
INTRODUCTION
Osteosarcoma is the most frequent primary malignancy bone tumor and occurs mainly in adolescents and young adults [1]. Neoadjuvant chemotherapy with surgical resection has markedly improved the prognosis in the past few decades [2]. However, the prognosis of osteosarcoma is still poor due to a relatively high rate of metastasis [3, 4]. The 5-year survival rate of patients with lung metastasis is only 30% [5]. Therefore, identification of the biological process and molecular mechanism involved in the metastasis of osteosarcoma is of significant value for improving the patients’ prognosis.
MicroRNAs (miRNAs) are small non-coding single stranded RNAs that suppress a large number of target genes [6, 7]. Accumulating evidence has indicated that miRNAs participate in a broad range of biological processes, such as proliferation, apoptosis, invasion and metastasis of different kind of tumors [8]. MiR-495, as a tumor-suppressive miRNA in most cases, has been shown to be under-expressed and to repress various target genes to inhibit the malignant processes of several types of tumor [9, 10]. However, there are different views on the role of miR-495 in regulating tumor invasion and migration. It has been reported that miR-495 suppresses the aggressive behaviors of glioma [11], prostate cancer [12] and esophageal squamous cell carcinoma cells [13]. However, in breast [14], bladder [15] and gastric [16] cancer cells, this miRNA promotes tumor invasion and migration. Importantly, there has been no research focusing on the role of miR-495 in osteosarcoma metastasis. Therefore, to explore how miR-495 participates in osteosarcoma progression, especially its effect on the invasion and migration of osteosarcoma cells, aiming at discovering novel targets for treatment becomes our goal of study.
In this study, for the first time, we confirmed the suppressive role of miR-495 in osteosarcoma metastasis both in vitro and in vivo. Also, we are the first to demonstrate that miR-495 directly suppresses HSP90AA1 to inhibit the invasion and migration of osteosarcoma cells. This metastasis-suppressive function of miR-495 is achieved by inhibiting the epithelial-mesenchymal transition (EMT) via HSP90AA1-mediated the PI3K/Akt/mTOR pathway.
RESULTS
miR-495 is downregulated in osteosarcoma tissues and cell lines
Paraffin-embedded tissue specimens of matched normal soft tissues and primary tumor tissues were collected from 10 osteosarcoma patients in XiJing Hospital. We extracted the total RNA including miRNAs in these tissues and qRT-PCR analyses showed that compared with the average expression level in normal soft tissues, miR-495 was significantly downregulated in tumor tissues (Figure 1A). Similarly, we found that compared with the normal osteoblast cell line hFOB1.19, osteosarcoma cell lines HOS, MG-63 and Saos-2 all showed decreased level of miR-495, among which MG-63 expressed the lowest level and HOS expressed the highest level (Figure 1B).
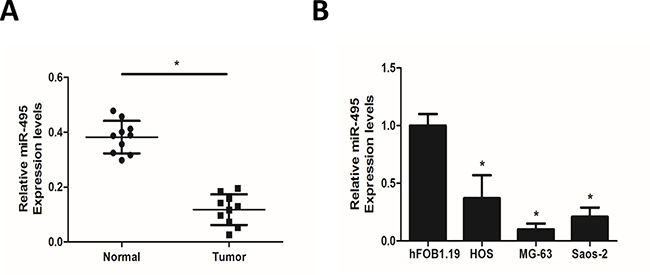
Figure 1: Expression of miR-495 in osteosarcoma tissue samples and cell lines. (A) qRT-PCR analyses of miR-495 in matched normal soft tissues and primary tumor tissues from 10 osteosarcoma patients. (B) Expression levels of miR-495 in normal osteoblast cell line hFOB1.19 and osteosarcoma cell lines HOS, MG-63 and Saos-2. Data represent the mean ± SD of three independent experiments. *P < 0.05.
miR-495 inhibits the invasion and migration of osteosarcoma cells in vitro
To test the effect of miR-495 on the invasion and migration of osteosarcoma cells, we introduced mimic NC/miR-495 mimic into MG63 cells which had lower level of miR-495 and transfected HOS and Saos-2 cells which had higher levels of miR-495 with inhibitor NC/miR-495 inhibitor (Figure 2). Wound-healing assays and matrigel invasion assays indicated that when exogenous miR-495 was introduced, MG-63 cells showed decreased capability of migrating to the monolayer of wounded cells (Figure 2A) and invading through the matrigel treated trans-well chambers (Figure 2B). On the contrary, when the endogenous miR-495 was inhibited, HOS and Saos-2 cells presented increased migration and invasion ability compared with inhibitor NC-treated cells (Figure 2A and 2B). These results demonstrated that miR-495 inhibits the invasion and migration of osteosarcoma cells in vitro.
