
INTRODUCTION
Ovarian cancer (OC) is the most lethal gynecologic malignancy [1]. In Japan, ovarian clear cell carcinoma (OCCC) is the second most common histologic subtype of ovarian cancer [2]. S everal reports have described an increase prevalence of OCCC in Japan compared with that in other countries [2, 3]; however, the reason for this is unknown. Previously, various studies have demonstrated an association between OCCC or ovarian endometrioid carcinoma (OEC) and endometriosis, and it is widely recognized that gene alterations associated with carcinogenesis occur in endometriosis. Common molecular genetic alterations in endometriosis-related ovarian neoplasms (ERONs) have already described [4–12], [15–18].
Recently, several reports have suggested that numerous genetic alterations are associated with carcinogenesis, leading from endometriosis to ovarian cancer. A KRAS mutation has been detected in OEC tissue, but not in atypical endometriosis bordering the cancerous region [8]. Common molecular genetic alterations in ERON, such as PTEN deletion and microsatellite instability, may also be detected in normal-appearing epithelial cells of endometriotic cysts [7, 9]. Among all molecular genetic changes identified to date, inactivating mutations of the ARID1A tumor suppressor gene are the most common in ERON. The role of ARID1A alterations in the early molecular pathogenesis of OCCC is demonstrated [10, 11]. Furthermore, it was identified that atypical endometriosis and OCCC share molecular alterations, such as inactivating mutations for ARID1A, activating mutations for PIK3CA, and the hypomethylation of HNF1 homeobox B (HNF1B) [12].
Thus, molecular biological analyses of ERONs have identified several molecular genetic alterations in ARID1A, PTEN, PIK3CA, KRAS, as well as in other genes. Additionally, numerous reports describe the relationship of carcinogenesis from endometriosis to OEC or OCCC; however, much less is known about the pathogenetic pathways and the order in which alterations occur. Moreover, the correct frequency of gene alterations could not be determined as the DNA used in these studies was purified by several different methods.
Recently, a novel treatment strategy, in which inhibition of enhancer of zeste homology 2 (EZH2) activity electively suppressed the growth of ARID1A-mutated OCCC cells, has been reported [13]. EZH2 is the functional enzymatic component of the Polycomb Repressive Complex 2 (PRC2), which is responsible for healthy embryonic development through the epigenetic maintenance of genes responsible for regulating development and differentiation [14].
GSK126, a highly specific EZH2 inhibitor, caused the regression of established ARID1A-mutated OCCC and decreased the number of disseminated tumor nodules in xenograft models [13]. Thus, GSK126exhibits potential as a molecular targeted drug that inhibits the proliferation of ARID1A-mutated ovarian clear cell carcinoma cells by targeting and inhibiting EZH2. The clinical application of drugs that target and inhibit EZH2 require further examination of the status of ARID1A mutations in OCCC. ARID1A is a large gene that contains 20 exons, and mutations are distributed evenly across the whole gene [15, 16]. It has already been shown in previous studies that the ARID1A mutation frequency in patients with OCCC and OEC hadis about 30-60% [15, 16]. For the utilization of EZH2 to the advantage of patients, the correct frequency of ARID1A mutation must be considered. Here, we attempted to show that the frequency of the ARID1A mutation is higher than that previously reported, using pure DNA purified from tumor epithelial cells, using a method we termed ‘liquid microdissection’.
Polymerase epsilon (POLE), which has recently become the focus for endometrial carcinoma, is a DNA polymerase with a proofreading exonuclease domain. It is responsible for the recognition and excision of mispaired bases, thereby allowing high-fidelity DNA replication. The Cancer Genome Atlas (TCGA) research network recently identified an ultra-mutated group of endometrial carcinomas characterized by mutations in POLE and exceptionally high substitution rates [17]. Previous reports have demonstrated the prevalence and patterns of POLE mutations in OEC [18]; however, these have not been studied in detail in other histological types. In comparison, relatively little known is known about the POLE mutation status of ERONs.
However, molecular analyses remain challenging: it is difficult to obtain DNA from pure tumor cells and to avoid contamination from fibroblasts. As such, no reports have utilized purified tumor samples to determine sequence mutations in ERONs. The analysis of such genetic alterations is typically complicated by contamination of fibroblast DNA. Thus, the quality of DNA products and the sensitivity of detection techniques for analyzing ovarian carcinomas must be improved. In this study, we purified DNA from only tumor epithelial cells after removing fibroblasts from seeded primary cultured cells [19]. We called this method “liquid microdissection.” Using this technique, we investigated the carcinogenesis of OEC and OCCC using high-quality purified DNA.
RESULTS
Clinical and pathological features
The clinical and histological features of nine clear cell and 13 endometrioid ovarian carcinomas are described (Table 1). Patient age ranged from 47 to 80 years of age with an average of 58.0 years, and all patients were post-menopausal. Fourteen patients had FIGO (2009) stage 1 disease, one patient had stage 2 disease, four patients had stage 3 disease, and three had stage 4 disease.
