
INTRODUCTION
Cancer is a multifactor disease and it represents leading causes of death for both men and women all over the world [1], with approximately more than 18 billion cancer deaths projected to occur in the next five years [2]. In recent decades, an increase in mortality and morbidity rates of cancer has been demonstrated in developed or developing countries because of different types of pollutions (especially environmental), carcinogenic agents, unhealthy lifestyles, anxiety, and heavy workload [1]. Presently, quick changing cancer prevalence occurs throughout the world. National Cancer Institute of USA projected that, a total of 1,688,780 new cancer cases, as well as 600,920 cancer related deaths will be occur in 2017 in USA (National Cancer Institute SEER Data. [(Accessed on 25 June 2017)]; Available online: http://seer.cancer.gov/statfacts/).
Despite the significant development made in diagnosis and treatment, the current clinical therapies including radiotherapy, phototherapy, chemotherapy, immunotherapy and surgery are limited as indicated by the salient morbidity and mortality rate as well as low outcome from cancer, suggesting crucial demands for novel efficient therapeutic agents [2, 3]. Current treatment options mainly based on chemotherapy that significantly reduce therapeutic success in cancer owing to the presence of several disadvantages like multiple forms of drug resistance, severe adverse effects and complications, such as fatigue, pain, diarrhea, nausea, vomiting, and hair loss [1, 4]. Therefore, much attention has been paid for exploring a novel drug from natural sources with few side effects that is highly effective for cancer treatment is of great importance.
Recently, about 80% commercially available anticancer new drugs derived from natural resources such as from marine organisms, micro-organism and plants sources [5]. However, the less success rates of chemotherapies, multiple forms of drug resistance, severe adverse effects and complicacy emphasize the importance of discovering new compounds, which is considered as safe, cost effective and alternative to cancer treatment [6-8].
Complementary and alternative medicine (CAM), also known as “other than” conventional medicine, is a group of diverse medical and healthcare systems, practices and products [3]. Globally, approximately 9.8–76.0% of the populations have used some form of CAM that greatly varies from country to country [9]. Among the different kinds of CAM, ginseng has been extensively used in the oriental and presently in western medicines whose history stretches back nearly 5,000 years [10]. Etymologically, Panax means “all-healing” in Greek and people’s belief that it is a panacea that promotes longevity [11]. The name “ginseng” came from a Chinese word “rénshen” which literally means “essence of men” [4]. Genus Panax belongs to the family Araliaceae, which comprises of 11 different species, including P. ginseng C.A. Meyer (known as Asian or Korean ginseng), P. quinquefolius L. (American ginseng), P. notoginseng (Burkill) F.H. Chen (Chinese ginseng, also called Sanchi or notoginseng), are vital herbs that used to treat different medical conditions [12].
Pharmacological actions of ginseng are mainly imputed to a variety of ginsenosides, a triterpenoid saponins that are secondary metabolites uniquely present in Panax species. Other than saponins, ginseng also contains flavonoids, polyacetylenes, phytosterols, essential oils, acids, polysaccharides, nitrogen-containing compounds and vitamins [10, 13]. The ginseng root contains 2–3% ginsenosides and also found in other parts of this herb such as leaves, flowers and berries [14]. More than 180 known different ginsenosides with various numbers, linkage positions and types of sugar moiety have been isolated and identified since their first discovered in the 1960s by Shibata’s group [15, 16]. Amazingly, the antitumor activities of ginsenosides negatively linked to the number of sugar groups, the number and position of hydroxyl groups and stereoselectivity [17].
Ginsenosides can be divided into two major types based on the functional groups at C6 position, namely (1) protopanaxadiol type (PPD) (e.g., ginsenoside Rb1, Rb2, Rc, Rd, Rg3 and Rh2) contains a hydrogen atom at C6, (2) protopanaxatriol type (PPT) (e.g., ginsenoside Re, Rf, Rg1 and Rh1) contains a C-6 sugar chain. The minor types are oleanolic acid type (e.g., ginsenoside Ro) and ocotillol type (e.g., pseudoginsenoside) (Figure 1). Protopanaxadiol ginsenosides exhibits higher anticancer activities than protopanaxatriol type. Moreover, the anticancer ability of ginsenosides depends on the position of sugar linkages in the order: C-3 > C-6 > C-20 [18]. American ginseng manifested a greater affinity for free radicals scavenging and capable of delaying lipid peroxidation as well as higher content of ginsenosides compared to Asian ginseng [19, 20].
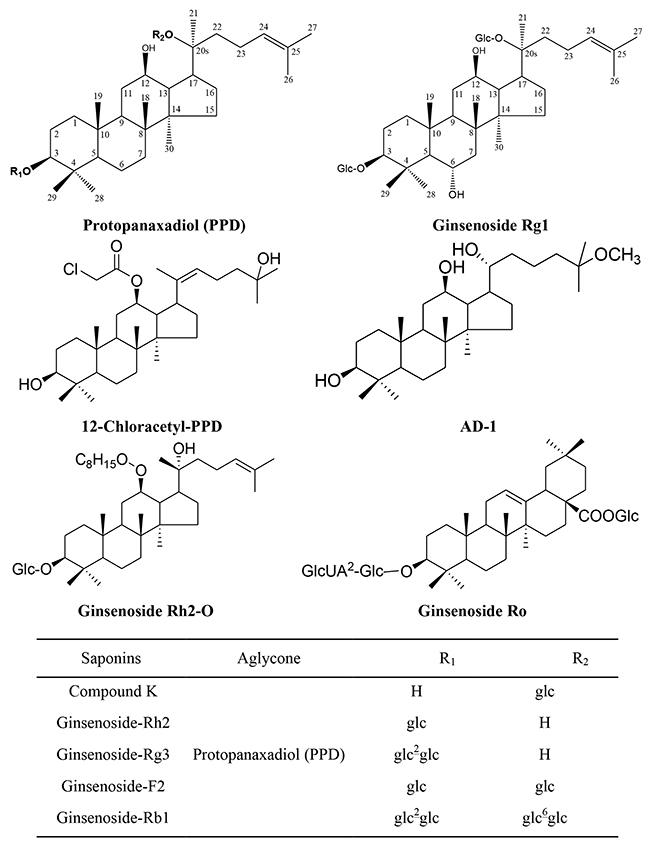
Figure 1: Chemical Structure of Ginsenosides Included in This Review. Glc: β-D-glucopyranosyl; GlcUA: β-D-glucuronyl-.
The scavenging effects of ginsenosides on intracellular ROS were ranked in the decreasing order: Rc > Rb2 > Rg2 > Rh2 > Rh1 > Rf > Rg3 > Rg1 > Rb1 > Re > Rd [21]. Recently, various reports have demonstrated that ginsenosides may be capable of enhancing therapeutic potential of conventional chemotherapeutic agents in case of cancer patients through defending normal tissues from chemotherapy-induced damage [4].
Ginsenosides and ginseng extract have been proved to possess an antioxidant, anti-inflammation, anti-fatigue, anti-stress, cure for respiratory problems, hemocytopenia therapy, cardioprotection, anti-obesity, anti-diabetic, immunomodulation, neuroprotection, antimicrobial action, sexual potentiating, antitumor [22], anti-allergic [23], antihypertensive [24] activities. Ginsenosides have been postulated to exert anticancer activities through cellular and molecular targets via modulation of diverse signaling pathways, thereby inhibiting the tumor by regulation of cell proliferation, inhibition of oxidative damage and inflammation, induction of apoptosis and autophagy, preventing angiogenesis, delaying invasion and metastasis, regulating tumor-related immune suppression, and increasing the sensitivity of resistant cells to chemotherapeutic agents [4, 17, 25].
Ginseng is popularly used in cancer patients, and several studies have assorted with ginseng consumption and risk of cancer prevention and treatment [4]. Various case-control studies proved that clinical uses of ginsengs have cancer prevention ability in the oral cavity, stomach, lung, liver, pancreas, ovary and colon by regular consumption [26]. Yun and Choi conducted a prospective cohort study to evaluate the preventive effect of ginseng against cancer on a population residing in a ginseng cultivation area on the basis of the result of case-control studies, and drew the conclusions that P. ginseng has non-organ specific preventive effect against cancer [27]. A 5 years follow-up cohort study of about 4600 non-cancer individuals revealed that risk of cancer considerably lower among ginseng intakers than non-intakers [28]. In a large, population-based cohort study of 1455 breast cancer patients in China showed that regular consumption of ginseng at a dose of 1.3 g per day before cancer diagnosis and nonstop to use it after the diagnosis had significantly decreased risk of death and recurrence equated with patients who never used ginseng [29]. Regular use of ginseng after cancer diagnosis was also positively linked with the quality of life scores, significant result in the psychological and social well-being domains [29]. In other study involving a meta-analysis of 274 female breast cancer patients in China also found that the combining treatment group showed considerably attenuated leucopenia and progress Karnosfsky Performance Scale score after chemotherapy [30].
It was reported that red ginseng powder inhibited the recurrence of AJCC stage III gastric cancer and showed immunomodulatory activities during postoperative chemotherapy, after a curative resection with D2 lymph node dissection [31]. In a randomized, double-blind, placebo-controlled pilot trial of 53 cancer patients, sun ginseng (3000 mg/day) was given daily for 12 weeks resulting in helpful ameliorating some expressions of mental and physical functioning, in case of gynecologic cancer and hepatobiliary cancer patients [32]. Another randomized, double-blind clinical study of 290 cancer patients also revealed that regular consumption of 750 mg of P. quinquefolius (American ginseng) does not contribute any benefit compared to the placebo, but two high doses (1000–2000 mg/day) did seem to reduce cancer-related fatigue more than a placebo [33]. In a recent randomized, double-blinded, placebo-controlled trial, 643 patients with chronic atrophic gastritis in China was given red ginseng extract powder (1 g/week) for 3 years and carried out for 8 years, and found that the administration of red ginseng extract powder for 3 years exerted significant preventive effects on the incidence of non-organ-specific human cancers in males [34].
A recent meta-analysis involving 7,436 cancer patients and 334,544 participants showed that ginseng consumption is associated with a significantly decreased risk of cancer (16%) and that the effect is not organ specific [35]. In a more recent randomized, double-blinded, placebo-controlled trial, 30 patients with epithelial ovarian cancer in Korea were given placebo or red ginseng (3000 mg/day) for three months. The results suggested that red ginseng may be capable of decreasing genotoxicity and enhancing health-related quality of life despite no benefit of survival in patients with epithelial ovarian cancer who received chemotherapy [36].
However, some contradictory and puzzling results were also come forward. For example, a large case–control study and a small cohort study have together proposed that ginseng consumption is related with a 60–70% decrease risk of gastric cancer in Korean populations. However, when the study was tried to be repeated in a large prospective cohort study of Chinese women, no relationship was found between ginseng consumption and risk of gastric cancer patients [37]. A retrospective case-control study of 949 breast cancer patients also showed that there is no significant relationship between ginseng consumption and risk of breast cancer patients [38]. The U.S. Food and Drug Administration (FDA) rates ginseng as GRAS (generally recognized as safe), which is generally known as a CAM in treatments of different types and stage of cancer in the U.S. and Europe [4].
