
INTRODUCTION
Cutaneous malignant melanoma (CMM), the most lethal form of skin cancer, also presents one of the most rapidly increasing incidences in the caucasian population [1]. A thorough profiling of the driver mutations leading melanoma progression has been a pivotal step for the identification of actionable genetic events and related targeted therapies [2]. Due to the high mutational somatic burden, CMM cannot be considered a single entity, but a set of genetically heterogeneous tumors with peculiar patterns of oncogenic mutations whose characterization could explain the molecular timing of tumorigenesis, the differences in pharmacosensitivity, the occurrence of resistance to pharmacological inhibitors and the diverse metastatic potentials [2, 3]. Further studies are still needed to appropriately interpret the mutational signatures [2, 4]. The mutations found in melanoma mostly involve the mitogen-activated protein kinase (MAPK) and the phosphoinositide 3-kinase/protein kinase B (PI3K/AKT) pathways, with BRAFV600E representing the most common mutation (about 60%) and mostly correlated with intermittent sun exposure [5, 6] followed by NRAS (28%) and NF1, encoding for the neurofibromin 1 protein (14%) [2, 7, 8]. Furthermore loss of expression of cyclin-dependent kinase inhibitor 2A (CDKN2A) gene, leading to the dysregulation of p53 or Rb activity, is observed in up to 40% of sporadic CMMs [9]. Somatic CDKN2A mutation was observed in 13% of melanoma cases in the TCGA analysis [7]. In familial melanoma, up to 40% of patients show germline mutations in the CDKN2A gene, hence, CDKN2A is considered the major melanoma susceptibility gene with high penetrance in the south-european area [10]. In an Italian cohort, about 10% of sporadic multiple CMM cases are CDKN2A mutation positive, with a further increase of the mutation rate related to the number of CMMs [11]. When CDKN2A founder germline mutations are present, as in the Italian population, the percentage of sporadic CMMs consecutively enrolled harboring a CDKN2A germline mutation is not negligible, ranging from 2 to 9% [11–13]. Variants in low to medium penetrance susceptibility gene Melanocortin-receptor type 1 (MC1R) [14] have been inconsistently associated with BRAF somatic mutation [15–20].
Recently, recurrent activating mutations in the core promoter of Telomerase Reverse Transcriptase (TERT), causing an increase in telomerase activity have been found in about 30–55% of primary melanomas depending on studies and up to 85% of metastatic cases and studied as an independent prognostic factor in different types of cancers including melanoma [21–28], with conflicting results regarding the association with BRAF mutations [29, 30]. Interestingly, the above described activating mutations in the TERT promoter, first described in melanoma cases, were identified following the finding of a TERT promoter germline mutation at nt –57 (T>G) shown to play a role in melanoma susceptibility, given its segregation in an extended melanoma family. This mutant created a new binding motif for Ets transcription factors and ternary complex factors (TCFs) near the transcription start and, in reporter gene assays, caused up to twofold increase in TERT transcription [22, 23]. This seminal finding stimulated the studies on the correlation and interplay between somatic and germline changes in main driver of melanomagenesis, not limited to the coding region of genes but extending to regulatory regions [29–31]. The first aim of this study was to establish the frequency of somatic mutations in some of the key driver genes of melanoma development, assessed by sequencing and/or IHC in 93 well studied [32] primary melanomas from an Italian cohort of CMMs characterized for germline status. The second aim was to correlate the germline status (CDKN2A/CDK4/MC1R) of patients with somatic mutations/variants or protein loss, assessed by molecular analyses and/or immunohistochemistry, in some of the key CMM drivers (BRAF, NRAS, TERT, NF1, p16).
RESULTS
Patients characteristics
Our cohort, described in a previous study [32], included 93 primary cutaneous melanomas. Of those, 59 (63%) were superficial spreading melanomas (SSM), whereas 20 (22%) were nodular melanomas (NM), and the remaining 14 (15%) were neither SSM nor NM.
Median age was 49 years, 41 were males and 52 females. The majority of melanomas were situated in the trunk (n = 50), followed by lower limbs (23), upper limbs (16) and head and neck (4). Median Breslow thickness was 1.6 mm. Most of the patients were stage I (52/93, 56%), followed by 30 at stage II, 7 at stage III and 3 at stage IV; one patient presented an in situ melanoma. Sixteen melanomas (17%) were from affected probands of melanoma families, while 74 (80%) were from sporadic cases. 12 cases (13%), 4 familial and 8 sporadic, developed multiple primary melanomas. In 3 cases (3%) information about familiarity or other primary lesions was not available.
CDKN2A/CDK4 and MC1R germline mutations/variants
Germline analysis for CDKN2A and CDK4 status, obtained by capillary sequencing, was available for 88/93 and 59/93 cases, respectively. All coding sequences of the CDKN2A gene were sequenced. We found 14 samples (16%) to be positive for mutations in the CDKN2A gene: 9 presented the p.G101W mutation, 2 the p.P48T, 1 the p.E27X, 1 the p.Q50R and 1 the p.A68L. We also found 2 variants of unknown significance (VUS) which were predicted as not pathogenic with in silico prediction tools (p.M1R and p.H98H which results as p.P113S on the p14 transcript). No germline mutations were found in CDK4 exon 2. Capillary sequencing of MC1R gene was performed on 88 out of 93 samples and the variants retrieved were classified as indicated by Davies et al. [33]. We found 22 wild type samples (score 0), 33 with score 1, 18 with score 2, 12 with score 3 and 3 with score 4. In total, in our cohort 66 out of 93 samples (71%) had a MC1R variant of any type (“r” or “R”) (Figure 1).
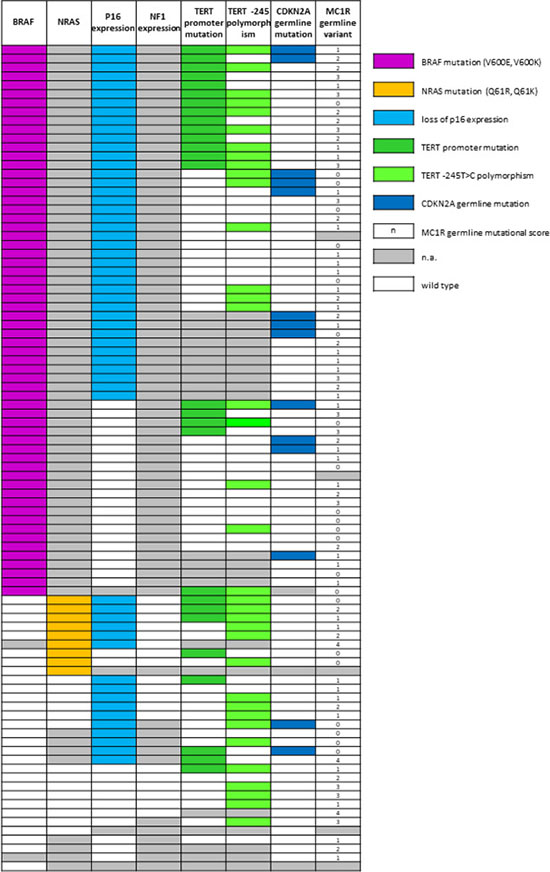
Figure 1: Combining results of germline and somatic analysis obtained in the study cohort. N.a.: not amplified. Wild-type: for p16 and NF1 expression, white cell means that the expression of the protein is maintained; for all the genes analysed by molecular methods white cell means wild-type sequence. For MC1R score, wild type corresponds to the 0 score.
