
INTRODUCTION
Osteosarcoma (OS) is a malignant solid bone tumor that is highly malignant with low survival rates [1, 2]. Nearly 30-40% of the patients with localized osteosarcoma undergo relapse due to lung metastasis [3]. The survival rates of patients with metastatic OS is below 30%[4, 5]. The 5-year survival rate of OS patients is 55-70% despite improved surgical procedures and multidrug chemotherapy [6, 7]. Therefore, there is an urgent need to identify new therapeutic and prognostic biomarkers associated with osteosarcoma progression and metastasis.
Many cancers show mutations of receptor tyrosine kinesis (RTKs) resulting in aberrant Ras/ERK signaling [8]. The Ras/ERK signaling pathway regulates cellular survival, growth and differentiation [9]. Nearly 67% osteosarcomas show aberrant ERK activation [10]. Osteosarcomas also show upregulated RTKs such as EGFR [11]. Targeted inhibition of overexpressed or mutant RTKs has diminished therapeutic outcomes due to compensatory activation of other RTKs [12]. MEK inhibitors demonstrate therapeutic potential in human osteosarcoma cells due to constitutive ERK activation [3].
Nuclear division cycle 80 (Ndc80) is a kinetochore complex made up of two heterodimers, CDCA1-KNTC2 and SPC24-SPC25 [13]. It is essential for stable kinetochore-microtubule anchoring and normal chromosomal segregation during mitosis [14]. During mitosis, the SPC24/SPC25 protein complex anchors the Ndc80 complex to the inner kinetochore with the nuclear spindle microtubules and kinetochores [13, 15]. Disruption of spindle checkpoint proteins alters chromosomal segregation resulting in genetic instability and aneuploidy [16]. The spindle checkpoint proteins, SPC24 and SPC25 are implicated in colorectal and hepatocellular carcinomas [14, 17]. In this study, we investigated the role of SPC24 in osteosarcomagenesis.
RESULTS
SPC24 knockdown decreases growth, proliferation and colony formation
Western blot analysis revealed that U2OS and 143B cell lines transfected with three different SPC24 siRNAs showed lower SPC24 protein levels than controls (Figure 1A-1B). MTT assay demonstrated that SPC24-knockdown (siSPC24) U2OS and 143b cells showed decreased cell proliferation than control cells at 24, 48, and 72 h (Figure 1C-1D). Moreover, SPC24 knockdown U2OS and 143b cells showed decreased colony formation (Figure 1E-1F, Supplementary Figure 1A).
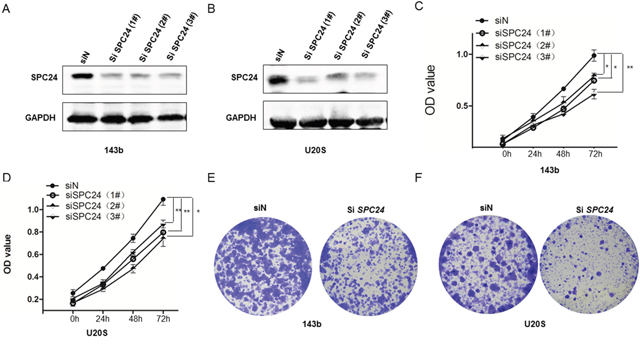
Figure 1: SPC24 knockdown decreases growth, viability and colony formation in 143B and U2OS cells. (A-B) Representative western blot images showing SPC24 expression in siSPC24 and siN transfected (A) 143B and (B) U2OS cells. β-actin is the loading control. (C-D) MTT assay showing viability of siSPC24 and siN transfected (C) 143B and (D) U2OS cells at 24, 48 and 72 h. (E-F) Soft-agar colony formation assay showing crystal violet stained colonies from siSPC24 and siN transfected (E) 143B and (F) U2OS cells. Note: * denotes p < 0.05 compared to control; **denotes p < 0.01 compared to control.
