
INTRODUCTION
Multidrug-resistant Gram-negative bacteria, particularly carbapenem-resistant Enterobacteriaceae (CRE), have spread globally into hospitals and communities, and thus have become a significant public health concern [1, 2]. For clinical infections caused by CRE, the treatment options are limited and the polymixins (colistin and polymyxin B) are the last-resort antibiotic. The recent identification of a plasmid-encoded polymyxin resistance mechanism (MCR-1) in Enterobacteriaceae from both human and animal samples suggests that this last-resort antibiotic may be under jeopardy [3, 4]. To date, mcr-1-harboring plasmids have been identified in a number of countries with a wide geographical distribution, and some MCR-1 producing strains were resistant to multiple antibiotics [5, 6].
Polymyxin resistance is the result of a 4′-phosphoethanolamine (PEA) or 4-amino-4-deoxy-L-arabinose (L-Arap4N) modification of bacterial lipid A, which is a component of the lipopolysaccharide (LPS), and results in a reduction in polymyxin affinity. Resistance is usually chromosomally mediated and involves modulation of two-component regulatory systems (e.g., pmrAB, phoPQ, and its negative regulator mgrB in Klebsiella pneumoniae) [7, 8]. The plasmid-transferrable mobilized colistin resistance gene mcr-1, encoding a novel PEA-transferase [3, 4], is able to mediate a PEA addition to the lipid A moiety at the 4′-phosphate group, thereby causing colistin resistance. So far, gene expression and transcriptomic analyses of chromosomal colistin resistance genes have been reported frequently. In contrast, the number of studies on gene regulation of mcr-1 is limited, since most of the mcr-1 studies have been focusing on epidemiological investigations of mcr-1 in different Enterobacteriaceae, including Escherichia coli, Salmonella enterica, K. pneumonia, Enterobacter aerogenes and Enterobacter cloacae [9–11].
Different incompatibility (InC) groups of plasmids have been found to carry mcr-1, while among them the IncI2 and IncX4 plasmids were most commonly reported. In the previous studies, we completely sequenced mcr-1-harboring plasmids pMCR1_IncX4 and pMCR1_IncI2 from clinical K. pneumoniae and E. coli strains [12]. In addition, we genomically characterized one of the first chromosomally encoded mcr-1 genes from an E. coli (BJ10) isolate [13]. In this study, we used quantitative reverse transcription PCR (qRT-PCR) to evaluate the expressions of mcr-1 of different plasmids (pMCR1_IncX4 and pMCR1_IncI2) within different species (E. coli and K. pneumoniae) and their changes in response to colistin challenge.
RESULTS
Analysis of the mcr-1 locus and its promoter
Sequence comparison of the mcr-1 neighboring regions in pMCR1_IncX4 (accession No. KU761327), pMCR1_IncI2 (accession No. KU761326) and BJ10 (accession No. LWQZ00000000) showed that the phosphoesterase encoding gene pap2 is universally present downstream of mcr-1 and forms a mcr-1-pap2 cassette, while two copies of ISApl1 flank the mcr-1-pap2 cassette on the chromosome of BJ10 (TnApl1) (Figure 1A). As shown in Figure 1B, the promoter sequences of mcr-1 in pMCR1_IncX4, pMCR1_IncI2 and BJ10 are similar to that of pAf23 and pAf48 reported by Poirel et al. [14].

Figure 1: Schematic map of the different mcr-1- bearing plasmids pMCR1_IncX4 and pMCR1_IncI2. (A) Model of mcr-1 locus. (B) Analyses of mcr-1 promoter. S, transcription start site; M, methionine and translation initiation site; The mcr-1 promoter sequences are indicated with the corresponding -10 and -35 boxes being underlined according to Poirel L’s work.
