
INTRODUCTION
Although the most important etiological agent in cervical cancer is human papillomavirus (HPV) infection, only a small proportion of infected women developed cervical cancer. HPV infection alone is insufficient to induce malignant changes. Copy number variation (CNV) is a very common phenomenon in cervical cancer and may be important in its pathogenesis [1]. Comparative genomic hybridization (CGH) studies for cervical cancer progression have shown that chromosome 3q gain was associated with the transition from pre-invasive to invasive cervical carcinoma. Subsequently, array-based CGH (aCGH), where arrays of genomic sequences replaced metaphase chromosomes as hybridization targets, was established. More detailed and precise genomic variations had been found in cervical cancer by using aGCH [2]. Lando et al. reported several potential driver genes for cervical carcinogenesis aCGH [3]. However, a recent study reported a limited correlation between chromosomal CNV and gene expression by single nucleotide polymorphisms array platform [4].
In this study, a high resolution Human Genome CGH Microarray Kit was used to detect genome-wide chromosomal CNV in 6 cervical cancer cell lines. We also performed gene expression studies in the cervical cancer cell lines, cervical carcinoma tissues and normal cervical epithelia by using Whole Human Genome Microarray Kit. Using appropriate bioinformatics software, we identified several chromosomal CNV regions and aberrantly expressed genes in cervical cancer. Statistical analysis and gene annotation analysis were also performed for the array data.
RESULTS
Fifty common chromosomal CNVs were identified in cervical cancer cell lines
Using aCGH at a genome-wide resolution of 250 kb, a total of 50 common chromosomal CNV regions were identified, ranging from 0.5 Mb to 80 Mb. Of these, 21 common amplification regions (including 13 significant amplification regions) and 29 deletion regions (including 2 significant deletion regions) were identified (Table 1).
Table 1: Physical location of the chromosome CNV regions identified by array CGH in cervical cancer cell lines
Region |
Region length |
Cytoband location |
Event |
Genes |
Frequency % |
P-value |
Reference |
|---|---|---|---|---|---|---|---|
chr1:10001-3752828 |
3,742,828 |
p36.33 - p36.32 |
Gain |
70 |
83.33333333 |
0.003 |
Gopeshwar Narayan, et al., 2007; Connie P. Matthews, et al., 2000; Y.W. CHOI, et al., 2007 |
chr2:75161025-85137285 |
9,976,261 |
p12 - p11.2 |
Loss |
17 |
83.33333333 |
0.006 |
Y.W. CHOI*, et al., 2007 |
chr2:137,395,646-170,227,851 |
32,832,206 |
q21.3 - q31.1 |
Loss |
84 |
100 |
>0.05 |
F.Y. Huang et al.2005; Y.W. CHOI, et al., 2007 |
chr2:178,374,598-197,908,813 |
19,534,216 |
q31.2 - q33.1 |
Loss |
67 |
83.33333333 |
>0.05 |
F.Y. Huang et al.2005; Y.W. CHOI, et al., 2007 |
chr2:209,391,516-216,053,786 |
6,662,271 |
q34 - q35 |
Loss |
15 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007; F.Y. Huang et al.2005; G Ng, et al., 2007 |
chr3:60,001-8,582,632 |
8,522,632 |
p26.3 - p25.3 |
Loss |
15 |
83.33333333 |
>0.05 |
F.Y. Huang et al.2005; Connie P. Matthews, et al., 2000; Y.W. CHOI, et al., 2007 |
chr3:16,297,340-37,460,259 |
21,162,920 |
p25.1 - p22.2 |
Loss |
60 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007; F.Y. Huang et al.2005; Connie P. Matthews, et al., 2000 |
chr3:58,690,297-90,181,487 |
31,491,191 |
p14.2 - p11.1 |
Loss |
52 |
83.33333333 |
0.028 |
Lockwood WW, et al., 2007; F.Y. Huang et al.2005; Connie P. Matthews, et al., 2000; Y.W. CHOI, et al., 2007; G Ng, et al., 2007 |
chr3:93,605,515-101,219,924 |
7,614,410 |
q11.2 - q12.3 |
Loss |
38 |
83.33333333 |
>0.05 |
N/A |
chr4:10,479,679-39,591,168 |
29,111,490 |