In the past 30 years, massive attempts have also been made to recognize the anti-cancer activity of ginseng compounds, such as ginsenoside Rg3 and Rh2. Among them, some compounds are currently accessible as over-the-counter drugs in China and universal. Shenyi capsule (ginsenoside Rg3 monomer preparation), an autonomously formulated class A new anti-cancer drug in traditional Chinese medicine, has been generally applied clinically in the treatment of different types of cancer, including breast cancer, gastrointestinal tumors and lung cancer in China [39]. A prospective, randomized, controlled study of 133 non-small cell lung cancer patients exhibited that Shenyi capsule, mainly in combination with chemotherapy, augmented the post-operative lifespan of patients was also positively linked with increasing the immune function and suppressing angiogenesis [40]. Another randomized trial on 60 patients with advanced esophageal cancer patients establish that Shenyi capsule in combination with chemotherapy was efficient in ameliorating the patients’ quality of life and 1 year survival rates [41].
In addition, some preparations, mainly consisted of ginseng, were evaluated for the effects on the human cancer. One clinical and experimental study on 176 cases of digestive tract cancer revealed that Shenqi injection enhanced immune function and body weight of the patients and decreased toxic effects of chemotherapy [42]. Another randomized trial of 63 stomach cancer patients showed that Shenmai injection augmented human immune function after chemotherapy [43].
To the best of our knowledge, this is the first systemic review about the roles of reactive oxygen species (ROS) in anticancer therapy with ginsenosides (Table 1). Based on the evidence manifesting anticancer properties of ginsenosides and the roles of ROS in cancer biology, this review summarizes recent advances about the ROS-mediated anticancer effects of ginsenosides and fetches new sheds light for further design of anticancer research or more conduct of preclinical and clinical trials with this king of herb plants.
Table 1: Panax herbs and their derivate or metabolites compounds of ginsenosides that modify ROS-related effects on cancer cells*
Components [Ref] |
Cancer cells |
Concentrations |
Molecular targets |
Signaling pathway |
Effects |
|
|---|---|---|---|---|---|---|
Conc. range |
IC50 |
|||||
Ginsenoside Rh2 and Rg3 [44] |
Leukemia Jurkat cells |
0-60 μM |
35 μM (24 h) |
ROS↑, MTP↓, caspase-3/9↑, Bax/Bcl-2↑, Cyt C↑ |
Mitochondria-dependent apoptotic pathway |
Inhibits cell proliferation and induces apoptosis by stimulating the mitochondrial ROS generation |
20(S)-Ginsenoside Rh2 [45] |
Acute lymphoblastic leukemia cells |
0-60 μM |
35 μM (24 h) |
ROS↑, MTP↓, Cyt C↑, caspase-3/9↑, LC3-I↑, LC3-II↑, Atg5↓, Beclin-1↑ |
Mitochondria-dependent apoptotic pathway |
Inhibits autophagy and induces apoptosis by mitochondrial ROS generation |
Reh cells |
40 μM (24 h) |
|||||
Ginsenoside Rg3 [46] |
Lewis lung |
0-600 ng/ml |
NA |
CDK2↓, cyclin E↓, CDK 4↓, cyclinD1↓, ERK↓, p38↓, JNK↓, BAX↑, PARP↑, Bcl-2↓ |
Cell proliferation-associated pathways |
Induces apoptosis by inhibiting activation of MAPK through the regulation of intracellular ROS. |
AD-1 [47] |
lung cancer A549 and H292 cells |
0-50 μM |
6.47 μM (A549) |
p38↑, ERK1/2↑, VEGF↓, MMP-9↓, CD34↓, MDM2↓, cyclin D1↓, cyclin E↓, p21↑, p27↑, Bax↑, Bcl-2↓, G0/G1↑ |
p38 MAPK pathway |
Inhibits cell proliferation and induces apoptosis through ROS generation and p38 MAPK activation |
3.46 μM (H292) |
||||||
Compound K [48] |
Lung cancer NCI-H460 cell |
30 μg/mL |
NA |
ROS↑, MMP↓, caspase-3↑ |
Intrinsic apoptotic pathways |
Enhances γ-ray induced apoptosis by increasing intracellular ROS generation, loss of MMP and activation of caspase-3 |
Ginsenoside Rh2 [49] |
Colorectal cancer HCT-116 |
0.5-50 μM |
2.5 μM/24 h (Sodium selenite) |
ROS↓, Bax/Bcl-2↑, capase-3↑, G1↓, S ↓ |
Intrinsic apoptotic pathways |
Induces anti-proliferative activity and autophagy by depletion of ROS production |
12.5 μM (24 h) |
||||||
Ginsenoside Rh2 [50] |
Colorectal carcinoma HCT-116 and SW-480 cells |
0-60 μM |
35 μM (72 h) |
Bax↑, Bad↑, Bcl-2↓, Bcl-XL↓, ROS↑, NF-κB↑, p53↓, Cytosolic vacuolization↓ |
p53 and NF-κB signaling pathway / ROS-NF-kB pathway |
Induces apoptosis and paraptosis by activating both p53 and NF-kB through up-regulation the levels of ROS |
Protopanaxadiol [51] |
Colorectal cancer HCT-116 and SW-480cells |
10–50 μM |
35 μM (24 h) |
ROS↑, NF-κB↑ |
NF-κB pathway |
Induces paraptosis through activation of the NF-κBpathway by ROS generation |
S4h [52] |
Colorectal cancer HCT-116 and SW-480 cells |
0.2 mg/ml (HCT-116) |
NA |
ROS↑, MTP↓, Bcl-xL↑, NF-κB↑, IκB↓ |
Both the apoptosis pathway and the ROS/NF-κB mediated survival pathway |
Induces apoptosis by activating both the apoptosis pathway and the ROS/NF-kB mediated survival pathway. |
0.4 mg/ml (SW-480) |
||||||
Compound K [53] |
Colon cancer HT-29 cells |
10-40 μg/mL |
20 μg/mL |
MTP↓, caspase-3/9↑, Cyt C↑, Bax↑, Bcl-2↓, ERK↓, JNK↑, p38↑ |
Mitochondria-dependent apoptotic pathway |
Induces apoptosis by generation of ROS via mitochondria-dependent apoptotic pathway and MAPK pathway |
Compound K [54] |
Colon cancer HT-29 cells |
NA |
20 μg/ml (24h) |
PERK↑, eIF2α↑, Ca2+↑, ER stress↑, IRE-1↑, XBP-1↑, ATF-6↑, GRP-78↑, CHOP↑, caspase-12↑ |
Endoplasmic reticulum (ER) stress signaling pathway |
Induces apoptosis which is mediated by ER stress signaling pathway |
Compound K [55] |
Colon cancer HCT-116 cells |
0-50 μg/ml |
20 μg/ml (24h) |
Caspase-3/9↑, GFP-LC3↑, LC3-I↑, LC3-II↑, Bcl-2↓, Bax↑, Atg5↑, Atg6↑, Atg7↑, JNK1/2↑ |
JNK pathway |
Induces autophagy and apoptosis through ROS generation and JNK activation |
Compound K [56] |
Colon cancer HCT-116 cells |
0-50 μM |
NA |
ROS↑, Mcl-1↓, Bcl-2↓, survivin↓, XIAP↓, cFLIP↓, Bax↑, tBid↑, Cyt C↑, LC3-II↑, Atg7↑, JNK↑, ERK↓, p38↓, p53↑, DR5↑, CHOP↑ |
p53-CHOP and ROS–JNK–autophagy pathways |
Enhance TRAIL-induced apoptosis through autophagy-dependent and–independentDR5 up-regulation. |
Ginsenoside Rg3 [57] |
Colon carcinoma CT-26 cells |
10-50 μM |
NA |
Ki-67↓, VEGF↓, CD34↓, HO-1↑, NQO-1↑, Nrf2↑ |
Nrf2-mediated HO-1/NQO-1 |
Promotes the efficacy of cisplatin via preventing cisplatin-induced intracellular ROS generation. |
Korean white ginseng extract [58] |
Hepatoma HepG2 Cells |
18.6 μg/mL |
NA |
Cyt C↑, c- Jun↑, SAPK↑, MDA↓, caspases-3↑, Iκ-b↓ |
JNK–NF- κB –cytochrome c apoptotic pathway |
Induces apoptosis via an antioxidative effect and JNK–NF-κB–Cyt C apoptotic pathway |
Fermented black ginseng [59] |
Hepatoma HepG2 Cells |
10-200 μg/ml |
50 μg/ml |
ROS↓, Bax/Bcl-2↑, capase-3↑, G1↓, S ↓ |
Intrinsic apoptotic pathways |
Induces anti-proliferative activity and autophagy by depletion of ROS production |
UGB [60] |
Hepatoma HepG2 Cells |
5-75 μg/ml |
20 μg/ml (24h) |
ROS↑, caspase-3↑, Bax↑, Bcl-2↓ |
Intrinsic apoptosis pathway |
Induces apoptosis through an intrinsic apoptosis pathway |
Ginsenoside Rh2 [61] |
Hepatoma HepG2 Cells |
5-55 μM |
42.12 μM (Rh2) |
Caspase-3/9↑, Bcl-2↓, Bax↑, PARP↑, tBid↑, Cyt C↑, MMP↓ |
Mitochondrial-mediated intrinsic pathway |
Induces apoptosis through a mitochondrial mediated intrinsic pathway via generation of intracellular ROS |
20.15 μM (Rh2-O) |
||||||
Ginsenoside Rh2 [62] |
Hepatoma HepG2 Cells |
0-50 μM |
42.12 μM (24h) |
Caspase-3/9↑, cytosol Cat B↓, leupeptin (Leu) ↑, MTP↓, Bid↑, tBid↑, Cyt C↓ |
Lysosomal-mitochondrial apoptotic pathway |
Induces apoptosis through ROS accumulation and mitochondrial apoptotic pathway |
Ginsenoside Rh2 [63] |
Hepatoma HepG2 Cells |
25-50 μM |
NA |
PARP↑, ROS↑, p-p38↑, p-AMPK↑ |
AMPK signaling pathway |
Induces apoptosis through activation of AMPK-mediated ROS generation. |
Ginsenoside Rg3 and Rh2 [64] |
Hepatoma Hep3B cells |
3-50 μM |
1-30 μM (24 h) |
ROS↑, caspase-3↑, Bcl-2↓, Bax↑, Cyt C↑, MTP↓ |
Mitochondria-mediated apoptosis pathway |
Induces apoptosis through intracellular ROS production and mitochondria-mediated apoptosis pathway |
Ginsenoside Rg3 [65] |
Hepatocellular carcinoma cells |
100 μmol/L |
12.5 ng/mL (TRAIL) |
Caspase-3↑, PARP↑, eIF2α↑, CHOP↑, GRP78↑, DR5↑ |
NA |
Induces sensitization of TRAIL-induced apoptosis via CHOP-mediated DR5 up-regulation. |
Compound K [66] |
Breast cancer MCF-7 cells |
10-50 μg/ml |
35 μg/ml (48 h) |
MTP↓, AMPK↑, COX-2↓, PGE2↓ |
AMPK–COX-2 pathway |
Induces apoptosis via generation of ROS and modulation of AMPK signaling |
Ginsenoside Rg3 [67] |
Breast Cancer MDA-MB-231 Cells |
0-50 μM |
30 μM (24 h) |
ROS↑, Bax /Bcl-2↓, MTP↓, caspase-3↑, PARP↑ |
Mitochondrial death pathway |