SPC24 knockdown promotes apoptosis and cell cycle arrest in OS cells
Dysregulated apoptosis is the hallmark of cancer cells. We evaluated the effects of SPC24 knockdown on OS cell apoptosis using AnnexinV/propidium iodide double staining. Flow cytometry analysis demonstrated that SPC24 knockdown increased early (AnnexinV+ PI−) and late (AnnexinV+ PI+) stage apoptosis (Figure 2A, Supplementary Figure 1B).
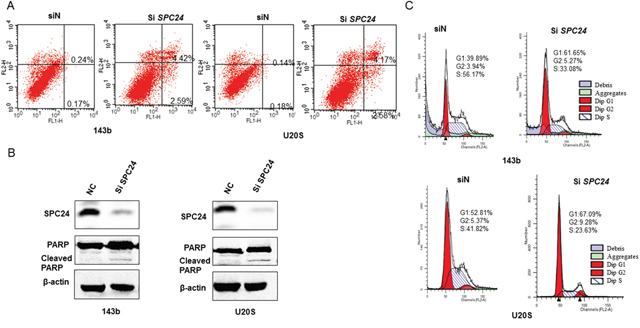
Figure 2: SPC24 knockdown promotes apoptosis and G1-S cell cycle arrest in 143B and U2OS cells. (A) Flow cytometry analysis of AnnexinV-FITC and propidium iodide stained control (siN) and SPC24 knockdown 143B and U2OS cells. AnnexinV+ PI- cells and AnnexinV+ PI+ cells represent early and late apoptotic cells, respectively. (B) Representative western blot showing SPC24 and PARP/cleaved PARP levels in control and siSPC24 transfected OS cells at 72 h. (C) Flow cytometry analysis of cell cycle distribution in control (siN) and SPC24 knockdown 143B and U2OS cells. Cells were stained with propidium iodide.
Moreover, SPC24 knockdown cells showed higher cleaved PARP levels than in control cells, further demonstrating increased apoptosis (Figure 2B).
Disrupted SPC24 results in uncontrolled mitosis because the cells are spindle checkpoint defective [18]. Therefore, we analyzed the cell cycle of SPC24 knockdown OS cells. Flow cytometry analysis showed that SPC24 knockdown demonstrated increased G1 phase cells and decreased S and G2-M phase cells than in control (siN) OS cells suggesting G1-S cell cycle arrest (Figure 2C and Supplementary Figure 1C).
SPC24 activated EGFR/Ras/Raf/MEK/ERK signal pathway
Pancreatic, colon, lung, ovarian and kidney tumors show aberrant epidermal growth factor receptor (EGFR) activation and MAP kinase signaling [19, 20]. We determined the effect of SPC24 knockdown on the EGFR/Ras/ERK signaling pathway. SPC24 knockdown decreased EGFR, Ras and p-ERK expression in OS cells than in controls (Figure 3A-3B). Howeve, the decreased level of p-ERK was not obvious with EGFR knockingdown when SPC24 was silenced (Figure 3C-3D).
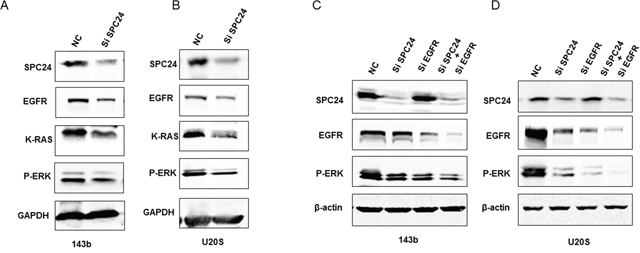
Figure 3: SPC24 knockdown inhibits EGFR/Ras/ERK signaling in 143B and U2OS cells. (A-B) Representative western blot showing SPC24, EGFR, K-Ras and p-ERK levels in siN or siSPC24 transfected (A) 143B and (B) U2OS cells. (C-D) Representative western blot showing SPC24, EGFR, K-Ras and p-ERK levels in siN or siEGFR transfected (A) 143B and (B) U2OS cells.