p16.1 - p14 |
Loss |
62 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007; F.Y. Huang et al.2005; Y.W. CHOI, et al., 2007; G Ng, et al., 2007 |
chr4:41,596,003-44,365,208 |
2,769,206 |
p13 |
Loss |
10 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007; F.Y. Huang et al.2005; G Ng, et al., 2007 |
chr4:58,367,789-139,871,504 |
81,503,716 |
q12 - q31.1 |
Loss |
300 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007; G Ng, et al., 2007 |
chr4:141,329,535-182,773,601 |
41,444,067 |
q31.1 - q34.3 |
Loss |
127 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007; Y.W. CHOI, et al., 2007; G Ng, et al., 2007 |
chr5:49,690,172-58,526,175 |
8,836,004 |
q11.1 - q11.2 |
Loss |
35 |
83.33333333 |
>0.05 |
Connie P. Matthews, et al., 2000 |
chr6:44,372,753-58,614,061 |
14,241,309 |
p21.1 - p11.2 |
Loss |
69 |
83.33333333 |
>0.05 |
Connie P. Matthews, et al., 2000; Y.W. CHOI, et al., 2007 |
chr6:61,982,931-73,725,450 |
11,742,520 |
q11.1 - q13 |
Loss |
16 |
83.33333333 |
>0.05 |
Connie P. Matthews, et al., 2000 |
chr6:75,067,155-105,246,238 |
30,179,084 |
q13 - q21 |
Loss |
83 |
83.33333333 |
>0.05 |
Connie P. Matthews, et al., 2000 |
chr6:112,398,800-148,256,165 |
35,857,366 |
q21 - q24.3 |
Loss |
143 |
83.33333333 |
>0.05 |
Connie P. Matthews, et al., 2000; Y.W. CHOI, et al., 2007 |
chr7:76,075,269-97,170,202 |
21,094,934 |
q11.23 - q21.3 |
Loss |
77 |
83.33333333 |
>0.05 |
N/A |
chr7:105,159,121-110,522,522 |
5,363,402 |
q22.3 - q31.1 |
Loss |
24 |
83.33333333 |
>0.05 |
N/A |
chr8:12,830,587-20,081,624 |
7,251,038 |
p22 - p21.3 |
Loss |
27 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007; G Ng, et al., 2007 |
chr8:75,336,800-85,510,468 |
10,173,669 |
q21.11 - q21.2 |
Loss |
27 |
83.33333333 |
>0.05 |
N/A |
chr8:142,141,881-146,304,022 |
4,162,142 |
q24.3 |
Gain |
92 |
83.33333333 |
0 |
Gopeshwar Narayan, et al., 2007; F.Y. Huang et al.2005; Connie P. Matthews, et al., 2000; Y.W. CHOI, et al., 2007; G Ng, et al., 2007 |
chr9:128,223,213-139,309,447 |
11,086,235 |
q33.3 - q34.3 |
Gain |
211 |
83.33333333 |
0.017 |
Lockwood WW, et al., 2007; Y.W. CHOI, et al., 2007 |
chr11:60,001-3,696,670 |
3,636,670 |
p15.5 - p15.4 |
Gain |
85 |
83.33333333 |
0 |
Y.W. CHOI, et al., 2007; G Ng, et al., 2007 |
chr11:20,658,997-31,795,373 |
11,136,377 |
p15.1 - p13 |
Loss |
27 |
83.33333333 |
>0.05 |
F.Y. Huang et al.2005 |
chr11:65,627,563-67,839,841 |
2,212,279 |
q13.1 - q13.2 |
Gain |
77 |
83.33333333 |
0.003 |
Y.W. CHOI, et al., 2007 |
chr12:38,766,104-42,827,028 |
4,060,925 |
q12 |
Loss |
12 |
83.33333333 |
>0.05 |
N/A |
chr13:45,697,630-55,761,181 |
10,063,552 |
q14.12 - q21.1 |
Loss |
56 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007; G Ng, et al., 2007 |
chr14:23,251,874-24,898,017 |
1,646,144 |
q11.2- q12 |
Gain |
64 |
83.33333333 |
0.001 |
N/A |
chr16:60,001-3,153,334 |
3,093,334 |
p13.3 |
Gain |
137 |
83.33333333 |
0.048 |
Y.W. CHOI, et al., 2007 |
chr16:3,970,244-5,071,063 |
1,100,820 |
p13.3 |
Gain |
23 |
83.33333333 |
0.048 |
Y.W. CHOI, et al., 2007 |
chr16:28,276,920-31,195,342 |
2,918,423 |
p11.2 |
Gain |
100 |
83.33333333 |
0.048 |
Y.W. CHOI, et al., 2007 |
chr16:66,279,668-70,782,910 |
4,503,243 |
q21 - q22.1 |
Gain |
103 |
83.33333333 |
0.021 |
Lockwood WW, et al., 2007; Connie P. Matthews, et al., 2000 |
chr16:83,959,097-90,294,753 |
6,335,657 |
q23.3 - q24.3 |
Gain |
78 |
83.33333333 |
0.021 |
Lockwood WW, et al., 2007; Connie P. Matthews, et al., 2000; Y.W. CHOI, et al., 2007 |
chr17:7,175,150-8,229,647 |
1,054,498 |
p13.1 |
Gain |
59 |
83.33333333 |
0.009 |
N/A |
chr17:72,693,870-81,060,000 |
8,366,131 |
q25.1 - q25.3 |
Gain |
166 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007; Y.W. CHOI, et al., 2007 |
chr18:18,510,899-43,242,321 |
