
INTRODUCTION
Lung cancer is the leading cause of cancer-related deaths worldwide, accounting for over 200,000 new cases and over 160,000 deaths per year in the United States, the most aggressive form being small cell lung cancer (SCLC) that accounts for >12% of all lung cancer diagnoses. In fact, SCLC patients have a median survival of 15-20 months and 5-year survival below 15% [1]. Treatment of SCLC is challenging, with a short disease-free survival after 1st line therapy [1, 2]. It is therefore of uttermost importance to gain a deeper understanding of the molecular events involved in SCLC tumorigenesis and progression in order to identify novel potential targets for therapy.
Bcl2-associated athanogene 3 (BAG3) belongs to a family of co-chaperones known to interact with the ATPase domain of the heat shock protein 70 (Hsp70) through the structural domain known as BAG domain (110-124 amino acids). In addition, BAG3 contains a WW domain, a proline-rich region (PXXP), and two conserved IPV (Ile-Pro-Val) motifs, that can also mediate binding to other proteins. While BAG3 can be induced in response to stress in cells of various origin it is constitutively expressed only in a few, including skeletal muscle and the heart. Importantly it is has also been shown to be constitutive in several primary tumors and tumor cell lines (pancreatic cancer, melanoma, leukemias, and others) [3, 4] and has been shown to play an important role in tumor biology [3, 4, 5, 6, 7, 8]. It has been suggested that its role in tumors is due to its anti-apoptotic properties in fact BAG3 has been shown to protect cells from death through a number of mechanisms that in general involve interaction with apoptosis- regulating proteins, including the IKK gamma subunit of the NF-κB- activating complex IKK [5], Bax [9], BRAF [10] and others [3].
Here we investigated BAG3 expression in SCLCs and its role in tumorigenesis in a xenograft mouse model. Our data suggest that indeed BAG3 is a potential target in a subset of tumors that express it.
RESULTS
Immunohistochemical analysis of BAG3 expression in human lung neoplastic samples
We analyzed BAG3 expression in 69 samples (36 small cell lung cancers, 13 squamous cell carcinomas, 13 adenocarcinomas, 4 large cell carcinomas and 3 normal lung samples as control) by immunoistochemistry (IHC), using an anti-BAG3 polyclonal antibody (TOS-2). As expected no BAG3 expression was detected in normal lung while all the NSCLC and 22 out of the 36 SCLC samples were BAG3- positive (Table 1 and Fig.1). Positive tumor samples showed a cytoplasmic staining in the majority of the cells.
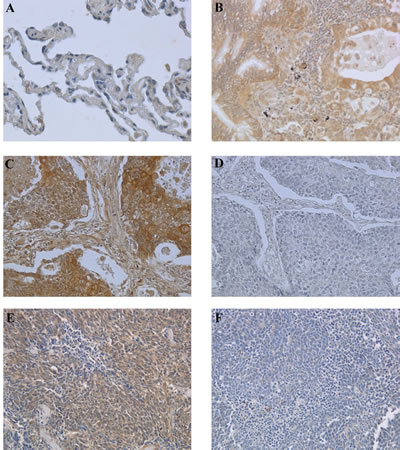
Figure 1: Immunohistochemical analysis of BAG3 expression in normal and neoplastic lung samples. Paraffin sections from 33 NSCLCs, 36 SCLCs and 3 normal lung specimens were analyzed by immunohistochemistry using anti-BAG3 rabbit polyclonal antibody. A) Normal lung tissue sample showing no immunoreactivity for BAG3 (200x) . B) Lung adenocarcinoma showing BAG3 cytoplasmic staining (200x). C) Squamous cell carcinoma sample showing cytoplasmic BAG3 staining (200x). D) Immunostaining of the same squamous cell carcinoma sample as in panel C stained without the primary antibody (200x). E) SCLC sample showing BAG3 cytoplasmic staining (200x). F) SCLC sample negative for BAG3 protein expression (200x).