Table 1: Clinical characteristics of patients with ERONs
Case no. |
Histology |
Age (years) |
FIGO Stage |
Residual tumor |
Recurrence |
ARID1A |
p53 |
POLE |
PTEN |
PIK3CA |
KRAS |
|---|---|---|---|---|---|---|---|---|---|---|---|
1 |
E |
58 |
IIIC |
N |
Y |
M |
WT |
M |
WT |
WT |
NA |
2 |
E |
57 |
IIIC |
Y |
Y |
M |
WT |
WT |
WT |
WT |
NA |
3 |
E |
72 |
IIIC |
N |
Y |
M |
M |
M |
M |
WT |
WT |
4 |
E |
60 |
IV |
Y |
Y |
M |
M |
M |
NA |
WT |
WT |
5 |
E |
80 |
IA |
N |
Y |
WT |
WT |
WT |
NA |
WT |
WT |
6 |
E |
59 |
IC |
N |
N |
M |
WT |
WT |
M |
M |
WT |
7 |
E |
58 |
IA |
N |
N |
M |
WT |
WT |
M |
M |
WT |
8 |
E |
61 |
IIC |
N |
N |
M |
WT |
M |
M |
WT |
NA |
9 |
E |
50 |
IC |
N |
N |
M |
M |
WT |
M |
WT |
M |
10 |
E |
47 |
IC |
N |
N |
M |
WT |
M |
WT |
WT |
NA |
11 |
E |
77 |
IC |
N |
N |
M |
M |
M |
M |
WT |
W |
12 |
E |
58 |
IC |
N |
N |
M |
WT |
WT |
M |
WT |
NA |
13 |
E |
76 |
IC |
N |
N |
M |
M |
WT |
NA |
WT |
NA |
14 |
C |
56 |
IV |
Y |
Y |
M |
NA |
M |
WT |
NA |
NA |
15 |
C |
63 |
IC |
N |
Y |
M |
WT |
WT |
M |
WT |
NA |
16 |
C |
55 |
IIIC |
Y |
Y |
M |
M |
M |
M |
M |
WT |
17 |
C |
50 |
IC |
N |
N |
NA |
NA |
WT |
WT |
NA |
NA |
18 |
C |
62 |
IC |
N |
Y |
M |
WT |
M |
M |
WT |
NA |
19 |
C |
61 |
IV |
Y |
Y |
M |
M |
WT |
M |
WT |
WT |
20 |
C |
50 |
IC |
N |
N |
NA |
NA |
WT |
WT |
NA |
NA |
21 |
C |
71 |
IC |
N |
N |
M |
WT |
WT |
M |
WT |
WT |
22 |
C |
55 |
IC |
N |
Y |
M |
WT |
WT |
M |
WT |
NA |
Mutation frequency % |
95 |
36.8 |
40.9 |
68.4 |
15.8 |
11.1 |
|||||
Previous Reports % |
E |
9(20)(21) |
20(18) |
8(7)(9) |
20(22)(23) |
10(24)(25) |
|||||
C |
25(20)(21) |
8(18) |
33-46(22)(23) |
||||||||
NA: Not available.
WT: Wild type.
M: Mutation.
E: endometroid carcinoma.
C: clear cell carcinoma.
Identification of ARID1A, p53, POLE, PTEN, PIK3CA, and KRAS mutations
The mutational status of ARID1A, p53, POLE, PTEN, PIK3CA, and KRAS in all 22 purified ovarian tumors is summarized in Table 1. The frequency of mutations of these genes were described comparing previous reports [7] [9] [15, 16] [18] [20–25]. Somatic mutations in ARID1A were identified in 19 (95.0 %) of 20 ERONs. Somatic mutations in POLE were identified in nine (40.9 %) of 22 ERONs. The mutations in ARID1A and POLE, and the fact that they were somatic, were confirmed by Sanger sequencing of DNA of tumor and normal tissues from the corresponding patients (Supplementary Figure 1). The frequency of ARID1A mutations was higher than that in previous reports, especially for clear cell carcinoma. These mutational types are described in Supplementary Table 2.
Clinical features of ERONs with POLE mutations
Univariate analysis of clinicopathologic factors showed that POLE mutations were only related to FIGO stage (Table 2). We then investigated the statistical correlation between mutations in these genes and POLE mutations (Table 3). Among these genes, there were no significant correlations regarding the mutation status.