SPC24 knockdown decreases OS cell invasiveness and EMT
Next, we investigated the role of SPC24 in osteosarcoma metastasis by Transwell invasion assays. SPC24 knockdown in U2OS and 143b cells decreased cell migration (Figures 4A-4B and Supplementary Figure 1D).
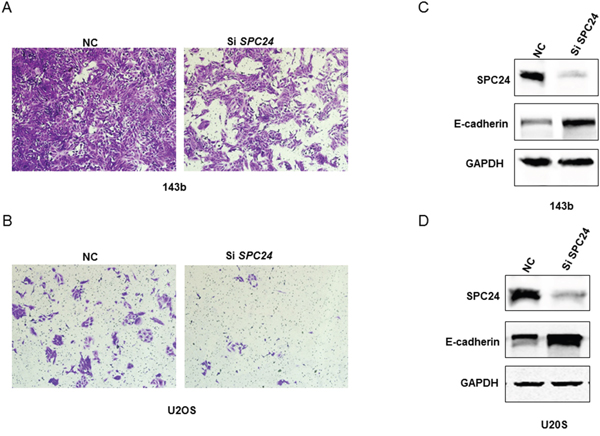
Figure 4: SPC24 knockdown increases invasiveness and EMT in 143B and U2OS cells. (A-B) Representative images (250X) showing transwell invasion assay results of siN or siSPC24 transfected 143B and U2OS cells. (C-D) Representative western blots showing E-cadherin and SPC24 levels in siN or siSPC24 transfected 143B and U2OS cells at 72 h.
E-cadherin plays a critical role in epithelial to mesenchymal transitioning (EMT), which is a key step in cancer metastasis and invasiveness. Therefore, we determined the effect of SPC24 knockdown on E-cadherin levels. SPC24 knockdown OS cells showed high E-cadherin expression than control cells (Figure 4C-4D). This suggested that SPC24 mediated osteosarcoma metastasis by downregulating E-cadherin.
SPC24 knockdown inhibits xenograft OS tumor growth in nude mice model
Next, we investigated the effects of SPC24 knockdown on in vivo osteosarcoma progression by xenografting control and SPC24 knockdown OS cells into dorsal flanking sites of 5 week old nude mice. We observed that the size and weight of shSPC24 tumors were highly reduced than shN tumors (p<0.01; Figure 5A-5D). Western bolt analysis demonstrated decreased EGFR and p-ERK as well as increased E-cadherin in shSPC24 tumors than shN tumors (Figure 5E-5F).
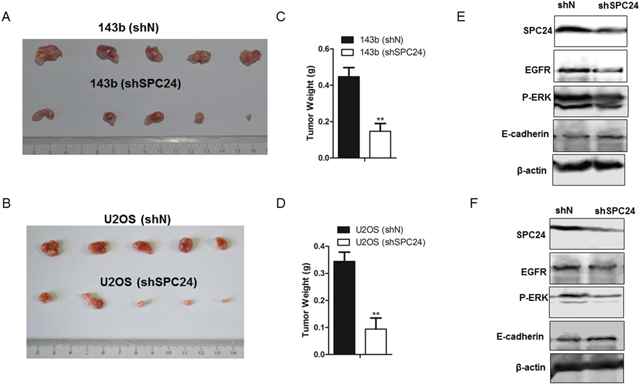
Figure 5: SPC24 knockdown inhibits osteosarcoma xenograft tumor growth in nude mice model. (A-B) Representative western blot analysis showing SPC24 expression in shN and shSPC24 transfected 143b and U2OS cells subcutaneously injected into 5-week old nude mice. (B) The tumor weights of xenograft tumors derived from control and SPC24 knockdown 143B and U2OS cells. Note: Values represent mean ± SEM; ** denotes p< 0.01 compared to control. (C-D) Representative western blot showing SPC24, EGFR, p-ERK and E-cadherin levels in xenograft tumors derived from control and SPC24 knockdown 143B and U2OS cells.
