
Introduction
Carcinogenesis is a multistep process [1], in which abnormal glucose metabolism may play an important role [2, 3]. Abnormal energy metabolism of malignant tumors is a subject of current research. Glucose provides mammalian cells with their main source of energy, via glucose transporters (GLUTs) expressed on the cell membrane, which mediate glucose entry into cells. Energy is accessible to cells via two pathways: mitochondrial oxidative phosphorylation and glycolysis. The first key enzyme involved in glycolysis is a member of the hexokinase (HK) family. Thus, GLUTs and HKs are important enzymes mediating glucose metabolism in tumorigenesis [4]. Glycolysis provides only 10% of the energy in normal cells and relatively few of the total ATP molecules (2 ATP) generated per molecule of glucose broken down, compared with aerobic respiration, which produces 38 ATP molecules. Rapid proliferation of malignant tumors requires high levels of energy; therefore, tumors must accelerate glucose uptake and increase their rate of glycolysis [5]. The use of glycolysis by malignant cells to generate energy, whether in an aerobic or anaerobic environment, is known as the Warburg effect. However, the precise mechanism of this effect is not clear. It may involve the interaction of many factors, such as abundant lactic acid production during glycolysis, the escape of malignant tumors from immune surveillance, prevention of apoptosis, etc. Studies have found that members of the GLUT family, GLUT-1 in particular, and of the HK family, especially HK-II, are associated with the transport of glucose and mediation of glucose metabolism. Abnormal expression of GLUT-1 and HK-II in malignant tumors is associated with invasion and metastasis of tumors including head and neck cancers [6]. The tumor microenvironment is hypoxic, which is related to abnormal expression of GLUT-1 and HK-II. Many studies have shown that inhibition of GLUT-1 and HK-II expression aberrations can improve the treatment efficacy of malignant tumors [7–9]. Thus, inhibition of GLUT-1 and HK-II may represent a novel treatment strategy for head and neck cancers.
GLUT-1
Research history of GLUT-1
The study of transmembrane glucose transport dates back approximately 100 years. In 1977, GLUT-1 was first isolated from red blood cells [10]. In 2014, Yan et al. were the first to analyze the crystal structure of human GLUT-1 [11].
Biological characteristics of GLUT-1
Glucose is the primary source of cellular energy. The first step in glucose metabolism is glucose entry into the cell, a process that relies on glucose transport proteins. GLUTs, of which GLUT-1 was the first discovered and is the most widely distributed, are embedded in the cell membrane to transport extracellular glucose into the cell. Reports have found that GLUTs consist of 12 transmembrane protein domains. GLUTs exist in almost every cell in the body and are important not only for maintenance of normal human body functions but also tumor metabolism [12]. Tumor cells need to consume excess glucose to maintain growth and proliferation. GLUT-1 transports glucose into cells without requiring energy. GLUT-1 is expressed at low levels in normal tissues and benign lesions, whereas high expression is often correlated with carcinogenesis and can indicate a poor prognosis or recurrence [5].
GLUT-1 and malignant tumors
GLUT-1 and development of malignant tumors
GLUT-1 may play an important role in carcinogenesis. High GLUT-1 expression has been reported in colorectal carcinoma [13], lung carcinoma [14], breast carcinoma [15], esophageal carcinoma [16], gastric carcinoma [17], ovarian carcinoma [18], cholangiocarcinoma [19], and head and neck squamous cell carcinoma [20]. In malignant melanoma, Koch et al. found that GLUT-1 expression increased correspondingly with tumor progression from nevus (early stage) to advanced stages. They reported that inhibition of GLUT-1 expression can reduce the growth, apoptosis, and migration of malignant melanoma [21]. Lu et al. reported positive GLUT-1 expression in 73.6% of 53 pancreatic cancer samples, which was significantly higher than that in a control group [22]. According to one study, expression of GLUT-1 in rectal cancer was 93.6% [23]. GLUT-1 expression in hyperplasia, endometrial carcinoma, type I endometrial carcinoma, and type II endometrial carcinoma was 88.9%, 98.5%, 98% and 100%, respectively. GLUT-1 expression was also associated with cancer grade in endometrial carcinoma [24].
GLUT-1 and tumor staging
Lu et al. found that high expression of GLUT-1 was associated with the clinical stage of pancreatic cancer and with the standard uptake value (SUV) and Ki67 expression [22]. In endometrial carcinoma, the expression of GLUT-1 was related to SUV and was significantly higher in the FIGO stage IB and IC phase than the IA phase (P=0.001, P=0.003) [25].
A study examining the expression of hypoxia inducible factor-1α (HIF-1α), carbonic anhydrase-IX (CA-IX), GLUT-1, and vascular endothelial growth factor (VEGF) in 54 advanced cervical cancers reported that GLUT-1 expression was associated with tumor stage and lymphovascular involvement [26].
However, controversy remains regarding the relationship between GLUT-1 expression and the tumor stage. A study that measured the expression of GLUT-1, GLUT-3, GLUT-6, and GLUT-10 in 150 gastric carcinomas by immunohistochemical (IHC) methods found that GLUT-1 was not associated with tumor stage or prognosis, but that GLUT-3 was associated with tumor stage (per the Union for International Cancer Control (UICC) guidelines) and survival [27]. In endometrial cancer, Sadlecki et al. found that neither GLUT-1 nor CA-IX was associated with the FIGO clinical stage, histological grade, Bokhman subtypes, lymph node involvement, distant metastases, deep myometrial invasion, or recurrence [28].
GLUT-1 and metastasis of malignant tumors
Koch et al. found that GLUT-1 was expressed more strongly in melanoma metastases than in primary melanomas [21]. In pancreatic neuroendocrine tumors, GLUT-1 expression in lymph node metastases was significantly higher than that in non-metastatic tumors [29]. High expression levels of GLUT-1 in cervical cancer [26], ovarian cancer [30], gastric cancer [31], and non-small-cell lung cancer [32] have been shown to be related to lymph node metastasis [30].
GLUT-1 and prognosis of malignant tumors
Several studies have reported that GLUT-1 is a prognostic factor in malignant tumors [22, 33, 34]. In a study of colorectal cancer, Lee et al. found that the SUV and retention index (RI) (a high RI score predicts poor prognosis) were significantly higher in specimens with high GLUT-1 expression than in those with low GLUT-1 expression [34]. Younes et al. found that the prognosis of patients with bladder cancer was worse for those with greater than 10% compared with less than 10% tumor positivity for GLUT-1 expression (P=0.0064) [33]. In pancreatic cancer, the expression of GLUT-1 was associated with not only tumor size, stage, and lymph node involvement but also tumor proliferation and a worse prognosis. Univariate analysis showed that the survival of patients with high GLUT-1 expression was 12.3 months, which was significantly shorter than that of those with low expression (22.2 months), suggesting that high GLUT-1 expression in pancreatic cancer predicts poor prognosis. Multivariate analysis showed that GLUT-1 was the only factor measured that was associated with poor prognosis in pancreatic cancer [22]. Several other studies yielded similar results for pancreatic cancer [35–37]. A meta-analysis of a selection of biomarkers including GLUT-1 found an association of GLUT-1 with poor prognosis in biliary tract cancers [38]. GLUT-1 expression has also been associated with the prognosis of many tumor types, including gastric cancer [39], osteosarcoma [40], meningioma [41], esophageal cancer [42], cervical cancer [43], endometrial cancer [28], and ovarian cancer [30].
The relationship between GLUT-1 and other factors
HIF-1α and GLUT-1 are important markers of hypoxia in the cancer microenvironment. GLUT-1 expression is influenced by HIF-1α [44], as well as by mitochondrial oxidative phosphorylation [45]. GLUT-1 and CA-IX have been coined “endogenous hypoxia markers” in some scholarly reports [26]. Studies have shown that the expression of GLUT-1 is affected by many factors, such as glucose deprivation, cellular carcinogenesis, and osmotic pressure [46]. While some studies have found a relationship between GLUT-1 and hypoxia [26, 44–46], others have not found a direct correlation [47, 48]. This divergence may be related to difficulties in identifying hypoxic regions of tissue when examining gangrenous versus living tissue by Polarography observation; the hypoxic regions identified may be inaccurately high [49]. One study showed that GLUT-1 expression is related to Ki-67 expression, a marker of cell proliferation, which is closely related to tumor differentiation, invasion, metastasis, and prognosis [22].
Possible mechanisms of resistance to cancer radiotherapy and chemotherapy mediated by GLUT-1
The underlying mechanism of GLUT-1 expression, which induces chemo-radioresistance in cancer cells, remains unclear. It is likely caused by interplay among multiple factors. First, the higher energy metabolic rate of malignant tumor cells compared with their nonmalignant counterparts, even in aerobic glycolysis, is called the Warburg effect. Increased GLUT-1 expression plays an important role in cancer glucose metabolism. We suggest that GLUT-1 overexpression results in satisfying the higher energy requirement of cancer and leads to chemo-radioresistance. Second, hypoxia is a common phenomenon in solid tumors. GLUT-1 is a potential intrinsic marker of hypoxia in cancer, and HIF-1α regulates the expression of several hypoxia-response genes, including GLUT-1. As mentioned above, HIF-1α expression is associated with GLUT-1 expression in many cancers, and both markers are associated with the poor prognosis of many cancers. We suggest that GLUT-1 expression is associated with chemo-radioresistance via HIF-1α regulation. Third, GLUT-1 expression is also regulated by the PI3K/Akt pathway. This pathway plays a role in chemo-radioresistance/intrinsic resistance, tumor cell proliferation and hypoxia. The results of our previous and others’ studies suggest that activation of the PI3K/Akt pathway plays a role in GLUT-1-mediated chemo-radioresistance in solid cancers, including laryngeal carcinoma. Fourth, CD133+ cancer stem cells may be involved in chemo-radioresistance caused by GLUT-1. In our previous study, we found that chemo-radioresistance in CD133+ laryngeal carcinoma was higher than that in CD133- laryngeal carcinoma cells [50–53] (Figure 1). However, the mechanism of cancer chemo-radioresistance needs further study.
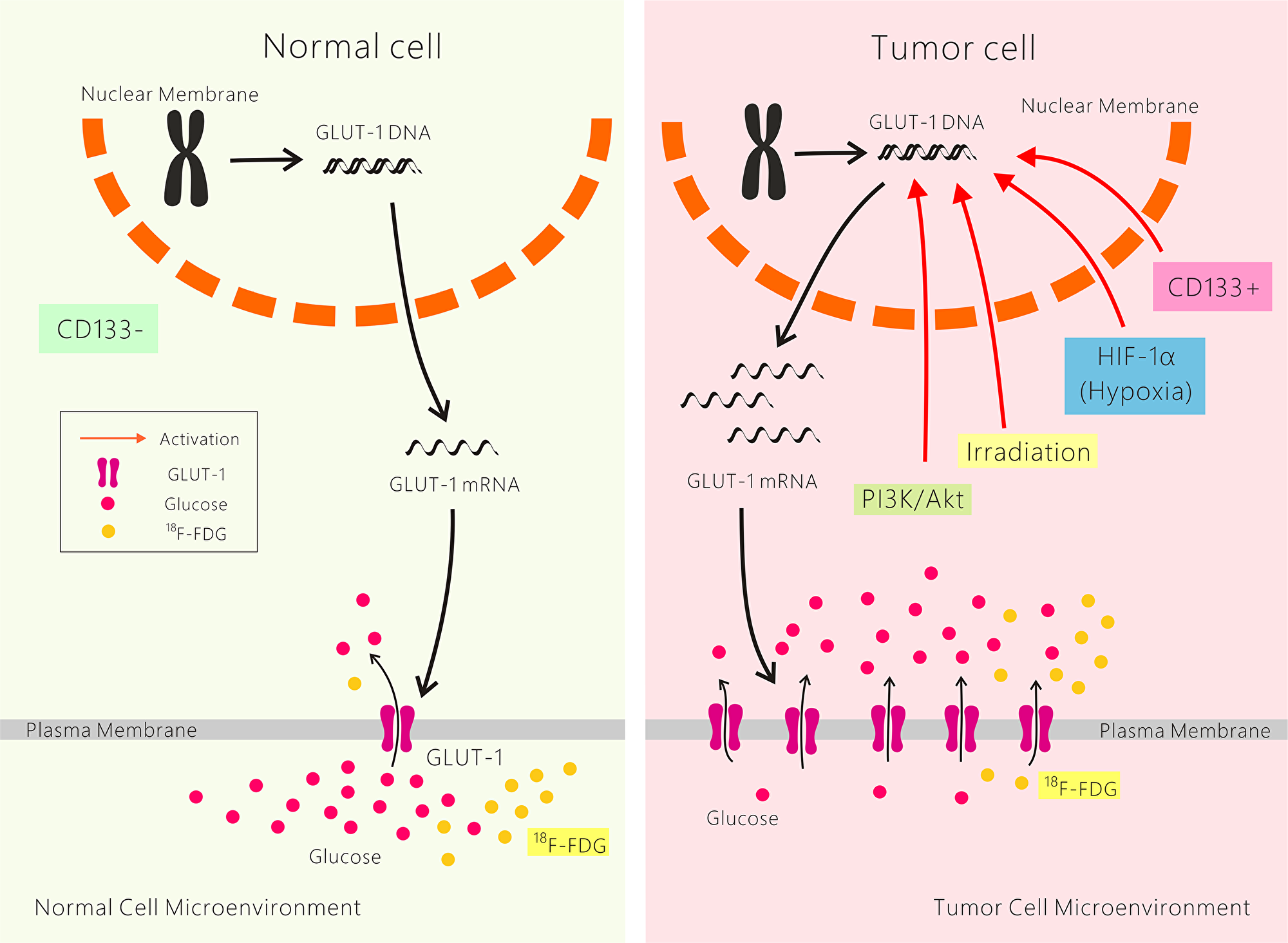
Figure 1: Possible mechanisms of resistance to cancer radiotherapy and chemotherapy mediated by GLUT-1.