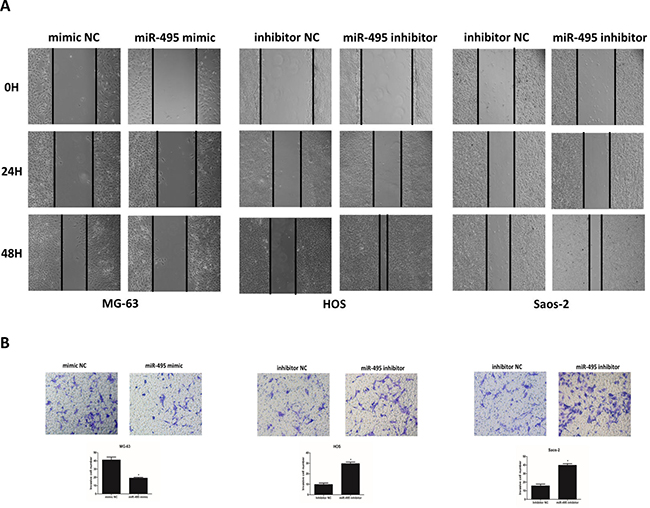
Figure 2: miR-495 inhibits the invasion and migration of osteosarcoma cells in vitro. (A, B) Osteosarcoma cell line MG-63 was transfected with mimic NC/miR-495 mimic, and cell lines HOS and Saos-2 were transfected with inhibitor NC/miR-495 inhibitor, followed by wound-healing assays (A) and matrigel invasion assays (B). Data are represented as the mean ± SD of three independent experiments. *P < 0.05.
miR-495 suppresses EMT as well as the invasion and migration of osteosarcoma cells in vivo
To further explore if the anti-metastatic miR-495 also inhibits epithelial-mesenchymal transition (EMT) of osteosarcoma cells, we tested levels of epithelial protein E-cadherin, mesenchymal protein Vimentin and transcription factors Snail1 and zinc finger e-box binding homeobox (ZEB1) which are known EMT markers under the above-mentioned treatments (Figure 3A and 3B). Results showed that miR-495 mimic treatment significantly drove the MG-63 cells towards an anti-EMT direction, indicated by decreased Snail1 and ZEB1 on mRNA level (Figure 3A) and decreased Vimentin and increased E-cadherin on both mRNA and protein levels (Figure 3A and 3B). Whereas miR-495 inhibitor treatment resulted in significant changes towards EMT in both HOS and Saos-2 cells (Figure 3A and 3B). Since Lung is the main metastatic site of osteosarcoma patients, nude mice model for lung metastasis was established to test the anti-metastatic role of miR-495 in vivo. We injected MG-63/Lenti-NC and MG-63/Lenti-miR-495 cells into the tail vein of nude mice. After sacrificing the mice at Day 28, we found that H&E staining of the lung tissues and counting under the microscope both showed smaller and less metastatic nodules in the MG-63/Lenti-miR-495 group compared with MG-63/Lenti-NC group (Figure 3C). These data suggested that miR-495 suppresses EMT as well as the invasion and migration of osteosarcoma cells in vivo.
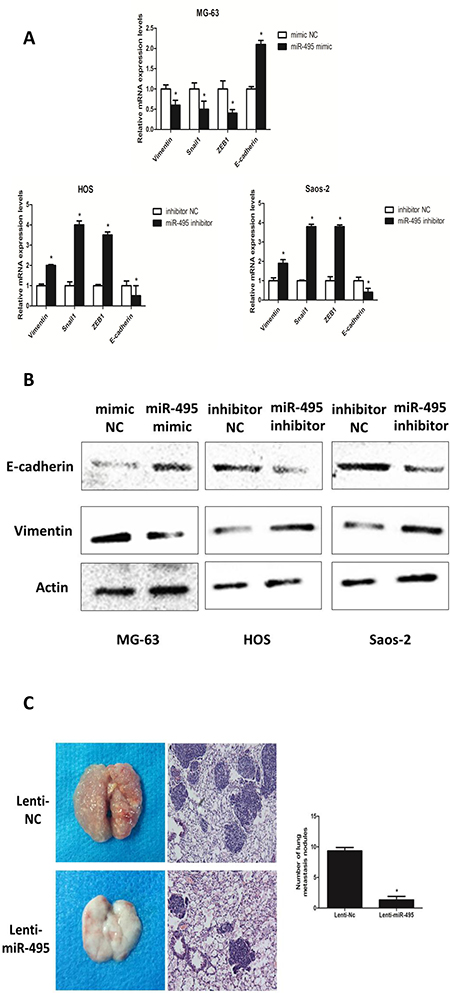
Figure 3: miR-495 inhibits EMT and the invasion and migration of osteosarcoma cells in vivo. (A, B) EMT markers (Vimentin, Snail 1, ZEB1 and E-cadherin) were analyzed by qRT-PCR (A) and levels of E-cadherin and Vimentin were also determined by western blot analyses (B) in mimic NC/miR-495 mimic-treated MG-63 cells and inhibitor NC/miR-495 inhibitor-treated HOS and Saos-2 cells. (C) lungs of nude mice in Lenti-NC/Lenti-miR-495 groups were analyzed by H&E staining of the metastatic tumor tissues. The numbers of metastatic lung nodules per nude mouse were counted under a microscope. Data are represented as the mean ± SD of three independent experiments. *P < 0.05.
HSP90AA1 is a direct target of miR-495
Using publicly available algorithms of TargetScan, PicTar and miRanda, we found HSP90AA1 the sole target gene of miR-495 predicted by all of the three algorithms. To verify this prediction, we cloned the 3′-untranslational region (3′ UTR) of HSP90AA1 with wild type (WT) and mutated (MT) miR-495 target site into a PGL3-msc2 vector (Figure 4A). In the luciferase reporter assays, readings of the relative activity markedly decreased in the miR-495 mimic- and WT 3′ UTR vector co-treated MG-63 cells and significantly increased in the miR-495 inhibitor- and MT 3′ UTR vector co-transfected HOS cells compared with the NC groups (Figure 4B). No significant differences were found in the MT 3′UTR vector-treated groups (Figure 4B). Furthermore, in Lenti-miR-495- and miR-495 mimic-treated MG-63 cells, the expression of HSP90AA1 is significantly downregulated and in miR-495 inhibitor-treated HOS and Saos-2 cells, the HSP90AA1 expression increased (Figure 4C). These data indicated that miR-495 inhibits HSP90AA1 expression by directly binding to its 3′ UTR.