OS patient tissues show high SPC24 protein levels
Next, we analyzed the correlation between SPC24 and EGFR/Ras/ERK signaling pathway in human OS samples. OS patient tissue samples showed high SPC24 mRNA and protein expression than in normal tissues (Figure 6A-6B). Moreover, OS patient tissues showed increased SPC24 and p-ERK, which correlated with decreased E-cadherin based on IHC staining (Figure 6C-6D).
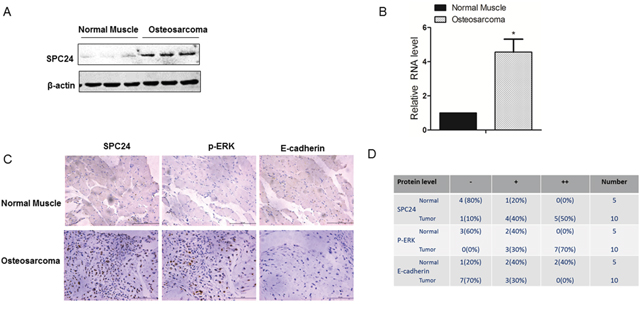
Figure 6: Human osteosarcoma patient tissues show increased SPC24 and p-ERK and decreased E-cadherin levels. (A) Representative western blot shows SPC24 levels in normal and OS patient tissue samples. B-actin was used as control. (B) QRT-PCR analysis of SPC24 mRNA levels in normal and OS patient tissue samples. (C) Representative images (magnification: 20x) show immunohistochemical staining of SPC24, p-ERK and E-cadherin in normal and OS tissue samples (Scale bar, 125μm). (D) Semiquantitative analysis of SPC24, p-ERK and E-cadherin expression in normal and OS cancer tissue samples based on immunohistochemical staining.
DISCUSSION
Osteosarcoma (OS) is the most common bone tumor. Osteosarcoma patients with pulmonary metastasis have a survival rate of <20% despite aggressive surgery and intensive chemotherapy [21]. Hence, identifying critical pathways regulating osteosarcoma metastasis is of paramount importance to develop novel therapeutic strategies. Human osteosarcoma xenograft models have been used to identify genetic factors and proteins that are associated with tumor progression and metastasis [22]. In this study, we demonstrate the oncogenic role of SPC24 in osteosarcomagenesis.
Disruption of spindle checkpoint proteins disrupts chromosomal segregation resulting in genetic instability and aneuploidy [16]. The changes in chromosomal copy number (aneuploidy) and aberrant chromosomal segregation contribute directly to the development and metastasis of malignant tumors [14, 23, 24]. SPC24 is a core component of the Ndc80 kinetochore complex, which is essential for directional movement of the chromosomes to the spindle poles during anaphase [25]. We demonstrate that SPC24 knockdown OS cell lines show decreased cell growth and proliferation and increased apoptosis.
Yu et al. reported that aberrant hyperactivation of the Ras/Raf/MEK/ERK pathway promoted lung metastasis in mice osteosarcoma model. Our study demonstrated that SPC24 activated EGFR/Ras/ERK signal pathway in osteosarcoma cells. The Ras/Raf/MEK/ERK pathway is upregulated in nearly 30% of human cancers [26, 27] and regulates metastasis in various cancers [28]. EMT is a key process that regulates cancer metastasis [29]. SPC knockdown in OS cells results in higher E-cadherin expression. These data demonstrate that SPC24 promotes invasiveness in OS by downregulating E-cadherin.
We also demonstrated that SPC24 knockdown OS cells reduced xenograft tumor growth. Moreover, they showed reduced p-ERK and higher E-cadherin levels. Furthermore, human OS samples showed high SPC24 and p-ERK expression as well as low E-cadherin. In conclusion, we demonstrate that SPC24 is a potential therapeutic target because it regulates OS progression and metastasis by activating EGFR/Ras/ERK signaling and downregulating E-cadherin.