Table 2: Association between POLE mutation and clinicopathological factors in patients with ERONs
Factors |
Patiens |
POLE mutation |
P |
|
|---|---|---|---|---|
Negative |
Positive |
|||
FIGO stage |
||||
I, II |
13 |
11 |
4 |
0.047 |
III, IV |
9 |
2 |
5 |
|
Age (y) |
||||
<60 |
12 |
8 |
4 |
0.429 |
≧60 |
10 |
5 |
5 |
|
Residual tumor |
||||
Negative |
17 |
11 |
6 |
0.323 |
Positive |
5 |
2 |
3 |
|
Recurrence |
||||
Yes |
11 |
5 |
6 |
0.193 |
No |
11 |
8 |
3 |
|
Secondline treatment sensitivity |
||||
Yes |
3 |
1 |
3 |
0.303 |
No |
8 |
4 |
3 |
|
Dead of disease |
||||
Yes |
8 |
5 |
3 |
0.806 |
No |
14 |
8 |
6 |
|
Table 3: Association between POLE mutation and other gene mutations in patients with ERONs
Factors |
Patiens |
POLE mutation |
P |
|
|---|---|---|---|---|
Negative |
Positive |
|||
ARID1A |
||||
Negative |
1 |
1 |
0 |
0.353 |
positive |
19 |
10 |
9 |
|
p53 |
||||
Negative |
12 |
8 |
4 |
0.311 |
positive |
7 |
3 |
4 |
|
PTEN |
||||
Negative |
6 |
3 |
3 |
0.636 |
positive |
13 |
8 |
5 |
|
PIK3CA |
||||
Negative |
16 |
9 |
7 |
0.737 |
positive |
3 |
2 |
1 |
|
KRAS |
||||
Negative |
9 |
5 |
4 |
0.389 |
positive |
1 |
1 |
0 |
|
Finally, Kaplan-Meier analyses were performed to determine potential correlations between POLE mutations and patient prognosis. However, Kaplan-Meier analysis similarly did not show a significant difference for PFS and OS (P = 0.938; log-rank test; P = 0.391, log-rank test; Figure 1).
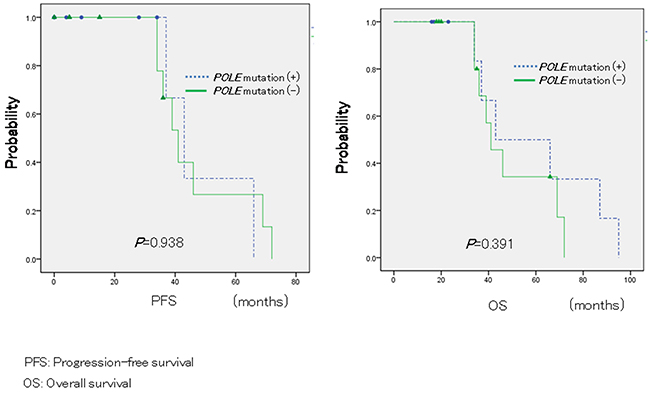
Figure 1: Relationship between POLE mutations and patient prognosis in ovarian carcinomas. In 22 patients with endometrioid and clear cell carcinoma, Kaplan-Meier survival analysis was performed, showing that POLE mutation positivity (blue line; n = 9) and negativity (green line; n = 11) are not associated with progression-free survival and overall survival (P = 0.938, log-rank test; P = 0.391, log-rank test).
DISCUSSION
The present study delivers three major findings. First, we showed that the prevalence of ARID1A mutations in ERONs is 95.0% (19/20). Previous reports showed rates of 30% in OEC and 46–57% in OCCC [15, 16]. At the onset of this study, we were concerned about complications due to the contamination of fibroblasts that were present with tumor epithelial cells in primary cultures from tumor tissue. In studies that examine gene alterations in various tumors, it is important to ensure robust DNA quality. In this study, DNA was purified from only tumor epithelial cells, which were isolated using a cell sorting system. Our results showed that DNA purified by “liquid microdissection” from tumor tissue could be used to identify the exact prevalence of ARID1A mutations, which was found to be higher than that stated in previous reports. We used a small sample in this study; however, it should be emphasized that this limitation was overcome by using a strategy for the purification of high-quality DNA from tissue culture sorting without contamination of fibroblasts, which makes the present findings more robust.
ERONs are the most important ovarian tumors in Japanese patients, based on the enhanced prevalence in this population [7, 26]. In Japan, a study showed that endometriosis-related ovarian carcinoma (ERON) is not rare, with an estimated incidence of 0.72% [27]. The carcinogenesis of ERONs from the endometriosis stage onwards was unclear; however, recently, several reports have demonstrated the carcinogenesis of ERONs. In a model of genetically engineered mice harboring an oncogenic allele of KRAS resulting in benign lesions reminiscent of endometriosis, a deletion of PTEN caused progression towards the OEC, but not the OCCC [28]. The role of ARID1A alterations in early molecular pathogenesis of CCC has been demonstrated in two recent reports [10, 11].
Regarding the carcinogenesis of ERONs, it is not clear which step is essential for the initiation or progression from benign tumor to cancer. The actual frequencies of certain genetic alterations have also not beendetermined. As ARID1A mutations are detected in endometrioma, it has been suggested that they are involved in the onset and progression of cancer [29]. Thus, based on the observed high frequency of ARID1A mutations, we suggest that such alterations form the basis of carcinogenesis for ERONs, and that other gene mutations might be gained subsequently to facilitate the progression of carcinogenesis.