Induces apoptosis by the activation of the mitochondrial death pathway. |
Ginsenoside Rg3 [68] |
Breast Cancer MDA-MB-231 Cells |
10-30 μM |
30 μM (24 h) |
mutant p53↓, p-ERK↓, Akt↓, NF-κB↓, Bcl-2↓, p65↓, p53↑, IκBα↓, MDM2↑, IKKβ↓, |
NF-κB pathway |
Inhibits NF-κB signaling via inactivation of ERK and Akt as well as destabilization of mutant p53. |
BG-AuNps and BG-AgNps [69] |
Breast cancer MCF-7 cells |
1-100 μg/mL |
3 μg/mL |
ROS↑ |
NA |
Induced oxidative cell damage and apoptosis through ROS generation. |
2.05 μg/mL |
||||||
12-Chloracetyl-PPD [70] |
Prostate cancer C4-2B cells |
0-30 μM |
9.85± 0.62 μM |
ROS↑, MDM2↓, p53↑, p21↑, cdc2↓, cdc25C↓, cyclin B1↓, G1 phase↓, G2/M phase↑ cdc2-Tyr15↑, cdc25-Ser216↑, caspase-3/8/9↑, PARP↑ |
G2/M cell cycle arrest |
Inhibits proliferation and induces ROS-mediated cell apoptosis via down-regulated MDM2 expression and up-regulated p53 expression. |
Ginsenoside Rh2 [71] |
Cervical carcinoma HeLa cells |
7.5 μg/mL |
NA |
ROS↑, MTP↓, Caspase-3↓, JNK1↑, SEK1↑, JNK2↑, c-Jun↑, Smac↑, Bax↑, Ca2+↑ |
ROS-JNK1 pathway |
Induces apoptosis by Ca2+ and ROS generation leading to the activation of SEK1 and JNK1 |
Compound K [72] |
Bladder cancer T24 cells |
0–25 μM |
5 μM (4 h) |
Cyt C↑, Bax↑, Bcl-2↓, p-p38MAPK↑, procaspase-3/9↑, p38↑, ROS↑, glutathione↓ |
p38MAPK pathway |
Induces apoptosis via ROS-mediated p38 MAPK pathway |
Ginsenoside F2 [73] |
Gastric carcinoma SGC7901 cells |
10-40 μg/mL |
20 μg/mL |
ROS↑, MTP↓, PARP↓, ASK-1↑, JNK↑, Bcl-2↓, Cyt C↑, Caspase-3/9↑ |
ASK-1/JNK pathway |
Induces apoptosis through ROS-mitochondria pathway and modulation of ASK-1/JNK signaling cascade |
GinsenosideRo [74] |
Esophageal cancer cells |
25-200 μM |
NA |
ROS↑, CYBB/Nox2↑, LC3B-II↑, ATG7↑, ESR2↓, NCF1↑, SQSTM1/p62↑, CSTB↓ CSTD↓, p-CHEK1↑, EGFR↑, DDIT3↑, ATM↑, ATR↑, BRCA1↑, GFP-LC3B puncta↑, Lysosomal pH↑, autophagic vacuoles↑ |
ESR2-NCF1-ROS pathway |
Suppresses autophagy via the ESR2-NCF1-ROS signal pathway and sensitized 5-fluorouracil-induced cell death through reducing CHEK1 degradation. |
Ginsenoside Rh2 [75] |
Rat C6 gliomalcells |
7.5 -10 μg/mL |
7.5 μg/mL (6 h) |
PARP↓, Bcl-XL↑ |
Caspase pathway |
Accelerates apoptosis is mediated by the generation of ROS, DNA fragmentation and the initiation of caspase pathway in a Bcl-XL-independent manner |
Ginsenoside Rg1 [76] |
Neuroblastoma SH-SY5Y cell |
5-20 μM |
10 μM |
ROS↓, JNK↑, caspase-3↑ |
JNK and caspase-3 pathway |
Prevents MPP+-induced apoptosis by inhibiting production of ROS and activating JNK pathway |
Compound K, Ginsenoside Rh2 [77] |
Astrocytoma CRT-MG cells |
0~50 mg/L |
25 mg/L (6 h) |
ROS↑, caspase-3↑, Cyt C↑, p-p38↑, Fas↑, MIP↓ |
Distinct apoptotic pathways |
Enhance Fas-mediated apoptosis in a caspase-, mitochondria-, and ROS-dependent manner. |
Ginsenoside Rb1 [78] |
Neuroblastoma SH-SY5Y cell |
10–100 μg/ml |
NA |
HO-1↑, caspase-3/9↓, PI3K↑, ROS↓, Akt↓, Nrf2↑ |
Gβ1/PI3K/Akt-Nrf2 pathway |
Prevents dopamine-induced oxidative stress through estrogen receptor-dependent Gβ1/PI3K/Akt-Nrf2 signaling pathway |
WEG [79] |
Neuroblastoma SH-SY5Y cell |
0.01-0.2 mg/mL |
NA |
ROS↑, Bcl-2↓, Bax↑, Cyt C↑, caspase-3↑ |
Mitochondria-dependent apoptotic pathway |
Prevents MPP+-induced apoptosis by alleviating oxidative stress and mitochondria-dependent apoptotic pathway. |
20(S)-ginsenoside Rg3 [80] |
GliomaU87cells |
10-100 μM |
20 μM |
ROS↑, p21↑, p16↑, p53↑, Akt↑ |
Akt and p53/p21-dependent pathways |
Induces senescence-like growth arrest by increasing ROS generation via Akt and p53/p21-dependent pathways |
Ginsenoside Rg3 [81] |
Glioblastoma U87MG cells |
10 μM |
NA |
Bcl-2↓, Bax↑, pro-caspase3↓, MEK1/2↑, ROS↑ |
MEK pathway |
Induces apoptosis through the MEK signaling pathway and ROS |
*S4h, 4 h-steamed American ginseng root extract; UGB, ultrasonication processed Panax ginseng berry extract; WEG, water extract of ginseng; Cyt C, cytochrome C; MIP, mitochondrial intermembrane potential; MPP+, 1-methyl-4-phenylpyridinium ion; MTP, mitochondrial transmembrane potentials; ↑, up-regulation; ↓, down-regulation,; NA, Not available; IC50, half maximal inhibitory concentration.
MATERIALS AND METHODS
We conducted a literature search for relevant articles in PubMed, Ovid Technologies, Excerpta Medica (EMBASE), Scopus databases, Google scholar, pubmed.cn, Web of Science SCI, Cochrane Library, Chinese National Knowledge Infrastructure (CNKI), and for published papers until May 2017 using the following search terms: “reactive oxygen species and cancer and Ginsenoside OR Ginseng” or “reactive oxygen species and cancer and Ginseng saponin”. Studies were screened for relevance. The contents of the identified articles were summarized and the current review focused on the ROS-mediated anticancer effects of ginsenosides. After removing duplicate publications and excluding information that was unrelated to ROS, we collected 38 articles about the ROS-related anticancer effects of ginsenosides.
ROLE OF ROS IN CANCER
Reactive oxygen species (ROS) are a group of highly reactive chemicals containing oxygen owing to the presence of their radicals, ions or molecules that have a single unpaired electron in their outermost shell of electrons. More than 20 types of ROS, among them, hydrogen peroxide (H2O2), superoxide anion radical (O2·−), and the highly reactive hydroxyl radical (OH·) play a crucial role in cancer [82]. Mounting evidence has demonstrated that ROS actively participate in multistep tumorigenesis, which is related to tumor initiation, transformation, progression, promotion, angiogenesis as well as metastasis [83-86]. Oxidative stress occurs when the imbalance between ROS production and elimination, which may be showed by the overproduction of ROS in the cells or the impairment of the antioxidant defense system [87], while abnormally increased ROS production accelerates irreversible damage to cellular macromolecules including proteins, lipids and DNA, contribute to deadly lesions in cell that initiates carcinogenesis [88]. Cancer cells produce increased numbers of ROS compared to their normal cell counterpart owing to the presence of their increased metabolism [89] and their elevation dependence on an antioxidant defense system [90].
The role of ROS in cancer is two-sided. First, ROS can triggers cancer initiation, progression as well as distributing through the activation and repairs of signaling pathways, which influence proliferation of cells, survival, angiogenesis and metastasis [91-93]. Second, the overproduction of ROS in malignant cells can also initiate signaling of cell death, senescence and cell cycle arrest [94, 95]. Mitochondria in malignant cell characteristically have a massive production of ROS and differ structurally and functionally from their normal cell counterpart, while oncogenic ROS enhance cancer generation via inducing genomic instability, altering gene expression, and affects in different signaling pathways [96]. In general, low concentrations of ROS are generated from oxygen through mitochondrial electron transport chain and from NADPH oxidases (NOXs) [97] that are involved for many cellular process and signal transduction. At low to moderate levels, ROS may participate in tumor generation through promotes the mutation of genomic DNA and/ or active signaling molecules. At high levels, ROS induce severe cell damage and death, especially at the beginning stage of tumor growth [98].
The increased ROS levels in cancer cells are a results of changes in mitochondrial metabolism [96, 99] that trouble cellular signaling pathways [100, 101], which are mainly mediated by the transcription factors NF-κB and STAT3, hypoxia-inducible factor-1α, kinases, growth factors, cytokines, and other enzymes [90] associated with cellular transformation, inflammation, tumor cell survival, proliferation, invasion, angiogenesis as well as metastasis of cancer [90].
ROS induce transformation of non-malignant to malignant cells through cellular DNA damage and DNA methylation [102], leading ultimately to mutations that impair oxidative phosphorylation process [96]. Some cancer cells abnormally express ROS-removing antioxidant enzymes and ROS-producing NADPH oxidases. Moreover, increasing evidence has suggested that cancer cell can produce massive ROS that triggers cancer cell death through autophagy [103, 104] and/or apoptosis [101, 105]. Owing to great amount of ROS production cancer cells showed more sensitivity than their normal counterpart [98]. Thus, increasing oxidative stress by exogenous ROS generation could be an efficient approach for the specific killing of cancer cells without causing significant toxicities to normal cells [106].