GLUT-1 in head and neck cancer
Our previous studies showed high expression of GLUT-1 in head and neck cancer [54], ceruminous adenoma [55], head and neck inflammatory myofibroblastic tumors [56], and nasal and laryngeal neuroendocrine carcinoma [57]. Li detected GLUT-1 expression in all 25 cases of head and neck cancer examined, with expression found mainly in the cell membrane. Poorly differentiated non-keratinizing squamous cell carcinoma cells of the head and neck were located in the central and peripheral nests, while differentiated keratinizing squamous cell carcinoma cells of the head and neck cancer were found in the nest. In addition, the GLUT-1 staining index (SI) was significantly higher in recurrent than in primary head and neck squamous cell carcinoma (P=0.03); the GLUT-1 SI was significantly lower in well-differentiated than in poorly differentiated head and neck squamous cell carcinoma (P=0.02) [58]. However, GLUT-1 expression in different head and neck cancers showed varying relationships with parameters such as progression and prognosis. These differences are reviewed as follows.
Salivary gland tumors
Only a few reports have focused on the clinical significance of GLUT-1 expression in salivary gland tumors. These studies demonstrated that GLUT-1 expression was related to the biological behavior of salivary gland tumors.
Features of GLUT-1 expression in salivary tumors
GLUT-1 expression was not detected in normal salivary gland tissues but wasfound mainly in the cytoplasm of pleomorphic adenomas (PAs), adenoid cystic carcinomas (ACCs), and mucoepidermoid carcinomas (MECs) of the salivary glands by de Souza et al. (2017) [59]. Some cases showed membrane staining only in focal areas. The intensity of immunostaining in the squamous metaplasia areas of PAs, epidermoid cells of MECs, and necrotic areas of ACCs was significantly higher than that in other areas. These findings were also observed by Mori et al. [60] and Bonfitto et al [61]. However, Demasi et al. detected GLUT-1 expression mainly in the cell membrane of higher grade MECs [62].
GLUT-1 may be related to salivary tumor progression
Mori et al. detected higher expression of GLUT-1 mRNA in ACC than in normal salivary glands by RT-PCR [60]. In our previous study, GLUT-1 mRNA and protein showed high expression in ACC-2 cell lines [63]. Demasi et al. found higher expression of GLUT-1 in high-grade MEC than in low-grade MEC of the salivary gland as detected by IHC. They suggested GLUT-1 to be a potential prognostic factor for MEC of the salivary gland. The possible mechanism of GLUT-1 overexpression in some intermediate- and in all high-grade MECs could require a greater energy supply by acquiring the bioenergetic phenotype of increased glycolysis, leading to the outgrowth of MEC cells under hypoxia [62]. Kim et al. (2011) detected GLUT-1 expression in 17 carcinoma ex PAs (CXPAs) of the salivary glands by IHC, with higher expression in the carcinoma components of CXPAs with luminal differentiation than in residual PA components (P=0.024) [64].
GLUT-1 may be related to 18F-fluorodeoxyglucose (FDG) uptake
In the study by Kim et al. (2011), 6 of 17 patients underwent 18F-FDG positron emission tomography (18F-FDG PET) [64], and a relationship between high 18F-FDG uptake and GLUT-1 overexpression was revealed [64].
GLUT-1 may be a poor prognostic factor of salivary tumors
Mori et al. used a Cox proportional hazard model to show that GLUT-1 expression (hazard ratio: 4.739; P=0.022) and lymph metastasis (hazard ratio: 3.944; P=0.042) are significant prognostic factors of malignant salivary tumors. They also found that some clinicopathological parameters, including age (P=0.003), tumor size (P=0.002), and distant metastasis (P=0.007), were significantly correlated with high GLUT-1 expression [60]. The authors suggested the following potential underlying mechanisms: i) overexpression of GLUT-1 leads to a higher glucose supply for malignant salivary tumors to accelerate and progress, ii) hypoxia within malignant salivary tumors are highly invasive, causing metastasis and a poor prognosis, and iii) hypoxia also results in resistance to chemo-radiotherapy [60].
Laryngeal carcinoma
Features of GLUT-1 expression in laryngeal carcinoma
In our previous study, GLUT-1 immunostaining was diffuse in the membranes of laryngeal cancer cells, with a rate of GLUT-1 positivity of 55.1% [65].
Relationship between GLUT-1 overexpression and progression, metastasis, and prognosis of laryngeal carcinoma
Whether the expression of GLUT-1 is a prognostic factor of laryngeal carcinoma remains debatable. In our previous study, we determined the expression of GLUT-1 and GLUT-3 in 38 tissue samples of primary head and neck cancers, including four laryngeal carcinomas, by RT-PCR and Western blotting. We found that high gene and protein expression of GLUT-1, but not GLUT-3, was associated with poor survival [54]. We also investigated the relationship between HIF-1α or GLUT-1 expression and clinicopathological features or prognosis in 49 tissue samples of laryngeal carcinoma. On univariate analysis, GLUT-1 expression was significantly associated with recurrence (P=0.02) and metastasis (P=0.01). On multivariate analysis, GLUT-1 expression (P=0.006), the primary site in supraglottic and subglottic areas (P=0.038), lymph node metastasis (P=0.007), and distant metastasis were associated with poor survival [65]. Starska et al. (2015) found that the GLUT-1 mRNA level was significantly associated with the degree of histological differentiation (P=0.0005), tumor grade (P<0.001), and 5-year survival rate (P=0.012) in 106 laryngeal squamous cell carcinoma cases [66]. However, GLUT-1 and GLUT-3 levels were not associated with prognosis. In their study, a worse overall survival rate was found in cases with high GLUT-1 and GLUT-3 expression, but this was only a trend. At the protein level, higher-grade tumors exhibited higher expression of GLUT-1 [66]. Zuo et al. (2016) evaluated whether hypoxia enhanced the invasion and migration of laryngeal carcinoma in vitro and investigated the relationships between GLUT-1 expression and biological behavior and prognosis in 57 tissue samples of laryngeal carcinoma [67]. They found that 85.96% of the laryngeal carcinoma samples were positive for GLUT-1, whereas 8.0% of normal laryngeal squamous epithelial tissue and 21.05% of laryngeal carcinoma adjacent tissues were positive for GLUT-1 (P<0.05). GLUT-1 overexpression was associated with clinical stage, lymph node metastasis, and poor prognosis. GLUT-1 expression also associated with the expression of epithelial-mesenchymal transition (EMT)-related proteins [67]. Furthermore, molecular mechanism studies showed that hypoxia enhanced laryngeal carcinoma SCC10A cell invasion and migration via EMT [67]. However, Kwon et al. (2014) examined the predictive values of HIF-1α, carbonic anhydrase IX (CAIX), GLUT-1, cyclooxygenase-2, Ki-67, and erythropoietin receptor in 42 tissue samples of T1/T2 early-stage laryngeal carcinomas treated with radiotherapy alone by tissue microarray analysis and IHC and found that only high CAIX was significantly associated with residual tumors on multivariate analysis [68]. Schrijvers et al. (2008) detected three hypoxia markers, HIF-1α, CAIX, and GLUT-1, in 91 tissue samples of T1/T2 glottic carcinoma by IHC and found no significant relationship between GLUT-1 and HIF-1α expression, and GLUT-1 overexpression was not a predictor of worse local control or prognosis [69]. In another study, we investigated the expression of HIF-1α, GLUT-1, and proteins of the PI3K/Akt pathway in 24 tissue samples from patients with laryngeal carcinoma who received PET/computed tomography (CT) preoperatively [70]. However, we found that the maximum SUV (SUVmax) (P=0.043) and PI3K (P=0.012) were significantly poor prognostic factors, and that GLUT-1, HIF-1α, and p-Akt were not correlated with prognosis, recurrence, or metastasis [70].
We suggest that the differential findings regarding the relationship between GLUT-1 expression and prognosis of laryngeal carcinoma may be attributed to differences in histopathological type, IHC technique, tumor stage, and sample number, among studies.
Relationship between the overexpression of GLUT-1 and 18F-fluoroazomycinarabinoside (18F-FAZA) or FDG uptake
Bruine de Bruin L (2015) evaluated the relationships of three endogenous hypoxia markers (HIF-1α, CAIX, and GLUT-1) with FAZA uptake in 11 patients with laryngeal carcinoma indicated for total laryngectomy and found positive immunostaining for GLUT-1 in all biopsies but no relationship of any of these three markers with FAZA uptake [71]. In our previous study, we found that 18F-FDG uptake (SUVmax) was associated with GLUT-1 (r=0.577; P=0.003), HIF-1α (r=1.0; P<0.0001), PI3K (r=1.0; P<0.0001) and p-Akt (r=0.577; P=0.003) expression [70]. in vivo, however, the SUV (tumor/normal tissue) was not associated with GLUT-1 or HIF-1α expression [72].
Targeted GLUT-1 expression as a therapeutic strategy for laryngeal carcinoma in vitro and in vivo
The mechanisms underlying GLUT-1 as a new therapeutic target in laryngeal carcinoma have been discussed in our published review [73]. In vitro, we found that antisense oligodeoxynucleotides (AS-ODNs) against GLUT-1 decreased glucose uptake and proliferation of Hep-2 laryngeal carcinoma cells by reducing GLUT-1 mRNA and protein expression [74]. In a previous study, we successfully isolated CD133+ cancer stem cells (CSCs) from Hep-2 cells and found higher GLUT-1 mRNA and protein expression in CD133+ than in CD133- Hep-2 cells. The proliferation of CD133+ Hep-2 cells was higher than that of CD133- Hep-2 cells. These results suggest that GLUT-1 is the main energy supply for laryngeal CD133+ Hep-2 cells and is a potential therapeutic target for inhibition of laryngeal CSC proliferation [50]. Our subsequent studies demonstrated that inhibition of GLUT-1 expression in laryngeal carcinoma cells may enhance the chemo-radiosensitivity of laryngeal carcinoma. In a previous study, resistance or insensitivity of Hep-2 cells to cisplatin was associated with GLUT-1 expression. Apigenin, a natural phytoestrogen flavonoid present in various fruits, vegetables (especially celery), beans, and tea, significantly inhibited the expression of GLUT-1 and p-Akt expression to improve the sensitivity of Hep-2 cells to cisplatin [75]. In vitro and in vivo, we found that the proliferation of Hep-2 cells did not increase with an increasing X-ray dose, and this accorded with the change in GLUT-1 mRNA expression in Hep-2 cells observed with different X-ray radiation doses. We suggest that GLUT-1 expression plays a role in the radioresistance of Hep-2 cells. Inhibition of GLUT-1 expression in Hep-2 cells via AS-ODNs may gradually decrease survival rate and increase the apoptotic rate with prolonged culture time and increased radiation in vitro. In vivo, the weight of tumors treated with 8 Gy radiation and GLUT-1 AS-ODNs was significantly lower than that after 8 Gy radiation alone. The underlying mechanism was the AS-ODN-induced decrease in GLUT-1 expression [51]. Another mechanistic study of laryngeal carcinoma radioresistance revealed that the PI3K/Akt pathway is involved, mediated by GLUT-1 overexpression [53]. Apigenin may enhance laryngeal carcinoma radiosensitivity through the inhibition of GLUT-1 expression and the PI3K/Akt pathway in vivo [52]. We first demonstrated co-inhibition of GLUT-1 expression via AS-ODNs and the PI3K/Akt pathway via specific inhibitors including Ly294002 and wortmannin. After 10 Gy X-ray radiation, Ly294002, wortmannin, Ly294002 plus GLUT-1 AS-ODNs, and wortmannin plus GLUT-1 AS-ODNs reduced the tumor size significantly compared with tumors treated with 10 Gy X-ray radiation only (P<0.05). Similarly, the expression levels of GLUT-1, p-Akt, and PI3K were significantly decreased by GLUT-1 AS-ODNs and PI3K/Akt inhibitors [53]. The mechanism of laryngeal carcinoma radioresistance may involve multiple factors. In our recent study using Hep-2 and Tu212 laryngeal carcinoma cells, 10 Gy X-ray radiation decreased the weight of Hep-2 and Tu212 xenografts, while GLUT-1 AS-ODNs decreased the weight of Tu212 xenografts only. Combined with HIF-1α, GLUT-1 AS-ODNs may significantly increase apoptosis and decrease microvessel density, the apoptotic index, and necrosis after X-ray irradiation in vivo [72].