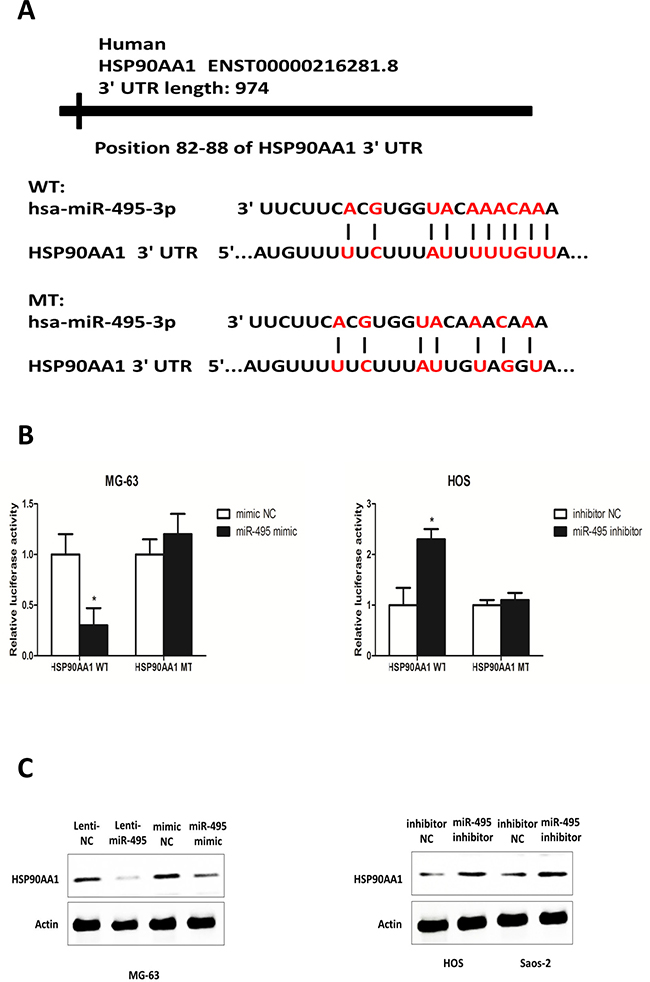
Figure 4: HSP90AA1 is a direct target of miR-495 in osteosarcoma cells. (A) Diagram of HSP90AA1 3′UTR containing the putative conserved target sequence for miR-495. WT: wild type; MT: mutant type. The seed regions of miR-495 and the seed-recognizing sites in the HSP90AA1 3′UTR are indicated in red. (B) Results of luciferase reporter assays in MG-63 cells and HOS cells co-transfected with HSP90AA1 WT/MT 3′UTR vectors and miR-495 mimic or miR-495 inhibitor as indicated for 48 h. (C) Expression levels of HSP90AA1 analyzed by western blot after transfection of mimic NC/miR-495 mimic into MG-63 cells and inhibitor NC/miR-495 inhibitor into HOS and Saos-2 cells. Data are represented as the mean ± SD of three independent experiments. *P < 0.05.
miR-495 inhibits the invasion and migration of osteosarcoma cells through HSP90AA1
To determine the role of HSP90AA1 in the miR-495-mediated anti-metastatic effect on osteosarcoma cells, we transfected MG-63 cells with HSP90AA1 shRNA with or without miR-495 inhibitor (Figure 5A and 5C) and treated HOS and Saos-2 cells with pHBLV-HSP90AA1 which contained the coding sequence of HSP90AA1 with or without miR-495 mimic (Figure 5B, 5D, 5E). Results from wound-healing assays and metrigel invasion assays showed that inhibition of endogenous HSP90AA1 suppressed the invasion and migration of MG-63 cells (Figure 5C) and expression of exogenous HSP90AA1 enhanced these two malignant features of HOS and Saos-2 cells (Figure 5D and 5E). In addition, the silencing of endogenous HSP90AA1 by shRNA partially reversed the miR-495 inhibitor-induced cell migration and invasion (Figure 5C), whereas the introduction of exogenous HSP90AA1 by pHBLV-HSP90AA1 partially neutralized the miR-495 mimic-mediated suppression on the migration and invasiveness of osteosarcoma cells (Figure 5D and 5E). Collectively, these data suggested that miR-495 inhibits the invasion and migration of osteosarcoma cells through HSP90AA1.
