
INTRODUCTION
Patients suffering of cancer often receive an external ionizing radiation therapy in association with surgery/chemotherapy or alone. This ionizing radiation can involve damages in tumor cells but also in healthy tissue in the radiation field. Radiation therapy induces several skin changes such as inflammation, edema, dermatitis, ulceration, late radiation-induced fibrosis (RIF), and necrosis [1, 2]. Radiation dose, fraction size and volume treated will vary the late side effects of radiation.
Radiation-induced fibrosis is marked by fibroblasts proliferation, myofibroblast differentiation, and synthesis of collagen, proteoglycans and extracellular matrix. [3, 4]. RIF can occur 4 to 12 months after radiation therapy, and this process is worsening with time and years. Every radiation-exposed part of the body can be affects by RIF, and the type of tissue exposed will influence the clinical presentation. RIF manifestations are multiple, such as skin induration and thickening, muscle shortening and atrophy, limited joint mobility, lymphedema, mucosal fibrosis, ulceration, fistula, hollow organ stenosis, and pain [5] and generally impact the quality of life [6, 7]. Fibrosis present an active retraction of the granulation tissue which is induced by contractile non-muscle cells, named myofibroblasts [8–10]. Fibroblasts and myofibroblasts are key effectors involved in the development of fibrosis due to excessive deposition of collagen and inappropriate extracellular matrix (ECM).
RIF is characterized by a DNA damage [11–14] and an inflammation [15–18]. Prolonged alteration of nuclear factor kappa B (NF-κB) pathway, a major effector of inflammation, leads to increase pro-fibrotic and pro-inflammatory cytokines which participate to the onset and the progression of RIF [6, 19, 20].
DNA damage and inflammation stimulate transforming growth factor β1 (TGF-β1) activity, which induces fibrosis mechanism [21, 22]. Furthermore, progression of radiation doses increases levels of TGF-β [14, 23].
TGF-β1 interacts with the canonical WNT/β-catenin pathway and peroxisome proliferator activated receptor γ (PPAR γ) which act in an opposite manner in several pathologies [24]. TGF-β1 stimulates myofibroblast differentiation by the activation of canonical WNT pathway and the downregulation of PPAR γ expression [25]. In response to TGF-β1, resident fibroblasts transdifferentiate into contractile myofibroblasts which express α -smooth muscle actin (α -SMA) and synthesize extracellular matrix proteins, particularly collagen.
We focus this review on the positive link between TGF-β1 and canonical WNT/β-catenin pathway and the opposite role of PPAR γ.
Radiation-induced fibrosis (RIF)
Radiation-induced fibrosis (RIF) is a major late effect which contributes to patient morbidity and occur in the skin, and subcutaneous tissue, lungs, gastrointestinal as well as any other organs in the treatment field. The mechanisms linking radiation to tissue sclerosis, fibrosis and atrophy are complex. In both tumors and normal tissues, radiation causes the induction of apoptosis or clonogenic cell death through free radical-mediated DNA damage [26]. In normal tissues, radiation toxicity induces changes in cell functions and causes activation of coagulation system, inflammation, epithelial regeneration, and tissue remodeling, which are precipitated by several signaling such as cytokines, chemokines and growth factors [27].
Etiology of RIF
Several factors can increase the risk of RIF. Total dose of radiotherapy and dose per fraction, volume of tissue treated, and time of treatment delivery are considered as primary factors. Increased levels of radiation dose, hypofractionation and increased field size are directly correlated with degree of RIF [28–32]. Patients with certain diseases are more susceptible to develop severe RIF, such as systemic scleroderma, systemic lupus erythematosus (SLE), or Marfan syndrome [33, 34]. Genetics factors have also a role in the predisposition to RIF [35, 36]. Single-nucleotide polymorphisms have been observed in genes encoding proteins such as TGF-β1, and superoxide dismutase 2 (SOD2) [37, 38]. Additional genes like IL18 (interleukin 18), MMP12 (matrix metalloproteinase 12), PER3 (period circadian protein homolog 3 protein), LTF (lactoferrin) stimulate the degradation of post-radiation ECM [39]. Several DNA modifications have been associated with RIF, like epigenetic modifications to DNA and histones [40]. Mitochondrial DNA damage enhance the removal of reactive oxygen species (ROS) [41].
Clinical presentation of RIF
RIF usually occurs 4 to12 months after radiation therapy and can progress over many years. The type of tissue exposed to irradiation is responsible for the clinical presentation. In general, RIF can manifest as skin induration and thickening, muscle shortening and atrophy, limited joint mobility, lymphedema, mucosal fibrosis, ulceration, fistula, hollow organ stenosis, and pain [5]. Other manifestations more regionally and specific include trismus, xerostomia, decreased vocal quality, osteoradionecrosis, dysphagia, and aspiration in patients with head and neck malignancy [42–47]; cervical plexopathy, brachial plexopathy, interstitial fibrosis, dyspnea, and oxygen requirement in patients with breast or lung malignancy [48, 49]; and urinary urgency, increased urinary frequency, diarrhea, loss of reproductive function, and dyspareunia in patients with abdominopelvic malignancy [50–52]. Currently, there is no uniform consensus to objectively quantify the degree of fibrosis in RIF [53].
Pathogenesis of RIF
Three histopathological phases of RIF are described. The prefibrotic phase shows chronic inflammation in which endothelial cells have a major role. The organized fibrosis phase contains a high density of myofibroblasts in an unorganized matrix adjacent to poorly cellularized fibrotic areas of senescent fibrocytes in a dense sclerotic matrix. The third phase named late fibroatrophic phase shows retractile fibrosis and gradual loss of parenchymal cells [54].
RIF is initially characterized by an injury which incites an acute response leading to inflammation, followed by the accumulation of fibroblasts, differentiation into myofibroblasts, and activation of extracellular matrix proteins like collagen [22]. Radiation induces direct DNA damages and the apparition of reactive oxygen species (ROS) [55] resulting in oxidative stress [56]. ROS involves interactions of ionizing radiation with water molecules and then the formation of free radicals such as superoxide, hydrogen peroxide and hydroxyl radical [57]. Hydroxyl radical production is responsible for the major part of damages [58, 59]. ROS generation and free radicals lead to a deterioration of cellular compounds such as DNA, RNA, proteins, lipids and membranes [58–60]. Superoxide dismutase, glutathione peroxidase and catalase control free radical damages [61]. Several studies have shown that a depletion of these enzymes stimulate oxidative stress [62–64]. During RT, injured cells lead to the release of chemoattractant molecules which can stimulate inflammation [55, 65, 66]. Furthermore, release of inflammatory cytokines and chemokines is exacerbated by thrombosis and ischemia [67, 68].
The first inflammatory cells which arrived at injured sites are neutrophils [69]. Neutrophils encounter fibronectin and collagen fragments and then lead to the release of inflammatory cytokines such as tumor necrosis factor alpha (TNF-α), interleukin 1 (IL-1), and interleukin 6 (IL-6) for the initiation of ROS and local inflammation [3, 70–74]. Theses inflammatory cytokines are correlated with high collagen deposition and with the onset of RIF [19, 75–78]. Monocytes and lymphocytes then interact with injured cells and stimulate the differentiation of monocytes into two subset of macrophages (M1 and M2) [79–81]. Subset M2 of macrophages secrete platelet-derived growth factor (PDGF) which stimulate the migration of fibroblast into injured tissue and the promotion of neo-angiogenesis [82]. Subset M2 of macrophages also secrete TGF-β, which is the main effector of Rif [83]. PDGF and TGF-β cascades are increased in lung tissues after RT [84–87].