MATERIALS AND METHODS
Cell lines
U2OS cells (ATCC) were grown in DMEM (Gibco) with 10% fetal bovine serum (FBS). 143B cells (Beijing Jishuitan Orthopaedic laboratory) were grown in RPMI 1640 (Hyclone) with 10% FBS.
SPC24 knockdown in U2OS and 143B cell lines
The stable SPC24 knockout U20S and 143B cell lines were generated by transfecting retroviral shRNA vectors specific for SPC24 (OriGene, Rockville, MD).
For transient SPC24 knockdown, 3 siRNAs against SPC24, (1) 5’-GAGCCUUCUCAAUGCGAAGTT-3; (2) 5’-CCGAGAAGCAGCUGCGAGATT-3’; and (3) 5’-UACCACCAA GUUAGUAAAATT-3’ [14] were transfected with TransExcellent (Cenji Biotech, Shanghai, China) according to manufacturer’s instructions. SPC24 mRNA and protein expression in the transfected cells was analyzed by qRT-PCR and western blot at 72 h.
Transwell invasion assay
Transwell matrigel assay was used to determine OS cell invasiveness as described previously [30]. In the upper part of the transwell chamber, 5×104 cells were seeded in DMEM or RPMI medium without FBS, whereas DMEM or RPMI medium with 10% FBS was added in the lower transwell chamber. After 24 h, the cells on the upper side of the transwell membrane were removed by cotton swabs. The migrating cells on the lower side of the insert filter were fixed with 10% buffered formaldehyde, stained with 1% crystal violet and quantified with a light microscope (Olympus).
MTT assay
OS cells (1 x 104 cells per 0.5 ml per well) were grown in 24-well plates at 37°C and 5% CO2. The cells were transiently transfected with control or SPC24 siRNAs for 24, 48, and72 h. At each time point, the cells were incubated with 500μg 3-(4,5-dimethyl-thiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT). Then, the MTT-formazan crystals were dissolved in 500 μl DMSO and the absorbance was measured at 570 nm in an ELISA reader.
Apoptosis assay
Cell apoptosis was determined by AnnexinV-FITC/PI Apoptosis Detection Kit (Abcam, Cambridge, MA, USA) according to the manufacturer’s protocols. The cells were washed in ice-cold PBS, re-suspended in 200 μl binding buffer and incubated with 5 μl Annexin V-FITC and 5 μl propidium iodide (PI) for 15 min at 4°C in the dark. Then, the cells were analyzed by flow cytometry.
Xenograft animal model
SPC24-knockdown and control cells (2×106 cells/100 μl) were injected into the dorsal flanking sites of 5 week old female BALB/c nude mice. After 3 weeks, the mice were euthanized and the tumors were harvested and used for both biochemical as well as immunohistochemical (IHC) staining. IHC staining was performed on paraffin embedded samples that were sectioned and stained as described.
Immunohistochemistry (IHC)
Serial paraffin embedded sections from tumor and normal tissues were deparaffinized in xylene and rehydrated in a graded ethanol series. Then, the sections were incubated with antibodies against SPC24, p-ERK and E-Cadherin overnight at 4°C. After PBS washes, the sections were incubated with HRP-conjugated secondary antibody for 1 h at room temperature. Then, the sections were developed with 3, 3- diaminobenzidine tetrahydrochloride (DAB) and counterstained with hematoxylin.
Statistical analysis
Data are presented as mean ± s.e.m. from at least three independent experiments. Difference between 2 groups was determined by student’s t-test. Difference at different time points among groups was evaluated by ANOVA. P < 0.05 was considered statistically significant.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
1. Lascelles BD, Dernell WS, Correa MT, Lafferty M, Devitt CM, Kuntz CA, Straw RC, Withrow SJ. Improved survival associated with postoperative wound infection in dogs treated with limb-salvage surgery for osteosarcoma. Ann Surg Oncol. 2005; 12:1073-1083.