ROS-MEDIATED ANTICANCER EFFECTS OF GINSENOSIDES ON VARIOUS CANCER CELLS
Panax herbs and their derivates or metabolite compounds including ultrasonication processed P. ginseng berry extract (UPB), fermented black ginseng (FGB), Korean white ginseng (KWG), Black ginseng-gold nanoparticles (BG-AuNps) and Black ginseng-silver nanoparticles (BG-AgNps), 4 h-steamed American ginseng root extract (S4h), water extract of ginseng (WEG) contain the active ingredients, aglycone such as protopanaxadiol (PPD), 12-chloracetyl-PPD, AD-1, and glycosides such as ginsenoside Rh2, Rg3, F2, Rb1, Rg1, Ro, and compound K (Figure 1). These compounds have demonstrated anticancer activity (Table 1) with notable dose- and time-dependent inhibitory effects on the viability of leukemia, lung, colorectal, hepatoma, breast, prostate, cervical, esophageal, gastric, bladder, glioma, glioblastoma, neuroblastoma and astrocytoma cancer cells. These effects, in terms of ROS, are described in more detail for each cell type in the following sections.
ROS-mediated anticancer effects of ginsenosides on leukemia cells
Treatment of human leukemia Jurkat cell with 35μM GRh2 or GRg3 for 24 hours inhibited cell growth in a dose and time-dependent manner. It induces apoptosis by enhancing cleaved-caspase-3 and -9, ratio of Bax to Bcl-2, cytochrome c and mitochondrial ROS generation. Notably, ginsenoside Rh2 was more effective than ginsenoside Rg3 [44]. Xia et al. also reported that 20(S)-ginsenoside Rh2 inhibited human acute lymphoblastic leukemia cells growth in a dose and time-dependent manner with a half maximal inhibitory concentration (IC50) of 35μM for 24 hours, and induced mitochondria-dependent apoptosis and autophagy by mitochondrial ROS generation [45].
ROS-mediated anticancer effects of ginsenosides on lung cancer cells
Ginsenoside Rg3 inhibited the proliferation of Lewis lung carcinoma (LLC) cell via suppression of cell cycle-associated proteins, including CDK2, cyclin E, CDK4, and cyclin D1, and cell growth-associated mitogen-activated protein kinases (MAPKs), including extracellular signal-regulated kinases (ERK), p38, and c-Jun NH2-terminal kinase (JNK). The treatment with Rg3 (200 ng/ml) for 48 h induces apoptosis through activation of pro-apoptotic proteins, including BCL-2-associated X protein (BAX), cleaved poly [ADP-ribose] polymerase 1 (PARP1), and cleaved caspase-3 and decreasing of anti-apoptotic proteins, including B-cell lymphoma 2 (BCL-2) via the regulation of intracellular ROS level [46].
Another study reported that 20(R)-25-methoxyl-dammarane-3β,12β,20-triol (AD-1) induced G0/G1 cell cycle arrest in A549 and H292 lung cancer cells followed by decreasing of mouse double minute 2 homolog (MDM2), cyclin D1 and cyclin E, while increasing p21 and p27. Moreover, in vivo oral treatment of AD-1 (at a doses of 10-40 mg/kg/day for 6 weeks) dose-dependently suppressed the growth of xenograft tumors by more than 55% (40 mg/kg) without disturbing body weight of athymic nude mice, in association with decreasing the expression of vascular endothelial growth factor (VEGF), CD34, and matrix metalloproteinase-9 (MMP-9) in tumor tissue via ROS generation and p38 MAPK activation [47]. In addition, compound K was also reported to enhance γ-ray (10 Gy)-induced apoptosis in NCI-H460 human lung cancer cells in vitro by increasing intracellular ROS generation, loss of mitochondrial membrane potential (MMP) and activation of caspase-3. Compound K subcutaneously injected at dose of 30 mg/kg inhibits the growth of xenograft tumors in athymic nude mice in vivo. [48].
ROS-mediated anticancer effects of ginsenosides on colorectal cancer cells
A recent study has shown that the combination of sodium selenite (2.5 μM) and ginsenoside Rh2 (12.5 μM) could induce anti-proliferative activity in a dose- and time-dependent manner. This combination also induce autophagy mediated by cell cycle arrest at the G1 and S phase, elevation of Bax/Bcl-2ratio and capase-3 level and reduction of ROS production in human colorectal carcinoma HCT-116 Cells [49]. Ginsenoside Rh2 induced apoptosis and paraptosis-like cell death in colorectal cancer HCT-116 and SW-480 cells via the activation of both the p53 and NF-κB signaling pathway through up-regulation the levels of ROS as well as Bax and Bad, while down-regulation the levels of Bcl-2 (Figure 3) [50].
Protopanaxadiol (PPD) also induced paraptosis in colorectal cancer HCT-116 and SW-480 cells in a concentration-dependent manner through activation of the NF-κB pathway via ROS generation [51]. Li et al. [52] demonstrated that 4h-steamed American ginseng root extract (S4h) accelerated apoptosis in colorectal cancer HCT-116 and SW-480 cells, which is mediated by mitochondria damage that initiates both the apoptosis pathway and the ROS/NF-κB mediated survival pathway.
Compound K inhibited human colon cancer HT-29 cell growth in a dose-dependent manner with IC50 values being 20 μg/mL. It significantly induced ROS generation to in turn led to mitochondria-dependent and caspase-dependent apoptosis through regulating the expression of Bcl-2 and Bax, disruption of MMP, release of cytochrome c (Cyt C), elevation of caspase-3, -9, and concomitant PARP cleavage via activation of p38 MAPK and JNK pathways [53].
It was also reported that compound K inhibited human colon cancer HT-29 cell growth and induced apoptotic cell death by induction of cytosolic and mitochondrial Ca2+ overloading and initiation of endoplasmic reticulum (ER) stress signaling pathway, which is mediated by phosphorylation of protein-kinase-like endoplasmic reticulum kinase (PERK) and eukaryotic initiation factor-2α (eIF-2α) and IRE-1, splicing of ER stress-specific X-box transcription factor-1 (XBP-1), cleavage of activating transcription factor-6 (ATF-6), elevation of glucose-regulated protein-78 (GRP-78/BiP) and CCAAT/enhancer-binding protein-homologous protein (CHOP), and cleavage of caspase-12 [54].
Kim et al. [55] demonstrated that compound K (IC50=20 μg/ml) inhibited human colon cancer HCT-116 cell growth in a time and dose-dependent manner. Compound K elicited autophagy and apoptosis through the generation of ROS and initiation of JNK pathway association with elevation of autophagy-related protein (Atg, such as Atg5, Atg6, Atg7), microtubule-associated protein 1 light chain 3 (LC3), cleavage of caspase-3 and caspase-9, and modulation of Bcl-2 and Bax protein expression. The pretreated with 50 μM compound K for 24 h enhanced tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) induced apoptosis in human colon cancer HCT-116 and HT-29 cells through down-regulated Mcl-1, Bcl-2, survivin, X-linked inhibitor of apoptosis protein (XIAP), and Fas-associated death domain-like IL-1-converting enzyme-inhibitory protein (cFLIP), increased Bax, truncated Bid (tBid), Cyt C, Atg7, LC3-II, and via up-regulating autophagy-dependent and autophagy-independent death receptors 5 (DR5) (Figures 3 & 4) [56].
Oral administration of ginsenoside Rg3 (oral administration at 5 or 10 mg/kg, once daily for 15 days) significantly inhibited tumor growth of 87% and promoted the anti-neoplastic efficacy of cisplatin in mice inoculated with CT-26 colon cancer cells. This combination dose-dependently induced apoptosis and down-regulated the expression of Ki-67, VEGF and CD34 in tumor tissues. Ginsenoside Rg3 promoted the efficacy of cisplatin through inhibiting the basal level of nuclear factor erythroid 2-related factor2-(Nrf2) mediated heme oxygenase-1(HO-1)/NAD(P)H quinone oxidoreductase-1(NQO-1) expression in cancer cells [57].
ROS-mediated anticancer effects of ginsenosides on hepatoma cells
Korean white ginseng extract (KWG) reduced the cell viability in a concentration- and time-dependent manner, induced anti-oxidative effect and stress-induced apoptotic cell signaling, such as JNK and stress-activated protein kinase (SAPK) expressions, release of mitochondrial cytochrome c and activation of caspase-3 in H2O2-induced HepG2 cells [58]. Fermented black ginseng (FBG) was also showed to defend HepG2 cells against H2O2-induced oxidative stress by scavenging ROS and increasing both the activity and the expression of cellular antioxidant enzymes, such ascatalase (CAT), glutathione peroxidase (GPx) and manganese-superoxide dismutase (Mn-SOD) via the suppression of MAPK signaling pathways [59]. A recent study showed that ultrasonication processed Panax ginseng berry extract (UGB) dose-dependently decreased the cell viability of HepG2 cells with IC50 values was 20 μg/mL, and induced apoptotic cell death as associated with Bax activation and Bcl-2 inhibition, increased expression of the cleaved form of caspase-3 and intracellular ROS levels which are responsible for the intrinsic apoptosis pathway [60].
Ginsenosides Rh2 and its octyl ester derivative (Rh2-O) showed concentration- and time-dependent inhibitory effects against the proliferation of HepG2 cells with IC50 values of 20.15 and 42.12 μM for Rh2-O and Rh2, respectively. This combination also accelerated apoptosis was associated with disruption of MMP, release of Cyt C, modulation of Bcl-2 family proteins, elevation of caspase-3/-9 activation and PARP cleavage via intracellular ROS generation. Notably, Rh2-O was better antitumor activity than Rh2 plausibly due to its higher cellular uptake [61]. Ginsenosides Rh2 also induces apoptosis in HepG2 cells through the generation of ROS leads to lysosomal membrane permeabilization involving the release of cathepsins B (Cat B), cleavage of Bcl-2 family protein Bid and activation of a caspase-dependent pathway (Figure 3) [62], and through activation of AMP-activated protein kinase (AMPK) via ROS generation [63].
Park et al. [64] reported that ginsenosides (GRg3 and GRh2) dose-dependently inhibited the growth of human hepatoma Hep3B cells, and induced apoptosis via the generation of intracellular ROS, elevation of Bax/Bcl-2 ratio and initiation of mitochondria-mediated apoptosis pathway (Cyt C and caspase-3). The pretreatment of ginsenosides Rg3 enhanced TRAIL-induced apoptosis in human hepatocellular carcinoma cells via CHOP-mediated DR5 up-regulation. The combination of Rg3 (oral dose of 20 mg/kg, daily for 21 days) and TRAIL (intraperitoneal 3 times per week at dose of 3 mg/kg, for 21 days) reduced tumor volume in in vivo mouse xenograft model compared with control group along with enhancing TUNEL-positive cells and cleaved caspase-3–positive cells in tumor sections [65].