24,731,423 |
q11.1 - q12.3 |
Loss |
70 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007; Y.W. CHOI, et al., 2007 |
chr18:62,450,433-71,009,737 |
8,559,305 |
q22.1 - q22.3 |
Loss |
11 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007; G Ng, et al., 2007 |
chr19:1,271,138-4,752,741 |
3,481,604 |
p13.3 |
Gain |
105 |
83.33333333 |
>0.05 |
Y.W. CHOI, et al., 2007 |
chr19:12,747,550-14,740,086 |
1,992,537 |
p13.13 - p13.12 |
Gain |
56 |
83.33333333 |
>0.05 |
Gopeshwar Narayan, et al., 2007 |
chr19:16,170,761-19,780,245 |
3,609,485 |
p13.12 - p13.11 |
Gain |
103 |
83.33333333 |
>0.05 |
N/A |
chr19:45,216,651-51,316,691 |
6,100,041 |
q13.32 - q13.33 |
Gain |
201 |
83.33333333 |
>0.05 |
Lockwood WW, et al., 2007 |
chr19:55,542,540-56,189,743 |
647,204 |
q13.42 |
Gain |
31 |
83.33333333 |
>0.05 |
N/A |
chr19:58,530,030-59,114,839 |
584,810 |
q13.43 |
Gain |
23 |
83.33333333 |
>0.05 |
N/A |
chr20:60,195,293-62,965,520 |
2,770,228 |
q13.33 |
Gain |
63 |
83.33333333 |
>0.05 |
Connie P. Matthews, et al., 2000; Y.W. CHOI, et al., 2007; G Ng, et al., 2007 |
chr21:14,417,523-32,339,619 |
17,922,097 |
q11.2 - q22.11 |
Loss |
61 |
83.33333333 |
>0.05 |
N/A |
chrX:77,966,491-93,063,726 |
15,097,236 |
q21.1 - q21.32 |
Loss |
25 |
83.33333333 |
>0.05 |
Connie P. Matthews, et al., 2000 |
chrX:120,138,580-127,769,411 |
7,630,832 |
q24 - q25 |
Loss |
10 |
83.33333333 |
>0.05 |
Connie P. Matthews, et al., 2000 |
chrX:152,449,419-153,711,912 |
1,262,494 |
q28 |
Gain |
47 |
83.33333333 |
0 |
Gopeshwar Narayan, et al., 2007; Connie P. Matthews, et al., 2000 |
Eleven of these (5 amplification regions and 6 deletion regions) have not been previously described in cervical cancer (Table 1). A total of 3514 genes were identified, and many tumor related genes, such as ABL1, BCL3, CDH1, CDKN1C, EPHA3, ERBB4, FOSL1, JUNB, MLH1, MYB, p53, RB1, ROS1, SKI, TGFBR1 and THRB, were located in these 50 chromosomal CNV regions.
Chromosomal CNVs could contribute to their transcript expression in cervical cancer
To evaluate if there is any association between chromosomal CNVs and gene expression changes in cervical cancer, we analyzed the gene expression profiles of the cervical cancer cell lines and normal cervical epithelium samples. 17.52% of transcripts (7,211 out of 41,152) exhibited a 2-fold over-expression and 9.02% of transcripts (3,712 out of 41,152) displayed a 2-fold down-regulation in 6 cervical cancer cell lines compared with 3 normal cervical epithelium tissues. Within the 21 common genomic amplification regions, 27.94% of the transcripts (772 out of 2,794) showed 2-fold over-expression. In the 13 significant amplification regions, the percentage was 29.56% (459 out of 1,553). In the 29 deletion regions, 10.46% (287 out of 2744) revealed 2-fold down-regulation. In the 2 significant deletion regions, the percentage was 11.67% (7 out of 60) (Figure 1A and 1B). Statistical analysis showed that gene up-regulation or down-regulation was significantly correlated with genomic amplification (P < 0.01, Spearsman correlation test) or deletion (P < 0.01) events. Thus, chromosomal CNVs can contribute to their transcript expression in cervical cancer. Two tumor related genes, ABL1 and p53, which were located in the genomic amplification regions, were found to be over-expressed by at least 2-fold in cervical cancer.
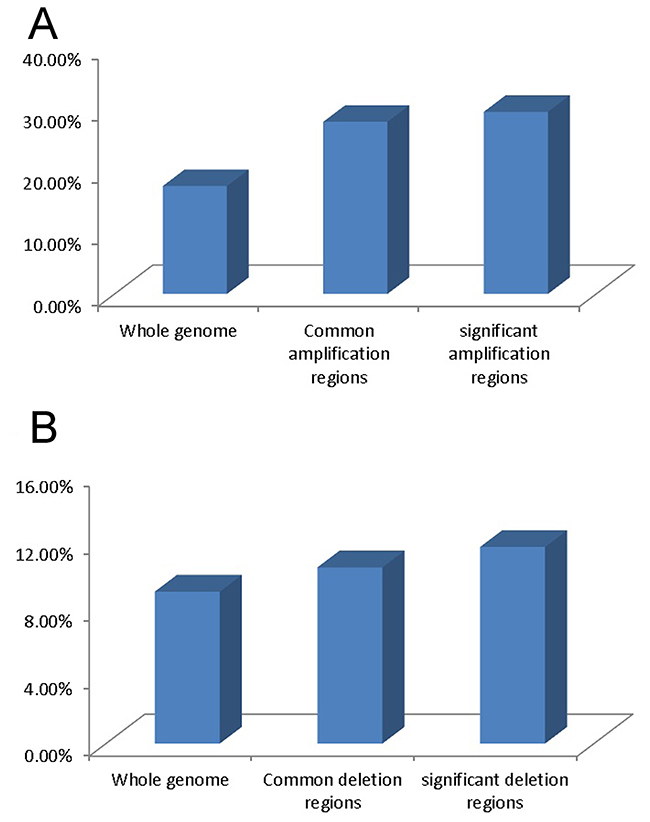
Figure 1: Gene expression variation in different genomic regions. (A) Percentage of up-regulated transcripts in whole genomic regions, common amplification regions and significant amplification regions. (B) Percentage of down-regulated transcripts in whole genomic regions, common deletion regions and significant deletion regions.