Oral carcinoma
Expression of GLUT-1 in oral carcinoma
GLUT-1 expression was confirmed in 100% of 50 cases of oral squamous cell carcinoma by IHC staining [76]. Pereira et al. (2016) detected GLUT-1 in 15 samples from patients with oral epithelial dysplasia (OED) and 15 samples from patients with oral squamous cell carcinoma (OSCC) by IHC. GLUT-1 expression was positive in all cases of OED and OSCC. GLUT-1 immunostaining was greater in OED than that in OSCC, suggesting that GLUT-1 is expressed during the initial stages of oral carcinoma [77]. Leite et al. (2017) detected GLUT-1 and GLUT-3 expression in both keratocystic odontogenic tumors associated with Gorlin syndrome (SKOTs) and non-syndromic keratocystic odontogenic tumors (NSKOTs) by IHC. They revealed positive GLUT-1 expression in the epithelial component in all cases [78]. They found that GLUT-1 and GLUT-3 were not associated with the angiogenic index in SKOTs, primary NSKOTs, or recurrent NSKOTs [78].
Relationship among GLUT-1, differentiation of oral carcinoma, and cellular distribution
Azad et al. found that the expression of GLUT-1 in oral squamous cell carcinoma was also closely related to smoking history (P<0.001), Bryne grade (P<0.001), tumor size (P=0.001), lymph node metastasis (P=0.022), and clinical stage (P<0.001).GLUT-1 also shows a progressive switch from membranous to cytoplasmic to a combined location and is correlated with histopathologic grade and pTNM stage. As tumors become more malignant, higher levels of GLUT-1 accumulate at the cell membrane to allow transport of glucose into cells, so that tumors can proliferate rapidly [76]. That report suggested that mature squamous epithelium is associated with glycogen accumulation during tumor formation. The presence of glycogen is related to the maturation of squamous epithelial cells, in that it disappears with the loss of differentiation during neoplastic transformation. In well-differentiated tumors, increased accumulation of glycogen in keratin pearls is inversely correlated with GLUT-1 immunostaining, suggesting that differentiated and mature cells within keratinized regions lack GLUT-1 expression. In contrast to poorly differentiated tumors, hypoxia-stimulated GLUT-1 creates an antistromal staining pattern in the absence of squamous epithelial differentiation or keratinization [76]. Among 57 cases of tongue squamous cell carcinoma, 52.6% had positive GLUT-1 expression in the tumor periphery and 47.4% in the tumor center, and this was associated with the classification of the tongue cancer tissue. In advanced tongue squamous cell carcinoma, 90.7% of samples were strongly positive for GLUT-1, with low levels of GLUT-1 in tumor cells in the tumor periphery and high levels in the central tumor [79]. A recent interesting study investigated the role of oral brush biopsy and GLUT-1 staining in 72 patients, including 24 healthy patients, 15 with carcinoma, 18 with leukoplakia, and 15 with oral lichen planus. The results showed positive GLUT-1 expression in 30 (41.7%) patients, including 80% (12) with HNSCC, 60% (9) with oral lichen planus, and 50% (9) with leukoplakia. No GLUT-1 staining was found in the healthy patients [80]. The authors suggested that GLUT-1 overexpression is likely an early event in carcinogenesis with prognostic and therapeutic value [80].
GLUT-1 and TNM stage of oral carcinoma
Ohba et al. found that GLUT-1 expression was related to the depth of oral cancer invasion. A significant difference was found in GLUT-1 staining intensity between tumors with < 4 and those with > 4 mm invasion depth (P=0.023), indicating that GLUT-1 expression is a marker of the extent of tumor invasion [81].
GLUT-1 expression and prognosis of oral carcinoma
Ayala et al. found that GLUT-1 was highly expressed in 142 patients with oral squamous cell carcinoma, while GLUT-3 was highly expressed in 21.1% of these patients, and high protein expression of GLUT-1 and GLUT-3 was associated with poor prognosis [82]. Studies have found that GLUT-1 expression alone cannot be used as an independent prognostic marker for head and neck cancer. Grimm found that increased expression levels of both GLUT-1 and transketolase-like protein 1 are markers of poor prognosis in oral cancer [83]. Eckert found that high expression levels of both GLUT-1 and HIF-1a are markers of poor prognosis in oral cancer [84].
Thyroid cancer
GLUT-1 expression
GLUT-1 expression in thyroid cancer is controversial. In vitro, GLUT-1 expression is significantly higher in anaplastic thyroid cancer cells than in normal cells [85]. Musholt et al. (1997) detected GLUT-1–5 in 10 medullary thyroid carcinoma (MTC) samples by Western blotting, one of which exhibited very weak GLUT-1 expression [86]. Kaida et al. (2011) evaluated the relationship between 18F-FDG uptake and GLUT expression or clinicopathological factors in 54 patients with papillary thyroid cancer (PTC). The results revealed positive expression of GLUT-1, GLUT-3, and GLUT-4 in the cytoplasm and/or membrane of PTC cells [87]. GLUT-3 and GLUT-4 expression was stronger than GLUT-1 expression. A GLUT-1 expression score of 3 was found in only one tumor with anaplastic features, suggesting that strong GLUT-1 expression may occur only in PTC with anaplasia [87]. Among 10 patients with thyroid tissue showing thymus-like elements (CASTLE), GLUT-1 expression was positive in all [88]. The authors suggested that GLUT-1 is a novel biomarker for CASTLE with potential diagnostic value [88]. Kim et al. (2013) detected a rate of GLUT-1 positivity of 66.7% in 188 patients with PTC who underwent 18F-FDG-PET/CT examination [89]. However, Chandan et al. (2006) detected CD57 and GLUT-1 expression in 50 thyroid fine-needle aspiration samples, including 15 papillary carcinoma, 14 atypical cytology, and 21 benign thyroid cases. They found that 20 of 29 cases with malignant lesions were positive, while all 21 benign thyroid cases were negative, for CD57 expression; however, GLUT-1 expression was negative in all 50 malignant and benign thyroid lesions [90].
GLUT-1 expression, biological behavior, and prognosis of thyroid cancer
Kim et al. (2014) investigated the expression of GLUT-1 and major thyroid-specific genes in 24 PTC tissue samples by RT-PCR [92]. They detected higher GLUT-1 expression in the less-differentiated group than in the well-differentiated group among 23 cases [91]. Lodewijk et al. (2017) detected HIF-1α, CAIX, and GLUT-1 expression in 111 MTC tissue samples by IHC. On univariate analysis, HIF-1α was associated with overall survival and progression-free survival (PFS), and GLUT-1 was significantly associated with PFS, as well as TNM stage, the presence of lymph node metastasis, heritability, and necrosis. However, only HIF-1α, TNM stage, and heritability were prognostic factors on Cox regression analysis (r=0.21; P=0.147) [92]. Kaida et al. (2011) found that the prognosis of PTC was not correlated with age, sex, extrathyroid extension, SUVmax, or GLUT-1, GLUT-3, or GLUT-4 expression [87]. Kim et al. (2013) found that GLUT-1 expression was not associated with SUVmax, extrathyroidal extension, lymph node metastasis, or advanced tumor stage in patients with primary thyroid cancer [93].
18F-FDG uptake
The findings of Kaida et al. (2011) showed that the SUVmax was associated with GLUT-3 (r=0.38; P=0.008) and GLUT-4 (r=0.46; P=0.001) but not GLUT-1 expression (r=0.21; P=0.147) [87]. Among 38 patients with thyroid papillary cancer with recurrent cervical nodal metastases, 21 (55%) exhibited positive 18F-FDG uptake, which was related to thyroid globulin expression and GLUT-1 membrane expression with luminal accentuation. However, 18F-FDG uptake was not associated with GLUT-1 cytoplasmic expression [94]. Kim et al. (2013) found that 18F-FDG uptake was not associated with GLUT-1 or HIF-1α expression [89].
HK-II
HK-II biological characteristics
HK is a key enzyme in glucose metabolism [95]. Mammals have four isoforms of HK (HKs I–IV). HK isoforms exhibit some organizational differences and specific distribution patterns in vivo [96]. HK-I is expressed mainly in the brain, HK-II mainly in insulin-sensitive tissue such as myocardial and skeletal muscle and adipose tissue, HK-III in kidney, liver, and intestinal tissues, and HK-IV in the liver and pancreas. When glucose enters the cell, the first step is its phosphorylation into glucose-6 phosphate, which is not able to cross the cell membrane. The first key rate-limiting enzyme in this process is HK [96]. In normal tissue, free HK molecules are predominant. However, in tumor tissues, HK can combine with mitochondria, forming particles of HK, of which the N-terminal domain has a hydrophobic end connected to the outer mitochondrial membrane; this then forms a complex with the mitochondrial permeability tunnel complex of the voltage dependent anion channel protein (VDAC) binding to HK and forming HK-VDAC [51]. HK-VDAC can enhance the ability of ATP to bind to mitochondria and to supply tumor cells with energy. HK-VDAC is also a major contributor to the immortalization of cancer cells [97].
Studies have shown that the disruption of HK-VDAC can lead to apoptosis via the PI3K/Akt signaling pathway [98]. However, the high levels of lactic acid produced by glycolysis can help tumor cells escape from immune detection and allow their rapid proliferation [96]. The four subtypes of HK (I-IV) are highly expressed in malignant tumors, with HK-II being the most highly expressed, and the proportion of HK-IIb in microparticles is higher than that of the other subtypes [97].
HK-II and malignant tumors
HK-II expression in malignant tumors
Several research reports have shown increased HK-II expression in many malignant tumors, including nasopharyngeal cancer, ovarian cancer, renal cell carcinoma, hepatocellular carcinoma, colon cancer, and glioma [98–100]. A five-fold increase in the gene expression of HK-II, but not the other HK isoforms, was detected in liver tumors, and this is thought to accelerate glycolysis in hepatoma cells to provide extra energy [97]. Guzman et al. found that the expression of HK-II was significantly higher in hepatocellular carcinoma than in the control group, and that the high HK-II expression was correlated with invasiveness and high tumor grade [100]. Wolf et al. found that in 25 patients with pleomorphic gliomas of the brain, 20 showed HK-II expression in the brain but not in normal human brain white matter [101].
The expression level of HK-II is 200-fold higher in malignant tumor tissues than in normal tissues. Moreover, it was found that the rate of glycolysis in hepatocytes was significantly increased after the introduction of mitochondrial binding HK-II [102]. Guzman also found that the expression of HK-II increased incrementally from normal liver tissue to the compensated and decompensated stages of liver cirrhosis to the development of liver cancer [100]. This pattern would suggest that HK-II increases during tumor development from normal tissue to precancerous lesions, playing an important role in tumor development.