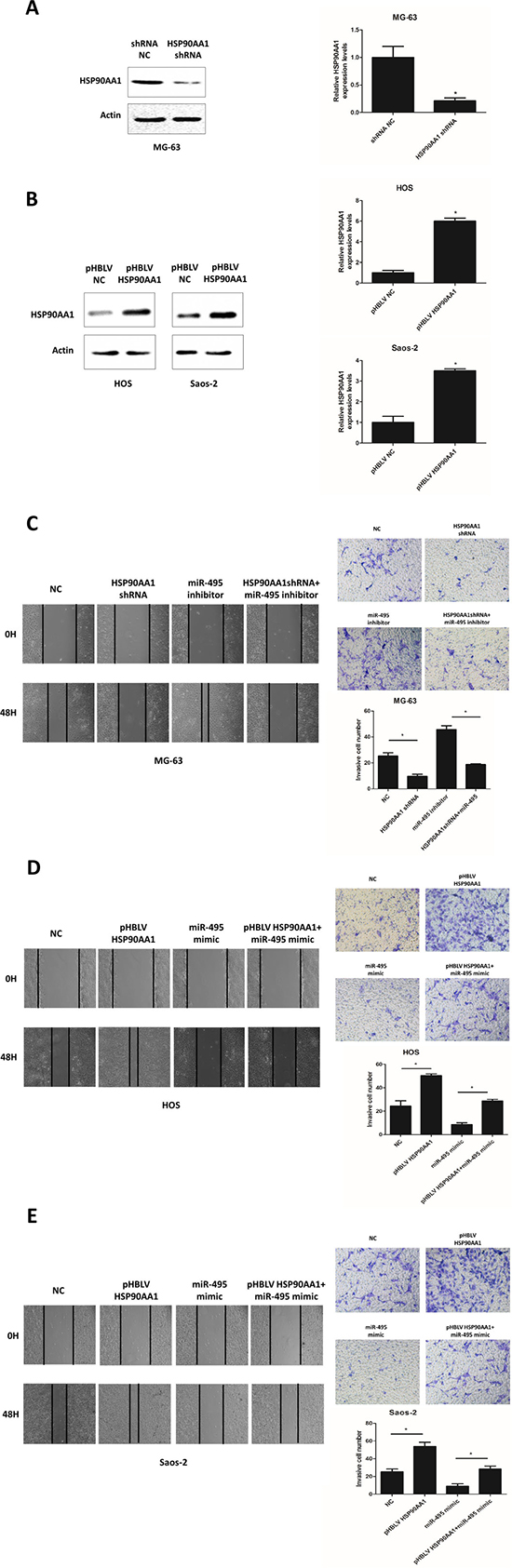
Figure 5: HSP90AA1 is involved in miR-495 mediated tumor suppressive effects. (A, B) Expression levels of HSP90AA1 assessed by Western blot and qRT-PCR in MG-63 cells transfected with control shRNA (shRNA NC) and HSP90AA1 shRNA for 48 hours (A), and HOS and Saos-2 cells transfected with pHBLV control vector (pHBLV NC) and pHBLV HSP90AA1 for 48 hours (B). (C–E) miR-495 inhibitor and shHSP90AA1 treated MG-63 cells (C) and miR-495 mimic and pHBLV HSP90AA1 treated HOS and Saos-2 cells (D, E) were subjected to wound-healing assays and matrigel invasion assays. Data are represented as the mean ± SD of three independent experiments. *P < 0.05.
Effects of HSP90AA1 on the PI3K/AKT signaling pathway
PI3K/AKT/mTOR is one of the most important signaling pathways in tumor progression and promotes invasion and metastasis in many cancers [17–19]. To explore if this pathway promotes osteosarcoma metastasis, we determined the expression of key molecules AKT and mTOR, and the changes in their phosphorylation levels by western blot analysis. As depicted in Figure 6A, HSP90AA1 knockdown was able to reduce the AKT and mTOR phosphorylation levels and to drive an anti-EMT process on MG-63 cells. Meanwhile, exogenous HSP90AA1 expression increased the phosphorylation level of these two key molecules and an EMT alteration could also be seen in HOS cells under pHBLV HSP90AA1 treatment (Figure 6A). Moreover, we treated the pHBLV HSP90AA1-transfected cells with the PI3K/AKT signaling pathway inhibitor LY-294002 (Figure 6B). The results indicated that the inhibitor could significantly inhibit the PI3K/AKT pathway, as well as the EMT process implicated by increased E-cadherin and decreased Vimentin levels, both of which were activated by HSP90AA1 upregulation (Figure 6A and 6B). These data demonstrated that HSP90AA1 induces EMT to promote osteosarcoma invasion and metastasis by the PI3K/AKT signaling pathway. Taken together, miR-495 inhibit HSP90AA1 expression and, then the downregulated-HSP90AA1 inhibits EMT by blocking PI3K/AKT signaling pathway.
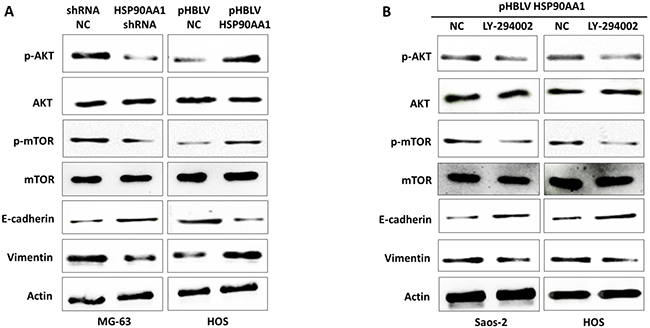
Figure 6: Effects of HSP90AA1 on the PI3K/AKT signaling pathway. (A) Western blot analysis for total and phosphorylated key proteins in the PI3K/Akt/mTOR pathway and EMT in MG-63 cells transfected with shRNA NC and HSP90AA1 shRNA and HOS cells transfected with pHBLV NC and pHBLV HSP90AA1 for 48 hours. (B) Western blot analysis for key proteins in the PI3K/Akt/mTOR pathway and EMT in Saos-2 and HOS cells treated with pHBLV HSP90AA1 with or without the PI3K/AKT pathway inhibitor LY294002. Data are represented as the representative of three independent experiments.