TGF-β is responsible for the production of fibroblasts from bone marrow progenitors [88, 89] and for the differentiation of fibroblast into myofibroblasts [14]. The differentiation of fibroblasts results in activation of the expression of α-smooth muscle actin (α-SMA) which is responsible for the transformation of proto-myofibroblasts into matured myofibroblasts [90]. Fibrocytes (bone marrow-derived progenitor cells) and epithelial cells during epithelial-mesenchymal transition (EMT) can also be at the origin of myofibroblasts [91]. Activated myofibroblasts, by overexpression of TGF-β, secrete too much collagen, fibronectin and proteoglycans [92]. This phenomenon is responsible for stiffness and thickening of tissues under RT [68, 93]. In addition, TGF-β stimulates inhibitors of matrix metalloproteinases (MMP) and then decreases the activity of MMP-2 and MMP-9, which participates to the aggravation of excessive ECM deposition [94]. TGF-β stimulates ECM production and collagen architecture alterations [95, 96]. Excessive accumulation of collagen reduces the vascularization with time [68], although basic fibroblast growth factor (bFGF) is activated by myofibroblasts and participates for endothelial cell proliferation and neoangiogenesis [97]. Loss of cell functions, tissue atrophy, and then necrosis are induced by damages and gradual ischemia suffered by radiation-induced fibrosis [49, 65, 98–100]. However, the apparition of early fibrosis is not correlated with the development of lasted effects of RIF [101–103].
TGF-β and radiation therapy (cf. Figure 1)
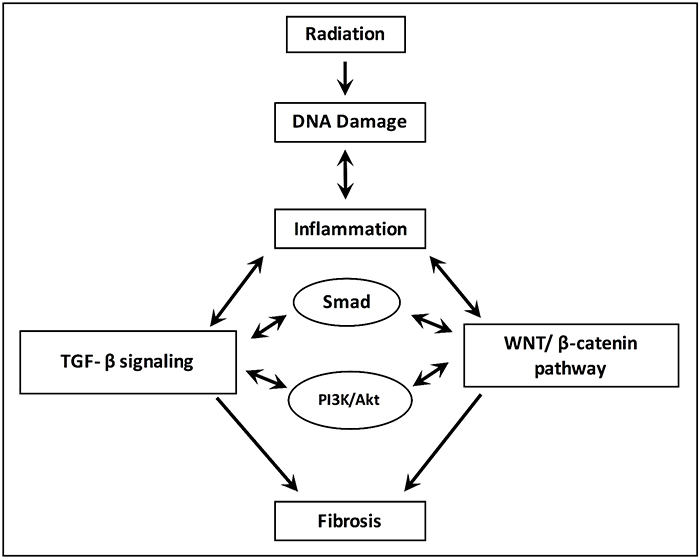
Figure 1: Processes for the development and progression of radiation-induced fibrosis.
Transforming growth factor β (TGF-β) is made of three similar proteins: TGF-β1, TGF-β2 and TGF-β3. TGF-β receptor 1 and TGF-β receptor 2 are transmembrane proteins. TGF-β1 binds TGF-βR2 which recruits TGF-βR1. TGF-β1 is secreted and then deposited into ECM and considered as a large latent complex. ECM is a main reservoir of cytokines [104]. TGF-β1 stimulates the Smad pathway and non-Smad pathways such as phosphatidylinositol 3-kinase/serine/threonine kinase (protein kinase B) (PI3K/Akt), Rho, and MAPK. Radiation-induced fibrosis is characterized by an upregulation of TGF-β1 [21, 23, 84, 105, 106].
TGF-β is responsible for the control of many functions, like breakdown of connective tissue, inhibition of epithelial cell proliferation and synthesis of extracellular matrix proteins (collagen and MMPs) [106].
Several studies have shown a weakly activation of TGF-β1 on day 1 after irradiation whereas its activation is strongly on day 14 after irradiation [106]. Thus, the irradiation-induced activation of TGF-β1 is rapid [107–110]. EMT is initiated by a prolonged irradiation exposure to TGF-β1 and this activates several transcriptional signaling pathways [111–113]. Radiation increases fibroblast differentiation-induced alpha-smooth muscle actin (α-SMA) activation [114]. Myofibroblast activation releases α-SMA or myofibroblasts activate the release α-SMA and α-SMA confers to myofibroblasts a strong contractile property [115, 116]. The interplay between TGF-β and several molecules, like extracellular signal-related kinase (EK), mitogen-activated protein kinase (MAPK), WNT, and PI3K, can explain the extent and the reversibility of EMT) [113].
A close relationship is observed between irradiation dose and liver cell viability [117]. Indeed, ionizing irradiation induces a progressive change in mRNA levels which gradually increase with time [118, 119]. Evolution of α-SMA in liver fibrosis induction suggests a time-dependent effect of irradiation [120, 121]. TGF-β1 expression gradually increases in a dose-dependent manner and peaking 4 weeks after irradiation [122].
The contractile non-muscle cells: myofibroblasts
Myofibroblasts contain actin filaments bundles with α-SMA. α-SMA represents peripheral focal adhesions and gap junctions for the connection between myofibroblats and granulation tissues [123]. Fibroblasts become protomyofibroblats after this interaction.
Protomyofibroblasts synthetize ECM, collagen, fibronectin which are essential for differentiation [124]. Then, protomyofibroblasts differentiate into myofibroblasts containing α-SMA. α-SMA is responsible for the retractile role of myofibroblasts [25]. Apoptosis is the final stage of myofibroblasts [125].
Mesenchymal stem cells (MSC) and fibroblasts are at the precursors of myofibroblasts, and these cells are found in normal tissues, like lung alveolar septa, uterine submucosa, lymph nodes, spleen, adrenal capsule, periodontal ligament, intestinal crypts and bone marrow stroma [115, 125].
Several diseases show myofibroblasts such as idiopathic pulmonary fibrosis and epithelial cancers [116]. In hypertrophic scar, myofibroblasts can persist after wound closure [126]. Precursors of myofibroblasts can be smooth muscle cells in coronary atheromatous plaque [127], perisinusoidal cells in liver [128], keratocytes in eyes [115], pericytes in kidneys [129], and bone marrow-derived fibrocytes [130]. Myofibroblasts are also observed in several pathological tissues like cancers (mammary carcinoma, epithelial cells in cancerous mammary glands), and fibrotic lesions [131].
Moreover, non-fibroblastic cell lineages [132–134] can differentiate into myofibroblasts through EMT process [135]. Differentiation of fibroblasts into myofibroblasts needs the participation of physical and chemical factors with increased stiffness of tissues [90, 136] and association of TGF-β1 with extra domain A (EDA) fibronectin [124, 137]. α-SMA increases contractile property of myofibroblasts and is synthesized through TGF-β1 activation [138]. The transmission of force generated by α-SMA and the molecular motor myosin are allowed by ECM through focal adhesions containing transmembrane integrins [139]. TGF-β1 is activated through an integrin-dependent process in ECM [126].