The relationships between HK-II and the clinical stage, differentiation, metastasis, and prognosis of malignant tumors
HK-II was found to be related to the clinical stage, differentiation, metastasis, and poor prognosis of malignant tumors [103–108]. Hamabe et al. analyzed 104 cases of colorectal cancer by IHC, dividing the samples into HK-II expression-positive and -negative groups. They found that the expression of HK-II was related to tumor diameter (P=0.046), depth of tumor invasion (P=0.0395) and lymph node metastasis (P=0.0409) in the HK-II-positive group [83]. Jin et al. found that HK-II expression was significantly higher in phase III/IV than phase III ovarian cancer (P<0.001), and HK-II expression was significantly higher in the poorly differentiated than the highly differentiated group (P=0.008) [98]. Other reports have shown that the expression of HK-II in phase II to IV was significantly higher than that in phase I cancer (P=0.044) [100]. The high expression of HK-II in colorectal cancer was related to tumor size, depth of tumor invasion, liver metastasis, recurrence, and TNM stage. Multivariate analysis showed that HK-II was an independent prognostic factor for colorectal cancer [109]. Similar results were found in liver cancer [103], pancreatic cancer [110], cervical cancer [111], gastric cancer [106], breast cancer [112], and lung cancer [113].
HK-II in head and neck cancer
To our knowledge, few studies have focused on HK-II in head and neck cancer. Thus, this section has not been subdivided according to different tissues and cell types of head and neck tumors.
The relationship between HK-II and head and neck cancer
Chen et al. found that the expression of HK-II in laryngeal carcinoma tissue, laryngeal papilloma, and laryngeal polyps was 100%, 37%, and 10%, respectively [114]. They suggested that the expression of HK-II was related to the occurrence and progression of laryngeal carcinoma [114]. Tian et al. detected HK-II expression in all 19 cases of oral squamous cell carcinoma examined [115].
The relationship between HK-II and the stage, differentiation, metastasis, and prognosis of head and neck cancer
Chen et al. found that the expression of HK-II was associated with the tumor site and TNM stage of laryngeal carcinoma [114]. A possible mechanism may be that glucose phosphorylation and HK-II binding to the mitochondrial membrane were increased, which inhibited cytochrome C release and apoptosis and reduced glucose-6-phosphate negative feedback on glycolysis; this may have increased glycolysis and substrate product biosynthesis, increasing the growth of cancer cells and acidulating the cancer microenvironment to promote tumor invasion [114].
Xiao et al. performed a retrospective analysis of 41 patients with nasopharyngeal carcinoma who received radiotherapy or chemoradiotherapy. They found that the median survival of 21 patients with high HK-II expression was 62.29 months, and that of 20 patients with low HK-II expression was 93.60 months. The high expression of HK-II was associated with prognosis. It was also found that inhibition of HK-II expression enhanced the radiosensitivity of nasopharyngeal carcinoma [116].
Possible mechanism of radioresistance caused by HK-II in cancer
The underlying mechanism of HK-II-induced radioresistance in laryngeal carcinoma has been described in our previous review. HIF-1α, suppressor oncogenes, miR-155 may involve in the mechanisms of radioresistance.
GLUT-1 and HK-II
GLUT-1, HK-II, and the Warburg effect
The Warburg effect has been confirmed by uptake of 18F-FDG), an analogue of glucose, on PET/CT. 18F-FDG is transported into the cell by GLUT after intravenous injection and is phosphorylated by HK to generate 18F-FDG -6-phosphate, which is not further metabolized and thus deposited in cells, showing a high metabolic concentration on PET images [117].
Expression of GLUT-1 and HK-II in malignant tumors and their correlation with FDG uptake
Several studies have shown that GLUT-1 and HK-II expression levels are related to tumor FDG uptake, but there are differences among tumor types. The uptake of FDG by some malignant tumors was related to tumor expression levels of GLUT-1 or HK-II, or both; however, uptake of FDG by other tumors was independent of either GLUT-1 or HK-II expression [120]. Therefore, the mechanism of FDG uptake in malignant tumors requires further study.
Several studies have reported a relationship between GLUT-1 and HK-II expression in malignant tumors [119, 120]. The rapid uptake of glucose via GLUT-1 in cancer cells is maintained mainly by HK-II. Grimm et al. found that GLUT-1 and HK-II expression significantly increased with the progression from normal oral mucosa, dysplasia, and neoplasia at stages I, II, and III to invasive squamous cell carcinoma [119]. The expression of GLUT-1 in oral squamous cell carcinoma was nearly four-fold higher than that in normal tissues [119]. Pauday et al. found that GLUT-1 expression was associated with HK-II expression in hepatobiliary cancer cells (P=0.002, P=0.3), and FDG uptake was also associated with GLUT-1 or HK-II expression [120]. In the breast cancer cells of nude mice, Kristian et al. used dynamic PET/CT to show that a high initial FDG uptake was positively correlated with increased expression of GLUT-1, and FDG uptake during the late phase was associated with HK-II expression. Therefore, GLUT-1 and HK-II may play roles during different periods of FDG uptake [121]. Cancer cells significantly increase their demand for glucose as malignancy progresses, and GLUT-1 and HK-II simultaneously increase in expression, which is associated with the SUV. These studies suggest that GLUT-1 and HK-II play a synergistic role in FDG uptake during the progression of malignant tumors.
Inconsistent results were reported for some malignant tumors, however. While HK-II was associated with GLUT-1 expression during stages T3 and T4 of oral squamous cell carcinoma (R=0.99, P=0.0001), there was no correlation between the expression of HK-II and FDG uptake [115]. In rectal cancer, Izuishi et al. found that GLUT-1 and HK-II expression was significantly higher in cancerous tissue than in normal tissue surrounding the mucosa, whereas FDG uptake was not associated with GLUT-1 or HK-II expression. They suggested that aberrant glucose-metabolizing pathways in cancer may not be regulated by a single molecule [122]. Although Yoon et al. found that GLUT-1 and HK-II expression levels were as high as 81% and 77%, respectively, in extrahepatic bile duct carcinoma, there were no correlation between their expression levels [4]. The SUV or SUV tumor-to-liver ratio was associated with GLUT-1 expression (ρ=0.648, P=0.0003 and ρ=0.703, P<0.0001, respectively), but not with HK-II expression [4]. Yang et al. detected the expression of HK-II and GLUT-1 using IHC in 50 patients with pancreatic cancer who underwent preoperative PET/CT and found that the expression of GLUT-1 and HK-II in cancer tissues was higher compared with the adjacent tissue (P<0.001). However, there was no correlation between FDG uptake and the expression of GLUT-1 or HK-II or between GLUT-1 and HK-II [118]. Higashi et al. also found a high correlation between GLUT-1 and HK-II expression in pancreatic cancer but no association with FDG uptake (P=0.055 and P=0.1852, respectively) [123]. Cho et al. found that GLUT-1 and HK-I expression levels were increased with higher grades of gastrointestinal stromal tumor risk. However, HK-II and several other markers (i.e., GLUT-2, -3, and -4) were not correlated with tumor risk grade. FDG absorption was related to the expression of GLUT-1 and HK-I [117]. Baschnagel et al. found that the expression of HK-II was correlated with the epidermal growth factor receptor, but not with p16 or GLUT-1, in 97 patients with locally advanced head and neck squamous cell carcinoma. The SUVmax was associated with HK-II expression (P=0.021), but not GLUT-1 expression [124]. Zhou et al. found that FDG uptake was associated with the expression of GLUT-1 and lactate dehydrogenase (LDHA), but not HK-II, in 51 cases of lung adenocarcinoma. In vitro, they found that knockdown of LDHA can reduce FDG uptake, GLUT-1 expression, and cell proliferation [125].
Park et al. found an absence of HK-II expression in 19 cases of malignant melanoma, and FDG uptake was correlated with the expression of GLUT-1 and GLUT-3, mainly GLUT-1 [126]. In five different breast cancer xenograft models, HK-II protein expression was significantly related to FDG absorption (r2=0.339, P=0.001), and HK-II was an independent predictor of FDG uptake. There was a relationship between GLUT-1 and FDG uptake only in tumors driven by Akt or the HER2/neu gene [127]. Takebayashi et al. reported high expression of HK-I and HK-II in 50 cases of gastric carcinoma; however, no relationship was detected between FDG uptake and GLUT-1 or HK-II expression [128].
These results suggest that in some malignant tumors, GLUT-1 and HK-II are not the only critical rate-limiting enzymes, and that HK-II expression is not completely consistent with GLUT-1 expression. Yang et al. suggested that glucose metabolism is a complex process involving multiple factors, including GLUT-2, -3, and -4 and HK-I; however, the actual mechanism requires further study [118].
The effect of hypoxia on the expression of GLUT-1 and HK-II in head and neck cancer
Hypoxia is a key regulator of the expression of GLUT-1 and HK-II. Malignant tumors, including head and neck cancers, arise in hypoxic environments [66]. Hypoxia can alter tumor cells physiologically and biochemically by regulating the expression of a variety of target genes, including GLUT-1 and HK-II, involved in adaption to the hypoxic environment.
HIF-1α plays an important role in the adaptive cellular response to hypoxia [26]. Co-expression of GLUT-1, HK-II, and HIF-1α has been observed in numerous tumor types [42, 129, 130]. HIF-1α can activate the expression of target genes such as GLUT-1, HK-II, and VEGF [26, 42, 129, 130].
In vitro, Iwamoto et al. found that 18F-FDG was significantly accumulated in KRAS-mutated cells compared with wild-type cells under normoxic conditions, and that GLUT-1 and HK-II mRNA levels were high. FDG uptake was associated mainly with GLUT-1 expression. A possible mechanism may be that the Raf/MEK/ERK pathway is correlated with GLUT-1 expression in KRAS-mutated cells, while the PI3K/Akt pathway is correlated with HK-II expression in both KRAS-mutated and wild-type cells [131]. Under hypoxia, the expression of HIF-1α was higher in KRAS-mutated cells compared with wild-type cells. Overexpression of HIF-1α resulted in higher GLUT-1 expression and 18F-FDG accumulation. That same study showed that increased FDG uptake was related mainly to GLUT-1 expression in vivo [131]. In acute myeloid leukemia, HIF-1α, GLUT-1, and HK-II were highly expressed [132]. HIF-1, GLUT-1, and HK-II were also highly expressed in 36 cases of stage T1 and T2 squamous cell carcinoma of the oral cavity and were significantly correlated with FDG uptake; furthermore, the expression of HIF-1 was related to the expression of GLUT-1 and HK-II [133]. Kim et al. (2017) detected glycolysis markers, including GLUT-1, HK-II, CAIX, and MCT4, in 265 follicular neoplasm (FN) samples and 108 Hürthle cell neoplasm (HCN) samples by tissue microarray and IHC staining and found that these markers were highly expressed in HCNs compared with in FNs, with the highest expression in Hürthle cell carcinoma (HCC) followed by Hürthle cell adenoma, follicular carcinoma (FC) and follicular adenoma (FA), in that order (all P<0.001). HK-II expression was correlated with a larger HCC tumor size (> 4 cm) (r=0.384; P=0.046). Expression of the Ki67 proliferation index was related to GLUT-1 expression in FC (r=0.419; P=0.029). In this study, however, the expression of the glycolysis markers GLUT-1, HK-II, CAIX, and MCT4 was not significantly related to the prognosis of FC or HCC on multivariate analysis [134].
GLUT-1, HK-II, and tumor-targeting therapy
As mentioned above, the development, metastasis, and poor prognosis of some malignant tumors were correlated with abnormal expression of GLUT-1 and HK-II. Many studies have found that GLUT-1 and HK-II expression is related to chemo- and radioresistance in some malignant tumors, and GLUT-1 and/or HK-II may represent anti-cancer therapeutic targets (Figure 2) [73, 95].