Table 1: Immunohistochemical analysis of BAG3 expression in normal and pathological human lung tissues
Histological type |
N. of total cases analized |
BAG3 staining score* |
|||
0 |
1 |
2 |
3 |
||
Normal lung tissue |
3 |
3 |
|||
Squamous cell carcinoma |
13 |
0 |
2 |
5 |
6 |
Adenocarcinoma |
13 |
0 |
4 |
7 |
2 |
Large cell carcinoma |
4 |
0 |
1 |
1 |
2 |
Small cell lung cancer |
36 |
14 |
10 |
8 |
4 |
*Human lung specimens were stained with anti-BAG3 polyclonal antibody. The percentage of malignant cells stained was scored from 0 to 3: 0, no positive cells; 1+,<10% of positive cells; 2+,11-50% of positive cells; 3+, 51–100% of positive cells.
Effect of BAG3 down-modulation on SCLC cell apoptosis
We then investigated the possibility that in BAG3 positive SCLCs this protein played a pro-survival role as reported for other tumor types [3, 4]. To this end we first evaluated BAG3 expression by western blot in five human SCLC cell lines. As shown in Fig. 2A, H69 and H446 cell lines displayed the highest BAG3 protein levels and were chosen for the subsequent experiments to test if BAG3 silencing sensitized cells to cisplatin treatment. Both cell lines were transfected with a bag3- specific small interfering (si) RNA or with a non targeted (NT) siRNA. As shown in figures 2B and C transfection with 200 nM of Bag3 specific siRNA resulted in silencing of BAG3 in both cell lines. As shown in figure 3B and D silencing of BAG3 results in increased response to cisplatin with an increase in both cell lines of almost 40% of apoptosis after 48 hours of treatment with a 100 µM cisplatin. Furthermore, BAG3 silencing in H446 results also a significant increase of basal apoptosis (Fig. 3D).
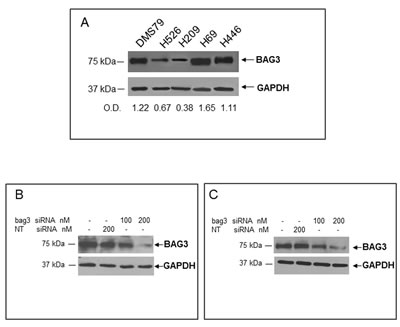
Figure 2: Analysis of BAG3 protein levels in human SCLC cell lines. A) Total protein extracts obtained from DMS79, H526, H209, H69 and H446 cell lines were analyzed by western blot using an anti-BAG3 polyclonal antibody. Densitometry data of samples are expressed as fractions of BAG3/GAPDH. H69 cells B) and H446 cells C) were transfected with a bag3-specific (bag3-siRNA) (100 and 200 nM) or a NT siRNA (200 nM). After 72 hrs BAG3 protein levels were analyzed by western blot using an anti-BAG3 polyclonal antibody. Anti-GAPDH antibody was used as loading control.
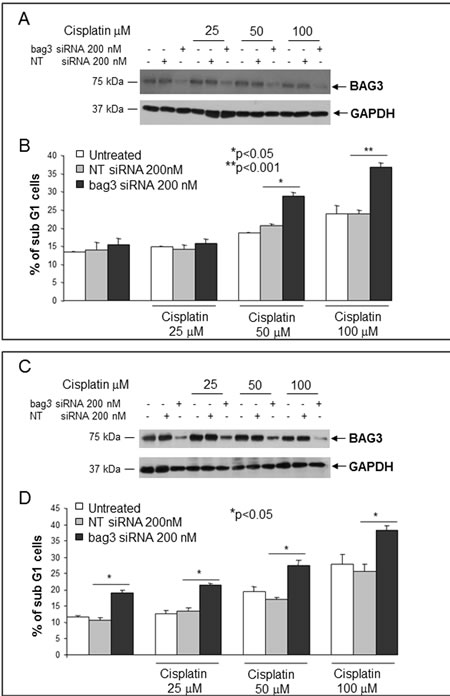
Figure 3: Effect of BAG3 down-regulation on SCLC cell apoptosis. H69 (A, B) or H446 (C, D) cells were transfected with bag3 siRNA (200 nM) or with NT siRNA (200 nM) and after 48 hrs treated with different doses of Cisplatin (25, 50, 100 microM) for 48 hrs. Cells were then collected and labeled with propidium iodide and analyzed by flow cytometry (B, D). Total protein extracts of cells treated as described above were analyzed by western blot using an anti-BAG3 polyclonal antibody. Anti-GAPDH antibody was used as loading control (A, C). *p<0.05; **p<0.001.