Myofibroblasts are contractile non-muscle cells, characterized by the type IIA non-muscle myosin (NMIIA) [140]. NMII are responsible for cell polarity, cell migration and cell-cell adhesion. NMIIA are highly observed in myofibroblasts of human placental stem villi [141, 142]. NMIIA is characterized by three pairs of chains, two heavy chains of 230 kDa, two 20 kDa regulatory light chains (RLCs) which stimulate the NMII activity and two 17 kDa essential light chains (ELCs) which stabilize the heavy chain structure. Calcium-calmodulin-myosin light chain kinase (MLCK) and Rho/ROCK/myosin light chain phosphatase increases NMII activity [90, 143, 144].
Myosin filaments link actin filaments in thick bundles like as stress fibers. NMMIIA molecules assemble into bipolar filaments. A title of the head enables a conformational change that moves actin filaments in an anti-parallel manner. The cross-bridge actin-myosin cycle of NMIIA is overall like smooth and striated muscle myosin. An ATP molecule binds the NMIIA-ATPase site on the myosin head. This allows the dissociation of actin from the NMIIA head. ATP is then hydrolyzed and subsequently, NMIIA binds to actin. Then, the power stoke occurs with a tilt of the NMIIA head, which generates a CB single force and a displacement of few nanometers. ADP is then released from the acto-NMIIA complex. A new ATP molecule dissociates actin from myosin head, and a novel CB cycle begins.
However, the major feature of NMIIA is its extreme slowness, with dramatic slow of its kinetics of contractile [145, 146]. The cross-bridge actin-myosin detachment constant, attachment constant, catalytic constant are widely low compared with striated or smooth muscles. Nevertheless, the cross-brigde actin-myosin of NMIIA single force is same order of magnitude compared with muscle myosin II (MII). Thermodynamic force, thermodynamic flow and thermodynamic entropy production rate are rarely low [147]. This explains why this stationary contractile system operates near-equilibrium. The low isometric tension observed in placental stem villi [141, 148] can be surely explained by the low placental myosin content [149, 150]. The extremely slow shortening velocity can be accounted for by the very low placental myosin ATPase activity which has an essential role for the association/dissociation of actin from the NMIIA head [145, 149]. In placental stem villi, a low isometric tension has been reported [147]. This can be explained by the low placental myosin content and the low placental myosin ATPase activity [148]. The acto-myosin apparatus functions as in smooth muscles in myofibroblasts of human placenta. In experimental bath, addition of KCl or electrical field induce the contraction phase. 2,3-butanedione monoxime, or isosorbide dinitrate, two inhibitors of NMII, can induce the relaxation phase [146]. Changes in the volume of the intervillons chamber can alter the length of the placental stem villi. Contraction of myofibroblasts induces a modulation of the distal resistance of the umbilical artery and then a modulation of the umbilical blood flow, due to the Starling phenomenon. In fibrotic processes, myofibroblasts generate rather a phenomenon of contraction-retraction lasting without relaxation and the pathological tissue undergoes an irreversible retraction, evolving towards fibrosis favored by the synthesis of collagen.
Canonical WNT/β-catenin pathway
Wingless and integration site (named WNT) pathway is a cascade of numerous signaling which are involved in development, metabolism, growth cellular, and maintain of stem cells [151]. WNT pathway is formed by secreted lipid-modified glycoproteins [152]. Deregulation of the canonical WNT pathway is observed in several pathologies [24].
WNT extracellular ligands bind Frizzled (FZD) receptors, low density lipoprotein receptor-related protein 5 and 6 (LRP 5/6) and disheveled (DSH), which lead to stimulate β-catenin accumulation and then the nuclear β-catenin translocation for the bind to T-cell factor/lymphoid enhancer factor (TCF/LEF) [153]. TCF/LEF related to nuclear β-catenin activates numerous target genes such as c-Myc, cyclin D1 [154].
Downregulation of the WNT pathway is characterized by the absence of binding between WNT extracellular ligands and the complex FZD/LRP 5/6. Thus, adenomatous polyposis coli (APC), AXIN and glycogen synthase kinase-3 (GSK-3β) form the β-catenin destruction complex and mediate degradation of β-catenin in the proteasome [155]. GSK-3β inhibits β-catenin accumulation and its nuclear translocation [155, 156].
WNT/β-catenin pathway is increased in fibroblasts in response to radiation during skin fibrogenesis [157]. WNT ligands are activated after irradiation and promote survival in head and neck cancers [158, 159]. Radiation stimulates ERK pathway through a reactive oxygen species (ROS) generation and inactivates GSK-3β [114]. Irradiation inhibits GSK-3β activity in mesenchymal cells [160] and increases WNT genes expression in fibroblasts [161]. Moreover, WNT/β-catenin pathway is overexpressed in liver, skin, lung, kidney, and heart presenting fibrosis [162–167].
Phosphatidylinositol 3-kinase/serine/threonine kinase (protein kinase B)/mammalian target of rapamycin (PI3K/Akt/mTOR) pathway is present in cell growth, proliferation, protein synthesis and energetic metabolism [168–171]. WNT/β-catenin pathway is considered as an upstream activator of PI3K/Akt/mTOR pathway [172] through the inhibition of GSK-3β [173]. In addition, diminution of bet-catenin signaling decreases the expression of PI3K/Akt/mTOR pathway [174, 175]. Moreover, in adipocyte differentiation, activated PI3K/Akt pathway inhibits GSK-3β [176, 177]. WNT/β-catenin pathway and PI3K/Akt/mTOR pathway mediate each other.
Inflammation and canonical WNT/β-catenin pathway
NF-κB signaling is a main effector of inflammation [178–180], its deregulation is implicated in numerous inflammatory processes [179–181]. Several studies have shown an interplay between canonical WNT/β-catenin pathway and NF-κB signaling [182]. Inflammation and immune response are modulated by this interaction between WNT/β-catenin pathway and NF-κB [182–185].
Overexpression of WNT/β-catenin pathway increases the NF-κB-mediated anti-apoptotic action [186, 187], and activates inflammatory processes through the stimulation of β-catenin targets genes [188, 189]. Activated β-catenin/TCF4 stimulates the NF-κB activity in vascular smooth muscle cells [190] and in colorectal cancer cells [191]. Activated WNT pathway in THP-1 cells induces release of inflammatory cytokines [192]. Nuclear activation of β-catenin associated with p50 and TNF-α, stimulates C-reactive protein (CRP) expression in 293T and HepG2 cells [193]. Nuclear β-catenin coupled with the TCF/LEF complex activates NF-κB target genes, such as CRP and matrix metalloproteinase 13 (MMP-13) in chondrocytes [194, 195], and then induces their genes transcription [193, 194]. In breast cancer, β-catenin/TCF and NF-κB signaling activate each other in a synergistic manner [196]. WNT/β-catenin pathway inhibits prolyl-hydroxylase and then activates NF-κB signaling [197, 198]. GSK3-β upregulation also decreases NF-κB signaling [199, 200].