2. Jaffe N. Osteosarcoma: review of the past, impact on the future. The American experience. Cancer Treat Res. 2009; 152:239-262.
3. Baranski Z, Booij TH, Kuijjer ML, de Jong Y, Cleton-Jansen AM, Price LS, van de Water B, Bovee JV, Hogendoorn PC, Danen EH. MEK inhibition induces apoptosis in osteosarcoma cells with constitutive ERK1/2 phosphorylation. Genes Cancer. 2015; 6:503-512. https://doi.org/10.18632/genesandcancer.91.
4. Meyers PA, Schwartz CL, Krailo M, Kleinerman ES, Betcher D, Bernstein ML, Conrad E, Ferguson W, Gebhardt M, Goorin AM, Harris MB, Healey J, Huvos A, et al. Osteosarcoma: a randomized, prospective trial of the addition of ifosfamide and/or muramyl tripeptide to cisplatin, doxorubicin, and high-dose methotrexate. J Clin Oncol. 2005; 23:2004-2011.
5. Ebb D, Meyers P, Grier H, Bernstein M, Gorlick R, Lipshultz SE, Krailo M, Devidas M, Barkauskas DA, Siegal GP, Ferguson WS, Letson GD, Marcus K, et al. Phase II trial of trastuzumab in combination with cytotoxic chemotherapy for treatment of metastatic osteosarcoma with human epidermal growth factor receptor 2 overexpression: a report from the children's oncology group. J Clin Oncol. 2012; 30:2545-2551.
6. Link MP, Goorin AM, Horowitz M, Meyer WH, Belasco J, Baker A, Ayala A, Shuster J. Adjuvant chemotherapy of high-grade osteosarcoma of the extremity. Updated results of the Multi-Institutional Osteosarcoma Study. Clin Orthop Relat Res. 1991; 8-14.
7. Provisor AJ, Ettinger LJ, Nachman JB, Krailo MD, Makley JT, Yunis EJ, Huvos AG, Betcher DL, Baum ES, Kisker CT, Miser JS. Treatment of nonmetastatic osteosarcoma of the extremity with preoperative and postoperative chemotherapy: a report from the Children's Cancer Group. J Clin Oncol. 1997; 15:76-84.
8. Schlessinger J. Receptor tyrosine kinases: legacy of the first two decades. Cold Spring Harb Perspect Biol. 2014; 6.
9. Zhang W, Liu HT. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002; 12:9-18.
10. Pignochino Y, Grignani G, Cavalloni G, Motta M, Tapparo M, Bruno S, Bottos A, Gammaitoni L, Migliardi G, Camussi G, Alberghini M, Torchio B, Ferrari S, et al. Sorafenib blocks tumour growth, angiogenesis and metastatic potential in preclinical models of osteosarcoma through a mechanism potentially involving the inhibition of ERK1/2, MCL-1 and ezrin pathways. Mol Cancer. 2009; 8:118.
11. Wen YH, Koeppen H, Garcia R, Chiriboga L, Tarlow BD, Peters BA, Eigenbrot C, Yee H, Steiner G, Greco MA. Epidermal growth factor receptor in osteosarcoma: expression and mutational analysis. Hum Pathol. 2007; 38:1184-1191.
12. Xu AM, Huang PH. Receptor tyrosine kinase coactivation networks in cancer. Cancer Res. 2010; 70:3857-3860.
13. Cheeseman IM, Desai A. Molecular architecture of the kinetochore-microtubule interface. Nat Rev Mol Cell Biol. 2008; 9:33-46.
14. Zhu P, Jin J, Liao Y, Li J, Yu XZ, Liao W, He S. A novel prognostic biomarker SPC24 up-regulated in hepatocellular carcinoma. Oncotarget. 2015; 6:41383-41397. https://doi.org/10.18632/oncotarget.5510.