BRAF mutational status
BRAF mutational status was assessed as detailed in our previous study, based on the concordance among IHC, PNA clamping real-time PCR (PNA) and, eventually, capillary sequencing [32]. Four samples were previously counted as undetermined at BRAF status due to unsuccessful capillary sequencing or invalid PNA results. These 4 samples were re-analyzed starting from a new DNA extraction from new slides from the same blocks. Due to the concordance among two methods, one case was deemed as BRAF positive, one as WT and the other two were still considered BRAF status undetermined (grey mark in Figure 1). Following the scheme proposed, we were able to classify 62 out of 91 patients (68%) as BRAF mutated and 29 out of 91 as wild type (32%). Of the 62 patients classified as mutated, 60 presented the classical c.1799 T>A missense mutation (p.V600E), one presented the substitution c.1799_1800delTGinsAA (p.V600E, often reported as p.V600E2) and one the c.1798_1799delGTinsAA mutation (p.V600K).
TERT promoter germline/somatic mutations
The analysis by capillary sequencing of the TERT gene promoter region was successful in 72 out of 93 tumor samples. We found 27 out of 72 samples (38%) with mutations previously described as pathological [24]. 8 (30%) showed the C>T substitution at –124 bp from the ATG start site (here called –124C>T, also indicated in literature as C228T), 17 (63%) the C>T transversion –146 bp from the ATG (here –146C>T, also described as C250T) and 2 samples (7%) both. Among the 72 amplified cases, 37 (51%) presented the –245T>C polymorphism (rs2853669) [34, 35]. In 17 out of 72 cases (24%) the variant was concomitant with TERT promoter mutations (Figure 1). In these patients we did not analyze germline DNA for the –57T>G mutation. This mutation, however, has never been detected in hundreds of probands from melanoma families analyzed from our cohort (data not shown).
p16 , NRAS, NF1 mutational status and expression
The expression of p16 was investigated in 89 samples by IHC with a highly specific antibody raised against full length recombinant p16. Thirty-three samples (37%) maintained the complete or partial expression of the protein while 56 (63%) showed negative staining (Figure 2). In the BRAF-wt and BRAF-undetermined samples we investigated the mutational status of the exon 3 of NRAS gene, with two methods, capillary sequencing and IHC. Capillary sequencing was successful in 15/31 samples (48%) and 4 resulted mutated: 3 presented the p.Q61R mutation and 1 the p.Q61K. These results were highly concordant with results obtained by IHC although the highly specific antibody is able to recognize only the NRAS Q61R mutation (Figure 2) [36]. These results were then completed with the data derived from sole IHC analysis, available in 8 additional samples. In total, 9 out 23 (39%) samples were mutated in NRAS: 8 were identified as Q61R by IHC (three confirmed by capillary sequencing) and 1 was identified as Q61K only by capillary sequencing, as expected. For the same BRAF-negative and undetermined samples, we checked the loss of expression of NF1 protein with an antibody designed to recognize the N terminal portion of the NF1 protein. All the samples in which the cell content was sufficient to be evaluated (19 out of 31) showed a positive staining (Figure 2), indicating that the NF1 protein expression was not lost in the samples available for testing.
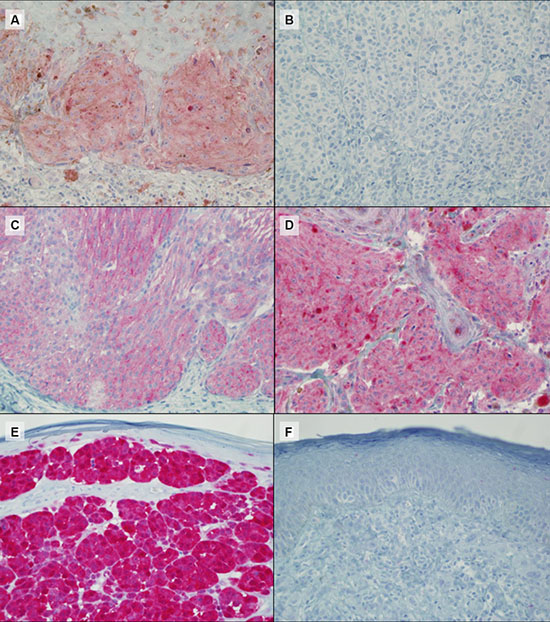
Figure 2: Representative IHC results for BRAF V600E (A: positive, B: negative), NRAS Q61R (C: positive), NF1 (D: positive) and p16 (E: positive and F: negative) protein expression. Magnification ×40.
Interplay between germline and somatic variants
Combining somatic and germline results, we observed that in 12 out of 86 (14%) cases there was co-occurrence of a somatic BRAF V600 mutation with a CDKN2A germline mutation. In line with previous studies [37], CDKN2A germline status (positive) showed an association with BRAF mutations, albeit without reaching statistical significance (OR = 3.16, p = 0.209; Table 1). In 45 out of 86 (52%) cases we found a somatic BRAF mutation combined with MC1R variation (any type), 18 of them (40%) included at least one “R” variant. The distribution of MC1R variants was not associated with BRAF mutations, either considering MC1R as a dichotomous variable (Table 1) or using the MC1R score as described by Davies et al. [33] (Kruskal–Wallis chi-squared = 0.04, degrees of freedom = 1, p = 0.845). MC1R variants were neither associated with TERT somatic variations nor with p16 loss (Table 1). p16 loss of expression in melanoma tissue was found in 10 out of 14 (71%) CDKN2A germline mutated cases, as compared to 45 out of 73 (62%) CDKN2A wild type cases, but this difference was not statistically significant (p = 0.559, Table 1). None of the NRAS mutated cases was CDKN2A germline mutation positive. Finally, only 3 out of 69 (4%) cases showed two somatic mutations (BRAF and TERT promoter) and a CDKN2A germline mutation (Figure 3).