Moreover, the NF-κB signaling activates canonical WNT/β-catenin pathway [201]. NF-κB activation stimulates the expression of the TCF/LEF complex and provides an indirect positive control of WNT/β-catenin pathway [202]. In human adipose tissue and bone marrow stroma cells, NF-κB decreases the expression of LZTS2 which inhibits β-catenin nuclear translocation and transcriptional activity [203, 204]. Stimulation of IKKB kinase (IKK) α, an activator of NF-κB, leads to cytosolic accumulation of β-catenin and then activates WNT/β-catenin pathway targets genes [201, 205]. Overexpression of interleukin 1 β (IL-1β) in mouse chondrocytes leads to a positive direct binding between NF-κB and LEF to induce β-catenin/LEF transcription [202]. NF-κB-induced TNF-α macrophages stimulates β-catenin cytosolic accumulation [206].
Ionizing radiation activates NF-κB which contributes to cell sensitivity to radiation [207]. NF-κB is activated in endothelial cells after irradiation and is necessary for radiation-induced IL-6 release [208]. Inflammation, through the activation of NF-κB, promotes the production of collagen and the release of inflammatory chemokines in the development of liver fibrosis [209]. NF-κB expression is stimulated by the activation of Akt pathway observed in RIF [210]. In fibrosis, CCN4, a WNT-inducible signaling pathway protein-1 (WISP1) is increased and participates to the fibroblast proliferation and ECM expression [211, 212]. CCN4 overexpression induces morphological transformation in skin fibrosis [213]. CCl4mAb, a specific inhibitor of CCN4, reduces NF-κB activity and then decreases the expression of pro-fibrotic factors, such as TGF-β1, in hepatic fibrosis [214].
Interactions between TGF-β1 and canonical WNT/β-catenin pathway (cf. Figure 2)
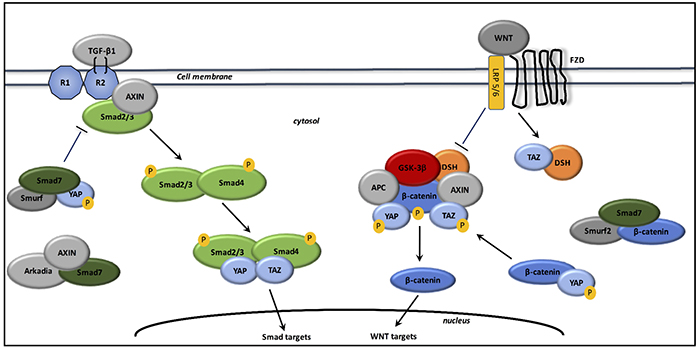
Figure 2: Interactions between TGF-β1, canonical WNT/β-catenin pathway, and YAP/TAZ signaling. In the absence of WNT ligands, Smad7 associates with β-catenin and Smurf2 to degrade β-catenin. Smad7 also binds phosphorylated YAP and Smurf to form a high affinity complex for TGF-β type 1 receptor to decrease TGF-β signaling expression. In the absence of WNT ligands, phosphorylated YAP and TAZ are associated with the β-catenin destruction complex. Phosphorylated YAP is also associated with β-catenin to inhibit its nuclear translocation and promote its degradation. Upon WNT stimulation, the beta-catenin destruction complex is inhibited and then beta-catenin accumulates in the cytosol and translocates to the nucleus. TAZ inhibits the phosphorylation of DSH and dissociates it from the β-catenin destruction complex. The destruction complex is inhibited because YAP and TAZ dissociate from the complex. β-catenin accumulates in the cytosol and then translocates to the nucleus for activating WNT targets. Upon TGF-β stimulation, Axin promotes the tail-phosphorylation of Smad2/3. Axin also forms a complex with Arkadia and Smad7 for enhancing TGF-β signaling. The activated Smad complex associates with TAZ and YAP and then translocates to the nucleus for activating Smad targets.
The canonical Smad pathway activates intracellular TGF-β1 signaling in activated myofibroblasts. TGF-β1 binds TGF-βR2 and then interacts with TGF-βR1 to form a heterotetramer for the phosphorylation of Smad2 and Smad3 which binds Smad4. The complex formed translocates to the nucleus for the activation of Smad binding element (SBE) DNA sequences and for recruit coactivators like histone acetyltransferase p300 [215]. Canonical WNT ligands, such as WNT3a, increase TGF-β1 through a Smad2 activation in a β-catenin-dependent manner and lead to myofibroblasts differentiation [216]. A non-Smad pathway is also observed and represented by MAPK, TGF-β activated kinase (TAK1), JNK, or PI3K/Akt [133, 217]. Phosphatase and tensin homolog (PTEN) is an inhibitor of PI3K/Akt pathway [218]. PTEN inhibits myofibroblasts differentiation, and the expression of collagen and α-SMA [219].
Fibroblasts stimulation with activated WNT3a enhances the expression of TGF-β and then the phosphorylation of the MH2 domain of Smad2 [216]. Conversely, the absence of WNT ligands decreases the expression of TGF-β and then attenuates the fibrotic response [220].
Without tail-phosphorylation by the TGF-β type I receptor, Smad2/3 cannot interact with Smad4 and then cannot engaged the DNA. Smads inactivated are phosphorylated by activated GSK-3β and then degraded [221]. Moreover, activation of TGF-β also enhances WNT pathway stimulation through the inhibition of dickkopf-related protein 1 (DKK1) [222].
Upon TGF-β and WNT stimulation, Axin facilitates the binding of Smad2/3 with TGF-β type 1 receptor and then activates Smad2/3 [223]. Moreover, upon WNT ligands activation, Axin forms a complex with Smad7 and the E3 ubiquitin ligase Arkadia to promote the degradation of Smad7 [224].
Smad7 is an inhibitor of Smad pathway [225]. Activated Smad7 binds YAP (yes-associated protein 1) and Smurf to increase affinity for TGF-β type 1 receptor, and then decreases TGF-β signaling [226]. Activated Smad7 also recruits Smurf 2 to induce ubiquitination and degradation of β-catenin [227].
Myofibroblast activation and fibrosis induction have been recently translated by the mechanical properties of YAP and TAZ. YAP and TAZ (transcriptional coactivator with PDZ-binding motif) elevated levels are observed in fibrosis [228]. YAP and TAZ knockdown in lung and liver fibroblasts cultured reduces the levels of protein associated with myofibroblast differentiation, such as pro-collagen and α-SMA [229]. During fibrosis, F-actin polymerization inactivates the Hippo core kinase complex leading to the dephosphorylation of YAP and TAZ [230].
Stimulation by WNT3a inhibits the destruction complex because YAP/TAZ dissociates from the β-catenin destruction complex and then allowed β-catenin nuclear translocation. TAZ binds DSH and dephosphorylates it upon stimulation with WNT3a to dissociate DSH from the destruction complex [231]. Indeed, in the absence of WNT ligands, phosphorylated YAP and TAZ bind β-catenin with activated GSK-3β and Axin to degraded β-catenin in the proteasome [232]. In fibrosis, activated Smad2/3-Smad4 complex is associated with YAP and TAZ for its translocation to the nucleus [233]. A crosstalk of several components of the TGF-β, WNT, and YAP/TAZ signaling is playing in the tuning of nucleocytoplasmic shuttling of fibrosis [230].
