-29a by hepatitis B virus X protein enhances hepatoma cell migration by targeting PTEN in cell culture model. PLoS One. 2011; 6:e19518.
18. Wei, X., et al., miR-101 is down-regulated by the hepatitis B virus x protein and induces aberrant DNA methylation by targeting DNA methyltransferase 3A. Cell Signal. 2013; 25:439-46.
19. Bandopadhyay, M., et al., Tumor suppressor micro RNA miR-145 and onco micro RNAs miR-21 and miR-222 expressions are differentially modulated by hepatitis B virus X protein in malignant hepatocytes. BMC Cancer. 2014; 14:721.
20. Ng, E.K., et al., MicroRNA-143 is downregulated in breast cancer and regulates DNA methyltransferases 3A in breast cancer cells. Tumour Biol. 2014; 35:2591-8.
21. Liu, S.G., et al., Differential expression of miRNAs in esophageal cancer tissue. Oncol Lett. 2013; 5:1639-1642.
22. Zhang, X., et al., Up-regulated microRNA-143 transcribed by nuclear factor kappa B enhances hepatocarcinoma metastasis by repressing fibronectin expression. Hepatology. 2009; 50:490-9.
23. Zhang, Z.Q., et al., Serum microRNA 143 and microRNA 215 as potential biomarkers for the diagnosis of chronic hepatitis and hepatocellular carcinoma. Diagn Pathol. 2014; 9:135.
24. Bai, J.W., H.Z. Xue, C. Zhang, Down-regulation of microRNA-143 is associated with colorectal cancer progression. Eur Rev Med Pharmacol Sci. 2016; 20:4682-4687.
25. Chen, J.H., et al., Functions of microRNA-143 in the apoptosis, invasion and migration of nasopharyngeal carcinoma. Exp Ther Med. 2016; 12:3749-3755.
26. Zhang, H.B., et al., miR-143 suppresses the proliferation of NSCLC cells by inhibiting the epidermal growth factor receptor. Exp Ther Med. 2016; 12:1795-1802.
27. Wang, L., et al., MiR-143 targets CTGF and exerts tumor-suppressing functions in epithelial ovarian cancer. Am J Transl Res. 2016; 8:2716-26.
28. Li, W.H., et al., MicroRNA-143 promotes apoptosis of osteosarcoma cells by caspase-3 activation via targeting Bcl-2. Biomed Pharmacother. 2016; 80:8-15.
29. Yang, Z., et al., MicroRNA143 targets CD44 to inhibit breast cancer progression and stem cell-like properties. Mol Med Rep. 2016; 13:5193-9.
30. Hirahata, M., et al., PAI-1, a target gene of miR-143, regulates invasion and metastasis by upregulating MMP-13 expression of human osteosarcoma. Cancer Med. 2016; 5:892-902.
31. Hu, Y., et al., miR-143 inhibits the metastasis of pancreatic cancer and an associated signaling pathway. Tumour Biol. 2012; 33:1863-70.
32. Song, T., et al., Expression of miR-143 reduces growth and migration of human bladder carcinoma cells by targeting cyclooxygenase-2. Asian Pac J Cancer Prev. 2011; 12:929-33.
33. Peng, X., et al., Identification of miRs-143 and -145 that is associated with bone metastasis of prostate cancer and involved in the regulation of EMT. PLoS One. 2011; 6:e20341.
34. Liu, X., J. Gong, B. Xu, miR-143 down-regulates TLR2 expression in hepatoma cells and inhibits hepatoma cell proliferation and invasion. Int J Clin Exp Pathol. 2015; 8:12738-47.
35. Wang, Y., et al., [miR-143 inhibits proliferation and invasion of hepatocellular carcinoma cells via down-regulation of TLR2 expression]. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2014; 30:1076-9.
36. Trepo, C., H.L. Chan, A. Lok, Hepatitis B virus infection. Lancet. 2014; 384:2053-63.
37. Lucifora, J., et al., Hepatitis B virus X protein is essential to initiate and maintain virus replication after infection. J Hepatol. 2011; 55:996-1003.
38. Neuveut, C., Y. Wei, M.A. Buendia, Mechanisms of HBV-related hepatocarcinogenesis. J Hepatol. 2010; 52:594-604.
39. Lupberger, J., E. Hildt, Hepatitis B virus-induced oncogenesis. World J Gastroenterol. 2007; 13:74-81.
40. Feitelson, M.A., L.X. Duan, Hepatitis B virus X antigen in the pathogenesis of chronic infections and the development of hepatocellular carcinoma. Am J Pathol. 1997; 150:1141-57.
41. Tu, H., et al., Biological impact of natural COOH-terminal deletions of hepatitis B virus X protein in hepatocellular carcinoma tissues. Cancer Res. 2001; 61:7803-10.
42. Jung, J.K., et al., Expression of DNA methyltransferase 1 is activated by hepatitis B virus X protein via a regulatory circuit involving the p16INK4a-cyclin D1-CDK 4/6-pRb-E2F1 pathway. Cancer Res. 2007; 67:5771-8.
43. Antoku, K., et al., Isolation of Bcl-2 binding proteins that exhibit homology with BAG-1 and suppressor of death domains protein. Biochem Biophys Res Commun. 2001; 286:1003-10.
44. Zhang, Y., et al., Bag3 promotes resistance to apoptosis through Bcl-2 family members in non-small cell lung cancer. Oncol Rep. 2012; 27:109-13.
45. Pagliuca, M.G., et al., Regulation by heavy metals and temperature of the human BAG-3 gene, a modulator of Hsp70 activity. FEBS Lett. 2003; 541:11-5.
46. Doong, H., et al., CAIR-1/BAG-3 forms an EGF-regulated ternary complex with phospholipase C-gamma and Hsp70/Hsc70. Oncogene. 2000; 19:4385-95.
47. Iwasaki, M., et al., BAG3 regulates motility and adhesion of epithelial cancer cells. Cancer Res. 2007; 67:10252-9.
48. Wang, H.Q., et al., Inhibition of the JNK signalling pathway enhances proteasome inhibitor-induced apoptosis of kidney cancer cells by suppression of BAG3 expression. Br J Pharmacol. 2009; 158:1405-12.
49. Du, F., et al., Implication of Bcl-2-associated athanogene 3 in fibroblast growth factor-2-mediated epithelial-mesenchymal transition in renal epithelial cells. Exp Biol Med (Maywood). 2015; 240:566-75.
50. Kong, D.H., et al., BAG3 sensitizes cancer cells exposed to DNA damaging agents via direct interaction with GRP78. Biochim Biophys Acta. 2013; 1833:3245-53.
51. Li, C., et al., BAG3 is upregulated by c-Jun and stabilizes JunD. Biochim Biophys Acta. 2013; 1833:3346-54.
52. Du, F., et al., BAG3 regulates ECM accumulation in renal proximal tubular cells induced by TGF-beta1. Am J Transl Res. 2015; 7:2805-14.