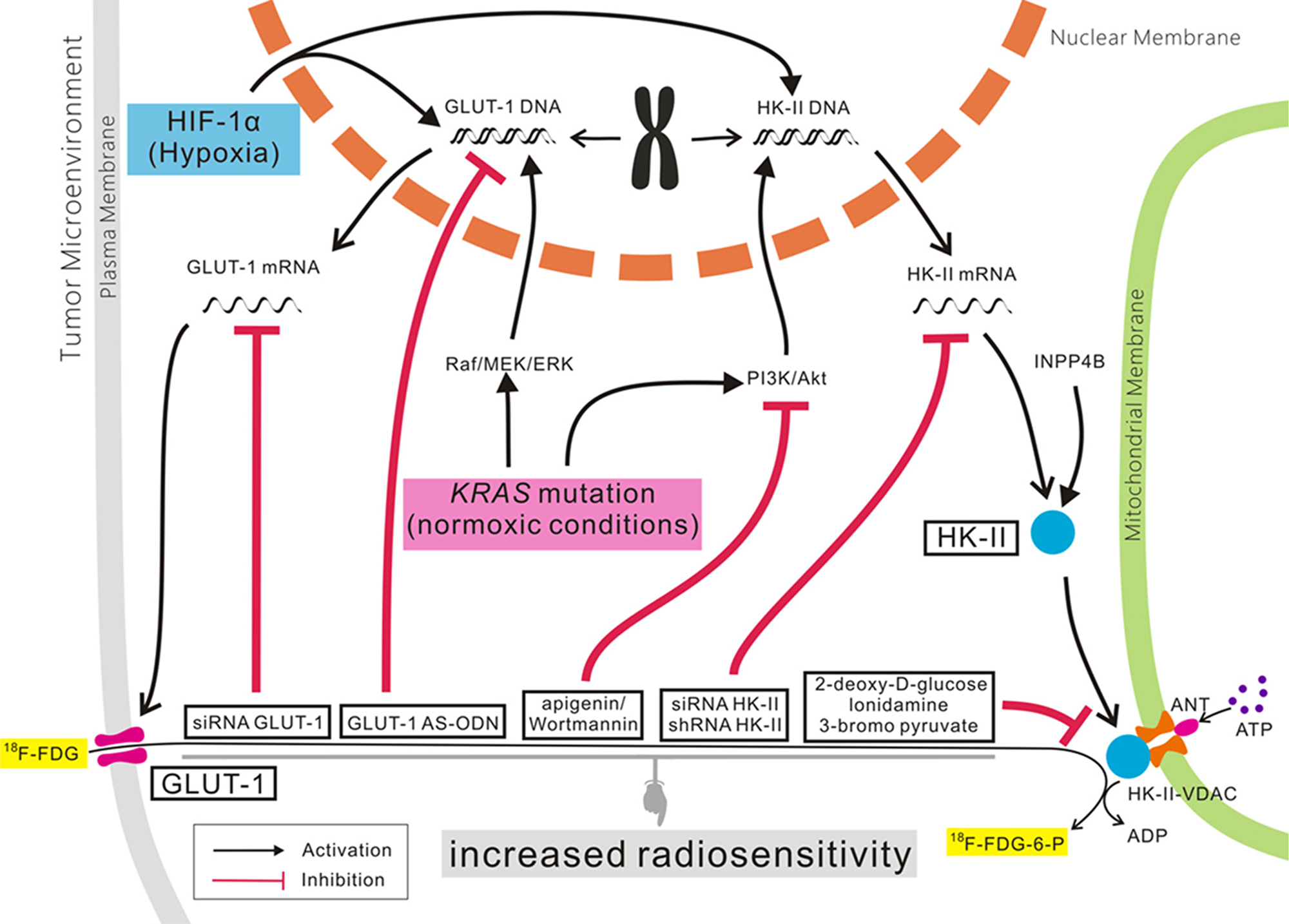
Figure 2: The role of GLUT-1 and HK-II expression in cancer.
Our previous studies found that high expression of GLUT-1 was associated with radioresistance in laryngeal carcinoma, and that inhibition of GLUT-1 expression by antisense oligodeoxynucleotides (AS-ODNs) may improve the radiosensitivity of laryngeal carcinoma, in vitro and in vivo [51, 74]. In vivo, apigenin/wortmannin or apigenin/wortmannin plus GLUT-1 AS-ODNs improved the radiosensitivity of laryngeal carcinoma [52, 53].
Similar results have been reported in other malignant tumors [135–137]. A GLUT-1-specific AS-ODN or siRNA, combined with inhibition of the PI3K/Akt signaling pathway, can improve the radiosensitivity of laryngeal cancer cells to some degree. However, this effect was found mainly in vitro, with unsatisfactory results in vivo. Therefore, some authors have suggested that it may be more effective to improve radiosensitivity via combined inhibition of GLUT-1 and other proteins involved in the Warburg effect, such as HIF-1 or HK-II.
As mentioned above, high expression of HK-II is associated with chemo- and radioresistance [138–145]. In recent years, research has increasingly focused on HK-II as a target for anti-tumor therapies, e.g., 2-deoxy-D-glucose [141], lonidamine [142], 3-bromo pyruvate [143]. Inhibition of HK-II expression in cancer cells using a targeted HK-II-VDAC complex [144] and siRNAs [145] induced apoptosis of cancer cells.
Chen et al. found that high expression of HK-II was related to the TNM stage of laryngeal carcinoma [114]. It was found in vivo and in vitro that inhibition of HK-II expression by shRNAs inhibited the growth of laryngeal carcinoma [114]. Min et al. found that inositol polyphosphate 4-phosphatase (INPP4B) can regulate HK-II-induced glycolysis, leading to radioresistance in laryngeal carcinoma. Inhibition of HK-II expression alone did not improve the sensitivity to radiotherapy and chemotherapy in laryngeal carcinoma cells, while co-inhibition of INPP4B and HK-II did [138].
Conclusions and therapeutic perspectives
These results suggest that simultaneous inhibition of HK-II and glycolysis-related genes may improve the radiosensitivity of laryngeal carcinoma. GLUT-1 and HK-II play important roles in different processes of glycolysis in malignant tumors and may be associated with radioresistance. Inhibition of GLUT-1 or HK-II can increase radiosensitivity to some extent. Therefore, we propose that combined inhibition of GLUT-1 and HK-II expression may enhance tumor chemo- and radiosensitivity and provide new therapeutic targets for malignant tumors.
Abbreviations
GLUT-1: Glucose transporter 1; HK-II: hexokinase-II; HIF-1α: hypoxia inducible factor-1α; CA-IX: carbonic anhydrase-IX; VDAC: voltage dependent anion channel; PET: protein, positron emission computed tomography; LDHA: lactate dehydrogenase; AS-ODN: antisense oligodeoxynucleotides; 3-BrPA: 3-bromo pyruvate; INPP4B: Inositol polyphosphate 4-phosphatase).
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
FUNDING
This research was supported by National Natural Science Foundation of China (No.81372903), and Science and Technology Department of Zhejiang Province, China (No.2016C33144).
References
1 . Morris LG, Chan TA. Therapeutic targeting of tumor suppressor genes. Cancer. 2015; 121:1357–68.
2 . San-Millán I, Brooks GA. Reexamining cancer metabolism: lactate production for carcinogenesis could be the purpose and explanation of the Warburg Effect. Carcinogenesis. 2017; 38:119–33.
3 . Hu ZY, Xiao L, Bode AM, Dong Z, Cao Y. Glycolytic genes in cancer cells are more than glucose metabolic regulators. J Mol Med (Berl). 2014; 92:837–45.
4 . Yoon SO, Jeon TJ, Park JS, Ryu YH, Lee JH, Yoo JS, Kim JK, Yoon DS, Oh EJ. Analysis of the roles of glucose transporter 1 and hexokinase 2 in the metabolism of glucose by extrahepatic bile duct cancer cells. Clin Nucl Med. 2015; 40:e178–82.
5 . Pereira KM, Chaves FN, Viana TS, Carvalho FS, Costa FW, Alves AP, Sousa FB. Oxygen metabolism in oral cancer: HIF and GLUTs (Review). Oncol Lett. 2013; 6:311–16.
6 . Kang F, Ma W, Ma X, Shao Y, Yang W, Chen X, Li L, Wang J. Propranolol inhibits glucose metabolism and 18F-FDG uptake of breast cancer through posttranscriptional downregulation of hexokinase-2. J Nucl Med. 2014; 55:439–45.
7 . Evans A, Bates V, Troy H, Hewitt S, Holbeck S, Chung YL, Phillips R, Stubbs M, Griffiths J, Airley R. Glut-1 as a therapeutic target: increased chemoresistance and HIF-1-independent link with cell turnover is revealed through COMPARE analysis and metabolomic studies. Cancer Chemother Pharmacol. 2008; 61:377–93.
8 . Vartanian A, Agnihotri S, Wilson MR, Burrell KE, Tonge PD, Alamsahebpour A, Jalali S, Taccone MS, Mansouri S, Golbourn B, Aldape KD, Zadeh G. Targeting hexokinase 2 enhances response to radio-chemotherapy in glioblastoma. Oncotarget. 2016; 7:69518–35. https://doi.org/10.18632/oncotarget.11680.
9 . Wang L, Wang J, Xiong H, Wu F, Lan T, Zhang Y, Guo X, Wang H, Saleem M, Jiang C, Lu J, Deng Y. Co-targeting hexokinase 2-mediated Warburg effect and ULK1-dependent autophagy suppresses tumor growth of PTEN- and TP53-deficiency-driven castration-resistant prostate cancer. EBioMedicine. 2016; 7:50–61.
10 . Kasahara M, Hinkle PC. Reconstitution and purification of the D-glucose transporter from human erythrocytes. J Biol Chem. 1977; 252:7384–90.
11 . Deng D, Xu C, Sun P, Wu J, Yan C, Hu M, Yan N. Crystal structure of the human glucose transporter GLUT1. Nature. 2014; 510:121–25.
12 . Berlth F, Mönig S, Pinther B, Grimminger P, Maus M, Schlösser H, Plum P, Warnecke-Eberz U, Harismendy O, Drebber U, Bollschweiler E, Hölscher A, Alakus H. Both GLUT-1 and GLUT-14 are Independent Prognostic Factors in Gastric Adenocarcinoma. Ann Surg Oncol. 2015; 22:S822–31.
13 . Wincewicz A, Baltaziak M, Kanczuga-Koda L, Koda M, Sulkowska U, Sulkowski S. GLUT1 and Bcl-xL in relation to erythropoietin in human colorectal adenocarcinomas. Hepatogastroenterology. 2010; 57:741–45.
14 . Sasaki H, Shitara M, Yokota K, Hikosaka Y, Moriyama S, Yano M, Fujii Y. Overexpression of GLUT1 correlates with Kras mutations in lung carcinomas. Mol Med Rep. 2012; 5:599–602.
15 . Jiwa LS, van Diest PJ, Hoefnagel LD, Wesseling J, Wesseling P, Moelans CB, and Dutch Distant Breast Cancer Metastases Consortium. Upregulation of Claudin-4, CAIX and GLUT-1 in distant breast cancer metastases. BMC Cancer. 2014; 14:864.
16 . Kobayashi M, Kaida H, Kawahara A, Hattori S, Kurata S, Hayakawa M, Hirose Y, Uchida M, Kage M, Fujita H, Hayabuchi N, Ishibashi M. The relationship between GLUT-1 and vascular endothelial growth factor expression and 18F-FDG uptake in esophageal squamous cell cancer patients. Clin Nucl Med. 2012; 37:447–52.
17 . Alakus H, Batur M, Schmidt M, Drebber U, Baldus SE, Vallböhmer D, Prenzel KL, Metzger R, Bollschweiler E, Hölscher AH, Mönig SP. Variable 18F-fluorodeoxyglucose uptake in gastric cancer is associated with different levels of GLUT-1 expression. Nucl Med Commun. 2010; 31:532–38.
18 . Semaan A, Munkarah AR, Arabi H, Bandyopadhyay S, Seward S, Kumar S, Qazi A, Hussein Y, Morris RT, Ali-Fehmi R. Expression of GLUT-1 in epithelial ovarian carcinoma: correlation with tumor cell proliferation, angiogenesis, survival and ability to predict optimal cytoreduction. Gynecol Oncol. 2011; 121:181–86.
19 . Zimmerman RL, Fogt F, Burke M, Murakata LA. Assessment of Glut-1 expression in cholangiocarcinoma, benign biliary lesions and hepatocellular carcinoma. Oncol Rep. 2002; 9:689–92.
20 . Kumar D. Regulation of glycolysis in head and neck squamous cell carcinoma. Postdoc J. 2017; 5:14–28.
21 . Koch A, Lang SA, Wild PJ, Gantner S, Mahli A, Spanier G, Berneburg M, Müller M, Bosserhoff AK, Hellerbrand C. Glucose transporter isoform 1 expression enhances metastasis of malignant melanoma cells. Oncotarget. 2015; 6:32748–60. https://doi.org/10.18632/oncotarget.4977.
22 . Lu K, Yang J, Li DC, He SB, Zhu DM, Zhang LF, Zhang XU, Chen XC, Zhang B, Zhou J. Expression and clinical significance of glucose transporter-1 in pancreatic cancer. Oncol Lett. 2016; 12:243–49.