ROS-mediated anticancer effects of ginsenosides on breast cancer cells
Compound K dose-and time-dependently inhibited the cell viability of human breast cancer MCF-7 cells with IC50 being 35 μg/mL for 48 h. Compound K also accelerated apoptosis MCF-7 cells through intracellular ROS generation as well as down-regulation cyclooxygenase-2 (COX-2) expression and prostaglandin E2 (PGE2) level and the modulation of AMPK signaling [66]. Ginsenoside Rg3 was also proved to induce the apoptosis of human breast cancer MDA-MB-231 cells through up-regulating intracellular ROS generation that was characterized by a down-regulating of Bcl-2/Bax ratio, perturbation of the MTP leading to Cyt C release and initiation of the caspase-3 [67], and inhibiting constitutive activation of NF-κB signaling via probably deactivation of ERK and Akt (Protein kinase B) as well as destabilization of mutant p53 through enhancement of Mdm2 binding to mutant p53 protein, which may be responsible for inhibition of Bcl-2 expression [68]. A recent study showed that black Ginseng-silver nanoparticles (BG-AgNps) showed oxidative cell damage and obvious apoptotic activity via generation of ROS and nuclear fragmentation in MCF-7 human breast cancer cells. However, black Ginseng-gold nanoparticles (BG-AuNps) and BG-AgNps revealed non-cytotoxicity in HaCaT and MCF-7 cells [69].
ROS-mediated anticancer effects of ginsenosides on prostate and cervical cancer cells
A recent study showed that 12-Chloracetyl-PPD concentration-dependently inhibited proliferation and induces G2/M cell cycle arrest and ROS-mediated cell apoptosis in C4-2B prostate cancer cells through down-regulated MDM2 expression, and up-regulated p53 expression [70]. Ginsenoside-Rh2 was showed to induce apoptosis in cervical carcinoma HeLa cells, which is mediated by Ca2+ and ROS generation leading to stress-activated protein kinase/extracellular signal-regulated kinase kinase1 (SEK1)/JNK1 initiation and proceeds through the intrinsic pathway that includes Bax-dependent Smac (second mitochondrial activator of caspases) release via mitochondrial depolarization (Figure 3) [71].
ROS-mediated anticancer effects of ginsenosides on esophageal, gastric, and bladder carcinoma cells
Compound K was showed to dose and time-dependently inhibit the growth and induce the apoptosis of bladder cancer T24 cells through ROS-mediated p38 MAPK pathway via the release of Cyt C, initiation of procaspases-3 and -9, and the alteration of Bax/Bcl-2 proteins ratio [72]. Ginsenoside F2 also induced the apoptosis through accumulation of ROS production followed by a decrease in MTP, Cyt C release, and modulation of apoptosis signal-regulated kinase-1 (ASK-1)/JNK signaling cascade, which triggered the caspase-dependent apoptotic pathway in humor gastric carcinoma SGC7901 cells (Figure 3). Moreover, in vivo treatment of ginsenoside F2 (intragastric administration at a dose of 1.6 mg/kg everyday) significantly reduced the tumor growth by 47.14% compared with control group in xenograft mouse model [73]. A recent study showed that ginsenoside Ro inhibits autophagy by hampering with autophagosome-lysosome fusion and lysosomal proteolytic activity by enhancing lysosomal pH and reducing lysosomal cathepsins through the estrogen receptor 2 (ESR2)-neutrophil cytosolic factor 1 (NCF1)-ROS signaling pathway (Figure 4) and thereby sensitized esophageal cancer cells to5-fluorouracil-induced cell death via reduces CHEK1 (checkpoint kinase 1) degradation, increases CHEK1-mediated DNA damage checkpoint arrest [74].
ROS-mediated anticancer effects of ginsenosides on glioma, glioblastoma, astrocytoma and neuroblastoma cells
It was first recognized that ginsenoside Rh2 inhibited rat C6 glioma cells growth in a dose-dependent manner and accelerated apoptosis mediated by the generation of ROS, DNA fragmentation and the initiation of caspase pathway in a Bcl-XL-independent manner [75]. Ginsenoside Rg1 prevents 1-methyl-4-phenylpyridinium ion (MPP+)-induced apoptosis which may be attributed to its antioxidant and anti-apoptotic properties via down-regulating ROS production, activation of JNK and caspase-3 pathway [76]. Ginsenosides CK and Rh2 was showed to accelerate Fas-mediated apoptosis in human astrocytoma CRT-MG cells in a caspase-, mitochondria-, and ROS-dependent manner through distinct apoptotic signaling pathways (Figure 3) [77].
Hwang and Jeong [78] demonstrated that ginsenoside Rb1 prevents 6-hydroxydopamine-induced cell death in human neuroblastoma SH-SY5Y cells through up-regulating heme oxygenase-1 expression, Nrf2 nuclear translocation via anon-genomic, estrogen receptor-mediated Gβ1/PI3K/Akt pathway, thus defending cells from oxidative stress.
The water extract of ginseng (WEG) prevented MPP+-induced apoptosis in SH-SY5Y human neuroblastoma cells probably through generation of ROS inhibition and the suppression of mitochondria-dependent apoptotic pathway via up-regulating Bax expression, cytosolic Cyt C and cleaved caspase-3, while down-regulating Bcl-2 expression [79].
Treatment with 20(S)-ginsenoside Rg3 at a ≥10 μM concentration showed a dose-dependent inhibitory effect on U87 human glioma cells proliferation for 3 days, however, chronic treatment at a 20 μM concentration completely inhibited cell proliferation for at least nine days. It also accelerates senescence-like growth arrest via increasing generation of intracellular ROS through Akt initiation and p53/p21-dependent signaling pathways [80]. In another study, treatment of human glioblastoma U87MG cells with 10μM ginsenoside Rg3 for 24 h exhibit a dose-and time-dependent inhibitory effect on their proliferation and elicits apoptosis via up-regulating Bax, while down-regulating Bcl-2 and pro-caspase 3 expression through initiation of ROS by antioxidant enzyme system and MEK signaling pathway [81].
CONCLUSIONS AND FUTURE PERSPECTIVES
A renewed interest issues in the study of complementary or alternative medicine therapy and less toxic cures for the treatment of different diseases, including cancer. One of the major goals of the treatment of cancers is maximizing efficacy and minimizing adverse effects. Ginsenosides may be a potential complementary or alternative therapy for various cancer patients. We found the potential utility of Panax herbs and their derivates, or their active constituents including aglycone such as PPD, 12-chloracetyl-PPD, AD-1, and glycosides such as ginsenoside Rh2, Rg3, F2, Rb1, Rg1, Ro and compound K (Figure 1). In the sheds light of growing literature, ginsenosides possess ample potential for providing diverse mechanisms for treating various cancers (Table 1) that’s why it may be tough for cancer cells to become resistance against ginsenosides. Moreover, the properties to kill tumor cells make ginsenosides attractive candidates for drug development. High-throughput expression analysis will help to identify molecular mechanisms and effects of different ginsenosides. The ROS-mediated anticancer effects of different ginsenosides depends on the specific type of various cancer cells involved (Table 1). Some ginsenosides may up-regulate Fas, Bid, tBid, Bax, Bad, p53, p21, p27, p16, Caspase-3, Caspase-8, Caspase-9, ER (endoplasmic reticulum) stress, DR5, CHOP, ATF-6, Smac, PERK, HO-1, EGFR, Beclin 1, Atg7, Atg5, LC3-I, LC3-II, AMP-activated protein kinase, and activate MEK, ASK-1/JNK, ESR2-NCF1-ROS, ER-dependent PI3K/Akt/ Nrf2, P53-CHOP, and ROS-JNK-autophagy signaling pathways. Conversely, these compounds can down-regulate Ki-67, cyclinD1, cyclin E, CDK2/4, cdc25c, cyclin B1, survivin, cFLIP, Mcl-1, XIAP, Bcl-2, cytosol Cat B, Cytosolic vacuolization, CD34, VEGF, COX-2, PGE2, MMP-9 and inhibit thePI3K/Akt signaling pathway (Figure 2). Overall, ginsenosides can inhibit cell proliferation and induce apoptosis, paraptosis or autophagy in vitro and in vivo by generating ROS in various human cancers. Collectively, these effects suppress cancer cell proliferation via arresting cell cycle progression, inducing cancer cell apoptosis, paraptosis and/or autophagy, and elicit anti-angiogenic and anti-metastatic effects (Figures 3 & 4). Although ROS-mediated effects of ginsenosides exhibit a clear anticancer activity in cancer cell lines, xenograft tumor models or in human clinical trials, it will be further necessary to investigate rigorous multicenter human studies and detailed large scale well-designed cohort clinical trials to confirm the exact effectiveness as an anticancer agents in human patients, thus improving treatment of cancer in near future.
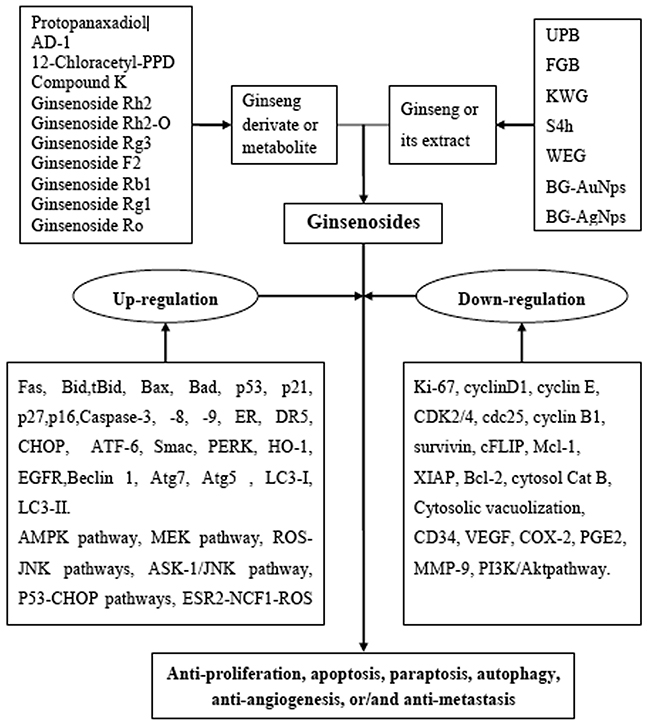
Figure 2: Schematic Diagram of Reactive Oxygen Species (ROS)-Related Anticancer Effects Mediated by Ginsenosides. Up-regulation: Fas, Bid, tBid, Bax, Bad, p53, p21, p27, p16, caspase-3, caspase-8, caspase-9, ER (endoplasmic reticulum) stress, DR5, CHOP, ATF-6, Smac, PERK, HO-1, EGFR, Beclin 1, Atg7, Atg5, LC3-I, LC3-II, AMP-activated protein kinase (AMPK), MEK signaling pathway, ASK-1/JNK signaling pathway, estrogen receptor 2 (ESR2)-neutrophil cytosolic factor 1 (NCF1)-ROS signaling pathway, ER (estrogen receptor)-dependent PI3K/Akt/ Nrf2 pathway, P53-CHOP pathways, and ROS–JNK–autophagy pathways. Down-regulation: Ki-67, cyclinD1, cyclin E, CDK2/4, cdc25, cyclin B1, survivin, cFLIP, Mcl-1, XIAP, Bcl-2, cytosol Cat B, cytosolic vacuolization, CD34, VEGF, COX-2, PGE2, MMP-9 and PI3K/Akt signaling pathway.