DISCUSSION
MiRNAs can function as tumor-suppressive genes or oncogenes associated with a broad range of tumor biological processes, such as recurrence and metastasis which can severely reduce the survival rate of patients with osteosarcoma. Dysregulation of miR-495 is frequently observed in human cancers, but the role of miR-495 in regulating tumor invasion and migration is controversial. In the present study, we provided the first evidence for the downregulation of miR-495 in osteosarcoma tissues and cell lines and revealed that miR-495 inhibits the invasion and metastasis of osteosarcoma cells both in vitro and in vivo.
We used strictly matched normal soft tissues and primary tumor tissues from 10 osteosarcoma patients to identify the expression levels of miR-495. The downregulation of miR-495 was found in both osteosarcoma tissues and cell lines, which demonstrated the anti-tumor effect of miR-495 in osteosarcoma. To validate the biological function of miR-495 downregulation, we transfected HOS and Saos-2 cells with miR-495 inhibitors and demonstrated that knockdown of miR-495 increased the invasive and metastatic capacities of osteosarcoma cells. Conversely, overexpression of miR-495 in MG-63 cells hampered cell invasion and migration.
To further prove the anti-metastatic role of miR-495, we analyzed the alterations of EMT markers following the increase or decrease of miR-495 level. Tumor metastasis begins with EMT, a highly conserved cellular program by which cells lose intercellular adhesion and obtain migratory and invasive capabilities [20–23]. This important process generates an aggressive tumor phenotype that leads to local metastasis via direct invasion or to distant metastasis via lymphatic and circulatory systems [24]. Tumor metastasis can presumably be inhibited by targeting factors involved in EMT [25–27]. Our results revealed that overexpression of miR-495 suppressed the process of EMT with upregulated expression of the epithelial marker E-cadherin, and decreased expression of the mesenchymal markers Vimentin. Conversely, decreased E-cadherin expression and increased expression of Vimentin were found after transfection with miR-495 inhibitors. The transcription factors Snail and ZEB1 which are involved in the process of EMT were downregulated in cells transfected with miR-495 mimics, and upregulated in cells transfected with miR-495 inhibitors. These results demonstrated that miR-495 could inhibit the process of EMT and thereby reduce the invasion and metastasis of osteosarcoma cells. In agreement with in vitro findings, our in vivo study also confirmed overexpression of miR-495 suppressed tumor metastasis in osteosarcoma xenografts.
We further demonstrated that HSP90AA1 is a direct target gene of miR-495. HSP90AA1 is a molecular chaperone that is up regulated in several cancers and promote the survival of cancer cells [28, 29]. It has been reported that increased level of HSP90AA1 also suggests poor prognosis of breast and pancreatic cancer patients [30, 31]. Therefore, HSP90AA1 is regarded essential for malignant transformation and progression in multiple cancer cells. We hypothesized that miR-495 may exert its anti-metastasis role in osteosarcoma via targeting of HSP90AA1 since bioinformatics analysis predicted that the 3′-UTR of HSP90AA1 mRNA had a complementary site for miR-495 binding. We used western blot analysis and luciferase reporter assay to further confirm that HSP90AA1 is indeed a target of miR-495 in osteosarcoma cells. Furthermore, Rescue experiments validated that depletion of HSP90AA1 reversed the promoted cell invasion and migration in miR-495 knockdown cells. Taken together, these results demonstrated for the first time that miR-495 inhibits osteosarcoma cell invasion and metastasis by directly targeting HSP90AA1.
The PI3K/AKT/mTOR pathway is an intracellular signaling pathway which plays important roles in regulating cell survival, growth and migration [32]. In many cancers, this signaling pathway is frequently activated and is essential for cancer invasion and metastasis [33–35]. It has been reported that EMT could be mediated by PI3K/AKT/mTOR pathway [36]. However, the relationship between HSP90AA1 and EMT remains unknow. The effect of HSP90AA1 on PI3K/AKT/mTOR pathway also remains unknown. In this study, we found that HSP90AA1 could upregulate the phosphorylation of AKT and mTOR, and at the same time induce EMT of osteosarcoma cells. In the meantime, inhibitor of the PI3K/AKT pathway not only blocked the activated PI3K/AKT/mTOR pathway upon HSP90AA1 upregulation, but also reversed the accompanying EMT process. These results further shed light on the molecular mechanisms underlying the tumor-suppressive effect of miR-495 targeting HSP90AA1 in osteosarcoma cells, which was mediated by downregulating the PI3K/Akt/mTOR signaling pathway.
In conclusion, we demonstrated significantly lower expression of miR-495 in human osteosarcoma tissues and cell lines. By targeting HSP90AA1, miR-495 suppresses osteosarcoma cell invasion and migration both in vitro and in vivo. Importantly, this metastasis-suppressive function of miR-495 is achieved by inhibiting the EMT via HSP90AA1-mediated the PI3K/Akt/mTOR pathway. Therefore, our study found that miR-495 may be a potential biomarker and therapeutic target in the clinical treatment of osteosarcoma.
MATERIALS AND METHODS
Human tissue samples
Ten human osteosarcoma tissues and adjacent normal soft tissues were collected from the Department of Orthopedics in Xijing Hospital affiliated to the Forth Military Medical University (Supplementary Table 1). All tissues were confirmed by histological examination and the diagnoses of these samples were verified by pathologists. The study was approved by the Medical Ethics Committee of Forth Military Medical University, and written informed consents for research purpose were obtained from each patient.