23 . Yang J, Wen J, Tian T, Lu Z, Wang Y, Wang Z, Wang X, Yang Y. GLUT-1 overexpression as an unfavorable prognostic biomarker in patients with colorectal cancer. Oncotarget. 2017; 8:11788–96. https://doi.org/10.18632/oncotarget.14352.
24 . Al-Sharaky DR, Abdou AG, Wahed MM, Kassem HA. HIF-1α and GLUT-1 Expression in Atypical Endometrial Hyperplasia, Type I and II Endometrial Carcinoma: A Potential Role in Pathogenesis. J Clin Diagn Res. 2016; 10:EC20–27.
25 . Lee DW, Chong GO, Lee YH, Hong DG, Cho YL, Jeong SY, Park JY, Lee YS. Role of SUVmax and GLUT-1 Expression in Determining Tumor Aggressiveness in Patients With Clinical Stage I Endometrioid Endometrial Cancer. Int J Gynecol Cancer. 2015; 25:843–49.
26 . Iwasaki K, Yabushita H, Ueno T, Wakatsuki A. Role of hypoxia-inducible factor-1α, carbonic anhydrase-IX, glucose transporter-1 and vascular endothelial growth factor associated with lymph node metastasis and recurrence in patients with locally advanced cervical cancer. Oncol Lett. 2015; 10:1970–78.
27 . Schlößer HA, Drebber U, Urbanski A, Haase S, Baltin C, Berlth F, Neiß S, von Bergwelt-Baildon M, Fetzner UK, Warnecke-Eberz U, Bollschweiler E, Hölscher AH, Mönig SP, Alakus H. Glucose transporters 1, 3, 6, and 10 are expressed in gastric cancer and glucose transporter 3 is associated with UICC stage and survival. Gastric Cancer. 2017; 20:83–91.
28 . Sadlecki P, Bodnar M, Grabiec M, Marszalek A, Walentowicz P, Sokup A, Zegarska J, Walentowicz-Sadlecka M. The role of Hypoxia-inducible factor-1 α, glucose transporter-1, (GLUT-1) and carbon anhydrase IX in endometrial cancer patients. Biomed Res Int. 2014; 2014:616850.
29 . Fujino M, Aishima S, Shindo K, Oda Y, Morimatsu K, Tsutsumi K, Otsuka T, Tanaka M, Oda Y. Expression of glucose transporter-1 is correlated with hypoxia-inducible factor 1α and malignant potential in pancreatic neuroendocrine tumors. Oncol Lett. 2016; 12:3337–43.
30 . Cho H, Lee YS, Kim J, Chung JY, Kim JH. Overexpression of glucose transporter-1 (GLUT-1) predicts poor prognosis in epithelial ovarian cancer. Cancer Invest. 2013; 31:607–15.
31 . Kim YH, Choi JY, Do IG, Kim S, Kim BT. Factors affecting 18F-FDG uptake by metastatic lymph nodes in gastric cancer. J Comput Assist Tomogr. 2013; 37:815–19.
32 . Nguyen XC, So Y, Chung JH, Lee WW, Park SY, Kim SE. High correlations between primary tumours and loco-regional metastatic lymph nodes in non-small-cell lung cancer with respect to glucose transporter type 1-mediated 2-deoxy-2-F18-fluoro-D-glucose uptake. Eur J Cancer. 2008; 44:692–98.
33 . Younes M, Juarez D, Lechago LV, Lerner SP. Glut 1 expression in transitional cell carcinoma of the urinary bladder is associated with poor patient survival. Anticancer Res. 2001; 21:575–78.
34 . Lee JH, Lee WA, Park SG, Park DK, Namgung H. Relationship Between Dual-Time Point FDG PET and Immunohistochemical Parameters in Preoperative Colorectal Cancer: preliminary Study. Nucl Med Mol Imaging. 2012; 46:48–56.
35 . Davis-Yadley AH, Abbott AM, Pimiento JM, Chen DT, Malafa MP. Increased Expression of the Glucose Transporter Type 1 Gene Is Associated With Worse Overall Survival in Resected Pancreatic Adenocarcinoma. Pancreas. 2016; 45:974–79.
36 . Kitasato Y, Yasunaga M, Okuda K, Kinoshita H, Tanaka H, Okabe Y, Kawahara A, Kage M, Kaida H, Ishibashi M. Maximum standardized uptake value on 18F-fluoro-2-deoxy-glucose positron emission tomography/computed tomography and glucose transporter-1 expression correlates with survival in invasive ductal carcinoma of the pancreas. Pancreas. 2014; 43:1060–65.
37 . Basturk O, Singh R, Kaygusuz E, Balci S, Dursun N, Culhaci N, Adsay NV. GLUT-1 expression in pancreatic neoplasia: implications in pathogenesis, diagnosis, and prognosis. Pancreas. 2011; 40:187–92.
38 . Jones RP, Bird NT, Smith RA, Palmer DH, Fenwick SW, Poston GJ, Malik HZ. Prognostic molecular markers in resected extrahepatic biliary tract cancers; a systematic review and meta-analysis of immunohistochemically detected biomarkers. Biomark Med. 2015; 9:763–75.
39 . Yin C, Nan Y, Lu T, Cheng Z, Cai Y. Clinicopathological Parameters Influence Assessment of FDG SPECT in Gastric Cancer. Hepatogastroenterology. 2015; 62:762–65.
40 . Kubo T, Shimose S, Fujimori J, Furuta T, Arihiro K, Ochi M. Does expression of glucose transporter protein-1 relate to prognosis and angiogenesis in osteosarcoma? Clin Orthop Relat Res. 2015; 473:305–10.
41 . van de Nes JA, Griewank KG, Schmid KW, Grabellus F. Immunocytochemical analysis of glucose transporter protein-1 (GLUT-1) in typical, brain invasive, atypical and anaplastic meningioma. Neuropathology. 2015; 35:24–36.
42 . Schreurs LM, Smit JK, Pavlov K, Pultrum BB, Pruim J, Groen H, Hollema H, Plukker JT. Prognostic impact of clinicopathological features and expression of biomarkers related to (18)F-FDG uptake in esophageal cancer. Ann Surg Oncol. 2014; 21:3751–57.
43 . Huang XQ, Chen X, Xie XX, Zhou Q, Li K, Li S, Shen LF, Su J. Co-expression of CD147 and GLUT-1 indicates radiation resistance and poor prognosis in cervical squamous cell carcinoma. Int J Clin Exp Pathol. 2014; 7:1651–66.
44 . Bredell MG, Ernst J, El-Kochairi I, Dahlem Y, Ikenberg K, Schumann DM. Current relevance of hypoxia in head and neck cancer. Oncotarget. 2016; 7:50781–804. https://doi.org/10.18632/oncotarget.9549.
45 . Lapel M, Weston P, Strassheim D, Karoor V, Burns N, Lyubchenko T, Paucek P, Stenmark KR, Gerasimovskaya EV. Glycolysis and oxidative phosphorylation are essential for purinergic receptor-mediated angiogenic responses in vasa vasorum endothelial cells. Am J Physiol Cell Physiol. 2017; 312:C56–70.
46 . Williams KJ, Telfer BA, Airley RE, Peters HP, Sheridan MR, van der Kogel AJ, Harris AL, Stratford IJ. A protective role for HIF-1 in response to redox manipulation and glucose deprivation: implications for tumorigenesis. Oncogene. 2002; 21:282–90.
47 . De Schutter H, Barbé B, Spaepen M, Begg AC, Balm A, Gregoire V, Haustermans K, Mahy P, Vander Poorten V, Nuyts S. Microsatellite alterations in head and neck squamous cell carcinoma and relation to expression of pimonidazole, CA IX and GLUT-1. Radiother Oncol. 2006; 80:143–50.
48 . Rajaganeshan R, Prasad R, Guillou PJ, Scott N, Poston G, Jayne DG. Expression patterns of hypoxic markers at the invasive margin of colorectal cancers and liver metastases. Eur J Surg Oncol. 2009; 35:1286–94.
49 . Meier V, Guscetti F, Roos M, Ohlerth S, Pruschy M, Rohrer Bley C. Hypoxia-Related Marker GLUT-1, CAIX, Proliferative Index and Microvessel Density in Canine Oral Malignant Neoplasia. PLoS One. 2016; 11:e0149993.
50 . Chen XH, Bao YY, Zhou SH, Wang QY, Wei Y, Fan J. Glucose transporter-1 expression in CD133+ laryngeal carcinoma Hep-2 cells. Mol Med Rep. 2013; 8:1695–700.
51 . Yan SX, Luo XM, Zhou SH, Bao YY, Fan J, Lu ZJ, Liao XB, Huang YP, Wu TT, Wang QY. Effect of antisense oligodeoxynucleotides glucose transporter-1 on enhancement of radiosensitivity of laryngeal carcinoma. Int J Med Sci. 2013; 10:1375–86.
52 . Bao YY, Zhou SH, Lu ZJ, Fan J, Huang YP. Inhibiting GLUT-1 expression and PI3K/Akt signaling using apigenin improves the radiosensitivity of laryngeal carcinoma in vivo. Oncol Rep. 2015; 34:1805–14.
53 . Luo XM, Xu B, Zhou ML, Bao YY, Zhou SH, Fan J, Lu ZJ. Co-Inhibition of GLUT-1 Expression and the PI3K/Akt Signaling Pathway to Enhance the Radiosensitivity of Laryngeal Carcinoma Xenografts In Vivo. PLoS One. 2015; 10:e0143306.
54 . Zhou S, Wang S, Wu Q, Fan J, Wang Q. Expression of glucose transporter-1 and -3 in the head and neck carcinoma—the correlation of the expression with the biological behaviors. ORL J Otorhinolaryngol Relat Spec. 2008; 70:189–94.
55 . Shen WQ, Cheng KJ, Bao YY, Zhou SH, Yao HT. Expression of Glut-1, HIF-1α, PI3K and p-Akt in a case of ceruminous adenoma. Head Neck Oncol. 2012; 4:18.
56 . Wang KR, Jiang T, Wu TT, Zhou SH, Yao HT, Wang QY, Lu ZJ. Expression of hypoxia-related markers in inflammatory myofibroblastic tumors of the head and neck. World J Surg Oncol. 2013; 11:294.
57 . Chai L, Ying HF, Wu TT, Zhou SH, Bao YY, Yao HT, You QH. Clinical features and hypoxic marker expression of primary sinonasal and laryngeal small-cell neuroendocrine carcinoma: a small case series. World J Surg Oncol. 2014; 12:199.
58 . Li SJ, Guo W, Ren GX, Huang G, Chen T, Song SL. Expression of Glut-1 in primary and recurrent head and neck squamous cell carcinomas, and compared with 2-[18F]fluoro-2-deoxy-D-glucose accumulation in positron emission tomography. Br J Oral Maxillofac Surg. 2008; 46:180–86.
59 . de Souza LB, de Oliveira LC, Nonaka CF, Lopes ML, Pinto LP, Queiroz LM. Immunoexpression of GLUT-1 and angiogenic index in pleomorphic adenomas, adenoid cystic carcinomas, and mucoepidermoid carcinomas of the salivary glands. Eur Arch Otorhinolaryngol. 2017; 274:2549–56.
60 . Mori Y, Tsukinoki K, Yasuda M, Miyazawa M, Kaneko A, Watanabe Y. Glucose transporter type 1 expression are associated with poor prognosis in patients with salivary gland tumors. Oral Oncol. 2007; 43:563–69.
61 . Bonfitto VL, Demasi AP, Costa AF, Bonfitto JF, Araujo VC, Altemani A. High-grade transformation of adenoid cystic carcinomas: a study of the expression of GLUT1 glucose transporter and of mitochondrial antigen. J Clin Pathol. 2010; 63:615–19.
62 . Demasi AP, Costa AF, Altemani A, Furuse C, Araújo NS, Araújo VC. Glucose transporter protein 1 expression in mucoepidermoid carcinoma of salivary gland: correlation with grade of malignancy. Int J Exp Pathol. 2010; 91:107–13.
63 . Fang J, Bao YY, Zhou SH, Fan J. Apigenin inhibits the proliferation of adenoid cystic carcinoma via suppression of glucose transporter-1. Mol Med Rep. 2015; 12:6461–66.
64 . Kim JW, Kwon GY, Roh JL, Choi SH, Nam SY, Kim SY, Cho KJ. Carcinoma ex pleomorphic adenoma of the salivary glands: distinct clinicopathologic features and immunoprofiles between subgroups according to cellular differentiation. J Korean Med Sci. 2011; 26:1277–85.