The identification of every association between carcinogenesis and gene alteration of ERONs was impossible in the present study alone. There are some limitations to this study: it is difficult to generalize whether a high mutation frequency leads to carcinogenesis. In this study, a large number of mutations were detected in ARID1A. With regard to this, there are various types of mutations, which may be either pathogenic or of uncertain importance. For example, variants of uncertain significance (VUS) were reported amongBRCA1/2 gene alterations upon examining a HBOC family cohort [30]. The identification of the specific alteration that leads to pathogenic change requires further investigation. The identification of pathogenic VUS requires the accumulation of large amounts of VUS data.
Another limitation is that there are numerous gene alterations that result not only from gene mutations, but are also affected by micro RNA, gene methylation, and so on. In this study, we only performed mutational analysis by Sanger sequencing, which may be insufficient to identify epigenetic changesassociated with carcinogenesis. Further studies will be needed to be established the association between carcinogenesis and epigenetic changes in this context.
Second, we evaluated the clinicopathological and prognostic significance of POLE mutations in ERONs. POLE is a DNA polymerase with a proofreading exonuclease domain, and is responsible for the detection and excision of mispaired bases, thus facilitating high-accuracy DNA replication [31]. In the TCGA series, POLE mutations were found in 3% of colorectal cancers [32] and 7% of endometrial cancers (ECs). Improved PFS in patients with POLE ultra-mutated EC has been reported [17]. Recently, Hoang et al [18] described that POLE mutations are found in 6% of low-grade and 17% of high-grade endometrioid carcinomas in OC. However, little is known about the POLE mutation status in ERONs.
In the present study, there was no significant relationship between POLE mutations and PFS or OS. We first reported the relationship between POLE mutations and patient prognosis in ERONs in the current study. Prior to this study, we hypothesized that patients with POLE mutations how good prognosis. In this study, some patients with POLE mutations exhibited good response to second-line chemotherapy with disease recurrence (date not shown). However, because of the small sample size, we did not have sufficient statistical power to determine its prognostic significance for ERONs. In the future, studies with a greater number of cases will be needed to determine the prognostic significance of POLE mutations.
There was another limitation regarding the mutation analysis of POLE. We could not confirm which mutations are responsible for the pathogenesis of the disease. In addition, hot spot mutations represent a possible reason for the difference in results between our study and TCGA data. Zou et al [33] recently reported the same missense mutations in POLE at p.S297F in OEC in a Chinese population. They were also unable to identify any hot spot mutations that are frequently identified in endometrioid EC based on TCGA data [17]. Mutation hot spots might be different between races or types of cancer. The results are also difficult to interpret based on the fact that mutations other than hot spot mutations have not been described in detail. Whether the mutation is significant could be defined by functional or configuration analyses. If the role of several mutations could be precisely proven, it may be possible to better infer correlations between POLE mutations and patient prognosis.
Recently, Church et al [34, 35] reported POLE-ultra-mutated and MSI groups serve as biomarkers of the blockade of immune checkpoints in cancer immunotherapy. These two groups are characterized by an active immune microenvironment. The interaction between programmed death 1 (PD-1) receptor and programmed death ligand 1 (PD-L1) is an important pathway for inhibiting the immune checkpoint system. These pathways are already accepted as a target for melanoma and lung cancer [36, 37]. Hamanishi et al [38] also reported that the anti-PD-1 antibody nivolumab has positive effects on recurrent ovarian cancer, especially for clear cell carcinoma. Therein, a patient with multiple peritoneal dissemination exhibited complete response without recurrence after treatment with nivolumab. We expect that an anti-PD-1 antibody might show efficacy in patients with ovarian carcinoma such as clear cell carcinoma from ERONs. The mechanism associated with the effect of the anti-PD-1 antibody was also previously demonstrated [38].
Previous studies noted that POLE-mutated ECs are typically associated with strong lymphocyte infiltration [34, 35, 39]; thus, patients with POLE-ultra mutated and MSI ECs may receive the maximum benefit from drugs such as immuno-checkpoint inhibitors [40]. Overexpression of PD-1/PD-L1 or loss of MMR proteins might be associated with response to checkpoint blockade immunotherapies; thus, these factors potentially represent useful biomarkers.
After the initial TCGA report, multiple studies have demonstrated that POLE-ultra mutated ECs are characterized by excellent prognosis, despite the high histological grade; however, the reasons for such favorable outcomes are not completely understood [35, 39, 41, 42]. In the future, research on biomarkers that predict response to checkpoint blockade immunotherapies could provide the main benefit in terms of durable responses or survival. If POLE-mutated ERONs also have excellent prognosis, this knowledge will likely affect the course of immunotherapy for these patients, and perhaps lead to investigation into treatment using immuno-checkpoint inhibitors for ERONs with disease recurrence. Overexpression of PD-1/PD-L1 or loss of MMR proteins might be associated with the response to checkpoint blockade immunotherapies; thus, these markers would be useful biomarkers. We aim to conduct a further examination of the association between checkpoint blockade immunotherapies and POLE mutations.