Expression of mcr-1 in plasmids pMCR-1_InX4 and pMCR1_IncI2 in parental strains
As shown in Figure 2, mcr-1 expressions in pMCR1_IncX4 varied from 1.81 × 10–5 to 1.05 × 10–4 (pmol per μg total RNA) in the two ST25 K. pneumoniae strains SZ03 and SZ04, and the two E. coli strains CDA6 and SZ01 (ST167 and ST2448, respectively), with the ST167 E. coli strain CDA6 showing the highest expression levels. The mcr-1 expression of pMCR1_IncI2 in the ST2085 E. coli strain SZ02 and ST457 E. coli strain BJ13 was 5.27 × 10–5 and 2.58 × 10–5, respectively. In addition, the expression of chromosomal mcr-1 (with ISApl1 inserted in the two flanking regions) in the ST156 E. coli strain BJ10 was 5.49 × 10–5. Our results showed that mcr-1 expression from the same plasmid may vary significantly when expressed in the different genetic backgrounds of the different strains. In addition, we investigated the MICs for colistin in these mcr-1 positive parental strains (Table 1) and found that the MICs for colistin were not consistent with the differences in expression of mcr-1 in these strains. These results suggest that colistin resistance not solely depends on the expression levels of mcr-1 and the structural modification of lipid A mediated by mcr-1 [15].
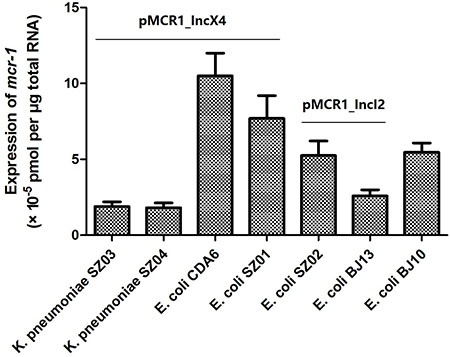
Figure 2: The mcr-1 expression of plasmids pMCR1_IncX4 and pMCR1_IncI2 in parental strains. Bacteria were grown to an OD600 of 0.5 and total RNA were extracted and subsequently used to do the qRT-PCR experiments. The data were analyzed by using Student’s t-test and shown as Mean with SEM.
Table 1: Strains and plasmids tested in this study
Strain |
Background |
mcr-1-harboring Plasmid |
Colistin MIC (μg/ml) |
Reference |
|---|---|---|---|---|
K. pneumoniae SZ03 |
ST25 |
pMCR1_IncX4 |
32 |
[12] |
K. pneumoniae SZ04 |
ST25 |
pMCR1_IncX4 |
32 |
[12] |
E. coli CDA6 |
ST167 |
pMCR1_IncX4 |
8 |
[13] |
E. coli SZ01 |
ST2448 |
pMCR1_IncX4 |
4 |
[12] |
E. coli SZ02 |
ST2085 |
pMCR1_IncI2 |
8 |
[12] |
E. coli BJ13 |
ST457 |
pMCR1_IncI2 |
4 |
[13] |
E. coli BJ10 |
ST156 |
$ |
8 |
[13] |
E. coli E600 |
# |
pMCR1_IncX4 |
4 |
This study |
E. coli E600 |
# |
pMCR1_IncI2 |
8 |
This study |
$: mcr-1 located in the chromosome, #: genetic engineering strain
Expression dynamics of mcr-1 in parental strains after colistin treatment for different times
We next examined mcr-1 expression from different plasmids in different bacterial hosts after colistin treatment. As shown in Figure 3, upon colistin treatment, the expression of mcr-1 in pMCR1_IncX4 increased 2- and 4-fold after 20 and 120 mins, respectively, among all pMCR1_IncX4-harboirng strains, except for the ST2448 E. coli SZ01 strain, which showed a lower expression after 20 mins that restored to baseline levels after 120 mins. In contrast, the mcr-1 expression levels of pMCR1_IncI2-harboring parental strains SZ02 and BJ13 remained at baseline level after 20 and 120 mins. However, the expression of chromosomal mcr-1 in E. coli (BJ10) remained stable, regardless of colistin treatment. These results demonstrated various mcr-1 expression patterns of mcr-1-harboring plasmids in different bacterial hosts after colistin treatment, which are likely the result of differences in bacterial backgrounds and/or the different mcr-1-harboirng plasmids.
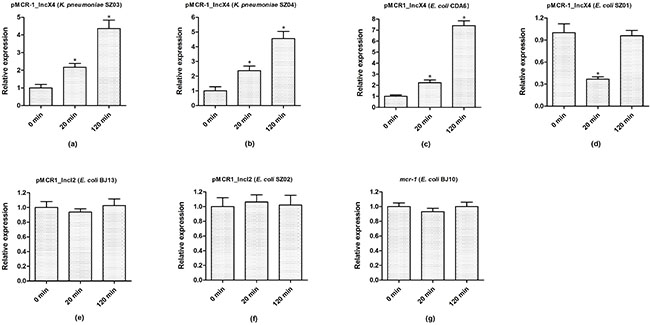
Figure 3: The expression dynamics of mcr-1 in parental strains under colistin treatment for different time. Bacteria were grown to an OD600 of 0.5, and then colistin was added into the cultures with the final concentration of 4 μg/ml. After 20 mins and 120 mins treatment by colistin, bacterial total RNA were isolated and subsequently used to do the qRT-PCR experiments. The data were analyzed by using Student’s t-test and shown as Mean with SEM. *P < 0.05 (compared to 0 mins treatment by colistin).