Cell culture
HOS, MG-63, Saos-2 and hFOB1.19 cell lines were purchased from the Cell Bank of Chinese Academy Sciences (Shanghai, China) and used for the in vitro experiments. All the cell lines used were authenticated and tested in the cell bank after the bank bought them from American Type Culture Collection (ATCC, Manassas, VA, USA) before sale. HOS and Saos-2 cells were cultured in McCoy’s 5A (Modified) Medium (Gibco, Los Angeles, CA, USA). MG-63 and hFOB1.19 were cultured in Dulbecco’s Modified Eagle Medium (DMEM, Gibco). All of the cell cultures were supplemented with 10% fetal bovine serum (FBS, Gibco) and were incubated in a humidified atmosphere of 5% CO2 at 37° C.
RNA isolation and quantitative real time-PCR (qRT-PCR)
Total RNA and miRNA were extracted from cultured cells or tissue samples with TRIzol Reagent (Invitrogen, Carlsbad, NM, USA) according to the manufacturer’s instructions. Reverse transcription reactions for mRNA and miRNA were then performed with PrimeScript™ RT Master Mix and SYBR® PrimeScript™ miRNA RT-PCR Kit (TaKaRa Bio Group, Shiga, Japan), respectively. qRT-PCR analyses were performed using SYBR® Premix Ex Taq™ II (TaKaRa) on a Bio-Rad CFX96 system (Bio-Rad, Hercules, CA, USA) according to the manufacturer’s protocols. β-actin mRNA was used as an endogenous control. The relative expressions of miRNAs were normalized by U6 SnRNA. Data were analyzed with comparative delta Ct method. The primers used for qRT-PCR analyses are in Supplementary Table 2.
Cell transfection
The overexpression or knockdown of miR-495 was accomplished by transfecting cells with synthetic miR-495 mimic or inhibitor purchased from Genephama (Shanghai, China). Overexpression vector pHBLV HSP90AA1 and HSP90AA1 shRNA were also obtained from GenePharma. Osteosarcoma cell lines were transfected with plasmids and oligonucleotides using Lipofectamine 3000 Transfection Reagent (Invitrogen) at final concentrations of 2 μg/ml and 50 nM respectively for 48 h, according to the manufacturer›s instructions. MG-63 cells were transfected with lentiviral particles of NC/miR-495 (Lenti-NC/Lenti-miR-495, Titer: 1.02 × 109 copies/ml, bought from GeneCopoeia, Guangzhou, China) according to the manufacturer’s instructions.
Cell migration and invasion assays
For the wound-healing assays, osteosarcoma cells were seeded into 6-well plate after transfected with miR-495 mimic, miR-495 inhibitor and mimic NC/inhibitor NC. A linear wound was carefully made by a sterile pipette tip across the confluent cell monolayer and then wound closures were observed by taking photographs under a microscope at 0, 24 and 48 hours. Matrigel invasion assays were performed with Matrigel (BD Biosciences, Heidelberg, Germany) and 8-μm, 24-well trans-well chambers (Millipore, Billerica, MA, USA) following the manufacturer’s instructions. Cells were diluted with serum-free culture medium and placed into the upper chamber. After cultured for 48 hours, migrated cells were stained with 0.1% crystal violet for 10 minutes at room temperature, and photographs were taken of ten randomly selected fields of fixed cells.
Western blot analysis
The protein concentration was quantified with the BCA method. Proteins were separated on sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), and then transferred onto polyvinyl fluoride (PVDF) membranes by using Mini-PROTEAN Tetra Electrophoresis System (Bio-Rad). The PVDF membrane was incubated with primary antibody against HSP90AA1 (1:1000 dilution, Proteintech 13171-1-AP), E-cadherin (1:100 dilution, Santa Cruz Biotechnology sc-7870), Vimentin (1:100 dilution, Santa Cruz Biotechnology sc-66002), p-Akt (1:1000 dilution, Abcam 38449), Akt (1:1000 dilution, Abcam ab8805), p-mTOR (1:1000 dilution, Abcam ab109268) and mTOR (1:1000 dilution, Abcam ab32028) overnight at 4°C, followed by an incubation with a horseradish peroxidase (HRP)-conjugated secondary antibody for 1 h at room temperature. β-actin (1:2000 dilution, Sigma-Aldrich A1978) was used as a loading control. The signals were detected by an enhanced chemiluminescence system (ECL System, Alpha Innotech, San Leandro, CA, USA) according to the manufacturer’s instructions.
Bioinformatics
The publicly available algorithms were used to predict potential miR-495 targets: TargetScan (http://www.targetscan.org/), PicTar (http://pictar.mdc-berlin.de/) and miRanda (http://www.microrna.org/microrna/home.do).
Luciferase reporter assay
Human HSP90AA1-3′ untranslational region (3′UTR) reporter plasmids containing the putative binding sequence of miR-495 (wild-type, WT) and its identical sequence with a mutation in the miR-495 seed sequence (mutant, MT) were synthesized by Hanbio (Shanghai, China). Both of these sequences were inserted between the EcoRI and EcoRV restriction sites of the pGL3-msc2 reporter vector (Promega, Madison, WI, USA), and validated by sequencing. Luciferase reporter assay was performed as previously described. In brief, MG-63 and HOS cells were seeded at 3 × 104/well in 48-well plate and each well was transfected with HSP90AA1-3′UTR WT/MT vectors (100 ng/well) and mimic NC/ miR-495 mimic (50 nM) for MG-63 cells or inhibitor NC/miR-495 inhibitor (50 nM) for HOS cells. A Renilla luciferase construct (pRL-TK; Promega; 10 ng/well) was transfected for normalization. The luciferase activity was measured 48 h after transfection using the Dual-Luciferase Reporter Assay System (Promega) according to the manufacturer’s instructions.