Table 1: Association between genes and relative statistics
Gene_1 |
Gene_2 |
N |
OR |
p |
Lower_CI |
Upper_CI |
|---|---|---|---|---|---|---|
BRAF |
CDKN2A |
86 |
3.16 |
0.209 |
0.63 |
31.23 |
BRAF |
P16 loss |
87 |
1.39 |
0.628 |
0.48 |
3.94 |
BRAF |
TERT |
72 |
1.09 |
1 |
0.36 |
3.44 |
BRAF |
TERT -245 |
72 |
0.3 |
0.025 |
0.09 |
0.93 |
BRAF |
TERT -245 (conservative) |
44 |
0.15 |
0.009 |
0.03 |
0.67 |
BRAF |
MC1R (wt/any r or R) |
86 |
1.1 |
1 |
0.33 |
3.47 |
CDKN2A |
TERT |
69 |
1.04 |
1 |
0.19 |
4.96 |
CDKN2A |
TERT -245 |
69 |
1.44 |
0.737 |
0.31 |
7.7 |
CDKN2A |
MC1R (wt/any r or R) |
87 |
0.55 |
0.331 |
0.14 |
2.38 |
CDKN2A |
p16 loss |
87 |
1.55 |
0.559 |
0.4 |
7.42 |
P16 loss |
TERT |
71 |
2.87 |
0.075 |
0.89 |
10.45 |
P16 loss |
TERT -245 |
71 |
2.16 |
0.144 |
0.73 |
6.65 |
P16 loss |
MC1R (wt/any r or R) |
88 |
1.29 |
0.618 |
0.42 |
3.85 |
TERT |
MC1R (wt/any r or R) |
70 |
2.33 |
0.179 |
0.67 |
9.5 |
TERT -245 |
MC1R (wt/any r or R) |
70 |
1.08 |
1 |
0.34 |
3.48 |
Abbreviations: N, OR odds ratio, CI. TERT -245 (conservative) = samples harboring TERT-245T>C polymorphism without concurrent TERT promoter pathogenic mutations.
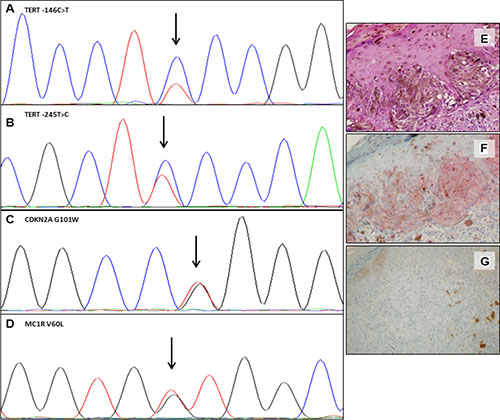
Figure 3: Somatic and germline mutations and variants in one representative case. Electropherograms showing the TERT promoter –146C>T somatic mutation (A) and the –245T>C polymorphism (B); CDKN2A p.G101W germline mutation (C) and of MC1R p.V60L germline variant (D). Hematoxylin and eosin (E), IHC positive staining for BRAF V600E (F) and IHC showing loss of expression of p16 protein (G). (Magnification ×40). The variant sequence is indicated by an arrow.
The TERT –245T>C polymorphism associates with BRAF-wt melanomas
Considering the total primary melanomas, we observed that BRAF mutation was concomitant in 19 out of 76 (25%) of samples with TERT promoter mutations and in 41 out of 62 (66%) with p16 loss of expression. In 14 out of 71 (20%) cases BRAF and TERT mutations were associated also with p16 loss of expression. The variant allele at −245 was observed in 21 out of 48 (44%) BRAF mutated and in 18 out of 28 (65%) BRAF wt. When examining the distribution of variants/mutations in our cohort, we found an association between TERT –245T>C polymorphism and the absence of BRAF pathogenic mutations (OR = 0.3, p = 0.025), as displayed in Figure 4A. To avoid any confounding effect by other mutations involving TERT, we then performed the same analysis on patients without concurrent TERT C228T and/or C250T mutations. In this subgroup, the inverse association between TERT –245T>C polymorphism and BRAF mutations was even stronger (OR = 0.15, p = 0.009, Figure 4B). In our cohort, the TERT –245T>C polymorphism was not associated with TERT promoter mutations (OR = 2.339589. p = 0.09542). Among the 9 primary melanoma samples which presented mutations at codon 61 of NRAS gene, 7 were successfully amplified for TERT: 4 (57%) showed mutations in TERT promoter and 6 (86%) presented the –245T>C polymorphism. A summary of all associations between genes and relative statistics is shown in Table 1. Due to the paucity of available data, we did not investigate the association of NRAS mutational status with other genes and with p16 expression.
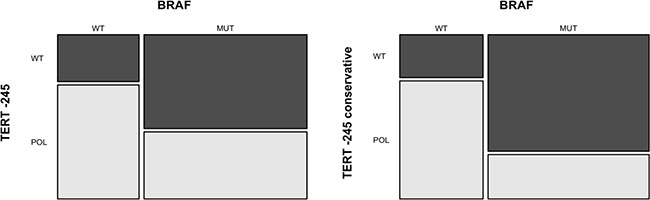
Figure 4: Association between the TERT –245T>C polymorphism and BRAF V600 mutation. The mosaic-plot shows an inverse association between BRAF V600 mutation and TERT –245 polymorphism in primary melanoma samples (A). This association is stronger when only samples without concurrent TERT promoter pathogenic mutations are analyzed (B) TERT –245 conservative = samples harboring TERT –245T>C polymorphism without concurrent TERT promoter pathogenic mutations. MUT= mutated samples; WT = wild-type samples; POL = samples with TERT –245T>C polymorphism ; WT = samples without TERT –245T>C polymorphism.