Profiles differed between cervical cancer cell lines, primary cervical carcinoma and normal cervical epithelium tissues
Expression profiles of transcripts across 6 different cervical cancer cell lines and 2 cervical carcinoma tissues and 3 normal, age-matched, cervical epithelium samples were analyzed using a hierarchical clustering algorithm (unsupervised K-means clustering). Cervical cancer cell lines, cervical carcinoma and normal cervical epithelium were divided two main groups (Figure 2A): cervical cancer cell lines for one group, and clinical cervical carcinoma and normal cervical epithelium for the other group. Interestingly, when gene tree clustering analysis was used to analyze genes with aberrant expression within the 50 common chromosome CNV regions and 15 significant chromosome CNV regions, similar results were obtained (Figure 2B and 2C)
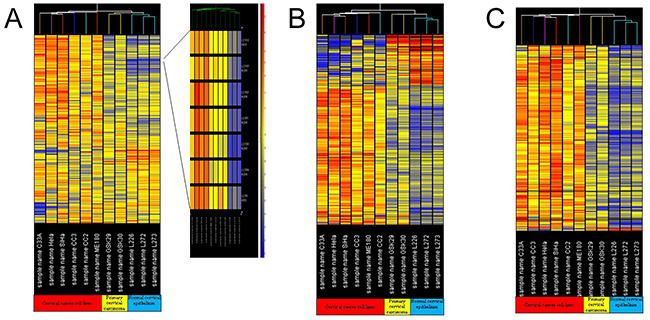
Figure 2: Gene tree clustering analysis. (A) Gene tree clustering analysis for the gene expression profiles of the cervical cancer cell lines, cervical carcinomas and normal cervical epithelium tissues; (B) gene tree clustering analysis for the aberrantly expressed genes in 50 common chromosomal CNV regions in cervical carcinoma; (C) gene tree clustering analysis for the aberrantly expressed genes in 15 significant chromosomal CNV regions in cervical carcinoma.
Gene ontology analysis for aberrantly expressed genes
By using a volcano plot in GeneSpring, 9,446 transcripts (27.4%) were found to be changed over two fold in the “Cancer group” (including 6 cervical cancer cell lines and 2 clinical cervical carcinomas) compared with the “Normal group” (including 3 cervical epithelium tissues). Among these genes, 6,001 transcripts were up-regulated by over 2 fold, and 3,445 transcripts were down-regulated by over 2 fold.
Pathway analysis showed that cell cycle pathways, cell communication pathways and DNA polymerase pathways were significantly affected pathways in cervical cancer
To further investigate the biological significance of these aberrantly expressed genes, pathway analysis was performed. The analysis results showed that cycle cycle pathways (P=1.15e-8), cell communication pathways (P=1.15e-8) and DNA polymerase pathways (P=1.15e-8) were significantly were affected in the cervical cancer (Table 2). Pathway analysis for the 446 differentially expressed transcripts in the 15 significant chromosomal CNV regions also showed that the cell cycle pathway involved the highest number of transcripts (including ABL1, DUSP9, E2F4, TP53, PKMYT1 and PPP1CA) (Figure 3), and the P value is 0.00062 (Table 3).
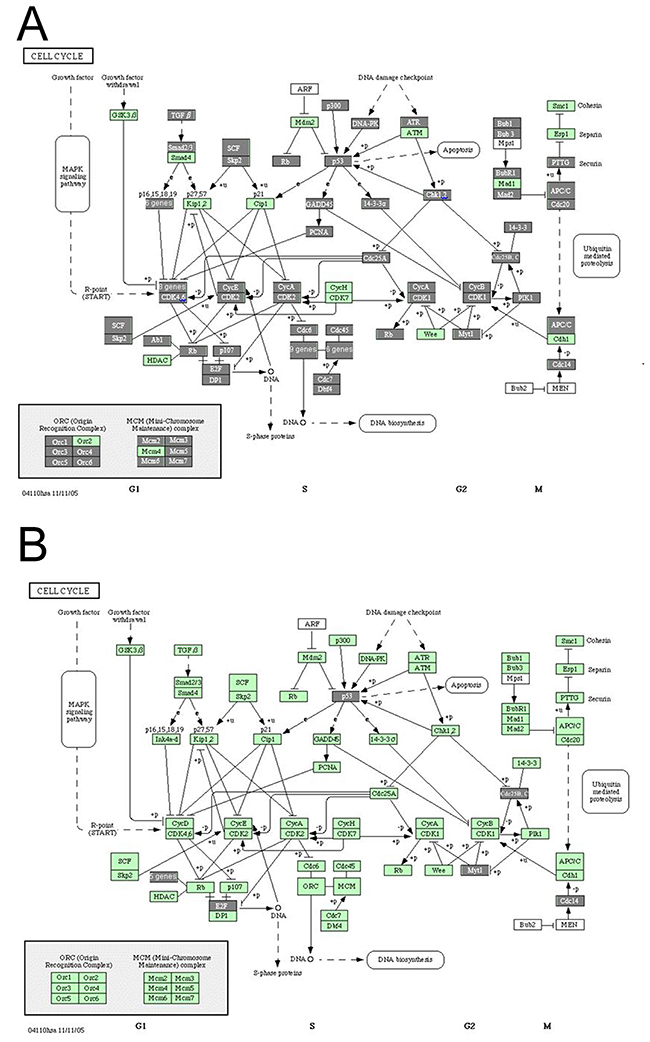
Figure 3: Cell cycle pathway analysis in cervical cancer. (A) Differently expressed genes involved in the cell cycle pathway in cervical carcinoma; (B) differently expressed genes within the significant chromosomal CNV regions involved in the cell cycle pathway in cervical carcinoma. Each rectangle represents one gene. The rectangle covered by gray color indicates that this gene is differently expressed in cervical carcinoma compared with normal cervix.