Expression of mcr-1 in E. coli E600 after colistin treatment for different times
To examine whether the differential expression observed above correlates with the different mcr-1-harboring plasmid backgrounds, we transferred plasmids pMCR1_IncX4 and pMCR1_IncI2 into the same E. coli host strain E600 via conjugation, and investigated the expression of mcr-1 after colistin treatment for different times. As shown in Figure 4, the expression of mcr-1 from the two different plasmids pMCR1_IncX4 and pMCR1_IncI2 was very similar without colistin challenge (1.31 × 10–4 and 1.49 × 10–4 pmol per μg total RNA, respectively) . However, mcr-1 expression after colistin treatment showed variations for plasmids pMCR1_IncX4 and pMCR1_IncI2. After treatment with colistin for 20 min, mcr-1 was significantly down-regulated in plasmid pMCR1_IncX4 and pMCR1_IncI2, while after treatment with colistin for 120 min, mcr-1 was up-regulated in plasmid pMCR1_IncX4 but significantly down-regulated in plasmid pMCR1_IncI2.
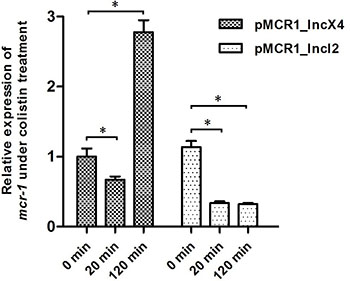
Figure 4: The mcr-1 expression of plasmids pMCR1_IncX4 and pMCR1_IncI2 in same host strain E. coli E600 under colistin treatment for different time. Bacteria were grown to an OD600 of 0.5, and then colistin was added into the cultures with the final concentration of 4 μg/ml. After 20 mins and 120 mins treatment by colistin, bacterial total RNA were isolated and subsequently used to do the qRT-PCR experiments. The data were analyzed by using Student’s t-test and shown as Mean with SEM. *P < 0.05 (compared to 0 mins treatment by colistin).
DISCUSSION
Colistin is a cationic polypeptide antibiotic which is regarded as one of the last antibacterial agents against CRE. In general, colistin resistance of Gram-negative bacteria like Klebsiella, E. coli and Salmonella enterica has been mediated by chromosomal mutations and was thought to be non-transferable. The emergence of colistin resistance mediated by the mcr-1 gene on a plasmid has become a matter of major concern since its first report in China. To date, the mcr-1 gene has been detected in Enterobacteriaceae from almost 35 countries all of the world, including E. coli, E. aerogenes, E. cloacae, K. pneumonia, Shigella sonnei and S. enterica [16–19]. A number of different plasmids including the most common, IncI2 and IncX4, have been associated with the spread of mcr-1. Recently, Wang et al. described over 10 mcr-1-harboring plasmids with diversified incompatibility in E. coli [20], and reported that the mcr-1 promoter of different origins exhibits similar activity through transcriptional analyses [21]. However, the mechanism of mcr-1 gene regulation is still unclear.
Previously, we identified seven CRE strains carrying the mcr-1 gene, including two K. pneumonia isolates and five E. coli isolates, which contained the two different mcr-1-harboring plasmids pMCR1_IncX4 and pMCR1_IncI2 [4, 12, 13]. In this study, we found that mcr-1 expression of the plasmids pMCR1_IncX4 and pMCR1_IncI2 may vary significantly in the different genetic background of different strains, although their promoters are highly similar. In general, gene expression is controlled by its promoter and the corresponding activators and/or inhibiters. Therefore, we suspect that genes encoding activators and/or inhibiters in the host chromosome may affect the expression of mcr-1 located on plasmids pMCR1_IncX4 and pMCR1_IncI2. In addition, we found that the phosphoesterase encoding gene pap2 is universally present downstream of mcr-1 and forms the mcr-1-pap2 cassette in pMCR1_IncX4 and pMCR1_IncI2. A previous study showed that pap2 is likely co-mobilized with the mcr-1 gene when it transferred from its original genetic context and does not impact colistin vsusceptibility [22].