These results indicate that down-regulation of BAG3 stimulates apoptosis in SCLC cells in vitro.
Down-regulation of BAG3 reduces in vivo tumor growth and induces apoptosis
Since in vitro data showed that down-regulation of BAG3 induces apoptosis in SCLS cells, we investigated the effects of bag3 siRNA-Ad treatment on tumor growth and apoptosis in vivo. As shown in Fig. 4A, we observed that intra-tumoral injection of bag3 siRNA-Ad was able to reduce in vivo tumor growth after 44 days of treatment as compared to the scramble-treated (scr siRNA-Ad) and control groups (p<0.001). Accordingly, mice treated with bag3 siRNA-Ad survived longer than control (PBS) and scr siRNA-Ad-treated mice (p<0.05) (Fig. 4B). Western blot and immunohistochemical analysis of tumor samples from xenografts treated for 12 days with bag3 siRNA-Ad specifically confirm a reduction of BAG3 protein levels (Fig. 5 A, B).
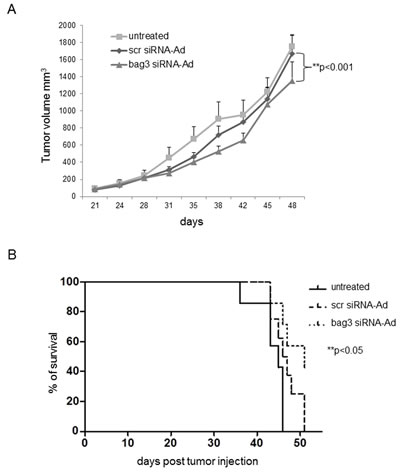
Figure 4: BAG3 down-regulation reduces tumor growth and increases animal survival in a xenograft model. H69 cells (3x 106) were injected subcutaneously onto the back of six/eight-week-old female athymic nude-Foxn1nu/nu mice. Two weeks later (100 mm3), animals were randomized into three groups (8 animals per group) and control PBS (100μl-untreated), bag3 siRNA-Ad or scrRNA-Ad (108 pfu/100 μl) were injected in the tumors twice a week. A) Tumor size was measured every week using a caliper in bag3 siRNA-Ad, scr siRNA-Ad treated or untreated mice. **p< 0.001. B) Kaplan-Meier analysis of animal survival. *p< 0.05.
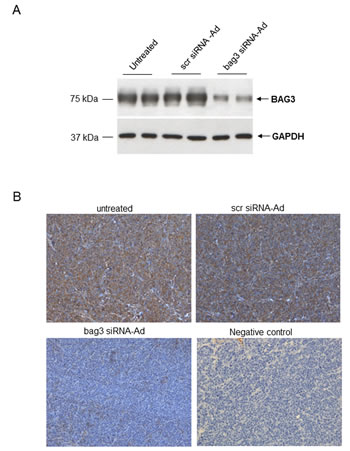
Figure 5: Analysis of BAG3 expression in H69 xenografts. A) Western blot analysis of protein extracted from H69 xenografts treated with PBS 100 μl (untreated), scr siRNA-Ad (108 pfu/100μl) or bag3 siRNA-Ad (108 pfu/100μl) by intratumoral injection, twice a week. B) Immunohistochemistry with anti-BAG3 monoclonal of xenograft tumors treated as in A (200x). Negative control: immunostaining of xenografted tumor samples stained without the primary antibody (200x).