DISCUSSION
The aim of this study was to investigate the genetic heterogeneity of primary melanomas by means of molecular analyses and/or IHC from patients characterized for the germline status. Despite the high incidence of CDKN2A mutations, the number of patients in our cohort was limited, so we did not investigate prognostic features, but instead we focused on the interplay between germline and somatic variants in the key genetic drivers of cutaneous melanoma. Concerning the relationship between germline and somatic mutations, we saw positive, but not significant association between CDKN2A and BRAF mutations. These results are consistent with those of Zebary et al., who found no differences in BRAF and NRAS mutation frequencies between CDKN2A carriers and matched sporadic cases [38]. Moreover, although a positive association between CDKN2A and either BRAF or NRAS mutations was initially found in a recent study by Staaf et al., statistical significance was lost after adjusting for age and tumor thickness [37]. However, as opposed to recent research, BRAF mutations were not significantly associated with MC1R variants (alternatively considered as dichotomous variable or scored) and TERT promoter mutations or p16 loss. Moreover, we did not observe any association between CDKN2A status, MC1R and mutations in other genes or p16 loss, despite an association between TERT promoter mutations and MC1R recently found by Nagore et al. [39]. When investigating genetic interplay at the somatic level, we discovered that BRAF mutations are less frequent in patients with the TERT –245T>C polymorphism, especially when only considering patients without concurrent TERT promoter mutations. The role of TERT –245T>C polymorphism in cancer is debated, and research on this topic has provided controversial result. TERT –245T>C polymorphisms has been investigated as a poor prognostic marker in several tumors, including hepatocellular carcinoma [40] and glioblastoma [41]. However, according to Nagore et al., this polymorphism is linked to improved survival in melanoma patients with TERT promoter mutations [28]. The hypothesis of a protective role of this particular polymorphism in cancer is supported by functional studies. In fact, Laboussiere et al. demonstrated that TERT –245T>C polymorphism downregulates TERT mRNA expression in gliomas, as opposed to TERT promoter mutations [42]. An association between the TERT –245T>C polymorphism and BRAF mutation was investigated, but not found, in a study on differentiated thyroid cancer [43], showing that BRAF mutation was not correlated to TERT –245T>C polymorphism as an additional prognostic factor. In our cohort, BRAF was the most frequently mutated gene, with a mutation rate of 68%. In particular, V600E was the most represented BRAF mutation with only 2 cases with V600K and V600E2. NRAS was mutated in 39% of BRAF WT or undetermined cases. Interestingly, BRAF and NRAS mutation rates were higher compared to previous large studies or metanalysis [7, 44] with a high concordance between capillary sequencing and IHC that strengthens the efficiency of NRAS antibody, though specific for Q61R mutation. The mutation rate of TERT promoter was lower if compared to a Mittle-European study [45] which found TERT mutations in 55% of the tumor samples with no effect on OS or association with pathologic features of aggressiveness as otherwise reported [25, 26, 24, 46, 47]. Nevertheless coexistence of a TERT promoter and BRAF mutation was detected in 19 out of 76 cases (25%), consistently with a recent Italian study [25] and occurrence of TERT –245T>C variant allele accounted up to 65% in BRAF WT cases. Our study also some limitations. As stated before, the size of our samples did not allow as to explore the relationships between germline/somatic mutations and patients’ prognosis in terms of survival and risk of relapse. Moreover, we could not analyze the association between mutations in NRAS and NF1 with other germline/somatic mutations, as the information on both genes was available for a very limited number of patients. In conclusion, we studied some of the most relevant somatic mutations described in a cohort of primary CMMs characterized for germline status and we found that germinal CDKN2A mutations are neither associated with specific somatic mutations, nor with p16 loss of expression, confirming previous findings obtained with a combination of molecular and IHC studies [48–51]. However the mutation rate of BRAF was higher than previous described in studies from other populations with a peculiar inversion between the TERT –245T>C polymorphism and BRAF mutations. To the best of our knowledge this is the first study to date that describes an association between TERT –245T>C polymorphism and BRAF mutations in cutaneous melanoma. This result is of particular interest considering that TERT –245T>C can be frequently found as a germline polymorphism in the general population, as the minor allele frequency (MAF) is estimated to be 30%, and highest population MAF reaches 50% [Source: https://www.ensembl.org/]. Therefore, it is possible to hypothesize that specific genotypes, including the TERT –245T>C polymorphism, besides the debated SNPS in MC1R (not confirmed by the present study), may influence the occurrence of somatic BRAF mutations in individuals who develop cutaneous melanoma. Larger studies are needed to confirm these findings and hypothesize its role as a germline predictor of BRAF somatic mutation status.
MATERIALS AND METHODS
Case selection
A total of 100 primary melanomas were selected for a previous study on BRAF mutation detection [32], based on formalin-fixed, paraffin embedded (FFPE) tissue availability at the Pathology department of Policlinic Hospital, San Martino, Genoa. All melanoma patients had been referred to our center for germline testing either for diagnostic (familial and multiple melanoma cases) or research purposes (sporadic cases from a case-control study) and were part of a consecutive series of both incident and prevalent cases. All the patients signed an informed consent under local IRB approved protocols for both germline testing and other research purposes dealing with the archived melanoma tissues in the pathology department. A local database collecting information from the pathology report and tumor cell content in the examined section was designed. In this study we focused on cutaneous melanoma so we excluded seven uveal, mucosal and acral lentiginous melanomas due to the known different genetic signatures, leaving a total of 93 cases for lecular, IHC and statistical analysis. In case of patients with multiple primary melanoma only one melanoma lesion was analyzed molecularly.
DNA extraction and capillary sequencing
Genomic DNA extraction, either from the sections or form blood withdrawal, and capillary sequencing were performed as previously described [32, 52–55]. We amplified exon 3 of NRAS gene, promoter region of the TERT gene (from –270 to –40), exon 15 of BRAF gene, the whole CDKN2A coding region (exons 1a, 1b, 2 and 3, including promoter and 3′ regions), CDK4 exon 2 and the exon 1 of the MC1R gene. Sequencing reactions were repeated at least twice by independent PCR, with forward and reverse primers, and the sample was scored as being mutated when the mutation was observed both times. The same primers were used for both PCR and capillary sequencing. The in silico prediction tools we used were Poly-Phen 2, P-Mut, SIFT and Mutation taster.
PNA clamping quantitative PCR analysis
BRAF V600 codon mutational status was also tested using the PNAClamp™ BRAF Mutation Detection Kit (Panagene, Daejeon, Korea) according to the manufacturer’s instructions, with slight modifications, as previously described [52]. The threshold cycle (Ct) was automatically calculated from the PCR amplification plots where fluorescence was plotted against the number of cycles. Delta-Ct values were calculated as the Ct values of the samples minus those of the controls. The higher delta-Ct values showed that the mutant was efficiently amplified. A cut-off value of 2.0 was used to determine the presence of mutant DNA.