After treatment withcolistin, various mcr-1 expression patterns of mcr-1-harboring plasmids from different bacterial hosts were detected, and we suspect that the differential expression changes may be due to the differences in bacterial backgrounds and/or the different mcr-1-harboirng plasmids. Moreover, we found that different plasmids had similar baseline mcr-1 expression within the same E. coli E600 strain, suggestin that the expression of mcr-1 from different plasmids is likely controlled by the host genetic background without colistin challenge. However, after colistin treatment, both the host bacteria and the mcr-1-harboring plasmids may contribute to mcr-1 gene expression regulation, therefore a complex regulation network of mcr-1 may be involved.
Taken together, this is one of the first studies focusing on the expression characteristics of mcr-1 from different plasmids and in different bacterial host backgrounds. It is suggested that different genetic signatures on the plasmids or in the bacterial strains may contribute to the observed variations, which warrant further investigations. Therefore, this study laid the foundation for further research on the regulation mechanism of mcr-1 expression.
MATERIALS AND METHODS
Bacterial strains and MIC measurements of colistin
Seven mcr-1 positive clinical isolates are used in this study, including two K. pneumoniae strains and five E. coli strains. All the bacterial strains used in this study are listed in Table 1. Minimal inhibitory concentrations (MICs) of colistin were determined using the agar dilution method and E. coli strain ATCC 25922 was used as a quality control strain.
Transfer of mcr-1 gene to the same host strain E. coli E600
The mcr-1-harboring plasmids pMCR1_IncX4 and pMCR1_IncI2 from E. coli SZ01 and E. coli SZ02, respectively, were transferred to the same host strain, E. coli E600, via conjugation. E. coli SZ01 and E. coli SZ02 isolates were used as donors, and E. coli E600 (resistant to rifampicin) was used as the recipient strain. Conjugation was carried out by broth mating and positive strains were selected by colistin and rifampicin dual resistance.
Expression of mcr-1 investigated by qRT-PCR
Full length mcr-1 from pMCR1_IncX4 was cloned into a T-vector in E. coli DH5α, and this recombinant T-vector was used as the standard for quantitative reverse transcription PCR (qRT-PCR). The absolute expression levels of mcr-1 of the pMCR1_IncX4 and pMCR1_IncI2 plasmids in their parental K. pneumoniae and E. coli isolates (Table 1) were investigated by qRT-PCR. In addition, the plasmid pMCR1_IncX4 and pMCR1_IncI2 were transferred into E. coli E600 via conjugation, and their transconjugants were evaluated by qRT-PCR. Moreover, the mcr-1 chromosomally integrated E. coli (BJ10) was included in the tests [13].
In the qRT-PCR experiments, the forward primer sequence for mcr-1 was AAATCAGCCAAACCTATCCC, and reverses primer sequence was CGTATCATAGACCGT GCCAT. The housekeeping genes rpoD and gapA were used for normalization for E. coli and K. pneumoniae, respectively. All the strains were cultured overnight at 37°C with shaking (250 rpm) in 1 ml LB broth, and then 1:100 diluted into 10 ml fresh medium. Cultures were incubated to exponential growth (OD 0.5 at 600 nm) and total RNA was extracted using an RNeasy kit (Qiagen, Germany) according to the manufacturer’s instructions. Extracted RNA was treated with 1 U of RNase-free DNase I at 37°C for 10 min to remove traces of DNA and incubated at 85°C for 15 min to inactivate the DNase. Subsequently, 1 μg total RNA was subjected to qRT-PCR as described previously [23]. Each experiment was performed with three RNA samples from three independent experiments. Differences between the two groups were assessed by Student’s t-test, P < 0.01 was considered to be statistically significant.
Expression dynamics of mcr-1 under colistin treatment investigated by qRT-PCR
To describe the expression dynamics of mcr-1 under colistin treatment, mcr-1 positive strains were cultured to exponential growth (OD 0.5 at 600 nm), and then colistin was added into the cultures with a final concentration of 4 μg/ml. After 20 mins and 120 mins treatment with colistin, bacterial total RNA were isolated and subsequently used to do the qRT-PCR experiments as above.