PPAR γ
Peroxisome proliferator receptor γ (PPAR γ) is a ligand activated transcriptional factor which forms a heterodimer with retinoid X receptor (RXR) to activate specific peroxisome response elements (PPRE) [234]. PPAR γ expression is involved in several mechanisms such as glucose and lipid metabolism, immune response, and inflammation [235, 236]. PPAR γ decreases NF-κB activity and then represses inflammation [237]. PPAR γ is expressed in several cells, such as adipocytes, muscle cells, brain cells, immune cells and in fibroblasts [238].
15d-prostaglandin J2 (15d-PGJ2), lysophosphatidic acid, and nitrolinoleic acid are natural activators of PPAR γ [239], whereas thiazolidinediones (TZDs) and oleanic acid derivatives such as triterpenoids (2-cyano-3,12-dioxoolean-1,9-dien-28-oic-acid (CDDO)) are synthetic activators of PPAR γ. PPAR γ expression mediates the functions of many signaling such as connective tissue regulation, mesenchymal cell activation, differentiation and cell survival creating a link between metabolism and fibrogenesis [134].
Numerous inflammatory cytokines, chemokines, or intracellular signaling decrease PPAR γ expression such as TFG-β1, canonical WNT/β-catenin pathway, TNF-α, interleukin (IL)-1, IL-13, Connective Tissue Growth Factor (CTGF), leptin, and lysophosphatidic acid (LPA) [240–242]. The transcription factor COUP II is a canonical WNT target and represses PPAR γ expression [243]. In adipocytes, adiponectin increases PPAR γ expression and decreases LPS-induced NF-κB expression and IL-6 production [244].
PPAR γ and fibrosis
An inverse relationship is observed between the expression of PPAR γ and the apparition of fibrosis. Aberrant downregulation of PPAR γ is correlated with the development of fibrosis in skin, lung, pancreas, heart, and liver [245]. PPAR γ□ expression is decreased in lung fibrosis [246], liver fibrosis [247], kidney fibrosis [248] and scarring alopecia fibrosis [249]. Furthermore, reduced PPAR γ expression precede fibrosis in several human diseases, suggesting a causal role [25, 250]. Fibroblasts, with a decreased level of PPAR γ, present an increase expression of TGF-β1, type 1 collagen, and α-SMA [251, 252].
Several studies have shown, that PPAR γ agonists can diminish pro-fibrotic signal-induced collagen synthesis and can blunt fibrosis [25, 241, 245, 251, 253–257]. PPAR γ natural and synthetic ligands, such as 15d-PGJ2 and rosiglitazone, can decrease the fibroblast-myofibroblast differentiation, synthesis of collagen and firbonectin and decrease the expression of TGF-β1 [247, 248, 253, 258, 259]. Moreover, 15d-PGJ2 and rosiglitazone decrease bleomycin-induced lung fibrosis [260]. PPAR γ agonists stop the TGF-β1-induced EMT of alveolar epithelial cells and decrease fibrosis in numerous organs such as heart [261, 262], lung [263, 264], liver [265, 266] and kidney [267, 268].
PPAR γ and radiation-induced fibrosis
Radiation therapy causes a downregulation of PPAR γ expression [59]. PPAR γ levels are decreased in 3 to 12 hours after a total body irradiation of C57BL/6 mice [59], after UVB irradiation in skin models [269] and in 3 days following a single abdominal dose of 10 Gy [270].
Oxidative stress and inflammation are involved in radiation-induced brain injury [271, 272]. Administration of PPAR γ agonists, such as pioglitazone, can decrease the severity of radiation-induced cognitive impairment in rat models [273]. PPAR γ ligands block the radiation-induced activation of NF-κB [274]. PPAR γ agonists administration can contribute to reduce the inflammation and oxidative stress [248, 275].
PPAR γ ligands enhance radiation-induced DNA damage in lung cancer cells in vitro [276, 277].
PPAR γ activators can prevent irradiation-induced inflammatory processes through the inhibition of NF-κB expression and the downregulation of STAT-3 pathway, an activator of canonical WNT/β-catenin pathway [270]. Diosmin, a citrus bioflavonoid which has antioxidant, anti-inflammatory and anti-apoptotic properties [278], can increase PPAR γ expression and can decrease canonical WNT/β-catenin pathway to attenuate radiation-induced hepatic fibrosis [9].
Interactions between PPAR γ and canonical WNT/β-catenin pathway
WNT/β-catenin pathway and PPAR γ act in an opposite manner in several diseases such as cancers [24, 279–284]. WNT/β-catenin pathway and PPAR γ interact through a TCF/LEF β-catenin domain and a catenin-binding domain within PPAR γ [285–288]. Downregulation of the WNT/β-catenin pathway induces PPAR γ stimulation while PPAR γ agonists decrease β-catenin expression in numerous cellular systems [235, 236, 289]. PPAR γ agonists can act as neuroprotective agents and promoting synaptic plasticity through a WNT/β-catenin/PI3K/Akt pathway interaction [290]. The regulation of mesenchymal stem cell differentiation also shows the existence of this crosstalk [291].
Indeed, in several diseases, β-catenin signaling inhibits PPAR γ [292–301], and using PPAR γ agonists is considered as promising treatment through this interplay [302]. Troglitazone, a PPAR γ agonist, can decrease c-Myc levels [303]. In intestinal fibrosis, the activation of WNT/β-catenin has been observed and the use of PPAR γ agonist can inhibit WNT/β-catenin pathway activation and then can repress fibrosis [304]. PPAR γ agonists stimulate Dickkopf-1 (DKK1) activity, which inhibits the canonical WNT pathway and then block fibroblast differentiation [305]. During adipogenesis in 3T3-L1 cells, PI3K/Akt pathway decreases PPAR γ expression [306]. Akt signaling participates to PPAR γ inhibition in adipocyte differentiation [307–309]. PI3K/Akt pathway phosphorylates GSK-3β and inhibits it, which decreases PPAR γ expression [176, 310]. β-catenin signaling, through its activation of Akt signaling, inhibits PPAR γ expression in adipocytes and 2T2-L1 preadipocytes [289, 311]. Moreover, inhibition of Akt pathway in 3T3-L1 cells activates the expression of PPAR γ [312]. PPAR γ agonists can decrease the activity of Akt signaling [313, 314]. Rosiglitazone and pioglitazone, PPAR γ agonists, inhibit both Akt signaling and GSK-3β activity in cardiac fibrosis to block TGF-β–induced collagen accumulation [315].
Interactions between PPAR γ and TGF-β1
TGF-β1 upregulation decreases PPAR γ expression in fibroblasts [245] and hepatic stellate cells [316], whereas PPAR γ agonists can directly inhibit TGF-β1 activity [317], prevent expression and synthesis of collagen in fibroblasts [253, 254, 257], and then prevent α-SMA expression [253, 257]. In addition, thiazolidinediones can reduce hepatic fibrosis through the inhibition of α-SMA and TGF-β expression [318]. PPAR γ agonists can inhibit both the Smad-dependent and Smad-independent TGF-β1 pathways.