Finally, in the present study, somatic mutations in both ARID1A and p53 were identified in four (40.0%) of 10 endometrioid carcinomas and in two (33.3%) of six clear cell carcinomas. A significant correlation between both ARID1A and p53 mutations and PFS was described for OCCC (p = 0.025, log-rank test); however, this was not described for OEC (p = 0.687, log-rank test) (data not shown). In previous reports, ARID1A was shown to be mutually exclusive of p53 [43, 44]; however, in recent reports, ARID1A mutations and p53 mutations have been described in the same case [45]. The explanation for this has been unclear; however, their gene products have been shown to form a complex that regulates the transcription of CDKN1A and SMAD4 [43]. This suggests that mutations in these genes may be sufficient to promote carcinogenesis through a common pathway.
In summary, the frequency of ARID1A mutations in ERONs may be higher than that reported previously. The “liquid microdissection” method, which was performed for the purification of DNA, yields high-quality sequencing results. ARIDA mutations may be the basis for carcinogenesis in ERONs; we speculate that other gene mutations are gained subsequently, thus facilitating the progression of carcinogenesis.
MATERIALS AND METHODS
Tissue samples and tumor cell isolation: cell sorting
Tissue samples were obtained from the Department of Obstetrics and Gynecology at Shimane University School of Medicine between 2008 and 2013. The acquisition of tumor tissues was approved by the Shimane University Institutional Review Board. The diagnoses were confirmed by a surgical pathologist before the tumor samples were harvested for experiments. All patients had endometriotic lesions in the abdominal cavity or had episodes of endometriosis. There were three patients in stage 4. Metastatic sites were not present invital organs such as liver and lung. Patients exhibited only lymphnode metastasis at distant lesions such as the mediastinum lymphnode or carcinomatous pleural effusion. All patients were primarily treated with cytoreductive surgery and adjuvant platinum and taxane chemotherapy (carboplatin AUC5, paclitaxel 175 mg/m2 or docetaxel 70 mg/m2) or platinum and topoisomerase inhibitor chemotherapy (cisplatin 60 mg/m2, irinotecan 60 mg/m2). All patients received 6–12 courses of this regimen. When patients had disease recurrence, the second line chemotherapy regimen comprised PLD and CBDCA for platinum-sensitive patients and GEM, PLD, TPT alone, and bevacizumab for platinum-resistant patients, For sequencing analysis, tumor cells from 13 endometrioid carcinomas and nine clear cell carcinomas were isolated using the following protocol, which is illustrated in Figure 2.
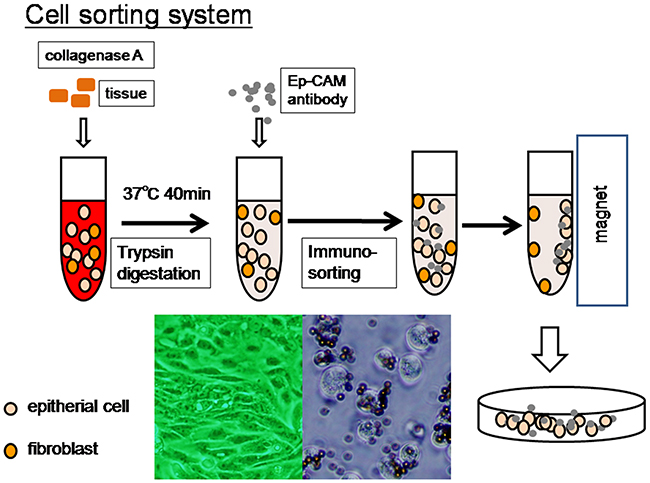
Figure 2: Overview of liquid microdissection technique. Fresh tumor tissues were minced to 2-3-mm fragments and liquefied with collagenase A (10 mg/ml) with mild rolling at 37°C for 40 min. Tumor cells were isolated, after the large tissue fragments sunk to the bottom of the tube, using magnetic beads coated with an Ep-CAM antibody. Tumor cells were directly harvested for genomic DNA isolation or cultured for 3 days to confirm that these cells maintained their epithelial characteristics.
The method used for cell sorting has been described in detail in our previous report [19]. Briefly, primary tumor cultures were derived from freshly isolated tumor samples by immune-sorting. Fresh tumor tissues were minced to 2-3-mm fragments and liquefied with collagenase A (10 mg/ml), with mild rolling at 37°C for 40 min. Small tumor cell clusters (< 10 cells) were gathered from the top part of centrifuge tubes after the large tissue fragments sank to the bottom of the tube. Tumor cells were isolated using magnetic beads coated with an Ep-CAM-specific antibody (Dynal, Oslo, Norway), anddirectly harvested for genomic DNA isolation. A small number of tumor cells were cultured for three days; we confirmed that these cells maintained the characteristics of epithelial cells.
Mutational analysis in ovarian cancers by Sanger sequencing
Nucleotide sequencing was used to analyze the mutational status of ARID1A, p53, POLE, PTEN, PIK3CA, and KRAS in tumor cells isolated from ovarian carcinomas. In this study, we focused on analyzing exons that have been reported to harbor the majority of mutations for each of the genes. The primer sequences and the PCR protocol have been described in our previous reports [15, 19].