Immunohistochemistry (IHC)
For all immunohistochemical reactions, four micron-thick tissue sections were freshly cut [56], dried, deparaffinised and rehydrated. Endogenous peroxidase was blocked with 5% H2O2 for 10 minutes. Immunoreactions were performed using the automated BenchMark XT immunostainer® (Ventana Medical Systems, Arizona, USA). The following antibodies were used: BRAF V600E mutation-specific antibody (Springer-Bio, clone VE1, 1:50 dilution, standard heat-based antigen retrieval was performed for 30 minutes), the NRAS antibody (LifeSpan BioSciences Inc, clone SP174, 1:50 dilution, standard heat-based antigen retrieval was performed for 60 minutes) [36], specific for Q61R mutation, NF1 antibody (Santa Cruz Biotechnology Inc., clone sc-20982, 1:20 dilution, standard heat-based antigen retrieval was performed for 60 minutes) and p16 antibody (Santa Cruz Biotechnology Inc, clone sc-56330, 1:100 dilution, standard heat-based antigen retrieval was performed for 30 minutes). The ultraVIEW Universal Alkaline Phosphatase Red Detection Kit (Ventana Medical Systems, Arizona, USA), was used. After immunostaining, slides were counterstained with haematoxylin and coverslipped. All reactions were carried out by adding positive [57] and negative controls for each run. Positive and negative controls used were chosen from metastatic samples already characterized for BRAF and NRAS molecular status for diagnostic purposes (both capillary sequencing and, for BRAF, PNA clamping real-time PCR, as performed by the manufacturer) whereas positive NF1 and p16 were chosen from the immunohistochemistry laboratory positive control library. All immunostained slides were simultaneously scored by two pathologists (LM and FG); disagreement was resolved by consensus. For BRAF, IHC was considered positive when cytoplasmic protein expression was scored according to the 3 categories score used by Tetzlaff et al. [58]. IHC evaluation for other markers was conducted as follows: NRAS (scored into 3 categories as used for BRAF, see above), NF1 (negative staining was defined as absence of any cytoplasmic staining, positive was defined as diffuse cytoplasmic positivity of any intensity) and p16 (positivity was defined using the two categories in Lade-Keller et al. [49].
Statistical analysis
To assess association between two categorical variables with binary outcomes, such as mutational status of one gene or p16 loss, we performed the Fisher’s Exact test. Association between a categorical variable with binary outcome and an ordinal variable (MC1R score as described by Davies et al. [33]) was calculated using the Kruskal–Wallis test. All tests were two-sided and significance level to reject the null hypothesis was set at p = 0.05. Statistical analysis was carried out within the R computational environment [R Core Team (2016). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria [59].
Abbreviations
CDKN2A: cyclin-dependent kinase inhibitor 2A; CMM: Cutaneous malignant melanoma; FFPE: paraffin embedded; IHC: immunohistochemistry; MAF: minor allele frequency; MAPK: mitogen-activated protein kinase; MC1R: Melanocortin-receptor type 1; PI3K/AKT: phosphoinositide 3-kinase/protein kinase B; PNA: PNA clamping real-time PCR; TCFs: ternary complex factors; TERT: Telomerase Reverse Transcriptase.
Author contributions
BW: study design and working hypothesis, acquisition and interpretation of data, drafting of manuscript, final approval of the version to be published; MC, DB: analysis and interpretation of data, scientific advice, drafting of manuscript, final approval of the version to be published; AV, PL: analysis and interpretation of data, scientific advice, final approval of the version to be published; CF, GM, SF, BA, QP: acquisition of data, scientific advice, final approval of the version to be published; GF, ML, GP: study planning and supervision of the entire project, analysis and interpretation of data, drafting of manuscript, scientific advice, final approval of the version to be published.
CONFLICTS OF INTEREST
The authors declare no conflict of interest. FS received lecture fees from Bristol-Myers Squibb, GlaxoSmithKline, Novartis, MSD and Roche. PQ served as advisor to Bristol-Myers Squibb, Roche-Genentech, MSD, GlaxoSmithKline and Novartis, and received lecture fees from Bristol-Myers Squibb, GlaxoSmithKline, Novartis, MSD and Roche.
FUNDING
This study was supported by AIRC IG 15460 to PC and Italian Ministry of Health 5 × 1000 per la Ricerca Corrente to San Martino Hospital to PG and PQ.
REFERENCES
1. Apalla Z, Lallas A, Sotiriou E, Lazaridou E, Ioannides D. Epidemiological trends in skin cancer. Dermatol Pract Concept. 2017; 7:1–6. https://doi.org/10.5826/dpc.0702a01.
2. Reddy BY, Miller DM, Tsao H. Somatic driver mutations in melanoma. Cancer. 2017; 123:2104–17. https://doi.org/10.1002/cncr.30593.
3. Lim SY, Menzies AM, Rizos H. Mechanisms and strategies to overcome resistance to molecularly targeted therapy for melanoma. Cancer. 2017; 123:2118–29. https://doi.org/10.1002/cncr.30435.
4. Fredriksson NJ, Elliott K, Filges S, Van den Eynden J, Ståhlberg A, Larsson E. Recurrent promoter mutations in melanoma are defined by an extended context-specific mutational signature. PLoS Genet. 2017; 13:e1006773. https://doi.org/10.1371/journal.pgen.1006773.
5. Bauer J, Büttner P, Murali R, Okamoto I, Kolaitis NA, Landi MT, Scolyer RA, Bastian BC. BRAF mutations in cutaneous melanoma are independently associated with age, anatomic site of the primary tumor, and the degree of solar elastosis at the primary tumor site. Pigment Cell Melanoma Res. 2011; 24:345–51. https://doi.org/10.1111/j.1755-148X.2011.00837.x.
6. Poynter JN, Elder JT, Fullen DR, Nair RP, Soengas MS, Johnson TM, Redman B, Thomas NE, Gruber SB. BRAF and NRAS mutations in melanoma and melanocytic nevi. Melanoma Res. 2006; 16:267–73. https://doi.org/10.1097/01.cmr.0000222600.73179.f3.
7. Akbani R, Akdemir K, Aksoy BA, Albert M, Ally A, Amin S, Arachchi H, Arora A, Auman JT, Ayala B, Baboud J, Balasundaram M, Balu S, et al, and Cancer Genome Atlas Network. Genomic Classification of Cutaneous Melanoma. Cell. 2015; 161:1681–96. https://doi.org/10.1016/j.cell.2015.05.044.
8. Manzano JL, Layos L, Bugés C, de Los Llanos Gil M, Vila L, Martínez-Balibrea E, Martínez-Cardús A. Resistant mechanisms to BRAF inhibitors in melanoma. Ann Transl Med. 2016; 4:237–237. https://doi.org/10.21037/atm.2016.06.07.