ACKNOWLEDGMENTS AND FUNDING
This study was supported by the National Natural Science Foundation of China (81572032, 81401636), Key research and development project of Jiangsu provincial science and Technology Department (BE2017654), Six talent peaks project in Jiangsu Province (2016-WSN-112), Gusu key health talent of Suzhou, the Natural Science Foundation for Colleges and Universities in Jiangsu Province (16KJB320006), Jiangsu youth medical talents program (QN-866, 867) and the Science and Technology Program of Suzhou (SZS201715, SYS201551, SS201638, SYS201619).
CONFLICTS OF INTEREST
The authors declare no conflicts of interests.
REFERENCES
1. Kumarasamy KK, Toleman MA, Walsh TR, Bagaria J, Butt F, Balakrishnan R, Chaudhary U, Doumith M, Giske CG, Irfan S, Krishnan P, Kumar AV, Maharjan S, et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: a molecular, biological, and epidemiological study. Lancet Infect Dis. 2010; 10:597–602. https://doi.org/10.1016/S1473-3099(10):70143-2.
2. Munoz-Price LS, Poirel L, Bonomo RA, Schwaber MJ, Daikos GL, Cormican M, Cornaglia G, Garau J, Gniadkowski M, Hayden MK, Kumarasamy K, Livermore DM, Maya JJ, et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect Dis. 2013; 13:785–96. https://doi.org/10.1016/S1473-3099(13)70190-7.
3. Liu YY, Wang Y, Walsh TR, Yi LX, Zhang R, Spencer J, Doi Y, Tian G, Dong B, Huang X, Yu LF, Gu D, Ren H, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016; 16:161–8. https://doi.org/10.1016/S1473-3099(15)00424-7.
4. Du H, Chen L, Tang YW, Kreiswirth BN. Emergence of the mcr-1 colistin resistance gene in carbapenem-resistant Enterobacteriaceae. Lancet Infect Dis. 2016; 16:287–8. https://doi.org/10.1016/S1473-3099(16)00056-6.
5. Gao R, Hu Y, Li Z, Sun J, Wang Q, Lin J, Ye H, Liu F, Srinivas S, Li D, Zhu B, Liu YH, Tian GB, et al. Dissemination and Mechanism for the MCR-1 Colistin Resistance. PLoS Pathog. 2016; 12:e1005957. https://doi.org/10.1371/journal.ppat.1005957.
6. Teo JW, Chew KL, Lin RT. Transmissible colistin resistance encoded by mcr-1 detected in clinical Enterobacteriaceae isolates in Singapore. Emerg Microbes Infect. 2016; 5:e87. https://doi.org/10.1038/emi.2016.85.
7. Cannatelli A, D’Andrea MM, Giani T, Di Pilato V, Arena F, Ambretti S, Gaibani P, Rossolini GM. In vivo emergence of colistin resistance in Klebsiella pneumoniae producing KPC-type carbapenemases mediated by insertional inactivation of the PhoQ/PhoP mgrB regulator. Antimicrob Agents Chemother. 2013; 57:5521–6. https://doi.org/10.1128/AAC.01480-13.
8. Gunn JS. The Salmonella PmrAB regulon: lipopolysaccharide modifications, antimicrobial peptide resistance and more. Trends Microbiol. 2008; 16:284–90. https://doi.org/10.1016/j.tim.2008.03.007.
9. Prim N, Rivera A, Rodríguez-Navarro J, Español M, Turbau M, Coll P, Mirelis B. Detection of mcr-1 colistin resistance gene in polyclonal Escherichia coli isolates in Barcelona, Spain, 2012 to 2015. Euro Surveill. 2016; 21. https://doi.org/10.2807/1560-7917.ES.2016.21.13.30183.
10. Doumith M, Godbole G, Ashton P, Larkin L, Dallman T, Day M, Day M, Muller-Pebody B, Ellington MJ, de Pinna E, Johnson AP, Hopkins KL, Woodford N. Detection of the plasmid-mediated mcr-1 gene conferring colistin resistance in human and food isolates of Salmonella enterica and Escherichia coli in England and Wales. J Antimicrob Chemother. 2016; 71:2300–5. https://doi.org/10.1093/jac/dkw093.
11. Zeng KJ, Doi Y, Patil S, Huang X, Tian GB. Emergence of plasmid-mediated mcr-1 gene in colistin-resistant Enterobacter aerogenes and Enterobacter cloacae. Antimicrob Agents Chemother. 2016; 60:3862–3. https://doi.org/10.1128/AAC.00345-16.