Interactions through the Smad pathway
Anti-fibrotic PPAR γ effects can be largely explained through the Smad pathway. PPAR γ promoter has two Smad binding elements and inhibition of PPAR γ favors canonical Smad2/3 signaling [316].
In human hepatic stellate cells, PPAR γ agonists can inhibit TGF-β1/Smad3 signaling [319]. Smad-dependent collagen production is suppressed by PPAR γ ligand-activated through targeting the p300 transcriptional coactivator [254]. CDDO can prevent TGF-β1-induced fibrosis by inhibition of Smad transcription and Akt pathway [257]. TG-Interacting Factor (TGIF) which is a Smad transcriptional co-repressor can be activated by PPAR γ agonists, such as troglitazone, ciglitazone, and 15d-PGJ2. This leads to repress TGF-β1-induced fibrosis in hepatocyte cells [320, 321]. TGF-β1-induced JNK pathway is decreased by troglitazone, which decreases Smad2 signaling and then impairs myofibroblasts differentiation [322]. In addition, adiponectin prevents fibrosis in liver in mice [323, 324].
Moreover, a PPAR γ downregulation shows an increase of (increases) Smad3 and α-SMA expression and an overproduction of collagen (collagen overproduction) [252]. Rosiglitazone, a PPAR γ agonist, could decrease fibrosis by stimulating Smad7, an inhibitor of TGF/Smad signaling pathway [325]. In liver fibrosis, overexpression of prolyl oligopeptidase (POP) by S17092 (a POP inhibitor) leads to activate Smad7 protein and PPAR γ and then inactivates TGF-β signaling [326].
Interactions through the non-Smad pathway
PPAR γ agonists can induce diminution of TGF-β1expression in alveolar epithelial cells and in tumor metastasis without interacting with the Smad pathway [327, 328]. In an independent Smad pathway manner, PPAR γ represses TGF-β1-induced fibrosis [253, 258, 317]. TGF-β1 expression can be inhibited by 15d-PGJ2 through a PTEN-mediated p70 ribosomal S6 kinase-1 inhibition [329]. PPAR γ agonists can decrease TGF-β1-induced myofibroblast differentiation via the inhibition of PI3K/Akt pathway [330]. PPAR γ agonists can stimulate PTEN expression for having anti-fibrosis effects [219, 329, 331], and leading to the inhibition of collagen production and myofibroblast differentiation [332].
Interactions between PPAR γ and canonical WNT/β-catenin pathway/TGF-β1 in radiation-induced fibrosis (cf. Figure 3)
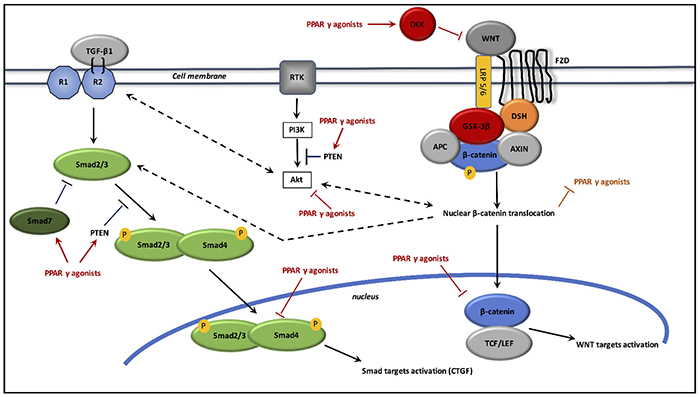
Figure 3: Interactions between TGF-β1, canonical WNT/β-catenin pathway and PPAR γ agonists in radiation-induced fibrosis. Inflammation activates WNT ligands. WNT ligand binds FZL and LRP5/6 receptors. This leads to inactivate the destruction complex AXIN/APC/GSK-3β. β-catenin phosphorylation is thus stopped, which prevents its degradation in the proteasome. Then, β-catenin accumulates in the cytosol and translocates to the nucleus for binding TCF/LEF co-transcription factor for inducing WNT target genes such as c-Myc and cyclin D1. WNT pathway and PI3K/Akt pathway increase each other. Inflammation and DNA damage also activate TGF-β1, which induces the Smad pathway. TGF-β1 binds type 2 TGF-β receptor, which recruits type 1 TGF-β receptor. The heterotetramer formed phosphorylates Smad2/3, which binds to Smad4. The Smad complex translocates to the nucleus for activation its target genes, such as CTGF. PPAR γ agonists activate Dickkopf-1 (DKK) for the inhibition of WNT ligands, and prevent β-catenin accumulation by activating GSK-3β. PPAR γ agonists inhibit Akt activity and stimulate PTEN, the inhibitor of PI3K. PPAR γ agonists also stimulate Smad7 and PTEN for inhibiting Smad pathway. Adenomatous polyposis coli (APC); dishevelled (DSH); frizzled (FZD); glycogen synthase kinase-3β (GSK-3β); low-density lipoprotein receptor-related protein 5/6 (LRP5/6); peroxisome proliferator-activated receptor γ (PPARγ); TGF-β1 receptors 1 and 2 (R1, R2); T-cell factor /lymphoid enhancer factor (TCF/LEF); transforming growth factor (TGF); phosphatase and tensin homolog (PTEN).
Several studies have shown a crosstalk between TGF-β1, canonical WNT/β-catenin pathway and PPAR γ [25, 250]. Indeed, canonical WNT/β-catenin is activated by TGF-β1, and this results in inhibition of PPAR γ. In liver fibrosis, TGF-β1 inhibits PPAR γ expression by increasing β-catenin expression [299]. In turn, activation of PPAR γ inhibits WNT/β-catenin pathway and then TGF-β1. PPAR γ targets PI3K/Akt for repress TGF-β1-induced myofibroblast differentiation [330]. PPAR γ agonists can inhibit fibrosis development through inactivation of TGF-β1 [333]. Moreover, 15-deoxy-delta12,14-prostaglandin J2, troglitazone, and rosiglitazone can suppress corneal myofibroblasts in eyes [294]. In addition, PPAR γ agonists can protect against excessive fibrosis [245]. Basic fibroblast growth factor (bFGF) can be repressed by using rosiglitazone and pioglitazone [313]. Inhibition of Akt pathway attenuates TGF-β signaling in rat kidney epithelial cells [334]. Troglitazone, a PPAR γ agonist, inhibits TGF-β signaling by phosphorylating Akt pathway at Ser437. LY294002, a PI3K inhibitor, inhibits Akt phosphorylation and α-SMA induction and then attenuates TGF-β1 signaling [335]. TGF-β-induced phosphorylation of β-catenin at Tyr654 associated with a conjunction between β-catenin and Smad3 stimulate α-SMA expression during EMT [336]. In fibrosis, troglitazone can inhibit TGF-β signaling by inhibiting β-catenin and PI3K/Akt pathway activation and by activating GSK-3β, the inhibitor of WNT pathway [335].