9. Hodis E, Watson IR, Kryukov GV, Arold ST, Imielinski M, Theurillat JP, Nickerson E, Auclair D, Li L, Place C, Dicara D, Ramos AH, Lawrence MS, et al. A landscape of driver mutations in melanoma. Cell. 2012; 150:251–63. https://doi.org/10.1016/j.cell.2012.06.024.
10. Leachman SA, Carucci J, Kohlmann W, Banks KC, Asgari MM, Bergman W, Bianchi-Scarrà G, Brentnall T, Bressac-de Paillerets B, Bruno W, Curiel-Lewandrowski C, de Snoo FA, Debniak T, et al. Selection criteria for genetic assessment of patients with familial melanoma. J Am Acad Dermatol. 2009; 61:677.e1–14. https://doi.org/10.1016/j.jaad.2009.03.016.
11. Bruno W, Pastorino L, Ghiorzo P, Andreotti V, Martinuzzi C, Menin C, Elefanti L, Stagni C, Vecchiato A, Rodolfo M, Maurichi A, Manoukian S, De Giorgi V, et al. Multiple primary melanomas (MPMs) and criteria for genetic assessment: MultiMEL, a multicenter study of the Italian Melanoma Intergroup. J Am Acad Dermatol. 2016; 74:325–32. https://doi.org/10.1016/j.jaad.2015.09.053.
12. Mantelli M, Pastorino L, Ghiorzo P, Barile M, Bruno W, Gargiulo S, Sormani MP, Gliori S, Vecchio S, Ciotti P, Sertoli MR, Queirolo P, Goldstein AM, Bianchi-Scarrà G, and Italian Melanoma Intergroup. Early onset may predict G101W CDKN2A founder mutation carrier status in Ligurian melanoma patients. Melanoma Res. 2004; 14:443–48. https://doi.org/10.1097/00008390-200412000-00002.
13. Pastorino L, Bonelli L, Ghiorzo P, Queirolo P, Battistuzzi L, Balleari E, Nasti S, Gargiulo S, Gliori S, Savoia P, Abate Osella S, Bernengo MG, Bianchi Scarrà G. CDKN2A mutations and MC1R variants in Italian patients with single or multiple primary melanoma. Pigment Cell Melanoma Res. 2008; 21:700–09. https://doi.org/10.1111/j.1755-148X.2008.00512.x.
14. Ghiorzo P, Bonelli L, Pastorino L, Bruno W, Barile M, Andreotti V, Nasti S, Battistuzzi L, Grosso M, Bianchi-Scarrà G, Queirolo P. MC1R variation and melanoma risk in relation to host/clinical and environmental factors in CDKN2A positive and negative melanoma patients. Exp Dermatol. 2012; 21:718–20. https://doi.org/10.1111/j.1600-0625.2012.01549.x.
15. Williams PF, Olsen CM, Hayward NK, Whiteman DC. Melanocortin 1 receptor and risk of cutaneous melanoma: a meta-analysis and estimates of population burden. Int J Cancer. 2011; 129:1730–40. https://doi.org/10.1002/ijc.25804.
16. Hacker E, Olsen CM, Kvaskoff M, Pandeya N, Yeo A, Green AC, Williamson RM, Triscott J, Wood D, Mortimore R, Hayward NK, Whiteman DC. Histologic and Phenotypic Factors and MC1R Status Associated with BRAF(V600E), BRAF(V600K), and NRAS Mutations in a Community-Based Sample of 414 Cutaneous Melanomas. J Invest Dermatol. 2016; 136:829–37. https://doi.org/10.1016/j.jid.2015.12.035.
17. Fargnoli MC, Pike K, Pfeiffer RM, Tsang S, Rozenblum E, Munroe DJ, Golubeva Y, Calista D, Seidenari S, Massi D, Carli P, Bauer J, Elder DE, et al. MC1R variants increase risk of melanomas harboring BRAF mutations. J Invest Dermatol. 2008; 128:2485–90. https://doi.org/10.1038/jid.2008.67.
18. Landi MT, Bauer J, Pfeiffer RM, Elder DE, Hulley B, Minghetti P, Calista D, Kanetsky PA, Pinkel D, Bastian BC. MC1R Germline Variants Confer Risk for BRAF-Mutant Melanoma. Science. 2006; 313:521–2. https://doi.org/10.1126/science.1127515.
19. Thomas NE, Kanetsky PA, Edmiston SN, Alexander A, Begg CB, Groben PA, Hao H, Busam K, Ollila DW, Berwick M, Conway K. Relationship between germline MC1R variants and BRAF-mutant melanoma in a North Carolina population-based study. J Invest Dermatol. 2010; 130:1463–65. https://doi.org/10.1038/jid.2009.410.
20. García-Casado Z, Traves V, Bañuls J, Niveiro M, Gimeno-Carpio E, Jimenez-Sanchez AI, Moragón M, Onrubia JA, Oliver V, Kumar R, Nagore E. BRAF, NRAS and MC1R status in a prospective series of primary cutaneous melanoma. Br J Dermatol. 2015; 172:1128–31. https://doi.org/10.1111/bjd.13521.
21. Bell RJ, Rube HT, Kreig A, Mancini A, Fouse SD, Nagarajan RP, Choi S, Hong C, He D, Pekmezci M, Wiencke JK, Wrensch MR, Chang SM, et al. Cancer. The transcription factor GABP selectively binds and activates the mutant TERT promoter in cancer. Science. 2015; 348:1036–39. https://doi.org/10.1126/science.aab0015.
22. Horn S, Figl A, Rachakonda PS, Fischer C, Sucker A, Gast A, Kadel S, Moll I, Nagore E, Hemminki K, Schadendorf D, Kumar R. TERT promoter mutations in familial and sporadic melanoma. Science. 2013; 339:959–61. https://doi.org/10.1126/science.1230062.
23. Huang FW, Hodis E, Xu MJ, Kryukov GV, Chin L, Garraway LA. Highly Recurrent TERT Promoter Mutations in Human Melanoma. Science. 2013; 339:957–9. https://doi.org/10.1126/science.1229259.
24. Heidenreich B, Nagore E, Rachakonda PS, Garcia-Casado Z, Requena C, Traves V, Becker J, Soufir N, Hemminki K, Kumar R. Telomerase reverse transcriptase promoter mutations in primary cutaneous melanoma. Nat Commun. 2014; 5:3401. https://doi.org/10.1038/ncomms4401.