DNA was extracted and amplified by polymerase chain reaction with primers for exon 2 of KRAS, exons 1–9 of PTEN, exons 9 and 20 of PIK3CA, exons 1–9 of p53, exons 9–14 of POLE, and the whole exome sequence of ARID1A. Gene mutations were analyzed using primers described in Supplementary Table 1.
All mutations identified in the tumors were confirmed by independent PCR and Sanger sequencing in the specific tumors and their paired normal tissue to determine their somatic nature. Sequencing was performed using the ABI BigDye Terminator v3.1 Cycle Sequencing kit (Applied Biosystems, Thermo Fisher K. K Yokohama Japan). We also performed sequencing analysis to detect mutations in benign tissue such as blood from each patient with cancer. We determined whether the same mutations were present in both tumor and benign tissue to ascertain germ line mutations.
In the current study, we did not use NGS as the aim was to determine whether the frequency of mutations was altered based on the quality of DNA; therefore, it was not necessary to attempt to identify new hot spots and mutation sites.
Statistical analysis of clinicopathological correlations
Statistical analyses were conducted using the Statistical Package for the Social Sciences for Windows software, Version 19.0 (IBM Corp., Armonk, NY, USA). All reported P-values were two-sided and P-values below 0.05 were considered to represent statistical significance.
Analysis of POLE and other gene mutations or alteration frequencies with clinicopathological parameters was performed using the chi-squared test.
Overall survival (OS) was defined as time from surgery to death from any cause. Patients were examined at the time of last follow-up. Progression-free survival (PFS) was defined as the time from surgery to the first recurrence or death from disease. For PFS analysis, patients were examined if they were alive with or without disease at the time of last follow-up. The data were plotted as Kaplan–Meier curves, and the statistical significance was determined by the log-rank test.
Abbreviations
OCCC: ovarian clear cell carcinoma; OEC: ovarian endometrioid carcinoma; ERONs: endometriosis-related ovarian neoplasms; TCGA: The Cancer Genome Atlas; POLE: polymerase epsilon; ARID1A:AT-rich interactive domain 1A; EC: endometrial cancer
Author contributions
Dr. Nakayama designed the study, and supervised all experiments. All other authors contributed to data collection. All authors approved the final version of the manuscript, and agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. 2015; 65:5e29.
2. Koshiyama M, Matsumura N, Konishi I. Recent concepts of ovarian carcinogenesis: type 1 and type 2. Biomed Res Int. 2014; 2014:934261.
3. Haruta S, Furukawa N, Yoshizawa Y, Tsunemi T, Nagai A, Kawaguchi R, Tanase Y, Yoshida S, Kobayashi H. Molecular genetics andepidemiology of epithelial ovarian cancer (review). Oncology reports. 2011; 26:1347–1356.
4. Maeda D, Shih IeM. Pathogenesis and the role of ARID1A mutation in endometriosis-related ovarian neoplasms. Adv Anat Pathol. 2013; 20:45–52.
5. Munksgaard PS, Blaakaer J. The association between endometriosis and ovarian cancer: a review of histological, genetic and molecular alterations. Gynecologic Oncology. 2012; 124:164–169.
6. Yamamoto S, Tsuda H, Takano M, Iwaya K, Tamai S, Matsubara O. PIK3CA mutation is an early event in the development of endometriosis-associated ovarian clear cell adenocarcinoma. J Pathol. 2011; 225:189–214.
7. Sato N, Tsunoda H, Nishida M, Morishita Y, Takimoto Y, Kubo T, Noguchi M. Loss of heterozygosity on 10q23.3 and mutation of the tumor suppressor gene PTEN in benign endometrial cyst of the ovary: possible sequence progression from benign endometrial cyst to endometrioid carcinoma and clear cell carcinoma of the ovary. Cancer Res. 2000; 60:7052–7056.
8. Amemiya S, Sekizawa A, Otsuka J, Tachikawa T, Saito H, Okai T. Malignant transformation of endometriosis and geneticalterations of K-ras and microsatellite instability. Int J Gynaecol Obstet. 2004; 86:371–376.
9. Ali-Fehmi R, Khalifeh I, Bandyopadhyay S. Patterns of loss of heterozygosity at 10q23.3 and microsatellite instability in endometriosis, atypical endometriosis, and ovarian carcinoma arising in association with endometriosis. Int J Gynecol Pathol. 2006; 25:223–229.
10. Yamamoto S, Tsuda H, Takano M, Tamai S, Matsubara O. Loss of ARID1A protein expression occurs as an early event in ovarian clear-cell carcinoma development and frequently coexists with PIK3CA mutations. Mod Pathol. 2012; 25:615–624.
11. Ayhan A, Mao TL, Seckin T, Wu CH, Guan B, Ogawa H, Futagami M, Mizukami H, Yokoyama Y, Kurman RJ, Shih IM. Loss of ARID1A expression is an early molecular event in tumor progression from ovarian endometriotic cyst to clear cell and endometrioid carcinoma. International journal of gynecological cancer. 2012; 22:1310-5.