65 . Wu XH, Chen SP, Mao JY, Ji XX, Yao HT, Zhou SH. Expression and significance of hypoxia-inducible factor-1α and glucose transporter-1 in laryngeal carcinoma. Oncol Lett. 2013; 5:261–66.
66 . Starska K, Forma E, Jóźwiak P, Bryś M, Lewy-Trenda I, Brzezińska-Błaszczyk E, Krześlak A. Gene and protein expression of glucose transporter 1 and glucose transporter 3 in human laryngeal cancer-the relationship with regulatory hypoxia-inducible factor-1α expression, tumor invasiveness, and patient prognosis. Tumour Biol. 2015; 36:2309–21.
67 . Zuo J, Wen J, Lei M, Wen M, Li S, Lv X, Luo Z, Wen G. Hypoxia promotes the invasion and metastasis of laryngeal cancer cells via EMT. Med Oncol. 2016; 33:15.
68 . Kwon OJ, Park JJ, Ko GH, Seo JH, Jeong BK, Kang KM, Woo SH, Kim JP, Hwa JS, Carey TE. HIF-1α and CA-IX as predictors of locoregional control for determining the optimal treatment modality for early-stage laryngeal carcinoma. Head Neck. 2015; 37:505–10.
69 . Schrijvers ML, van der Laan BF, de Bock GH, Pattje WJ, Mastik MF, Menkema L, Langendijk JA, Kluin PM, Schuuring E, van der Wal JE. Overexpression of intrinsic hypoxia markers HIF1alpha and CA-IX predict for local recurrence in stage T1-T2 glottic laryngeal carcinoma treated with radiotherapy. Int J Radiat Oncol Biol Phys. 2008; 72:161–69.
70 . Zhao K, Yang SY, Zhou SH, Dong MJ, Bao YY, Yao HT. Fluorodeoxyglucose uptake in laryngeal carcinoma is associated with the expression of glucose transporter-1 and hypoxia-inducible-factor-1α and the phosphoinositide 3-kinase/protein kinase B pathway. Oncol Lett. 2014; 7:984–90.
71 . Bruine de Bruin L, Bollineni VR, Wachters JE, Schuuring E, van Hemel BM, van der Wal JE, Slagter-Menkema L, de Bock GH, Steenbakkers RJ, Langendijk JA, Pruim J, van der Laan BF, Halmos GB. Assessment of hypoxic subvolumes in laryngeal cancer with (18)F-fluoroazomycinarabinoside ((18)F-FAZA)-PET/CT scanning and immunohistochemistry. Radiother Oncol. 2015; 117:106–12.
72 . Shen LF, Zhao X, Zhou SH, Lu ZJ, Zhao K, Fan J, Zhou ML. In vivo evaluation of the effects of simultaneous inhibition of GLUT-1 and HIF-1α by antisense oligodeoxynucleotides on the radiosensitivity of laryngeal carcinoma using micro 18F-FDG PET/CT. Oncotarget. 2017; 8:34709–26 https://doi.org/10.18632/oncotarget.16671.
73 . Luo XM, Zhou SH, Fan J. Glucose transporter-1 as a new therapeutic target in laryngeal carcinoma. J Int Med Res. 2010; 38:1885–92.
74 . Zhou SH, Fan J, Chen XM, Cheng KJ, Wang SQ. Inhibition of cell proliferation and glucose uptake in human laryngeal carcinoma cells by antisense oligonucleotides against glucose transporter-1. Head Neck. 2009; 31:1624–33.
75 . Xu YY, Wu TT, Zhou SH, Bao YY, Wang QY, Fan J, Huang YP. Apigenin suppresses GLUT-1 and p-AKT expression to enhance the chemosensitivity to cisplatin of laryngeal carcinoma Hep-2 cells: an in vitro study. Int J Clin Exp Pathol. 2014; 7:3938–47.
76 . Azad N, Kumari Maurya M, Kar M, Goel MM, Singh AK, Sagar M, Mehrotra D, Kumar V. Expression of GLUT-1 in oral squamous cell carcinoma in tobacco and non-tobacco users. J Oral Biol Craniofac Res. 2016; 6:24–30.
77 . Pereira KM, Feitosa SG, Lima AT, Luna EC, Cavalcante RB, de Lima KC, Chaves FN, Costa FW. Immunohistochemical Evaluation of Glucose Transporter Type 1 in Epithelial Dysplasia and Oral Squamous Cell Carcinoma. Asian Pac J Cancer Prev. 2016; 17:147–51.
78 . Leite RB, Cavalcante RB, Nogueira RL, Souza LB, Pereira Pinto L, Nonaka CF. Analysis of GLUT-1, GLUT-3, and angiogenic index in syndromic and non-syndromic keratocystic odontogenic tumors. Braz Oral Res. 2017; 31:e34.
79 . Vasconcelos MG, Vasconcelos RG, Pereira de Oliveira DH, de Moura Santos E, Pinto LP, da Silveira ÉJ, Queiroz LM. Distribution of Hypoxia-Inducible Factor-1α and Glucose Transporter-1 in Human Tongue Cancers. J Oral Maxillofac Surg. 2015; 73:1753-60.
80 . Brands RC, Köhler O, Rauthe S, Hartmann S, Ebhardt H, Seher A, Linz C, Kübler AC, Müller-Richter UD. The prognostic value of GLUT-1 staining in the detection of malignant transformation in oral mucosa. Clin Oral Investig. 2017; 21:1631–37.
81 . Ohba S, Fujii H, Ito S, Fujimaki M, Matsumoto F, Furukawa M, Yokoyama J, Kusunoki T, Ikeda K, Hino O. Overexpression of GLUT-1 in the invasion front is associated with depth of oral squamous cell carcinoma and prognosis. J Oral Pathol Med. 2010; 39:74–78.
82 . Ayala FR, Rocha RM, Carvalho KC, Carvalho AL, da Cunha IW, Lourenço SV, Soares FA. GLUT1 and GLUT3 as potential prognostic markers for Oral Squamous Cell Carcinoma. Molecules. 2010; 15:2374–87.
83 . Grimm M, Munz A, Teriete P, Nadtotschi T, Reinert S. GLUT-1(+)/TKTL1(+) coexpression predicts poor outcome in oral squamous cell carcinoma. Oral Surg Oral Med Oral Pathol Oral Radiol. 2014; 117:743–53.
84 . Eckert AW, Lautner MH, Schütze A, Taubert H, Schubert J, Bilkenroth U. Coexpression of hypoxia-inducible factor-1α and glucose transporter-1 is associated with poor prognosis in oral squamous cell carcinoma patients. Histopathology. 2011; 58:1136–47.
85 . Samih N, Hovsepian S, Notel F, Prorok M, Zattara-Cannoni H, Mathieu S, Lombardo D, Fayet G, El-Battari A. The impact of N- and O-glycosylation on the functions of Glut-1 transporter in human thyroid anaplastic cells. Biochim Biophys Acta. 2003; 1621:92–101.
86 . Musholt TJ, Musholt PB, Dehdashti F, Moley JF. Evaluation of fluorodeoxyglucose-positron emission tomographic scanning and its association with glucose transporter expression in medullary thyroid carcinoma and pheochromocytoma: a clinical and molecular study. Surgery. 1997; 122:1049–60.
87 . Kaida H, Hiromatsu Y, Kurata S, Kawahara A, Hattori S, Taira T, Kobayashi M, Uchida M, Yamada K, Mihashi H, Umeno H, Kage M, Nakashima T, et al. Relationship between clinicopathological factors and fluorine-18-fluorodeoxyglucose uptake in patients with papillary thyroid cancer. Nucl Med Commun. 2011; 32:690–98.
88 . Wang YF, Liu B, Fan XS, Rao Q, Xu Y, Xia QY, Yu B, Shi SS, Zhou XJ. Thyroid carcinoma showing thymus-like elements: a clinicopathologic, immunohistochemical, ultrastructural, and molecular analysis. Am J Clin Pathol. 2015; 143:223–33.
89 . Kim MH, Ko SH, Bae JS, Lee SH, Jung CK, Lim DJ, Baek KH, Kim SH, Lee JM, Kang MI, Cha BY. Non-FDG-avid primary papillary thyroid carcinoma may not differ from FDG-avid papillary thyroid carcinoma. Thyroid. 2013; 23:1452–60.
90 . Chandan VS, Faquin WC, Wilbur DC, Khurana KK. The role of immunolocalization of CD57 and GLUT-1 in cell blocks in fine-needle aspiration diagnosis of papillary thyroid carcinoma. Cancer. 2006; 108:331–36.
91 . Kim S, Chung JK, Min HS, Kang JH, Park DJ, Jeong JM, Lee DS, Park SH, Cho BY, Lee S, Lee MC. Expression patterns of glucose transporter-1 gene and thyroid specific genes in human papillary thyroid carcinoma. Nucl Med Mol Imaging. 2014; 48:91–97.
92 . Lodewijk L, van Diest P, van der Groep P, Ter Hoeve N, Schepers A, Morreau J, Bonenkamp J, van Engen-van Grunsven A, Kruijff S, van Hemel B, Links T, Nieveen van Dijkum E, van Eeden S, et al. Expression of HIF-1α in medullary thyroid cancer identifies a subgroup with poor prognosis. Oncotarget. 2017; 8:28650–59. https://doi.org/10.18632/oncotarget.15622.
93 . Kim BH, Lee CH, Kim SJ, Jeon YK, Kim SS, Kim YK, Kim IJ. Clinicopathologic characteristics of synchronous primary thyroid cancer detected by initial staging 18F-FDG PET-CT examination in patients with underlying malignancy. Thyroid. 2013; 23:1431–36.
94 . Lee JW, Min HS, Lee SM, Kwon HW, Chung JK. Relations Between Pathological Markers and Radioiodine Scan and (18)F-FDG PET/CT Findings in Papillary Thyroid Cancer Patients With Recurrent Cervical Nodal Metastases. Nucl Med Mol Imaging. 2015; 49:127–34.
95 . Zhong JT, Zhou SH. Warburg effect, hexokinase-II, and radioresistance of laryngeal carcinoma. Oncotarget. 2017; 8:14133–46. https://doi.org/10.18632/oncotarget.13044.
96 . Pedersen PL. Voltage dependent anion channels (VDACs): a brief introduction with a focus on the outer mitochondrial compartment’s roles together with hexokinase-2 in the “Warburg effect” in cancer. J Bioenerg Biomembr. 2008; 40:123–26.
97 . Mathupala SP, Ko YH, Pedersen PL. Hexokinase-2 bound to mitochondria: cancer’s stygian link to the “Warburg Effect” and a pivotal target for effective therapy. Semin Cancer Biol. 2009; 19:17–24.
98 . Jin Z, Gu J, Xin X, Li Y, Wang H. Expression of hexokinase 2 in epithelial ovarian tumors and its clinical significance in serous ovarian cancer. Eur J Gynaecol Oncol. 2014; 35:519–24.
99 . Kwee SA, Hernandez B, Chan O, Wong L. Choline kinase alpha and hexokinase-2 protein expression in hepatocellular carcinoma: association with survival. PLoS One. 2012; 7:e46591.
100 . Guzman G, Chennuri R, Chan A, Rea B, Quintana A, Patel R, Xu PZ, Xie H, Hay N. Evidence for heightened hexokinase II immunoexpression in hepatocyte dysplasia and hepatocellular carcinoma. Dig Dis Sci. 2015; 60:420–26.
101 . Wolf A, Agnihotri S, Micallef J, Mukherjee J, Sabha N, Cairns R, Hawkins C, Guha A. Hexokinase 2 is a key mediator of aerobic glycolysis and promotes tumor growth in human glioblastoma multiforme. J Exp Med. 2011; 208:313–26.
102 . Robey RB, Hay N. Mitochondrial hexokinases, novel mediators of the antiapoptotic effects of growth factors and Akt. Oncogene. 2006; 25:4683–96.
103 . Zhang ZF, Feng XS, Chen H, Duan ZJ, Wang LX, Yang D, Liu PX, Zhang QP, Jin YL, Sun ZG, Liu H. Prognostic significance of synergistic hexokinase-2 and beta2-adrenergic receptor expression in human hepatocelluar carcinoma after curative resection. BMC Gastroenterol. 2016; 16:57.