25. Macerola E, Loggini B, Giannini R, Garavello G, Giordano M, Proietti A, Niccoli C, Basolo F, Fontanini G. Coexistence of TERT promoter and BRAF mutations in cutaneous melanoma is associated with more clinicopathological features of aggressiveness. Virchows Arch. 2015; 467:177–84. https://doi.org/10.1007/s00428-015-1784-x.
26. Griewank KG, Murali R, Puig-Butille JA, Schilling B, Livingstone E, Potrony M, Carrera C, Schimming T, Möller I, Schwamborn M, Sucker A, Hillen U, Badenas C, et al. TERT promoter mutation status as an independent prognostic factor in cutaneous melanoma. J Natl Cancer Inst. 2014; 106:dju246. https://doi.org/10.1093/jnci/dju246.
27. Vinagre J, Pinto V, Celestino R, Reis M, Pópulo H, Boaventura P, Melo M, Catarino T, Lima J, Lopes JM, Máximo V, Sobrinho-Simões M, Soares P. Telomerase promoter mutations in cancer: an emerging molecular biomarker? Virchows Arch. 2014; 465:119–33. https://doi.org/10.1007/s00428-014-1608-4.
28. Nagore E, Heidenreich B, Rachakonda S, Garcia-Casado Z, Requena C, Soriano V, Frank C, Traves V, Quecedo E, Sanjuan-Gimenez J, Hemminki K, Landi MT, Kumar R. TERT promoter mutations in melanoma survival. Int J Cancer. 2016; 139:75–84. https://doi.org/10.1002/ijc.30042.
29. Hayward NK, Wilmott JS, Waddell N, Johansson PA, Field MA, Nones K, Patch AM, Kakavand H, Alexandrov LB, Burke H, Jakrot V, Kazakoff S, Holmes O, et al. Whole-genome landscapes of major melanoma subtypes. Nature. 2017; 545:175–80. https://doi.org/10.1038/nature22071.
30. Fredriksson NJ, Ny L, Nilsson JA, Larsson E. Systematic analysis of noncoding somatic mutations and gene expression alterations across 14 tumor types. Nat Genet. 2014; 46:1258–63. https://doi.org/10.1038/ng.3141.
31. Andreotti V, Bisio A, Bressac-de Paillerets B, Harland M, Cabaret O, Newton-Bishop J, Pastorino L, Bruno W, Bertorelli R, De Sanctis V, Provenzani A, Menin C, Fronza G, et al. The CDKN2A/p16(INK) (4a) 5'UTR sequence and translational regulation: impact of novel variants predisposing to melanoma. Pigment Cell Melanoma Res. 2016; 29:210–21. https://doi.org/10.1111/pcmr.12444.
32. Bruno W, Martinuzzi C, Andreotti V, Pastorino L, Spagnolo F, Dalmasso B, Cabiddu F, Gualco M, Ballestrero A, Bianchi-Scarrà G, Queirolo P, Grillo F, Mastracci L, Ghiorzo P, and Italian Melanoma Intergroup (IMI). Heterogeneity and frequency of BRAF mutations in primary melanoma: comparison between molecular methods and immunohistochemistry. Oncotarget. 2017; 8:8069–82. https://doi.org/10.18632/oncotarget.14094.
33. Davies JR, Randerson-Moor J, Kukalizch K, Harland M, Kumar R, Madhusudan S, Nagore E, Hansson J, Höiom V, Ghiorzo P, Gruis NA, Kanetsky PA, Wendt J, et al. Inherited variants in the MC1R gene and survival from cutaneous melanoma: a BioGenoMEL study. Pigment Cell Melanoma Res. 2012; 25:384–94. https://doi.org/10.1111/j.1755-148X.2012.00982.x.
34. Shen N, Lu Y, Wang X, Peng J, Zhu Y, Cheng L, Shen N, Lu Y, Wang X, Peng J, Zhu Y, Cheng L. Association between rs2853669 in TERT gene and the risk and prognosis of human cancer: a systematic review and meta-analysis. Oncotarget. 2017; 8:50864-50872. https://doi.org/10.18632/oncotarget.15140.
35. Pellegrini C, Maturo MG, Martorelli C, Suppa M, Antonini A, Kostaki D, Verna L, Landi MT, Peris K, Fargnoli MC. Characterization of melanoma susceptibility genes in high-risk patients from Central Italy. Melanoma Res. 2017; 27:258–67. https://doi.org/10.1097/CMR.0000000000000323.
36. Massi D, Simi L, Sensi E, Baroni G, Xue G, Scatena C, Caldarella A, Pinzani P, Fontanini G, Carobbio A, Urso C, Mandalà M. Immunohistochemistry is highly sensitive and specific for the detection of NRASQ61R mutation in melanoma. Mod Pathol. 2015; 28:487–97. https://doi.org/10.1038/modpathol.2014.137.
37. Staaf J, Harbst K, Lauss M, Ringnér M, Måsbäck A, Howlin J, Jirström K, Harland M, Zebary A, Palmer JM, Ingvar C, Olsson H, Newton-Bishop J, et al. Primary melanoma tumors from CDKN2A mutation carriers do not belong to a distinct molecular subclass. J Invest Dermatol. 2014; 134:3000–03. https://doi.org/10.1038/jid.2014.272.
38. Zebary A, Omholt K, van Doorn R, Ghiorzo P, Harbst K, Hertzman Johansson C, Höiom V, Jönsson G, Pjanova D, Puig S, Scarra GB, Harland M, Olsson H, et al. Somatic BRAF and NRAS mutations in familial melanomas with known germline CDKN2A status: a GenoMEL study. J Invest Dermatol. 2014; 134:287–90. https://doi.org/10.1038/jid.2013.270.
39. Nagore E, Reyes-Garcia D, Heidenreich B, Garcia-Casado Z, Requena C, Kumar R. TERT promoter mutations associate with MC1R variants in melanoma patients. Pigment Cell Melanoma Res. 2017; 30:273–75. https://doi.org/10.1111/pcmr.12567.
40. Ko E, Seo HW, Jung ES, Kim BH, Jung G. The TERT promoter SNP rs2853669 decreases E2F1 transcription factor binding and increases mortality and recurrence risks in liver cancer. Oncotarget. 2016; 7:684–99. https://doi.org/10.18632/oncotarget.6331.
41. Batista R, Cruvinel-Carloni A, Vinagre J, Peixoto J, Catarino TA, Campanella NC, Menezes W, Becker AP, de Almeida GC, Matsushita MM, Clara C, Neder L, Viana-Pereira M, et al. The prognostic impact of TERT promoter mutations in glioblastomas is modified by the rs2853669 single nucleotide polymorphism. Int J Cancer. 2016; 139:414–23. https://doi.org/10.1002/ijc.30057.