CONCLUSION
Radiation-induced fibrosis is characterized by DNA damage and inflammation. These two processes lead to the activation of TGF-β1 and canonical WNT/β-catenin pathway. TGF-β1 plays a major role in the differentiation of fibroblasts into myofibroblasts. Myofibroblasts appear able to physiological contraction and relaxation. However, in fibrosis myofibroblasts generate a phenomenon of contraction-retraction lasting without relaxation and with an irreversible retraction favored by the synthesis of collagen. Myofibroblasts show a main role in cellular fibrosis in numerous organs such as kidney, heart, lung, liver and wound. TGF-β1 operates in a canonical WNT/β-catenin pathway dependent manner. These two pathways stimulate each other through the Smad pathway or non-Smad pathways like PI3K/Akt pathway. TGF-β1 stimulates myofibroblast differentiation by the stimulation of canonical WNT pathway and the downregulation of PPAR γ expression. TGF-β1 appears to be an interesting therapeutic target in fibrosis [337, 338]. PPAR γ agonists stimulate Smad7 for inhibiting Smads pathway what blocks TGF-β signaling. PPAR γ agonist can decrease canonical WNT/β-catenin pathway by activating both Smad 7, GSK-3β and DKK. PPAR γ agonists can also stimulate PTEN, an inhibitor of PI3K/Akt pathway and inhibit the Smad pathway for downregulation TGF-β1 expression. Thus, by both inhibiting TGF-β1 and the canonical WNT/β-catenin pathway, PPAR γ agonists could be interesting preventive targets for radiation-induced fibrosis treatment (cf. Figure 4).
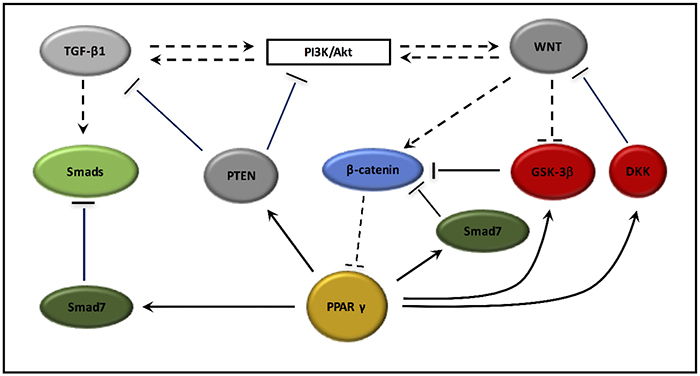
Figure 4: PPAR γ interactions with TGF-β1, PI3K/Akt pathway and the canonical WNT/β-catenin pathway. PPAR γ agonists activate Smad7, an inhibitor of Smads pathway. PPAR γ agonists activate PTEN, which inhibits both PI3K/Akt pathway and TGF-β1 signaling. PPAR γ agonists activate DKK and Smad7, two inhibitors of WNT/beta-catenin pathway leading to the β-catenin degradation in the proteasome. PPAR γ agonists activate GSK-3β activity, which forms the destruction complex to degrade β-catenin. In contrast, presence of WNT ligands inhibits GSK-3β and lead to cytosolic accumulation of β-catenin. Then nuclear β-catenin associates with TCF/LEF in the nucleus to inhibit PPAR γ expression.
Abbreviations
APC: adenomatous polyposis coli; DSH: disheveled; FZD: frizzled; GSK-3β: glycogen synthase kinase-3β; LRP 5/6: low-density lipoprotein receptor-related protein 5/6; PI3K-Akt: phosphatidylinositol 3-kinase-protein kinase B; PPAR γ: peroxisome proliferator-activated receptor γ; RIF: radiation-induced fibrosis; TCF/LEF: T-cell factor/lymphoid enhancer factor; TGF-β: transforming growth factor β.
Author contributions
All the authors have contributed equally to this review.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
1. Harper JL, Franklin LE, Jenrette JM, Aguero EG. Skin toxicity during breast irradiation: pathophysiology and management. South Med J. 2004; 97: 989–93. https://doi.org/10.1097/01.SMJ.0000140866.97278.87.
2. Pires AM, Segreto RA, Segreto HR. RTOG criteria to evaluate acute skin reaction and its risk factors in patients with breast cancer submitted to radiotherapy. Rev Lat Am Enfermagem. 2008; 16: 844–9.
3. Wynn TA. Fibrotic disease and the T(H)1/T(H)2 paradigm. Nat Rev Immunol. 2004; 4: 583–94. https://doi.org/10.1038/nri1412.
4. Wynn TA. Cellular and molecular mechanisms of fibrosis. J Pathol. 2008; 214: 199–210. https://doi.org/10.1002/path.2277.
5. Dörr W, Hendry JH. Consequential late effects in normal tissues. Radiother Oncol. 2001; 61: 223–31.
6. Flanders KC, Sullivan CD, Fujii M, Sowers A, Anzano MA, Arabshahi A, Major C, Deng C, Russo A, Mitchell JB, Roberts AB. Mice lacking Smad3 are protected against cutaneous injury induced by ionizing radiation. Am J Pathol. 2002; 160: 1057–68. https://doi.org/10.1016/S0002-9440(10)64926-7.
7. Müller K, Meineke V. Radiation-induced alterations in cytokine production by skin cells. Exp Hematol. 2007; 35: 96–104. https://doi.org/10.1016/j.exphem.2007.01.017.
8. Gabbiani G, Hirschel BJ, Ryan GB, Statkov PR, Majno G. Granulation tissue as a contractile organ. A study of structure and function. J Exp Med. 1972; 135: 719–34.
9. Hasan HF, Abdel-Rafei MK, Galal SM. Diosmin attenuates radiation-induced hepatic fibrosis by boosting PPAR-γ expression and hampering miR-17-5p-activated canonical Wnt-β-catenin signaling. Biochem Cell Biol. 2017; 95: 400–14. https://doi.org/10.1139/bcb-2016-0142.
10. Zampieri F, Coen M, Gabbiani G. The prehistory of the cytoskeleton concept. Cytoskelet (Hoboken). 2014; 71: 464–71. https://doi.org/10.1002/cm.21177.
11. Della Latta V, Cecchettini A, Del Ry S, Morales MA. Bleomycin in the setting of lung fibrosis induction: from biological mechanisms to counteractions. Pharmacol Res. 2015; 97: 122–30. https://doi.org/10.1016/j.phrs.2015.04.012.
12. Kunwar A, Haston CK. DNA damage at respiratory distress, but not acute time-points, correlates with tissue fibrosis following thoracic radiation exposure in mice. Int J Radiat Biol. 2015; 91: 360–7. https://doi.org/10.3109/09553002.2015.997897.
13. Povedano JM, Martinez P, Flores JM, Mulero F, Blasco MA. Mice with pulmonary fibrosis driven by telomere dysfunction. Cell Rep. 2015; 12: 286–99. https://doi.org/10.1016/j.celrep.2015.06.028.
14. Yarnold J, Brotons MC. Pathogenetic mechanisms in radiation fibrosis. Radiother Oncol. 2010; 97: 149–61. https://doi.org/10.1016/j.radonc.2010.09.002.
15. Chitra P, Saiprasad G, Manikandan R, Sudhandiran G. Berberine attenuates bleomycin induced pulmonary toxicity and fibrosis via suppressing NF-κB dependant TGF-β activation: a biphasic experimental study. Toxicol Lett. 2013; 219: 178–93. https://doi.org/10.1016/j.toxlet.2013.03.009.