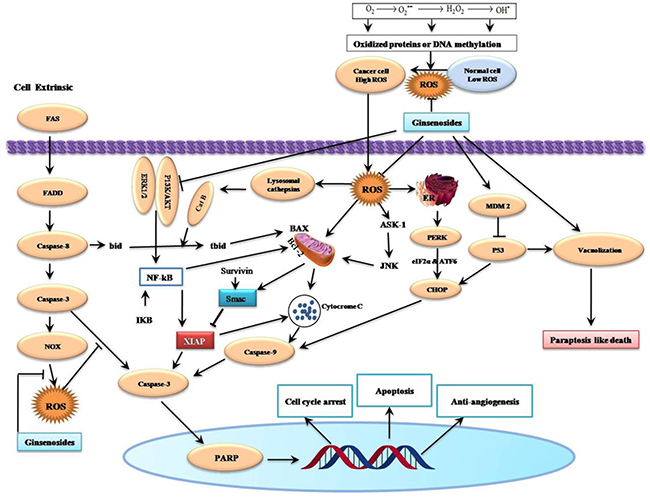
Figure 3: Schematic Diagram of Effects of Ginsenosides on Reactive Oxygen Species-Related Cell Cycle Arrest, Apoptosis, Paraptosis and Anti-angiogenesis of Cancer Cells. Ginsenosides can induce intrinsic cell death and they augment Fas-induced extrinsic cell death by ROS suppression. Production of ROS is the vital issues of ginsenosides mediated apoptosis from oxidized protein or DNA methylation. Generally, ROS are generated either through mitochondrial electron transport chain or from NADPH oxidases (NOXs). Aberrant mitochondrial functions are associated with the release of cytochrome c resulting in the activation of caspase-3 and PARP, ultimately leading to apoptosis and cell death. The final effects are cell cycle arrest, induction of apoptosis, paraptosis and anti-angiogenic activity. →, Activation; ┤, Inhibition.
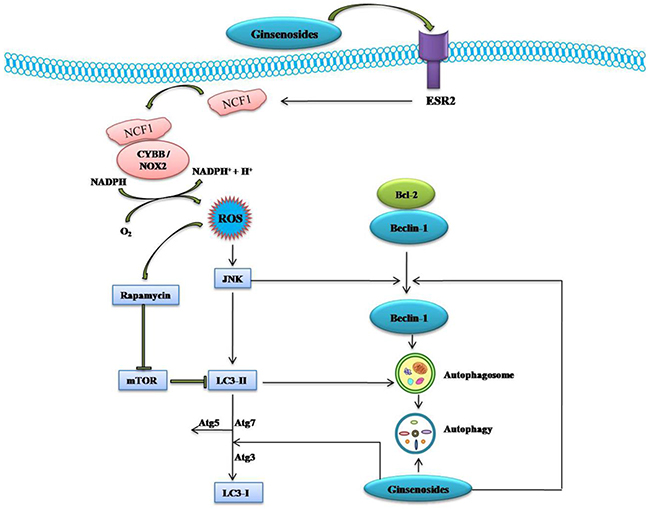
Figure 4: Schematic Diagram of Effects of Ginsenosides on ESR2-NCF1-ROS and ROS-JNK-Autophagy Pathway of Cancer Cells. Ginsenoside activate estrogen receptor 2 (ESR2)-neutrophil cytosolic factor 1 (NCF1)-ROS signaling pathway. ROS then activate JNK which in turns phosphorylate Bcl-2 and release Beclin 1 and induce autophagy through up-regulation of ATG5, Atg7, LC3-I and LC3-II. →, Activation; ┤, Inhibition.
Abbreviations
Complementary and alternative medicine, CAM; protopanaxadiol, PPD; protopanaxatriol, PPT; reactive oxygen species, ROS; Panax ginseng berry extract, UPB; fermented black ginseng, FGB; Korean white ginseng, KWG; Black ginseng-gold nanoparticles, BG-AuNps; Black ginseng-silver nanoparticles, BG-AgNps; 4 h-steamed American ginseng root extract, S4h; water extract of ginseng, WEG; 20(R)-25-methoxyldammarane-3β,12β,20-triol, AD-1; 12β-O-(L-Chloracetyl)-dammar-20(22)-ene-3β,25-diol, 12-Chloracetyl-PPD; mitogen-activated protein kinases, MAPK; mouse double minute 2 homolog, MDM2; vascular endothelial growth factor, VEGF; matrix metalloproteinase-9, MMP-9; nicotinamide adeninedinucleotide phosphate (NADPH) oxidase enzymes, NOX; mitochondrial transmembrane potential, MTP; mitochondrial membrane potential, MMP; endoplasmic reticulum, ER; c-Jun NH2-terminal kinase, JNK; mammalian target of rapamycin, mTOR; tumor necrosis factor (TNF)-related apoptosis-inducing ligand, TRAIL; X-linked inhibitor of apoptosis protein, XIAP; Fas-associated death domain-like IL-1-converting enzyme-inhibitory protein, cFLIP; truncated Bid, tBid, cytochrome c, Cyt C; C/EBP homologous protein, CHOP; death receptors 5, DR5; nuclear factor erythroid 2-related factor 2, Nrf2; heme oxygenase-1, HO-1; NAD(P)H quinone oxidoreductase-1, NQO-1; cathepsins B, Cat B; AMP-activated protein kinase, AMPK; cyclooxygenase-2, COX-2; prostaglandin E2, PGE2; extracellular signal-regulated kinase, ERK; protein kinase B, Akt; stress-activated protein kinase/extracellular signal-regulated kinase kinase1, SEK1; second mitochondrial activator of caspases, Smac; apoptosis signal-regulated kinase-1, ASK-1; estrogen receptor 2, ESR2; neutrophil cytosolic factor 1, NCF1; checkpoint kinase 1, CHEK1; Activating transcription factor 6, ATF6; poly (ADP-ribose) polymerase, PARP; epidermal growth factor receptor, EGFR; autophagy-related protein, Atg; autophagy-related protein 6, Beclin-1; microtubule-associated protein 1 light chain 3, LC3; Fas-associated death domain, FADD; PKR-like endoplasmic reticulum kinase, PERK; Reactive oxygen species, ROS; compound K, CK; phosphatidylinositol 3-kinase, PI3K.
CONFLICTS OF INTEREST
The authors declare no potential conflicts of interest.
FUNDING
This work was supported by Grant-in-Aid from the National Natural Science Foundation of China (Nos. 31272597, 31472229 and 31772783), the Zhejiang Provincial Natural Science Foundation of China (LZ13C180001), the National Major Scientific and Technological Special Project for “Significant New Drugs Development (2014ZX09102041-009), the National Key R&D Program of China (2017YFD0501505).
REFERENCES
1. Liu H, Dong Y, Gao Y, Du Z, Wang Y, Cheng P, Chen A, Huang H. The fascinating effects of baicalein on cancer: a review. Int J Mol Sci. 2016; 17:1681.
2. Bandhavkar S. Cancer stem cells: a metastasizing menace!. Cancer Med. 2016; 5:649–55.
3. Neergheen VS, Bahorun T, Taylor EW, Jen LS, Aruoma OI. Targeting specific cell signaling transduction pathways by dietary and medicinal phytochemicals in cancer chemoprevention. Toxicology. 2010; 278:229–41.
4. Wong AS, Che CM, Leung KW. Recent advances in ginseng as cancer therapeutics: a functional and mechanistic overview. Nat Prod Rep. 2015; 32:256–72.
5. Newman DJ, Cragg GM. Natural products as sources of new drugs from 1981 to 2014. J Nat Prod. 2016; 79:629–61.
6. Amin AR, Kucuk O, Khuri FR, Shin DM. Perspectives for cancer prevention with natural compounds. J Clin Oncol. 2009; 27:2712–25.
7. Weng CJ, Yen GC. Chemopreventive effects of dietary phytochemicals against cancer invasion and metastasis: phenolic acids, monophenol, polyphenol, and their derivatives. Cancer Treat Rev. 2012; 38:76–87.
8. Millimouno FM, Dong J, Yang L, Li J, Li X. Targeting apoptosis pathways in cancer and perspectives with natural compounds from mother nature. Cancer Prev Res (Phila). 2014; 7:1081–107
9. Harris PE, Cooper KL, Relton C, Thomas KJ. Prevalence of complementary and alternative medicine (CAM) use by the general population: a systematic review and update. Int J Clin Pract. 2012; 66:924–39.
10. Kim YJ, Zhang D, Yang DC. Biosynthesis and biotechnological production of ginsenosides. Biotechnol Adv. 2015; 33:717–35.
11. Kim JH. Cardiovascular diseases and Panax ginseng: a review on molecular mechanisms and medical applications. J Ginseng Res. 2012; 36:16–26.
12. Yang WZ, Hu Y, Wu WY, Ye M, Guo DA. Saponins in the genus Panax L. (Araliaceae): a systematic review of their chemical diversity. Phytochemistry. 2014; 106:7–24.
13. Wang CZ, Anderson S, Du W, He TC, Yuan CS. Red ginseng and cancer treatment. Chin J Nat Med. 2016; 14:7–16.
14. Qi LW, Wang CZ, Yuan CS. Isolation and analysis of ginseng: advances and challenges. Nat Prod Rep. 2011; 28:467–95.
15. Shibata S, Tanaka O, Soma K, Ando T, Iida Y, Nakamura H. Studies on saponins and sapogenins of ginseng the structure of panaxatriol. Tetrahedron Lett. 1965; 42:207–13.
16. Christensen LP. Ginsenosides chemistry, biosynthesis, analysis, and potential health effects. Adv Food Nutr Res. 2009; 55:1–99.
17. Nag SA, Qin JJ, Wang W, Wang MH, Wang H, Zhang R. Ginsenosides as anticancer agents: in vitro and in vivo activities, structure-activity relationships, and molecular mechanisms of action. Front Pharmacol. 2012; 3:25.
18. Quan K, Liu Q, Wan JY, Zhao YJ, Guo RZ, Alolga RN, Li P, Qi LW. Rapid preparation of rare ginsenosides by acid transformation and their structure-activity relationships against cancer cells. Sci Rep. 2015; 5:8598.