104 . He X, Lin X, Cai M, Zheng X, Lian L, Fan D, Wu X, Lan P, Wang J. Overexpression of Hexokinase 1 as a poor prognosticator in human colorectal cancer. Tumour Biol. 2016; 37:3887–95.
105 . Gao Y, Xu D, Yu G, Liang J. Overexpression of metabolic markers HK1 and PKM2 contributes to lymphatic metastasis and adverse prognosis in Chinese gastric cancer. Int J Clin Exp Pathol. 2015; 8:9264–71.
106 . Gong L, Cui Z, Chen P, Han H, Peng J, Leng X. Reduced survival of patients with hepatocellular carcinoma expressing hexokinase II. Med Oncol. 2012; 29:909–14.
107 . Nakata B, Nishimura S, Ishikawa T, Ohira M, Nishino H, Kawabe J, Ochi H, Hirakawa K. Prognostic predictive value of 18F-fluorodeoxyglucose positron emission tomography for patients with pancreatic cancer. Int J Oncol. 2001; 19:53–58.
108 . Hamabe A, Yamamoto H, Konno M, Uemura M, Nishimura J, Hata T, Takemasa I, Mizushima T, Nishida N, Kawamoto K, Koseki J, Doki Y, Mori M, Ishii H. Combined evaluation of hexokinase 2 and phosphorylated pyruvate dehydrogenase-E1α in invasive front lesions of colorectal tumors predicts cancer metabolism and patient prognosis. Cancer Sci. 2014; 105:1100–08.
109 . Katagiri M, Karasawa H, Takagi K, Nakayama S, Yabuuchi S, Fujishima F, Naitoh T, Watanabe M, Suzuki T, Unno M, Sasano H. Hexokinase 2 in colorectal cancer: a potent prognostic factor associated with glycolysis, proliferation and migration. Histol Histopathol. 2017; 32:351–60.
110 . Anderson M, Marayati R, Moffitt R, Yeh JJ. Hexokinase 2 promotes tumor growth and metastasis by regulating lactate production in pancreatic cancer. Oncotarget. 2016; 8:56081–56094. https://doi.org/10.18632/oncotarget.9760.
111 . Wu CH, Lin YW, Wu TF, Ko JL, Wang PH. Clinical implication of voltage-dependent anion channel 1 in uterine cervical cancer and its action on cervical cancer cells. Oncotarget. 2016; 7:4210–25. https://doi.org/10.18632/oncotarget.6704.
112 . Coelho RG, Calaça IC, Celestrini DM, Correia-Carneiro AH, Costa MM, Zancan P, Sola-Penna M. Hexokinase and phosphofructokinase activity and intracellular distribution correlate with aggressiveness and invasiveness of human breast carcinoma. Oncotarget. 2015; 6:29375–87. https://doi.org/10.18632/oncotarget.4910.
113 . Li XB, Gu JD, Zhou QH. Review of aerobic glycolysis and its key enzymes - new targets for lung cancer therapy. Thorac Cancer. 2015; 6:17–24.
114 . Chen J, Zhang S, Li Y, Tang Z, Kong W. Hexokinase 2 overexpression promotes the proliferation and survival of laryngeal squamous cell carcinoma. Tumour Biol. 2014; 35:3743–53.
115 . Tian M, Zhang H, Higuchi T, Oriuchi N, Nakasone Y, Takata K, Nakajima N, Mogi K, Endo K. Hexokinase-II expression in untreated oral squamous cell carcinoma: comparison with FDG PET imaging. Ann Nucl Med. 2005; 19:335–38.
116 . Xiao L, Hu ZY, Dong X, Tan Z, Li W, Tang M, Chen L, Yang L, Tao Y, Jiang Y, Li J, Yi B, Li B, et al. Targeting Epstein-Barr virus oncoprotein LMP1-mediated glycolysis sensitizes nasopharyngeal carcinoma to radiation therapy. Oncogene. 2014; 33:4568–78.
117 . Cho MH, Park CK, Park M, Kim WK, Cho A, Kim H. Clinicopathologic Features and Molecular Characteristics of Glucose Metabolism Contributing to 18F-fluorodeoxyglucose Uptake in Gastrointestinal Stromal Tumors. PLoS One. 2015; 10:e0141413.
118 . Yang HJ, Xu WJ, Guan YH, Zhang HW, Ding WQ, Rong L, Qiu ZB, Zhong L. Expression of Glut-1 and HK-II in Pancreatic Cancer and Their Impact on Prognosis and FDG Accumulation. Transl Oncol. 2016; 9:583–91.
119 . Grimm M, Cetindis M, Lehmann M, Biegner T, Munz A, Teriete P, Kraut W, Reinert S. Association of cancer metabolism-related proteins with oral carcinogenesis - indications for chemoprevention and metabolic sensitizing of oral squamous cell carcinoma? J Transl Med. 2014; 12:208.
120 . Paudyal B, Oriuchi N, Paudyal P, Higuchi T, Nakajima T, Endo K. Expression of glucose transporters and hexokinase II in cholangiocellular carcinoma compared using [18F]-2-fluro-2-deoxy-D-glucose positron emission tomography. Cancer Sci. 2008; 99:260–66.
121 . Kristian A, Nilsen LB, Røe K, Revheim ME, Engebråten O, Mælandsmo GM, Holm R, Malinen E, Seierstad T. Dynamic (18) F-FDG PET for Assessment of Tumor Physiology in Two Breast Carcinoma Xenografts. Nucl Med Mol Imaging. 2013; 47:173–80.
122 . Izuishi K, Yamamoto Y, Sano T, Takebayashi R, Nishiyama Y, Mori H, Masaki T, Morishita A, Suzuki Y. Molecular mechanism underlying the detection of colorectal cancer by 18F-2-fluoro-2-deoxy-D-glucose positron emission tomography. J Gastrointest Surg. 2012; 16:394–400.
123 . Higashi T, Saga T, Nakamoto Y, Ishimori T, Mamede MH, Wada M, Doi R, Hosotani R, Imamura M, Konishi J. Relationship between retention index in dual-phase (18)F-FDG PET, and hexokinase-II and glucose transporter-1 expression in pancreatic cancer. J Nucl Med. 2002; 43:173–80.
124 . Baschnagel AM, Wobb JL, Dilworth JT, Williams L, Eskandari M, Wu D, Pruetz BL, Wilson GD. The association of (18)F-FDG PET and glucose metabolism biomarkers GLUT1 and HK2 in p16 positive and negative head and neck squamous cell carcinomas. Radiother Oncol. 2015; 117:118–24.
125 . Zhou X, Chen R, Xie W, Ni Y, Liu J, Huang G. Relationship between 18F-FDG accumulation and lactate dehydrogenase A expression in lung adenocarcinomas. J Nucl Med. 2014; 55:1766–71.
126 . Park SG, Lee JH, Lee WA, Han KM. Biologic correlation between glucose transporters, hexokinase-II, Ki-67 and FDG uptake in malignant melanoma. Nucl Med Biol. 2012; 39:1167–72.
127 . Alvarez JV, Belka GK, Pan TC, Chen CC, Blankemeyer E, Alavi A, Karp JS, Chodosh LA. Oncogene pathway activation in mammary tumors dictates FDG-PET uptake. Cancer Res. 2014; 74:7583–98.
128 . Takebayashi R, Izuishi K, Yamamoto Y, Kameyama R, Mori H, Masaki T, Suzuki Y. [18F]Fluorodeoxyglucose accumulation as a biological marker of hypoxic status but not glucose transport ability in gastric cancer. J Exp Clin Cancer Res. 2013; 32:34.
129 . de Andrade Barreto E, de Souza Santos PT, Bergmann A, de Oliveira IM, Wernersbach Pinto L, Blanco T, Rossini A, Pinto Kruel CD, Mattos Albano R, Ribeiro Pinto LF. Alterations in glucose metabolism proteins responsible for the Warburg effect in esophageal squamous cell carcinoma. Exp Mol Pathol. 2016; 101:66–73.
130 . Massari F, Ciccarese C, Santoni M, Iacovelli R, Mazzucchelli R, Piva F, Scarpelli M, Berardi R, Tortora G, Lopez-Beltran A, Cheng L, Montironi R. Metabolic phenotype of bladder cancer. Cancer Treat Rev. 2016; 45:46–57.
131 . Iwamoto M, Kawada K, Nakamoto Y, Itatani Y, Inamoto S, Toda K, Kimura H, Sasazuki T, Shirasawa S, Okuyama H, Inoue M, Hasegawa S, Togashi K, Sakai Y. Regulation of 18F-FDG accumulation in colorectal cancer cells with mutated KRAS. J Nucl Med. 2014; 55:2038–44.
132 . Song K, Li M, Xu X, Xuan LI, Huang G, Liu Q. Resistance to chemotherapy is associated with altered glucose metabolism in acute myeloid leukemia. Oncol Lett. 2016; 12:334–42.
133 . Yamada T, Uchida M, Kwang-Lee K, Kitamura N, Yoshimura T, Sasabe E, Yamamoto T. Correlation of metabolism/hypoxia markers and fluorodeoxyglucose uptake in oral squamous cell carcinomas. Oral Surg Oral Med Oral Pathol Oral Radiol. 2012; 113:464–71.
134 . Kim HM, Koo JS. Differential Expression of Glycolysis-Related Proteins in Follicular Neoplasms versus Hürthle Cell Neoplasms: A Retrospective Analysis. Dis Markers. 2017; 2017:6230294.
135 . Zhang TB, Zhao Y, Tong ZX, Guan YF. Inhibition of glucose-transporter 1 (GLUT-1) expression reversed Warburg effect in gastric cancer cell MKN45. Int J Clin Exp Med. 2015; 8:2423–28.
136 . Jian F, Yuan F, Jiong M, Zhu XZ, Yu GR, Lu DD. Silencing of Glucose Transporter Protein-1 by RNA Interference Inhibits Human Osteosarcoma Mg63 Cells Growth in vivo. Technol Cancer Res Treat. 2015; 14:243–48.
137 . Shimanishi M, Ogi K, Sogabe Y, Kaneko T, Dehari H, Miyazaki A, Hiratsuka H. Silencing of GLUT-1 inhibits sensitization of oral cancer cells to cisplatin during hypoxia. J Oral Pathol Med. 2013; 42:382–88.
138 . Min JW, Kim KI, Kim HA, Kim EK, Noh WC, Jeon HB, Cho DH, Oh JS, Park IC, Hwang SG, Kim JS. INPP4B-mediated tumor resistance is associated with modulation of glucose metabolism via hexokinase 2 regulation in laryngeal cancer cells. Biochem Biophys Res Commun. 2013; 440:137–42.
139 . Lu CL, Qin L, Liu HC, Candas D, Fan M, Li JJ. Tumor cells switch to mitochondrial oxidative phosphorylation under radiation via mTOR-mediated hexokinase II inhibition—a Warburg-reversing effect. PLoS One. 2015; 10:e0121046.
140 . Fabian C, Koetz L, Favaro E, Indraccolo S, Mueller-Klieser W, Sattler UG. Protein profiles in human ovarian cancer cell lines correspond to their metabolic activity and to metabolic profiles of respective tumor xenografts. FEBS J. 2012; 279:882–91.
141 . Gupta P, Jagavelu K, Mishra DP. Inhibition of NADPH Oxidase-4 Potentiates 2-Deoxy-D-Glucose-Induced Suppression of Glycolysis, Migration, and Invasion in Glioblastoma Cells: Role of the Akt/HIF1α/HK-2 Signaling Axis. Antioxid Redox Signal. 2015; 23:665–81.
142 . Cervantes-Madrid D, Dueñas-González A. Antitumor effects of a drug combination targeting glycolysis, glutaminolysis and de novo synthesis of fatty acids. Oncol Rep. 2015; 34:1533–42.
143 . Gong L, Wei Y, Yu X, Peng J, Leng X. 3-Bromopyruvic acid, a hexokinase II inhibitor, is an effective antitumor agent on the hepatoma cells : in vitro and in vivo findings. Anticancer Agents Med Chem. 2014; 14:771–76.
144 . Krasnov GS, Dmitriev AA, Lakunina VA, Kirpiy AA, Kudryavtseva AV. Targeting VDAC-bound hexokinase II: a promising approach for concomitant anti-cancer therapy. Expert Opin Ther Targets. 2013; 17:1221–33.
145 . Xi F, Ye J. Inhibition of Lung Carcinoma A549 Cell Growth by Knockdown of Hexokinase 2 In Situ and In Vivo. Oncol Res. 2016; 23:53–59.