12. Li A, Yang Y, Miao M, Chavda KD, Mediavilla JR, Xie X, Feng P, Tang YW, Kreiswirth BN, Chen L, Du H. Complete Sequences of mcr-1-Harboring Plasmids from Extended-Spectrum-β-Lactamase- and Carbapenemase-Producing Enterobacteriaceae. Antimicrob Agents Chemother 2016; 60:4351–4. https://doi.org/10.1128/AAC.00550-16.
13. Yu H, Qu F, Shan B, Huang B, Jia W, Chen C, Li A, Miao M, Zhang X, Bao C, Xu Y, Chavda KD, Tang YW. et al. Detection of the mcr-1 Colistin Resistance Gene in Carbapenem-Resistant Enterobacteriaceae from Different Hospitals in China. Antimicrob Agents Chemother. 2016; 60:5033–5. https://doi.org/10.1128/AAC.00440-16.
14. Poirel L, Kieffer N, Brink A, Coetze J, Jayol A, Nordmann P. Genetic Features of MCR-1-Producing Colistin-Resistant Escherichia coli Isolates in South Africa. Antimicrob Agents Chemother. 2016; 60:4394–7. https://doi.org/10.1128/AAC.00444-16.
15. Liu YY, Chandler CE, Leung LM, McElheny CL, Mettus RT, Shanks RMQ, Liu JH, Goodlett DR, Ernst RK, Doi Y. Structural Modification of Lipopolysaccharide Conferred by mcr-1 in Gram-Negative ESKAPE Pathogens. Antimicrob Agents Chemother 2017; 61. https://doi.org/10.1128/AAC.00580-17.
16. Schwarz S, Johnson AP. Transferable resistance to colistin: a new but old threat. J Antimicrob Chemother. 2016; 71:2066–70. https://doi.org/10.1093/jac/dkw274.
17. Stoesser N, Mathers AJ, Moore CE, Day NP, Crook DW. Colistin resistance gene mcr-1 and pHNSHP45 plasmid in human isolates of Escherichia coli and Klebsiella pneumoniae. Lancet Infect Dis. 2016; 16:285–6. https://doi.org/10.1016/S1473-3099(16)00010-4.
18. Yang YQ, Zhang AY, Ma SZ, Kong LH, Li YX, Liu JX, Davis MA, Guo XY, Liu BH, Lei CW, Wang HN. Co-occurrence of mcr-1 and ESBL on a single plasmid in Salmonella enterica. J Antimicrob Chemother. 2016; 71:2336–8. https://doi.org/10.1093/jac/dkw243.
19. Pham Thanh D, Thanh Tuyen H, Nguyen Thi Nguyen T, Chung The H, Wick RR, Thwaites GE, Baker S, Holt KE. Inducible colistin resistance via a disrupted plasmid-borne mcr-1 gene in a 2008 Vietnamese Shigella sonnei isolate. J Antimicrob Chemother. 2016; 71:2314–7. https://doi.org/10.1093/jac/dkw173.
20. Wang Q, Sun J, Li J, Ding Y, Li XP, Lin J, Hassan B, Feng Y. Expanding landscapes of the diversified mcr-1-bearing plasmid reservoirs. Microbiome 2017; 5:70. https://doi.org/10.1186/s40168-017-0288-0.
21. Wang Q, Li Z, Lin J, Wang X, Deng X, Feng Y. Complex dissemination of the diversified mcr-1-harbouring plasmids in Escherichia coli of different sequence types. Oncotarget 2016; 7:82112–82122. https://doi.org/10.18632/oncotarget.12621.
22. Zurfluh K, Kieffer N, Poirel L, Nordmann P, Stephan R. Features of the mcr-1 Cassette Related to Colistin Resistance. Antimicrob Agents Chemother 2016; 60:6438–9. https://doi.org/10.1128/AAC.01519-16.
23. Zhang H, Jia Y, Xie X, Wang M, Zheng Y, Xu S, Zhang W, Wang Q, Huang X, Du H. RpoE promotes invasion and intracellular survival by regulating SPI-1 and SPI-2 in Salmonella enterica serovar Typhi. Future Microbiol. 2016; 11:1011–24. https://doi.org/10.2217/fmb.16.19.