19. Dharmananda S. The nature of ginseng: traditional use, modern research, and the question of dosage. HerbalGram. 2002; 54:34–51.
20. Hu C, Kitts DD. Free radical scavenging capacity as related to antioxidant activity and ginsenoside composition of asian and north american ginseng extracts. J Am Oil Chem Soc. 2001; 78:249–55.
21. Chae S, Kang KA, Youn U, Park JS, Hyun JW. A comparative study of the potential antioxidant activities of ginsenosides. J Food Biochem. 2010; 34:31–43.
22. Patel S, Rauf A. Adaptogenic herb ginseng (Panax) as medical food: status quo and future prospects. Biomed Pharmacother. 2017; 85:120–7.
23. Jung JH, Kang IG, Kim DY, Hwang YJ, Kim ST. The effect of korean red ginseng on allergic inflammation in a murine model of allergic rhinitis. J Ginseng Res. 2013; 37:167–75.
24. Rhee MY, Kim YS, Bae JH, Nah DY, Kim YK, Lee MM, Kim HY. Effect of korean red ginseng on arterial stiffness in subjects with hypertension. J Altern Complement Med. 2011; 17:45–9.
25. Choi JS, Chun KS, Kundu J, Kundu JK. Biochemical basis of cancer chemo-prevention and/or chemotherapy with ginsenosides (Review). Int J Mol Med. 2013; 32:1227–38.
26. Yun TK, Choi SY. A case-control study of ginseng intake and cancer. Int J Epidemiol. 1990; 19:871–6.
27. Yun TK, Choi SY. Non-organ specific cancer prevention of ginseng: a prospective study in Korea. Int J Epidemiol. 1998; 27:359–64.
28. Yun TK, Choi SY, Yun HY. Epidemiological study on cancer prevention by ginseng: are all kinds of cancers preventable by ginseng? J Korean Med Sci. 2001; 16:S19–27.
29. Cui Y, Shu XO, Gao YT, Cai H, Tao MH, Zheng W. Association of ginseng use with survival and quality of life among breast cancer patients. Am J Epidemiol. 2006; 163:645–53.
30. Wang XS, Wu Q, Liang L. Combined shenyi-jiao nang and chemotherapy in treatment of breast cancer: a systematic review. Pract J Clin Med. 2012; 9:192–5.
31. Suh SO, Kroh M, Kim NR, Joh YG, Cho MY. Effects of red ginseng upon postoperative immunity and survival in patients with stage III gastric cancer. Am J Chin Med. 2002; 30:483–94.
32. Kim JH, Park CY, Lee SJ. Effects of sun ginseng on subjective quality of life in cancer patients: a double-blind, placebo-controlled pilot trial. J Clin Pharm Ther. 2006; 31:331–4.
33. Barton DL, Soori GS, Bauer BA, Sloan JA, Johnson PA, Figueras C, Duane S, Mattar B, Liu H, Atherton PJ, Christensen B, Loprinzi CL. Pilot study of Panax quinquefolius (American ginseng) to improve cancer-related fatigue: a randomized, double-blind, dose-finding evaluation: NCCTG trial N03CA. Support Care Cancer. 2010; 18:179–87.
34. Yun TK, Zheng S, Choi SY, Cai SR, Lee YS, Liu XY, Cho KJ, Park KY. Non-organ-specific preventive effect of long-term administration of Korean red ginseng extract on incidence of human cancers. J Med Food. 2010; 13:489–94.
35. Jin X, Che DB, Zhang ZH, Yan HM, Jia ZY, Jia XB. Ginseng consumption and risk of cancer: a meta-analysis. J Ginseng Res. 2016; 403:269–77.
36. Kim HS, Kim MK, Lee M, Kwon BS, Suh DH, Song YS. Effect of red ginseng on genotoxicity and health-Related quality of life after adjuvant chemotherapy in patients with epithelial ovarian cancer: a randomized, double blind, placebo-controlled trial. Nutrients. 2017; 9:772.
37. Kamangar F, Gao YT, Shu XO, Kahkeshani K, Ji BT, Yang G, Li HL, Rothman N, Chow WH, Zheng W. Ginseng intake and gastric cancer risk in the shanghai women’s health study cohort. Cancer Epidemiol Biomarkers Prev. 2007; 16:629–30.
38. Rebbeck TR, Troxel AB, Norman S, Bunin GR, DeMichele A, Baumgarten M, Berlin M, Schinnar R, Strom BL. A retrospective case-control study of the use of hormone-related supplements and association with breast cancer. Int J Cancer. 2007; 120:1523–8.
39. Cheng Y, Hua HQ. Clinical research progress in anti-tumor effects of ginesenoside Rg3. Medical Recap. 2015; 21:2938–40.
40. Lu P, Su W, Miao ZH, Niu HR, Liu J, Hua QL. Effect and mechanism of ginsenoside Rg3 on postoperative life span of patients with non-small cell lung cancer. Chin J Integr Med. 2008; 14:33–6.
41. Huang JY, Sun Y, Fan QX, Zhang YQ. Efficacy of shenyi capsule combined with gemcitabine plus cisplatin in treatment of advanced esophageal cancer: a randomized controlled trial. J Chin Integr Med. 2009; 7:1047–51.
42. Li NQ. Clinical and experimental study on shen-qi injection with chemotherapy in the treatment of malignant tumor of digestive tract. Chin J Integr Tradit West Med. 1992; 12:588–92.
43. Lin SY, Liu LM, Wu LC. Effects of Shenmai injection on immune function in stomach cancer patients after chemotherapy. Chin J Integr Tradit West Med.1995; 15:451–3.
44. Xia T, Wang YN, Zhou CX, Wu LM, Liu Y, Zeng QH, Zhang XL, Yao JH, Wang M, Fang JP. Ginsenoside Rh2 and Rg3 inhibit cell proliferation and induce apoptosis by increasing mitochondrial reactive oxygen species in human leukemia Jurkat cells. Mol Med Rep. 2017; 15:3591–8.
45. Xia T, Wang J, Wang Y, Wang Y, Cai J, Wang M, Chen Q, Song J, Yu Z, Huang W, Fang J. Inhibition of autophagy potentiates anticancer property of 20(S)-ginsenoside Rh2 by promoting mitochondria-dependent apoptosis in human acute lymphoblastic leukaemia cells. Oncotarget. 2016; 7:27336–49. https://doi.org/10.18632/oncotarget.8285.
46. Sun HY, Lee JH, Han YS, Yoon YM, Yun CW, Kim JH, Song YS, Lee SH. Pivotal roles of ginsenoside Rg3 in tumor apoptosis through regulation of reactive oxygen species. Anticancer Res. 2016; 36:4647–54.
47. Zhang LH, Jia YL, Lin XX, Zhang HQ, Dong XW, Zhao JM, Shen J, Shen HJ, Li FF, Yan XF, Li W, Zhao YQ, Xie QM. AD-1, a novel ginsenoside derivative, shows anti-lung cancer activity via activation of p38 MAPK pathway and generation of reactive oxygen species. Biochim Biophys Acta. 2013; 1830:4148–59.
48. Chae S, Kang KA, Chang WY, Kim MJ, Lee SJ, Lee YS, Kim HS, Kim DH, Hyun JW. Effect of compound K, a metabolite of ginseng saponin, combined with gamma-ray radiation in human lung cancer cells in vitro and in vivo. J Agric Food Chem. 2009; 57:5777–82.
49. Zhu C, Liu F, Qian W, Zhang T, Li F. Combined effect of sodium selenite and ginsenoside Rh2 on HCT116 human colorectal carcinoma cells. Arch Iran Med. 2016; 19:23–9.
50. Li B, Zhao J, Wang CZ, Searle J, He TC, Yuan CS, Du W. Ginsenoside Rh2 induces apoptosis and paraptosis-like cell death in colorectal cancer cells through activation of p53. Cancer Lett. 2011; 301:185–92.
51. Wang CZ, Li B, Wen XD, Zhang Z, Yu C, Calway TD, He TC, Du W, Yuan CS. Paraptosis and NF-κB activation are associated with protopanaxadiol-induced cancer chemoprevention. BMC Complement Altern Med. 2013; 13:2.
52. Li B, Wang CZ, He TC, Yuan CS, Du W. Antioxidants potentiate american ginseng-induced killing of colorectal cancer cells. Cancer Lett. 2010; 289:62–70.
53. Lee IK, Kang KA, Lim CM, Kim KC, Kim HS, Kim DH, Kim BJ, Chang WY, Choi JH, Hyun JW. Compound K, a metabolite of ginseng saponin, induces mitochondria-dependent and caspase-dependent apoptosis via the generation of reactive oxygen species in human colon cancer cells. Int J Mol Sci. 2010; 11:4916–31.
54. Zhang R, Chung Y, Kim HS, Kim DH, Kim HS, Chang WY, Hyun JW. 20-O-(β-D-glucopyranosyl)-20(S)-protopanaxadiol induces apoptosis via induction of endoplasmic reticulum stress in human colon cancer cells. Oncol Rep. 2013; 29:1365–70.
55. Kim AD, Kang KA, Kim HS, Kim DH, Choi YH, Lee SJ, Kim HS, Hyun JW. A ginseng metabolite, compound K, induces autophagy and apoptosis via generation of reactive oxygen species and activation of JNK in human colon cancer cells. Cell Death Dis. 2013; 4:e750.
56. Chen L, Meng Y, Sun Q, Zhang Z, Guo X, Sheng X, Tai G, Cheng H, Zhou Y. Ginsenoside compound K sensitizes human colon cancer cells to TRAIL-induced apoptosis via autophagy-dependent and -independent DR5 upregulation. Cell Death Dis. 2016; 7:e2334.
57. Lee CK, Park KK, Chung AS, Chung WY. Ginsenoside Rg3 enhances the chemosensitivity of tumors to cisplatin by reducing the basal level of nuclear factor erythroid 2-related factor 2-mediated heme oxygenase-1/NAD(P)H quinone oxidoreductase-1 and prevents normal tissue damage by scavenging cisplatin-induced intracellular reactive oxygen species. Food Chem Toxicol. 2012; 50:2565–74.
58. Lee M, Sorn S, Baek S, Jang S, Kim S. Antioxidant and apoptotic effects of korean white ginseng extracted with the same ratio of protopanaxadiol and protopanaxatriols saponins in human hepatoma HepG2 cells. Ann N Y Acad Sci. 2009; 1171:217–27.
59. Bak MJ, Jeong WS, Kim KB. Detoxifying effect of fermented black ginseng on H2O2-induced oxidative stress in HepG2 cells. Int J Mol Med. 2014; 34:1516–22.
60. Jung H, Bae J, Ko SK, Sohn UD. Ultrasonication processed Panax ginseng berry extract induces apoptosis through an intrinsic apoptosis pathway in HepG2 cells. Arch Pharm Res. 2016; 39:855–62.