Table 2: Pathway analysis for the differentially expressed genes in cervical cancer
Pathway |
Number of genes with each pathway |
Genelist vs pathway random overlap p-value |
|---|---|---|
Cell cycle - Homo sapiens (human) |
111 |
1.15E-08 |
Proteasome - Homo sapiens (human) |
27 |
2.66E-08 |
One carbon pool by folate - Homo sapiens (human) |
22 |
5.11E-07 |
Cell Communication - Homo sapiens (human) |
60 |
5.85E-07 |
Pyrimidine metabolism - Homo sapiens (human) |
52 |
6.05E-06 |
Purine metabolism - Homo sapiens (human) |
77 |
2.27E-05 |
DNA polymerase - Homo sapiens (human) |
19 |
5.80E-05 |
Arginine and proline metabolism - Homo sapiens (human) |
35 |
0.000103 |
Riboflavin metabolism - Homo sapiens (human) |
14 |
0.00036 |
gamma-Hexachlorocyclohexane degradation - Homo sapiens (human) |
17 |
0.000515 |
Hematopoietic cell lineage - Homo sapiens (human) |
45 |
0.000654 |
Valine, leucine and isoleucine biosynthesis - Homo sapiens (human) |
10 |
0.000804 |
Glycosphingolipid biosynthesis - ganglioseries - Homo sapiens (human) |
13 |
0.000894 |
Pathogenic Escherichia coli infection - EHEC - Homo sapiens (human) |
33 |
0.000941 |
Pathogenic Escherichia coli infection - EPEC - Homo sapiens (human) |
33 |
0.000941 |
Selenoamino acid metabolism - Homo sapiens (human) |
23 |
0.00105 |
Valine, leucine and isoleucine degradation - Homo sapiens (human) |
31 |
0.00139 |
Urea cycle and metabolism of amino groups - Homo sapiens (human) |
17 |
0.00172 |
ECM-receptor interaction - Homo sapiens (human) |
46 |
0.00179 |
Glycan structures - biosynthesis 2 - Homo sapiens (human) |
36 |
0.00261 |
2,4-Dichlorobenzoate degradation - Homo sapiens (human) |
6 |
0.00264 |
Butanoate metabolism - Homo sapiens (human) |
24 |
0.00302 |
Lysine degradation - Homo sapiens (human) |
30 |
0.00449 |
Aminoacyl-tRNA biosynthesis - Homo sapiens (human) |
17 |
0.0047 |
Cell adhesion molecules (CAMs) - Homo sapiens (human) |
63 |
0.0062 |
Folate biosynthesis - Homo sapiens (human) |
22 |
0.00732 |
Ascorbate and aldarate metabolism - Homo sapiens (human) |
10 |
0.00742 |
Histidine metabolism - Homo sapiens (human) |
22 |
0.00918 |
Alkaloid biosynthesis II - Homo sapiens (human) |
14 |
0.0101 |
Limonene and pinene degradation - Homo sapiens (human) |
18 |
0.0102 |
Focal adhesion - Homo sapiens (human) |
91 |
0.0108 |
Methionine metabolism - Homo sapiens (human) |
11 |
0.0112 |
Oxidative phosphorylation - Homo sapiens (human) |
49 |
0.012 |
Citrate cycle (TCA cycle) - Homo sapiens (human) |
16 |
0.0153 |
Glycosaminoglycan degradation - Homo sapiens (human) |
11 |
0.016 |
Nitrobenzene degradation - Homo sapiens (human) |
10 |
0.0165 |
Olfactory transduction - Homo sapiens (human) |
17 |
0.0179 |
Glycolysis Gluconeogenesis - Homo sapiens (human) |
31 |
0.0197 |
Ethylbenzene degradation - Homo sapiens (human) |
11 |
0.0221 |
Ubiquitin mediated proteolysis - Homo sapiens (human) |
26 |
0.0236 |
Glyoxylate and dicarboxylate metabolism - Homo sapiens (human) |
9 |
0.0243 |
Arachidonic acid metabolism - Homo sapiens (human) |
25 |
0.0306 |
N-Glycan biosynthesis - Homo sapiens (human) |
21 |
0.033 |
Chondroitin sulfate biosynthesis - Homo sapiens (human) |
7 |
0.037 |
Linoleic acid metabolism - Homo sapiens (human) |
17 |
0.0415 |
Pentose phosphate pathway - Homo sapiens (human) |
14 |
0.0465 |
Apoptosis - Homo sapiens (human) |
38 |
0.0467 |
Protein export - Homo sapiens (human) |
8 |
0.0498 |
Table 3: Pathway analysis for the differentially expressed genes within the 15 significant chromosome CNV regions in cervical cancer
Pathway |
Number of common genes with each pathway |
Genelist vs pathway random overlap p-value |
|---|---|---|
Cell cycle - Homo sapiens (human) |
11 |
0.000622 |
Purine metabolism - Homo sapiens (human) |
7 |
0.0126 |
Axon guidance - Homo sapiens (human) |
6 |
0.0365 |
Insulin signaling pathway - Homo sapiens (human) |
6 |
0.0386 |
Selenoamino acid metabolism - Homo sapiens (human) |
5 |
0.000365 |
Tyrosine metabolism - Homo sapiens (human) |
5 |
0.00382 |
Glycerophospholipid metabolism - Homo sapiens (human) |
5 |
0.00419 |
Tryptophan metabolism - Homo sapiens (human) |
5 |
0.0137 |
Nitrobenzene degradation - Homo sapiens (human) |
4 |
0.000101 |
Aminophosphonate metabolism - Homo sapiens (human) |
4 |
0.000199 |
Histidine metabolism - Homo sapiens (human) |
4 |
0.00452 |
Androgen and estrogen metabolism - Homo sapiens (human) |
4 |
0.00747 |
Glycan structures - biosynthesis 2 - Homo sapiens (human) |
4 |
0.0281 |
Sulfur metabolism - Homo sapiens (human) |
3 |
0.000724 |
Ethylbenzene degradation - Homo sapiens (human) |
3 |
0.00309 |
RNA polymerase - Homo sapiens (human) |
3 |
0.00781 |