12. Gadducci A, Lanfredini N, Tana R. Novel insights on the malignant transformation of endometriosis into ovarian carcinoma. Gynecol Endocrinol. 2014; 30:612–617.
13. Bitler BG, Aird KM, Garipov A, Li H, Amatangelo M, Kossenkov AV, Schultz DC, Liu Q, Shih IeM, Conejo-Garcia JR, Speicher DW, Zhang R. Synthetic lethality by targeting EZH2 methyltransferase activity in ARID1A-mutated cancers. Nat Med. 2015; 21:231–238.
14. Morey L, Helin K. Polycomb group protein-mediated repression of transcription. Trends Biochem Sci. 2010; 35:323–32.
15. Jones S, Wang TL, Shih IeM, Mao TL, Nakayama K, Roden R, Glas R, Slamon D, Diaz LA Jr, Vogelstein B, Kinzler KW, Velculescu VE, Papadopoulos N. Frequent mutations of chromatin remodeling gene ARID1A in ovarian clear cell carcinoma. Science. 2010; 330:228–231.
16. Wiegand KC, Shah SP, Al-Agha OM, Zhao Y, Tse K, Zeng T, Senz J, McConechy MK, Anglesio MS, Kalloger SE, Yang W, Heravi-Moussavi A, Giuliany R, et al. ARID1A mutations in endometriosis-associated ovarian carcinomas. N Engl J Med. 2010; 363:1532–1543.
17. Kandoth C, Schultz N, Cherniack AD. Clinically, patiens in the POLE ultramutated group reportedly had improved progression-free survival (PFS). Cancer genome atlas research network integrated genomic characterization of endometrial carcinoma. Nature. 2013; 497:67–73.
18. Hoang LN, McConechy MK, Köbel M, Anglesio M, Senz J, Maassen M, Kommoss S, Meng B, Postovit L, Kelemen LE, Staebler A, Brucker S, Krämer B, et al. Polymerase Epsilon Exonuclease Domain Mutations in Ovarian Endometrioid Carcinoma. Int J Gynecol Cancer. 2015; 25:1187–1193.
19. Nakayama K, Nakayama N, Kurman RJ, Cope L, Pohl G, Samuels Y, Velculescu VE, Wang TL, Shih IeM. Sequence mutations and amplification of PIK3CA and AKT2 genes in purified ovarian serous neoplasms. Cancer Biol Ther. 2006; 5:779–785.
20. Singer G, Stohr R, Cope L, Dehari R, Hartmann A, Cao DF, Wang TL, Kurman RJ, Shih IM. Patterns of p53 mutations separate ovarian serous borderline tumors and low- and high-grade carcinomas and provide support for a new model of ovarian carcinogenesis: a mutational analysis with immunohistochemical correlation. Am J Surg Pathol. 2005; 29:218–224.
21. Leitao MM, Soslow RA, Baergen RN, Olvera N, Arroyo C, Boyd J. Mutation and expression of the TP53 gene in early stage epithelial ovarian carcinoma. Gynecol Oncol. 2004; 93:301–306.
22. Wang Y, Helland A, Holm R, Kristensen GB, Borresen-Dale AL. PIK3CA mutations in advanced ovarian carcinomas. Hum Mutat. 2005; 25:322.
23. Levine DA, Bogomolniy F, Yee CJ, Lash A, Barakat RR, Borgen PI, Boyd J. Frequent mutation of the PIK3CA gene in ovarian and breast cancers. Clin Cancer Res. 2005; 11:2875–2878.
24. Singer G, Oldt R 3rd, Cohen Y, Wang BG, Sidransky D, Kurman RJ, Shih IeM. Mutations in BRAF and KRAS characterize the development of low-grade ovarianserous carcinoma. J Natl Cancer Inst. 2003; 95:484–486.
25. Ho CL, Kurman RJ, Dehari R, Wang TL, Shih IM. Mutations of BRAF and KRAS precede the development of ovarian serous borderline tumors. Cancer Res. 2004; 64:6915–6918.
26. Pearce CL, Templeman C, Rossing MA, Lee A, Near AM, Webb PM, Nagle CM, Doherty JA, Cushing-Haugen KL, Wicklund KG, Chang-Claude J, Hein R, Lurie G, et al. Association between endometriosis and risk of histological subtypes of ovarian cancer: pooled analysis of case-control studies. Lancet Oncol. 2012; 13:385–394.
27. Kobayashi H, Sumimoto K, Moniwa N, Imai M, Takakura K, Kuromaki T, Morioka E, Arisawa K, Terao T. Risk of developing ovarian cancer among women with ovarian endometrioma: a cohort study in Shizuoka, Japan. Int J Gynecol Cancer. 2007; 17:37–43.
28. Dinulescu DM, Ince TA, Quade BJ, Shafer SA, Crowley D, Jacks T. Role of K-ras and Pten in the development of mousemodels of endometriosis and endometrioid ovarian cancer. Nat Med. 2005; 11:63–70.