Animal experiments
Male Nude nu/nu mice (4–6 weeks old) were purchased from the Experimental Animal Center of the Forth Military Medical University and housed under specific pathogen-free conditions. To evaluate the lung metastatic potential of osteosarcoma cells in vivo, 5 × 106 MG-63/Lenti-NC or MG-63/Lenti-miR-495 cells in 200 μl of serum-free medium were injected into nude mice through the tail vein. Mice were sacrificed 4 weeks after cell injection. Individual organs from the mice were removed, and metastatic tissues (lung) were analyzed with H&E staining. All animal experiments complied with ethical regulations and humane treatment and were approved by the Medical Ethics Committee of FMMU.
Statistical analysis
Data are expressed as the mean ± standard. Student’s t-test or one-way ANOVA were used to evaluate statistical significance between two groups. One representative experiment is shown in the triplicates used for the statistical analysis. A P-value of < 0.05 was considered statistically significant.
ACKNOWLEDGMENTS AND FUNDING
This work was supported by grants from National Key R&D Plan (No. 2017YFC1104901) and Shaanxi Industrial Science and Technology Project (No. 2016GY-198). The authors thank Cassandra C. Garbutt for editing the English language.
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest. All institutional and national guidelines for the care and use of laboratory animals were followed. All procedures followed were in accordance with the ethical standards of the Medical Ethics Committee of Forth Military Medical University, and written informed consents for research purpose were obtained from each patient.
REFERENCES
1. Siegel RL, Miller KD, Jemal A. Cancer Statistics, 2017. CA Cancer J Clin. 2017; 67:7–30. https://doi.org/10.3322/caac.21387.
2. Messerschmitt PJ, Rettew AN, Brookover RE, Garcia RM, Getty PJ, Greenfield EM. Specific tyrosine kinase inhibitors regulate human osteosarcoma cells in vitro. Clin Orthop Relat Res. 2008; 466:2168–75. https://doi.org/10.1007/s11999-008-0338-9.
3. Bacci G, Longhi A, Versari M, Mercuri M, Briccoli A, Picci P. Prognostic factors for osteosarcoma of the extremity treated with neoadjuvant chemotherapy: 15-year experience in 789 patients treated at a single institution. Cancer. 2006; 106:1154–61. https://doi.org/10.1002/cncr.21724.
4. Longhi A, Errani C, De Paolis M, Mercuri M, Bacci G. Primary bone osteosarcoma in the pediatric age: state of the art. Cancer Treat Rev. 2006; 32:423–36. https://doi.org/10.1016/j.ctrv.2006.05.005.
5. Rosen G, Murphy ML, Huvos AG, Gutierrez M, Marcove RC. Chemotherapy, en bloc resection, and prosthetic bone replacement in the treatment of osteogenic sarcoma. Cancer. 1976; 37:1–11.
6. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009; 136:215–33. https://doi.org/10.1016/j.cell.2009.01.002.
7. Fabian MR, Sonenberg N, Filipowicz W. Regulation of mRNA translation and stability by microRNAs. Annu Rev Biochem. 2010; 79:351–79. https://doi.org/10.1146/annurev-biochem-060308-103103.
8. Ambros V. microRNAs: tiny regulators with great potential. Cell. 2001; 107:823–6.
9. Lv C, Bai Z, Liu Z, Luo P, Zhang J. MicroRNA-495 suppresses human renal cell carcinoma malignancy by targeting SATB1. Am J Transl Res. 2015; 7:1992–9.
10. Chu H, Chen X, Wang H, Du Y, Wang Y, Zang W, Li P, Li J, Chang J, Zhao G, Zhang G. MiR-495 regulates proliferation and migration in NSCLC by targeting MTA3. Tumour Biol. 2014; 35:3487–94. https://doi.org/10.1007/s13277-013-1460-1.
11. Zhang B, Yuan F, Liu J, Li Y, Zhou F, Liu X, Hao Z, Li Q, Zheng Y, Wang W. Hsa-miR-495 acts as a tumor suppressor gene in glioma via the negative regulation of MYB. Mol Med Rep. 2016; 14:977–82. https://doi.org/10.3892/mmr.2016.5327.
12. Li JZ, Wang ZL, Xu WH, Li Q, Gao L, Wang ZM. MicroRNA-495 Regulates Migration and Invasion in Prostate Cancer Cells Via Targeting Akt and mTOR Signaling. Cancer Invest. 2016; 34:181–8. https://doi.org/10.3109/07357907.2016.1156690.
13. Mao Y, Li L, Liu J, Wang L, Zhou Y. MiR-495 inhibits esophageal squamous cell carcinoma progression by targeting Akt1. Oncotarget. 2016; 7:51223–36. https://doi.org/10.18632/oncotarget.9981.
14. Cao M, Nie W, Li J, Zhang Y, Yan X, Guan X, Chen X, Zen K, Zhang CY, Jiang X, Hou D. MicroRNA-495 induces breast cancer cell migration by targeting JAM-A. Protein Cell. 2014; 5:862–72. https://doi.org/10.1007/s13238-014-0088-2.
15. Tan M, Mu X, Liu Z, Tao L, Wang J, Ge J, Qiu J. microRNA-495 promotes bladder cancer cell growth and invasion by targeting phosphatase and tensin homolog. Biochem Biophys Res Commun. 2017; 483:867–73. https://doi.org/10.1016/j.bbrc.2017.01.019.