1- and 2-Methylnaphthalene degradation - Homo sapiens (human) |
3 |
0.0136 |
Benzoate degradation via CoA ligation - Homo sapiens (human) |
3 |
0.0153 |
Limonene and pinene degradation - Homo sapiens (human) |
3 |
0.0153 |
Pyruvate metabolism - Homo sapiens (human) |
3 |
0.0362 |
Bisphenol A degradation - Homo sapiens (human) |
2 |
0.0233 |
Glycosphingolipid biosynthesis - ganglioseries - Homo sapiens (human) |
2 |
0.03 |
Dentatorubropallidoluysian atrophy (DRPLA) - Homo sapiens (human) |
2 |
0.0323 |
Parkinson's disease - Homo sapiens (human) |
2 |
0.0323 |
DISCUSSION
In our genome-wide CNV analysis, we identified 50 frequently altered genomic regions (ranging from 0.5 Mb to 80 Mb), of which 11 have not been previously described in cervical cancer (Table 1). These differences in our results may be due to the different platforms of assay, different settings of analysis or the different cervical cancer cells. In these 50 commonly altered genomic regions, 3514 genes are included. Some of these, especially oncogenic or tumor suppressor genes, may be associated with the development of cervical cancer.
The gene tree clustering result suggested that during the development of cervical carcinoma, gene expression significantly changes, and cervical carcinoma can be distinguished from normal cervical epithelium tissue by clustering analysis of the gene expression profile. Since the cervical cancer cell lines were separate from primary cervical carcinoma and normal cervical epithelium tissue, we assumed that extended culturing of cervical cancer cell lines may also significantly alter their gene expression profiles.
The integrated analysis of genome-wide chromosomal copy number changes and gene expression profiling indicated that the identified CNVs could contribute to the expression of some but not all genes (Figure 1). This finding is consistent with a report by Vazquez-Mena et al. which used a different array platform to detect the correlation between CNVs and gene expression variation in cervical cancer cell lines. Other factors, such as epigenetic changes or transcription factors, may also contribute to variation of gene expression in cervical cancer [4]. Pathway analysis indicated that significant changes of some pathways, especially those involving the cell cycle, may be involved in the pathogenesis of cervical cancer.
ABL1 and p53, the two cell cycle pathway tumor-related genes which were located in the significant genomic amplification regions, were found to be overexpressed at least 2-fold in cervical cancer. ABL1, which was located in the chr9:128,223,213-139,309,447 genomic amplification region, plays a role in apoptosis. p53 is a well-known tumor suppressor gene, and an increase in p53 levels plays a critical role in the induction of genes that results in cell cycle arrest [7], allowing repair of damaged DNA or activation of apoptotic pathways [8]. In cervical cancer with high risk of HPV-infection, the E6 protein from high-risk HPV can bind to tumor suppressor protein p53 for rapid degradation via a cellular ubiquitin ligase [9]. Other studies have indicated that p53 protein over-expression is not common or associated with survival in cervical carcinoma [10, 11]. However, from our array CGH and gene expression array data, p53 was located in the chr17:7,175,150-8,229,647 genomic amplification region and was over-expressed at the mRNA level (the median of the expression of p53 in cervical cancer cell lines was 1.316 [from 0.82 to 3.38); the median of the expression of p53 in normal cervical epithelium was 0.295 [from 0.153 to 0.485]). This suggests that gene dosage of p53 contributes to RNA over-expression in some cervical cancers.
Our results demonstrated that over-expression of transforming growth factor-beta 1 (TGF-β1), a gene important in cell cycle pathways, may be due to a chromosomal CNV. TGF-β1 was amplified and was also over-expressed (>2 fold) in the cancer group compared with the normal group. TGF-β1 is involved in many different critical processes, such as embryonic development, cellular maturation and differentiation, wound healing, immune regulation and inflammation. TGF-β1 is a potent inhibitor of cell proliferation at the beginning of carcinogenesis [12, 13]. When cells become resistant to TGF-β1, tumor growth may be enhanced and metastasis promoted via immune evasion and angiogenesis. An increased expression of TGF-β1 has been found in cervical cancer. Kirma et al. suggested that TGF-β1 may be a factor in inducing over-expression of an oncogene, c-fms. Blocking c-fms has been demonstrated to result in increased apoptosis and decreased motility in cervical cancer [14].