15. DeLuca JG, Musacchio A. Structural organization of the kinetochore-microtubule interface. Curr Opin Cell Biol. 2012; 24:48-56.
16. Lee H. How chromosome mis-segregation leads to cancer: lessons from BubR1 mouse models. Mol Cells. 2014; 37:713-718.
17. Kaneko N, Miura K, Gu Z, Karasawa H, Ohnuma S, Sasaki H, Tsukamoto N, Yokoyama S, Yamamura A, Nagase H, Shibata C, Sasaki I, Horii A. siRNA-mediated knockdown against CDCA1 and KNTC2, both frequently overexpressed in colorectal and gastric cancers, suppresses cell proliferation and induces apoptosis. Biochem Biophys Res Commun. 2009; 390:1235-1240.
18. Bharadwaj R, Yu H. The spindle checkpoint, aneuploidy, and cancer. Oncogene. 2004; 23:2016-2027.
19. Markman B, Javier Ramos F, Capdevila J, Tabernero J. EGFR and KRAS in colorectal cancer. Adv Clin Chem. 2010; 51:71-119.
20. Hoshino R, Chatani Y, Yamori T, Tsuruo T, Oka H, Yoshida O, Shimada Y, Ari-i S, Wada H, Fujimoto J, Kohno M. Constitutive activation of the 41-/43-kDa mitogen-activated protein kinase signaling pathway in human tumors. Oncogene. 1999; 18:813-822.
21. Daw NC, Chou AJ, Jaffe N, Rao BN, Billups CA, Rodriguez-Galindo C, Meyers PA, Huh WW. Recurrent osteosarcoma with a single pulmonary metastasis: a multi-institutional review. Br J Cancer. 2015; 112:278-282.
22. Dass CR, Ek ET, Choong PF. Human xenograft osteosarcoma models with spontaneous metastasis in mice: clinical relevance and applicability for drug testing. J Cancer Res Clin Oncol. 2007; 133:193-198.
23. Pavelka N, Rancati G, Zhu J, Bradford WD, Saraf A, Florens L, Sanderson BW, Hattem GL, Li R. Aneuploidy confers quantitative proteome changes and phenotypic variation in budding yeast. Nature. 2010; 468:321-325.
24. Holland AJ, Cleveland DW. Boveri revisited: chromosomal instability, aneuploidy and tumorigenesis. Nat Rev Mol Cell Biol. 2009; 10:478-487.
25. McCleland ML, Kallio MJ, Barrett-Wilt GA, Kestner CA, Shabanowitz J, Hunt DF, Gorbsky GJ, Stukenberg PT. The vertebrate Ndc80 complex contains Spc24 and Spc25 homologs, which are required to establish and maintain kinetochore-microtubule attachment. Curr Biol. 2004; 14:131-137.
26. Wellbrock C, Karasarides M, Marais R. The RAF proteins take centre stage. Nat Rev Mol Cell Biol. 2004; 5:875-885.
27. Boldt S, Kolch W. Targeting MAPK signalling: Prometheus' fire or Pandora's box? Curr Pharm Des. 2004; 10:1885-1905.
28. Young A, Lyons J, Miller AL, Phan VT, Alarcon IR, McCormick F. Ras signaling and therapies. Adv Cancer Res. 2009; 102:1-17.
29. Onder TT, Gupta PB, Mani SA, Yang J, Lander ES, Weinberg RA. Loss of E-cadherin promotes metastasis via multiple downstream transcriptional pathways. Cancer Res. 2008; 68:3645-3654.
30. Liu KC, Huang AC, Wu PP, Lin HY, Chueh FS, Yang JS, Lu CC, Chiang JH, Meng M, Chung JG. Gallic acid suppresses the migration and invasion of PC-3 human prostate cancer cells via inhibition of matrix metalloproteinase-2 and -9 signaling pathways. Oncol Rep. 2011; 26:177-184.