16. Lee SH, Jung YD, Choi YS, Lee YM. Targeting of RUNX3 by miR-130a and miR-495 cooperatively increases cell proliferation and tumor angiogenesis in gastric cancer cells. Oncotarget. 2015; 6:33269–78. https://doi.org/10.18632/oncotarget.5037.
17. Aoki M, Fujishita T. Oncogenic Roles of the PI3K/AKT/mTOR Axis. Curr Top Microbiol Immunol. 2017; 407:153–89. https://doi.org/10.1007/82_2017_6.
18. Ersahin T, Tuncbag N, Cetin-Atalay R. The PI3K/AKT/mTOR interactive pathway. Mol Biosyst. 2015; 11:1946–54. https://doi.org/10.1039/c5mb00101c.
19. Mabuchi S, Kuroda H, Takahashi R, Sasano T. The PI3K/AKT/mTOR pathway as a therapeutic target in ovarian cancer. Gynecol Oncol. 2015; 137:173–9. https://doi.org/10.1016/j.ygyno.2015.02.003.
20. Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, Campbell LL, Polyak K, Brisken C, et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. 2008; 133:704–15. https://doi.org/10.1016/j.cell.2008.03.027.
21. Acloque H, Adams MS, Fishwick K, Bronner-Fraser M, Nieto MA. Epithelial-mesenchymal transitions: the importance of changing cell state in development and disease. J Clin Invest. 2009; 119:1438–49. https://doi.org/10.1172/jci38019.
22. Morales AV, Barbas JA, Nieto MA. How to become neural crest: from segregation to delamination. Semin Cell Dev Biol. 2005; 16:655–62. https://doi.org/10.1016/j.semcdb.2005.06.003.
23. Oft M, Heider KH, Beug H. TGFbeta signaling is necessary for carcinoma cell invasiveness and metastasis. Curr Biol. 1998; 8:1243–52.
24. Hong JH, Hwang ES, McManus MT, Amsterdam A, Tian Y, Kalmukova R, Mueller E, Benjamin T, Spiegelman BM, Sharp PA, Hopkins N, Yaffe MB. TAZ, a transcriptional modulator of mesenchymal stem cell differentiation. Science. 2005; 309:1074–8. https://doi.org/10.1126/science.1110955.
25. Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009; 139:871–90. https://doi.org/10.1016/j.cell.2009.11.007.
26. Huber MA, Kraut N, Beug H. Molecular requirements for epithelial-mesenchymal transition during tumor progression. Curr Opin Cell Biol. 2005; 17:548–58. https://doi.org/10.1016/j.ceb.2005.08.001.
27. Mimeault M, Batra SK. Interplay of distinct growth factors during epithelial mesenchymal transition of cancer progenitor cells and molecular targeting as novel cancer therapies. Ann Oncol. 2007; 18:1605–19. https://doi.org/10.1093/annonc/mdm070.
28. Gao J, Aksoy BA, Dogrusoz U, Dresdner G, Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, Cerami E, Sander C, Schultz N. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal. 2013; 6:pl1. https://doi.org/10.1126/scisignal.2004088.
29. Cerami E, Gao J, Dogrusoz U, Gross BE, Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, Antipin Y, Reva B, Goldberg AP, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012; 2:401–4. https://doi.org/10.1158/2159-8290.cd-12-0095.
30. Jameel A, Skilton RA, Campbell TA, Chander SK, Coombes RC, Luqmani YA. Clinical and biological significance of HSP89 alpha in human breast cancer. Int J Cancer. 1992; 50:409–15.
31. Gress TM, Muller-Pillasch F, Weber C, Lerch MM, Friess H, Buchler M, Beger HG, Adler G. Differential expression of heat shock proteins in pancreatic carcinoma. Cancer Res. 1994; 54:547–51.
32. Yap TA, Garrett MD, Walton MI, Raynaud F, de Bono JS, Workman P. Targeting the PI3K-AKT-mTOR pathway: progress, pitfalls, and promises. Curr Opin Pharmacol. 2008; 8:393–412. https://doi.org/10.1016/j.coph.2008.08.004.
33. Ni J, Cozzi P, Hao J, Beretov J, Chang L, Duan W, Shigdar S, Delprado W, Graham P, Bucci J, Kearsley J, Li Y. Epithelial cell adhesion molecule (EpCAM) is associated with prostate cancer metastasis and chemo/radioresistance via the PI3K/Akt/mTOR signaling pathway. Int J Biochem Cell Biol. 2013; 45:2736–48. https://doi.org/10.1016/j.biocel.2013.09.008.
34. Chang L, Graham PH, Hao J, Bucci J, Cozzi PJ, Kearsley JH, Li Y. Emerging roles of radioresistance in prostate cancer metastasis and radiation therapy. Cancer Metastasis Rev. 2014; 33:469–96. https://doi.org/10.1007/s10555-014-9493-5.
35. Chang L, Graham PH, Hao J, Ni J, Bucci J, Cozzi PJ, Kearsley JH, Li Y. PI3K/Akt/mTOR pathway inhibitors enhance radiosensitivity in radioresistant prostate cancer cells through inducing apoptosis, reducing autophagy, suppressing NHEJ and HR repair pathways. Cell Death Dis. 2014; 5:e1437. https://doi.org/10.1038/cddis.2014.415.
36. Zhang D, Wang S, Chen J, Liu H, Lu J, Jiang H, Huang A, Chen Y. Fibulin-4 promotes osteosarcoma invasion and metastasis by inducing epithelial to mesenchymal transition via the PI3K/Akt/mTOR pathway. Int J Oncol. 2017; 50:1513–30. https://doi.org/10.3892/ijo.2017.3921.