Matrix metalloproteinases (MMPs) play an important role in the enhancement of tumor-induced angiogenesis. Our aCGH data showed that 9 MMP genes (MMP1, 3, 7, 8, 10, 12, 13, 20 and 27) located within 11q22 are amplified in Caski and SiHa cell lines, consistent with Lockwood’s findings. Further analysis revealed that MMP14, 23B and 25 were located in significant genomic amplification regions and MMP1, 15, 17 and TIMP1 in all our 6 cervical cancer cell lines. This is consistent with previous work reporting over-expression of MMP12 and MMP15 [15-19].
In summary, we have identified several chromosomal CNV regions and demonstrated that chromosomal CNVs are a common phenomenon which can affect the level of RNA expression in cervical cancer. Pathway analysis for the aberrantly expressed genes suggested that significant changes of some pathways, especially those involving the cell cycle, may contribute to the pathogenesis of cervical cancer. This study provided some clinical significance for us to have a better understanding of cervical cancer pathogenesis.
MATERIALS AND METHODS
Cervical cancer cell lines and specimens
Six human cervical cancer cell lines (HeLa, SiHa, C33A, ME180, CC2 and CC3) were used for aCGH and gene expression array. Five clinical specimens, including three normal cervical tissues and two cervical carcinoma specimens (FIGO stage: IIA), were collected from patients (aged between 36-42 years old) at the Department of Obstetrics and Gynaecology at the Prince of Wales Hospital in Hong Kong from January 2014 to December 2014. Informed consent was obtained from all participating subjects, and Institutional Review Board approval was obtained.
Tissue micro-dissection
Micro-dissection was used as described in our previous study [5]. Briefly, the tissue specimens were frozen in OCT cryomoulds (SAKURA, Japan), sectioned (8 μm) by a cryostat at -20°C (Leica Corp., CRYOCUT 1800), and then mounted onto glass slides (SAIL BRAND, Cat No 7105) at room temperature. Sections were stained by 0.1% methyl green (Sigma) and micro-dissected using a sterile surgical blade (AESCULAP) and collected immediately for further experiments.
Microarray comparative genomic hybridization analysis
Microarray comparative genomic hybridization using Human Genome CGH Microarray Kit (4x44K) (Agilent Technologies, Santa Clara, CA, USA) was used for identifying chromosomal CNV of 6 cervical cancer cell lines and 3 normal cervical samples as per the manufacturer’s protocol. Briefly, genomic DNA of cervical cancer cell lines was extracted using a DNeasy Blood & Tissue Kit (QIAGEN, Cat No. 69506). 1 μg genomic DNA of test sample and 1 μg human sex-matched control DNA as a reference sample (Promega G1521A; Lot no. 20929604) were digested using Alu I and Rsa I. This was followed by fluorescent labeling, clean-up of labeled genomic DNA, microarray hybridization and scanning. The data were extracted using the Agilent Feature Extraction (FE) v11.0 program. After calculating the background signal, non-uniform signal and the average raw signal on each probe, the resulting data files were generated and transferred to the bioinformatics software, Nexus Copy Number version 6.1 (BioDiscovery, Inc., El Segundo, CA, USA) for analysis [6].
Gene expression analysis
The Whole Human Genome Microarray Kit, 4x44K (G4112F) was used to probe gene expression in 6 cervical cancer cell lines, 2 cervical carcinomas and 3 normal cervical epithelium tissue samples. The resulting data files were generated and transferred to GeneSpring GX version 11.5 (Agilent Technologies, Santa Clara, CA, USA) for further analysis.
Reverse transcription PCR (RT-RCR)
RT-RCR was peformed for the identified gain and loss genes both on expression level and on genome level, normalized by GAPDH and B-globin (Forward: 5’-GAAGAGCCAAGGACAGGTAC-3’, Reverse: 5’-CAACTTCATCCACGTTCACC-3’), B2M (Forward:5’-TGCTGTCTCCATGTTTGATGTATCT-3’;Reverse:5’-TCTCTGCTCCCCACCTCTAAGT-3’), respectively.
Statistical analysis
For the Microarray Comparative Genomic Hybridization Analysis, 0.37 was used as the cut off value for amplification or 0.5 for deletion of a single probe. Putative chromosome copy number changes were defined by intervals of three or more adjacent probes with log2 ratios suggestive of a deletion or duplication when compared with the log2 ratios of adjacent probes. The p value for significant difference was set to less than 0.05 to reduce the false discovery rate (FDR). For Gene Expression Analysis, the cut-off value defining an aberrant change of gene expression was set at 2 fold for data analysis. Pathway analysis for the gene expression data was performed by GeneSpring GX version 11.5, and the pathway was downloaded from the KEGG database (ftp://ftp.genome.jp/pub/kegg/).
Abbreviations
CNV Copy number variations
HPV Human papillomavirus
CGH Comparative genomic hybridization
TGF-β1 Transforming growth factor-beta 1
MMPs Matrix metalloproteinases
FDR False discovery rate
Author contributions
T.T., Y.S. and T.Y. designed the research. Y.D. and Y.S. performed the experiments and wrote the manuscript. C.C.W., H.K.W., T.K.H.C., L.X.H. and K.W.C. revised the manuscript. All authors reviewed the manuscript.
CONFLICTS OF INTEREST
None of the authors of this manuscript report conflicts of interest regarding the performance of this study or its publication.
FUNDING
This study was supported by Basic Research for 2015 Shenzhen Municipal Science and Technology Programme (JCY20150630165236963), Shenzhen Science and Technology Innovation Project (JCYJ20140414114853648), the Innovation and Technology Fund of the Hong Kong Special Administrative Region, China (Ref No. UIM/283) and grants from the National Natural Science Foundation of China (grant No. 81660150 and 81372750).