29. Kuhn E, Wu RC, Guan B, Wu G, Zhang J, Wang Y, Song L, Yuan X, Wei L, Roden RB, Kuo KT, Nakayama K, Clarke B, et al. Identification of molecular pathway aberrations in uterine serous carcinoma by genome-wide analyses. J Natl Cancer Inst. 2012; 104:1503–1513.
30. Grindedal EM, Heramb C, Karsrud I, Ariansen SL, Mæhle L, Undlien DE, Norum J, Schlichting E. Current guidelines for BRCA testing of breast cancer patients are insufficient to detect all mutation carriers. BMC Cancer. 2017; 17:438.
31. Heitzer E, Tomlinson I. Replicative DNA polymerase mutations in cancer. Curr Opin Genet Dev. 2001; 24:107Y113.
32. Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nature. 2012; 487:330–337.
33. Zou Y, Liu FY, Liu H, Wang F, Li W, Huang MZ, Huang Y, Yuan XQ, Xu XY, Huang OP, He M. Frequent POLE1 p.S297F mutation in Chinese patients with ovarian endometrioid carcinoma. Mutat Res. 2014; 761:49–52.
34. Church DN, Briggs SE, Palles C, Domingo E, Kearsey SJ, Grimes JM, Gorman M, Martin L, Howarth KM, Hodgson SV, Kaur K, Taylor J, Tomlinson IP; NSECG Collaborators. DNA polymerase epsilon and delta exonuclease domain mutations in endometrial cancer. Hum Mol Genet. 2013; 22:2820–2828.
35. Church DN, Stelloo E, Nout RA, Valtcheva N, Depreeuw J, ter Haar N, Noske A, Amant F, Tomlinson IP, Wild PJ, Lambrechts D, Jürgenliemk-Schulz IM, Jobsen JJ, et al. Prognostic significance of POLE proofreading mutations in endometrial cancer. J Natl Cancer Inst. 2015; 107:402.
36. Robert C, Schachter J, Long GV, Arance A, Grob JJ, Mortier L, Daud A, Carlino MS, McNeil C, Lotem M, Larkin J, Lorigan P, Neyns B, et al. Pembrolizumab versus ipilimumab in advanced melanoma. N Engl J Med. 2015; 372:2521–2532.
37. Brahmer J, Reckamp KL, Baas P, Crinò L, Eberhardt WE, Poddubskaya E, Antonia S, Pluzanski A, Vokes EE, Holgado E, Waterhouse D, Ready N, Gainor J, et al. Nivolumab versus docetaxel in advanced squamous-cell non-small-cell lung cancer. N Engl J Med. 2016; 373:123–135.
38. Hamanishi J, Mandai M, Ikeda T, Minami M, Kawaguchi A, Murayama T, Kanai M, Mori Y, Matsumoto S, Chikuma S, Matsumura N, Abiko K, Baba T, et al. Safety and antitumor activity of anti-PD-1 antibody, nivolumab, in patients with platinum-resistant ovarian cancer. J Clin Oncol. 2015; 33:4015–4022.
39. Stelloo E, Bosse T, Nout RA, MacKay HJ, Church DN, Nijman HW, Leary A, Edmondson RJ, Powell ME, Crosbie EJ, Kitchener HC, Mileshkin L, Pollock PM, et al. Refining prognosis and identifying targetable pathways for high-risk endometrialbcancer; a TransPORTEC initiative. Mod Pathol. 2015; 28:836–844.
40. Morice P, Leary A, Creutzberg C, Abu-Rustum N, Darai E. Endometrialcancer. Lancet. 2016; 387:1094–1108.
41. Meng B, Hoang LN, McIntyre JB, Duggan MA, Nelson GS, Lee CH, Köbel M. POLE exonuclease domain mutation predicts long progression-free survival in grade 3 endometrioid carcinoma of the endometrium. Gynecol Oncol. 2014; 134:15–19.
42. van Gool IC, Eggink FA, Freeman-Mills L, Stelloo E, Marchi E, de Bruyn M, Nijman HW, Bosse T. POLE proofreading mutations elicit an antitumor immune response in endometrial cancer. Clin Cancer Res. 2015; 21:3347–3355.
43. Guan B, Wang TL, Shih IeM. ARID1A, a factor that promotes formation of SWI/SNF-mediated chromatin remodeling, is a tumor suppressor in gynecologic cancers. Cancer Res. 2011; 71:6718–6727.
44. Bosse T, ter Haar NT, Seeber LM, v Diest PJ, Hes FJ, Vasen HF, Nout RA, Creutzberg CL, Morreau H, Smit VT. Loss of ARID1A expression and its relationship with PI3K-Akt pathway alterations, TP53 and microsatellite instability in endometrial cancer. Mod Pathol. 2013; 26:1525–1535.
45. Walton J, Blagih J, Ennis D, Leung E, Dowson S, Farquharson M, Tookman LA, Orange C, Athineos D, Mason S, Stevenson D, Blyth K, Strathdee D, et al. CRISPR/Cas9-mediated Trp53 and brca2 knockout to generate improved murine models of ovarian high-grade serous carcinoma. Cancer Res. 2016; 76:6118–6129.