61. Chen F, Deng ZY, Zhang B, Xiong ZX, Zheng SL, Tan CL, Hu JN. Esterification of ginsenoside Rh2 enhanced its cellular uptake and antitumor activity in human HepG2 cells. J Agric Food Chem. 2016; 64:253–61.
62. Chen F, Deng Z, Xiong Z, Zhang B, Yang J, Hu J. A ROS-mediated lysosomal-mitochondrial pathway is induced by ginsenoside Rh2 in hepatoma HepG2 cells. Food Funct. 2015; 6:3828–37.
63. Kim MJ, Yun H, Kim DH, Kang I, Choe W, Kim SS, Ha J. AMP-activated protein kinase determines apoptotic sensitivity of cancer cells to ginsenoside-Rh2. J Ginseng Res. 2014; 38:16–21.
64. Park HM, Kim SJ, Kim JS, Kang HS. Reactive oxygen species mediated ginsenoside Rg3- and Rh2-induced apoptosis in hepatoma cells through mitochondrial signaling pathways. Food Chem Toxicol. 2012; 50:2736–41.
65. Lee JY, Jung KH, Morgan MJ, Kang YR, Lee HS, Koo GB, Hong SS, Kwon SW, Kim YS. Sensitization of TRAIL-induced cell death by 20(S)-ginsenoside Rg3 via CHOP-mediated DR5 upregulation in human hepatocellular carcinoma cells. Mol Cancer Ther. 2013; 12:274–85.
66. Kim AD, Kang KA, Zhang R, Lim CM, Kim HS, Kim DH, Jeon YJ, Lee CH, Park J, Chang WY, Hyun JW. Ginseng saponin metabolite induces apoptosis in MCF-7 breast cancer cells through the modulation of AMP-activated protein kinase. Environ Toxicol Pharmacol. 2010; 30:134–40.
67. Kim BM, Kim DH, Park JH, Na HK, Surh YJ. Ginsenoside Rg3 induces apoptosis of human breast cancer (MDA-MB-231) Cells. J Cancer Prev. 2013; 18:177–85.
68. Kim BM, Kim DH, Park JH, Surh YJ, Na HK. Ginsenoside Rg3 inhibits constitutive activation of NF-κB signaling in human breast cancer (MDA-MB-231) cells: ERK and Akt as potential upstream targets. J Cancer Prev. 2014; 19:23–30.
69. Wang D, Markus J, Kim YJ, Wang C, Jiménez Pérez ZE, Ahn S, Aceituno VC, Mathiyalagan R, Yang DC. Coalescence of functional gold and monodisperse silver nanoparticles mediated by black Panax ginseng Meyer root extract. Int J Nanomedicine. 2016; 11:6621–34.
70. Wang X, Sun YY, Zhao C, Qu FZ, Zhao YQ. 12-Chloracetyl-PPD, a novel dammarane derivative, shows anti-cancer activity via delay the progression of cell cycle G2/M phase and reactive oxygen species-mediate cell apoptosis. Eur J Pharmacol. 2017; 798:49–56.
71. Ham YM, Lim JH, Na HK, Choi JS, Park BD, Yim H, Lee SK. Ginsenoside-Rh2-induced mitochondrial depolarization and apoptosis are associated with reactive oxygen species- and Ca2+-mediated c-Jun NH2-terminal kinase 1 activation in HeLa cells. J Pharmacol Exp Ther. 2006; 319:1276–85.
72. Wang H, Jiang D, Liu J, Ye S, Xiao S, Wang W, Sun Z, Xie Y, Wang J. Compound K induces apoptosis of bladder cancer T24 cells via reactive oxygen species-mediated p38 MAPK pathway. Cancer Biother Radiopharm. 2013; 28:607–14.
73. Mao Q, Zhang PH, Wang Q, Li SL. Ginsenoside F2 induces apoptosis in humor gastric carcinoma cells through reactive oxygen species-mitochondria pathway and modulation of ASK-1/JNK signaling cascade in vitro and in vivo. Phytomedicine. 2014; 21:515–22.
74. Zheng K, Li Y, Wang S, Wang X, Liao C, Hu X, Fan L, Kang Q, Zeng Y, Wu X, Wu H, Zhang J, Wang Y, He Z. Inhibition of autophagosome-lysosome fusion by ginsenoside Ro via the ESR2-NCF1-ROS pathway sensitizes esophageal cancer cells to 5-fluorouracil-induced cell death via the CHEK1-mediated DNA damage checkpoint. Autophagy. 2016; 12:1593–613.
75. Kim HE, Oh JH, Lee SK, Oh YJ. Ginsenoside Rh-2 induces apoptotic cell death in rat C6 glioma via a reactive oxygen- and caspase-dependent but Bcl-X(L)-independent pathway. Life Sci. 1999; 65:PL33–40.
76. Chen XC, Fang F, Zhu YG, Chen LM, Zhou YC, Chen Y. Protective effect of ginsenoside Rg1 on MPP+-induced apoptosis in SHSY5Y cells. J Neural Transm (Vienna). 2003; 110:835–45.
77. Choi K, Choi C. Proapoptotic ginsenosides compound K and Rh2 enhance Fas-induced cell death of human astrocytoma cells through distinct apoptotic signaling pathways. Cancer Res Treat. 2009; 41:36–44.
78. Hwang YP, Jeong HG. Ginsenoside Rb1 protects against 6-hydroxy- dopamine-induced oxidative stress by increasing heme oxygenase-1 expression through an estrogen receptor-related PI3K/Akt/Nrf2-dependent pathway in human dopaminergic cells. Toxicol Appl Pharmacol. 2010; 242:18–28.
79. Hu S, Han R, Mak S, Han Y. Protection against 1-methyl-4-phenylpyridinium ion (MPP+)-induced apoptosis by water extract of ginseng (Panax ginseng C.A. Meyer) in SH-SY5Y cells. J Ethnopharmacol. 2011; 135:34–42.
80. Sin S, Kim SY, Kim SS. Chronic treatment with ginsenoside Rg3 induces Akt-dependent senescence in human glioma cells. Int J Oncol. 2012; 41:1669–74.
81. Choi YJ, Lee HJ, Kang DW, Han IH, Choi BK, Cho WH. Ginsenoside Rg3 induces apoptosis in the U87MG human glioblastoma cell line through the MEK signaling pathway and reactive oxygen species. Oncol Rep. 2013; 30:1362–70.
82. Liou GY, Storz P. Reactive oxygen species in cancer. Free Radic Res. 2010; 44:479–96.
83. Sosa V, Moliné T, Somoza R, Paciucci R, Kondoh HL, Leonart ME. Oxidative stress and cancer: an overview. Ageing Res Rev. 2013; 12:376–90.
84. Kryston TB, Georgiev AB, Pissis P, Georgakilas AG. Role of oxidative stress and DNA damage in human carcinogenesis. Mutat Res. 2011; 711:193–201.
85. Wu WS. The signaling mechanism of ROS in tumor progression. Cancer Metastasis Rev. 2006; 25:695–705.
86. Hung YC, Pan TL, Hu WL. Roles of reactive oxygen species in anticancer therapy with Salvia miltiorrhiza Bunge. Oxid Med Cell Longev. 2016; 2016:5293284.
87. Schieber M, Chandel NS. ROS function in redox signaling and oxidative stress. Curr Biol. 2014; 24:R453–62.
88. Klaunig JE, Xu Y, Isenberg JS, Bachowski S, Kolaja KL, Jiang J, Stevenson DE, Walborg EF Jr. The role of oxidative stress in chemical carcinogenesis. Environ Health Perspect. 1998; 106:289–95.
89. Cairns RA, Harris IS, Mak TW. Regulation of cancer cell metabolism. Nat Rev Cancer. 2011; 11:85–95.
90. Prasad S, Gupta SC, Tyagi AK. Reactive oxygen species (ROS) and cancer: role of antioxidative nutraceuticals. Cancer Lett. 2017; 387:95–105.
91. Weinberg F, Chandel NS. Reactive oxygen species-dependent signaling regulates cancer. Cell Mol Life Sci. 2009; 66:3663–73.
92. Weinberg F, Hamanaka R, Wheaton WW, Weinberg S, Joseph J, Lopez M, Kalyanaraman B, Mutlu GM, Budinger GR, Chandel NS. Mitochondrial metabolism and ROS generation are essential for Kras-mediated tumorigenicity. Proc Natl Acad Sci U S A. 2010; 107:8788–93.
93. Wallace DC. Mitochondria and cancer. Nat Rev Cancer. 2012; 12:685–98.
94. Ichijo H, Nishida E, Irie K, ten Dijke P, Saitoh M, Moriguchi T, Takagi M, Matsumoto K, Miyazono K, Gotoh Y. Induction of apoptosis by ASK1, a mammalian MAPKKK that activates SAPK/JNK and p38 signaling pathways. Science. 1997; 275:90–4.
95. Moon DO, Kim MO, Choi YH, Hyun JW, Chang WY, Kim GY. Butein induces G(2)/M phase arrest and apoptosis in human hepatoma cancer cells through ROS generation. Cancer Lett. 2010; 288:204–13.
96. Yang Y, Karakhanova S, Hartwig W, D’Haese JG, Philippov PP, Werner J, Bazhin AV. Mitochondria and mitochondrial ROS in cancer: novel targets for anticancer therapy. J Cell Physiol. 2016; 231:2570–81.
97. Chandel NS, Tuveson DA. The promise and perils of antioxidants for cancer patients. N Engl J Med. 2014; 371:177–8.
98. Gorrini C, Harris IS, Mak TW. Modulation of oxidative stress as an anticancer strategy. Nat Rev Drug Discov. 2013; 12:931–47.
99. Sabharwal SS, Schumacker PT. Mitochondrial ROS in cancer: initiators, amplifiers or an Achilles’ heel? Nat Rev Cancer. 2014; 14:709–21.
100. Bauer G. Targeting extracellular ROS signaling of tumor cells. Anticancer Res. 2014; 34:1467–82.
101. Wu CC, Bratton SB. Regulation of the intrinsic apoptosis pathway by reactive oxygen species. Antioxid Redox Signal. 2013; 19:546–58.
102. Wu Q, Ni X. ROS-mediated DNA methylation pattern alterations in carcinogenesis. Curr Drug Targets. 2015; 16:13–9.
103. Dewaele M, Maes H, Agostinis P. ROS-mediated mechanisms of autophagy stimulation and their relevance in cancer therapy. Autophagy. 2010; 6:838–54.
104. Li L, Ishdorj G, Gibson SB. Reactive oxygen species regulation of autophagy in cancer: implications for cancer treatment. Free Radic Biol Med. 2012; 53:1399–410.
105. Yang Y, Karakhanova S, Werner J, Bazhin AV. Reactive oxygen species in cancer biology and anticancer therapy. Curr Med Chem. 2013; 20:3677–92.
106. Liu J, Wang Z. Increased oxidative stress as a selective anticancer therapy. Oxid Med Cell Longev. 2015; 2015:294303.