REFERENCES
1. Bodelon C, Vinokurova S, Sampson JN, den Boon JA, Walker JL, Horswill MA, Korthauer K, Schiffman M, Sherman ME, Zuna RE, Mitchell J, Zhang X, Boland JF, et al. Chromosomal copy number alterations and HPV integration in cervical precancer and invasive cancer. Carcinogenesis. 2016; 37: 188-96.
2. Lockwood WW, Coe BP, Williams AC, MacAulay C, Lam WL. Whole genome tiling path array CGH analysis of segmental copy number alterations in cervical cancer cell lines. Int J Cancer. 2007; 120: 436-43.
3. Lando M, Holden M, Bergersen LC, Svendsrud DH, Stokke T, Sundfor K, Glad IK, Kristensen GB, Lyng H. Gene dosage, expression, and ontology analysis identifies driver genes in the carcinogenesis and chemoradioresistance of cervical cancer. PLoS Genet. 2009; 5: e1000719.
4. Vazquez-Mena O, Medina-Martinez I, Juarez-Torres E, Barron V, Espinosa A, Villegas-Sepulveda N, Gómez-Laguna L, Nieto-Martínez K, Orozco L, Roman-Basaure E, Muñoz Cortez S, Borges Ibañez M, Venegas-Vega C, et al. Amplified genes may be overexpressed, unchanged, or downregulated in cervical cancer cell lines. PLoS One. 2012; 7: e32667.
5. Tang T, Wong HK, Gu W, Yu MY, To KF, Wang CC, Wong YF, Cheung TH, Chung TK, Choy KW. MicroRNA-182 plays an onco-miRNA role in cervical cancer. Gynecol Oncol. 2013; 129: 199-208.
6. Tusher VG, Tibshirani R, Chu G. Significance analysis of microarrays applied to the ionizing radiation response. Proc Natl Acad Sci U S A. 2001; 98: 10515.
7. Orth JD, Loewer A, Lahav G, Mitchison TJ. Prolonged mitotic arrest triggers partial activation of apoptosis, resulting in DNA damage and p53 induction. Mol Biol Cell. 2012; 23: 567-76.
8. Wang X, Simpson ER, Brown KA. p53: Protection against tumor growth beyond effects on cell cycle and apoptosis. Cancer Res. 2015; 75: 5001-07.
9. Hiller T, Stubenrauch F, Iftner T. Isolation and functional analysis of five HPVE6 variants with respect to p53 degradation. J Med Virol. 2008; 80: 478-83.
10. Tommasino M, Accardi R, Caldeira S, Dong W, Malanchi I, Smet A, Zehbe I. The role of TP53 in cervical carcinogenesis. Hum Mutat. 2003; 21: 307-12.
11. Koyamatsu Y, Yokoyama M, Nakao Y, Fukuda K, Saito T, Matsukuma K, Iwasaka T.. A comparative analysis of human papillomavirus types 16 and 18 and expression of p53 gene and Ki-67 in cervical, vaginal, and vulvar carcinomas. Gynecol Oncol. 2003; 90: 547-51.
12. Mantel PY, Schmidt-Weber CB. Transforming growth factor-beta: recent advances on its role in immune tolerance. Methods Mol Biol. 2011; 677: 303-38.
13. Kloth JN, Fleuren GJ, Oosting J, de Menezes RX, Eilers PH, Kenter GG, Gorter A. Substantial changes in gene expression of Wnt, MAPK and TNFalpha pathways induced by TGF-beta1 in cervical cancer cell lines. Carcinogenesis. 2005; 26: 1493-502.
14. Kirma N, Hammes LS, Liu YG, Nair HB, Valente PT, Kumar S, Flowers LC, Tekmal RR. Elevated expression of the oncogene c-fms and its ligand, the macrophage colony-stimulating factor-1, in cervical cancer and the role of transforming growth factor-beta1 in inducing c-fms expression. Cancer Res. 2007; 67: 1918-26.
15. Luo CF, Zhu RQ, Wang H, Lu YL. Expression of COX-2 and MMP-9 in cervical carcinoma and their clinical significance. Zhonghua Zhong Liu Za Zhi. 2007; 29: 526-30.
16. Rauvala M, Aglund K, Puistola U, Turpeenniemi-Hujanen T, Horvath G, Willen R, Stendahl U. Matrix metalloproteinases-2 and -9 in cervical cancer: different roles in tumor progression. Int J Gynecol Cancer. 2006; 16: 1297-302.
17. Abraham R, Schafer J, Rothe M, Bange J, Knyazev P, Ullrich A. Identification of MMP-15 as an anti-apoptotic factor in cancer cells. J Biol Chem. 2005; 280: 34123-32.
18. Vazquez-Ortiz G, Pina-Sanchez P, Vazquez K, Duenas A, Taja L, Mendoza P, Garcia JA, Salcedo M. Overexpression of cathepsin F, matrix metalloproteinases 11 and 12 in cervical cancer. BMC Cancer. 2005; 5: 68.
19. Nasr M, Ayyad SB, El-Lamie IK, Mikhail MY. Expression of matrix metalloproteinase-2 in preinvasive and invasive carcinoma of the uterine cervix. Eur J Gynaecol Oncol. 2005; 26: 199-202.