42. Labussière M, Di Stefano AL, Gleize V, Boisselier B, Giry M, Mangesius S, Bruno A, Paterra R, Marie Y, Rahimian A, Finocchiaro G, Houlston RS, Hoang-Xuan K, et al. TERT promoter mutations in gliomas, genetic associations and clinico-pathological correlations. Br J Cancer. 2014; 111:2024–32. https://doi.org/10.1038/bjc.2014.538.
43. Muzza M, Colombo C, Rossi S, Tosi D, Cirello V, Perrino M, De Leo S, Magnani E, Pignatti E, Vigo B, Simoni M, Bulfamante G, Vicentini L, Fugazzola L. Telomerase in differentiated thyroid cancer: promoter mutations, expression and localization. Mol Cell Endocrinol. 2015; 399:288–95. https://doi.org/10.1016/j.mce.2014.10.019.
44. Lee JH, Choi JW, Kim YS. Frequencies of BRAF and NRAS mutations are different in histological types and sites of origin of cutaneous melanoma: a meta-analysis. Br J Dermatol. 2011; 164:776–84. https://doi.org/10.1111/j.1365-2133.2010.10185.x.
45. Ofner R, Ritter C, Heidenreich B, Kumar R, Ugurel S, Schrama D, Becker JC. Distribution of TERT promoter mutations in primary and metastatic melanomas in Austrian patients. J Cancer Res Clin Oncol. 2017; 143:613–17. https://doi.org/10.1007/s00432-016-2322-1.
46. Nagore E, Heidenreich B, Requena C, García-Casado Z, Martorell-Calatayud A, Pont-Sanjuan V, Jimenez-Sanchez AI, Kumar R. TERT promoter mutations associate with fast-growing melanoma. Pigment Cell Melanoma Res. 2016; 29:236–38. https://doi.org/10.1111/pcmr.12441.
47. Seynnaeve B, Lee S, Borah S, Park Y, Pappo A, Kirkwood JM, Bahrami A. Genetic and Epigenetic Alterations of TERT Are Associated with Inferior Outcome in Adolescent and Young Adult Patients with Melanoma. Sci Rep. 2017; 7:45704. https://doi.org/10.1038/srep45704.
48. Ghiorzo P, Villaggio B, Sementa AR, Hansson J, Platz A, Nicoló G, Spina B, Canepa M, Palmer JM, Hayward NK, Bianchi-Scarrà G. Expression and localization of mutant p16 proteins in melanocytic lesions from familial melanoma patients. Hum Pathol. 2004; 35:25–33. https://doi.org/10.1016/j.humpath.2003.08.017.
49. Lade-Keller J, Riber-Hansen R, Guldberg P, Schmidt H, Hamilton-Dutoit SJ, Steiniche T. Immunohistochemical analysis of molecular drivers in melanoma identifies p16 as an independent prognostic biomarker. J Clin Pathol. 2014; 67:520–28. https://doi.org/10.1136/jclinpath-2013-202127.
50. Bosserhoff AK. Novel biomarkers in malignant melanoma. Clin Chim Acta. 2006; 367:28–35. https://doi.org/10.1016/j.cca.2005.10.029.
51. Straume O, Sviland L, Akslen LA. Loss of nuclear p16 protein expression correlates with increased tumor cell proliferation (Ki-67) and poor prognosis in patients with vertical growth phase melanoma. Clin Cancer Res. 2000; 6:1845–53.
52. Martinuzzi C, Pastorino L, Andreotti V, Garuti A, Minuto M, Fiocca R, Bianchi-Scarrà G, Ghiorzo P, Grillo F, Mastracci L. A combination of immunohistochemistry and molecular approaches improves highly sensitive detection of BRAF mutations in papillary thyroid cancer. Endocrine. 2016; 53:672–80. https://doi.org/10.1007/s12020-015-0720-9.
53. Ghiorzo P, Gargiulo S, Pastorino L, Nasti S, Cusano R, Bruno W, Gliori S, Sertoli MR, Burroni A, Savarino V, Gensini F, Sestini R, Queirolo P, et al. Impact of E27X, a novel CDKN2A germ line mutation, on p16 and p14ARF expression in Italian melanoma families displaying pancreatic cancer and neuroblastoma. Hum Mol Genet. 2006; 15:2682–89. https://doi.org/10.1093/hmg/ddl199.
54. Bruno W, Ghiorzo P, Battistuzzi L, Ascierto PA, Barile M, Gargiulo S, Gensini F, Gliori S, Guida M, Lombardo M, Manoukian S, Menin C, Nasti S, et al, and IMI, Italian Melanoma Intergroup. Clinical genetic testing for familial melanoma in Italy: a cooperative study. J Am Acad Dermatol. 2009; 61:775–82. https://doi.org/10.1016/j.jaad.2009.03.039.
55. Ghiorzo P, Pastorino L, Queirolo P, Bruno W, Tibiletti MG, Nasti S, Andreotti V, Paillerets BB, Bianchi Scarrà G, and Genoa Pancreatic Cancer Study Group. Prevalence of the E318K MITF germline mutation in Italian melanoma patients: associations with histological subtypes and family cancer history. Pigment Cell Melanoma Res. 2013; 26:259–62. https://doi.org/10.1111/pcmr.12047.
56. Grillo F, Pigozzi S, Ceriolo P, Calamaro P, Fiocca R, Mastracci L. Factors affecting immunoreactivity in long-term storage of formalin-fixed paraffin-embedded tissue sections. Histochem Cell Biol. 2015; 144:93–99. https://doi.org/10.1007/s00418-015-1316-4.
57. Bragoni A, Gambella A, Pigozzi S, Grigolini M, Fiocca R, Mastracci L, Grillo F. Quality control in diagnostic immunohistochemistry: integrated on-slide positive controls. Histochem Cell Biol. 2017; 148:569–73. https://doi.org/10.1007/s00418-017-1596-y.
58. Tetzlaff MT, Pattanaprichakul P, Wargo J, Fox PS, Patel KP, Estrella JS, Broaddus RR, Williams MD, Davies MA, Routbort MJ, Lazar AJ, Woodman SE, Hwu WJ, et al. Utility of BRAF V600E Immunohistochemistry Expression Pattern as a Surrogate of BRAF Mutation Status in 154 Patients with Advanced Melanoma. Hum Pathol. 2015; 46:1101–10. https://doi.org/10.1016/j.humpath.2015.04.012.
59. The R Project for Statistical Computing. Accessed 2017 Oct 12. Available from https://www.r-project.org/.