16. Fox J, Haston CK. CXC receptor 1 and 2 and neutrophil elastase inhibitors alter radiation-induced lung disease in the mouse. Int J Radiat Oncol Biol Phys. 2013; 85: 215–22. https://doi.org/10.1016/j.ijrobp.2012.02.024.
17. Graves PR, Siddiqui F, Anscher MS, Movsas B. Radiation pulmonary toxicity: from mechanisms to management. Semin Radiat Oncol. 2010; 20: 201–7. https://doi.org/10.1016/j.semradonc.2010.01.010.
18. Paun A, Kunwar A, Haston CK. Acute adaptive immune response correlates with late radiation-induced pulmonary fibrosis in mice. Radiat Oncol. 2015; 10: 45. https://doi.org/10.1186/s13014-015-0359-y.
19. Johnston CJ, Wright TW, Rubin P, Finkelstein JN. Alterations in the expression of chemokine mRNA levels in fibrosis-resistant and -sensitive mice after thoracic irradiation. Exp Lung Res. 1998; 24: 321–37. https://doi.org/10.3109/01902149809041538.
20. Johnston CJ, Williams JP, Okunieff P, Finkelstein JN. Radiation-induced pulmonary fibrosis: examination of chemokine and chemokine receptor families. Radiat Res. 2002; 157: 256–65.
21. Koerdt S, Rohleder NH, Rommel N, Nobis C, Stoeckelhuber M, Pigorsch S, Duma MN, Wolff KD, Kesting MR. An expression analysis of markers of radiation-induced skin fibrosis and angiogenesis in wound healing disorders of the head and neck. Radiat Oncol. 2015; 10: 202. https://doi.org/10.1186/s13014-015-0508-3.
22. Straub JM, New J, Hamilton CD, Lominska C, Shnayder Y, Thomas SM. Radiation-induced fibrosis: mechanisms and implications for therapy. J Cancer Res Clin Oncol. 2015; 141: 1985–94. https://doi.org/10.1007/s00432-015-1974-6.
23. de Andrade CB, Ramos IP, de Moraes AC, do Nascimento AL, Salata C, Goldenberg RC, de Carvalho JJ, de Almeida CE. Radiotherapy-induced skin reactions induce fibrosis mediated by TGF-β1 ytokine. Dose Response. 2017; 15. https://doi.org/10.1177/1559325817705019.
24. Lecarpentier Y, Claes V, Duthoit G, Hébert JL. Circadian rhythms, Wnt/beta-catenin pathway and PPAR alpha/gamma profiles in diseases with primary or secondary cardiac dysfunction. Front Physiol. 2014; 5: 429. https://doi.org/10.3389/fphys.2014.00429.
25. Wei J, Bhattacharyya S, Jain M, Varga J. Regulation of matrix remodeling by peroxisome proliferator-activated receptor-γ: a novel link between metabolism and fibrogenesis. Open Rheumatol J. 2012; 6: 103–15. https://doi.org/10.2174/1874312901206010103.
26. Stubblefield MD. Radiation fibrosis syndrome: neuromuscular and musculoskeletal complications in cancer survivors. PM R. 2011; 3: 1041–54. https://doi.org/10.1016/j.pmrj.2011.08.535.
27. Westbury CB, Yarnold JR. Radiation fibrosis—current clinical and therapeutic perspectives. Clin Oncol (R Coll Radiol). 2012; 24: 657–72. https://doi.org/10.1016/j.clon.2012.04.001.
28. Borger JH, Kemperman H, Smitt HS, Hart A, van Dongen J, Lebesque J, Bartelink H. Dose and volume effects on fibrosis after breast conservation therapy. Int J Radiat Oncol Biol Phys. 1994; 30: 1073–81.
29. Davis AM, O’Sullivan B, Turcotte R, Bell R, Catton C, Chabot P, Wunder J, Hammond A, Benk V, Kandel R, Goddard K, Freeman C, Sadura A, et al. Late radiation morbidity following randomization to preoperative versus postoperative radiotherapy in extremity soft tissue sarcoma. Radiother Oncol. 2005; 75: 48–53.
30. Geara FB, Komaki R, Tucker SL, Travis EL, Cox JD. Factors influencing the development of lung fibrosis after chemoradiation for small cell carcinoma of the lung: evidence for inherent interindividual variation. Int J Radiat Oncol Biol Phys. 1998; 41: 279–86.
31. Graham MV, Purdy JA, Emami B, Harms W, Bosch W, Lockett MA, Perez CA. Clinical dose-volume histogram analysis for pneumonitis after 3D treatment for non-small cell lung cancer (NSCLC). Int J Radiat Oncol Biol Phys. 1999; 45: 323–9.
32. Johansson S, Svensson H, Denekamp J. Dose response and latency for radiation-induced fibrosis, edema, and neuropathy in breast cancer patients. Int J Radiat Oncol Biol Phys. 2002; 52: 1207–19.
33. Gold DG, Miller RC, Petersen IA, Osborn TG. Radiotherapy for malignancy in patients with scleroderma: the Mayo Clinic experience. Int J Radiat Oncol Biol Phys. 2007; 67: 559–67. https://doi.org/10.1016/j.ijrobp.2006.09.003.
34. Suarez EM, Knackstedt RJ, Jenrette JM. Significant fibrosis after radiation therapy in a patient with Marfan syndrome. Radiat Oncol J. 2014; 32: 208–12. https://doi.org/10.3857/roj.2014.32.3.208.
35. Andreassen CN, Overgaard J, Alsner J, Overgaard M, Herskind C, Cesaretti JA, Atencio DP, Green S, Formenti SC, Stock RG, Rosenstein BS. ATM sequence variants and risk of radiation-induced subcutaneous fibrosis after postmastectomy radiotherapy. Int J Radiat Oncol Biol Phys. 2006; 64: 776–83. https://doi.org/10.1016/j.ijrobp.2005.09.014.
36. Edvardsen H, Tefre T, Jansen L, Vu P, Haffty BG, Fosså SD, Kristensen VN, Børresen-Dale AL. Linkage disequilibrium pattern of the ATM gene in breast cancer patients and controls; association of SNPs and haplotypes to radio-sensitivity and post-lumpectomy local recurrence. Radiat Oncol. 2007; 2: 25. https://doi.org/10.1186/1748-717X-2-25.
37. Azria D, Gourgou S, Sozzi WJ, Zouhair A, Mirimanoff RO, Kramar A, Lemanski C, Dubois JB, Romieu G, Pelegrin A, Ozsahin M. Concomitant use of tamoxifen with radiotherapy enhances subcutaneous breast fibrosis in hypersensitive patients. Br J Cancer. 2004; 91: 1251–60. https://doi.org/10.1038/sj.bjc.6602146.
38. Cheuk IW, Yip SP, Kwong DL, Wu VW. Association of XRCC1 and XRCC3 gene haplotypes with the development of radiation-induced fibrosis in patients with nasopharyngeal carcinoma. Mol Clin Oncol. 2014; 2: 553–8. https://doi.org/10.3892/mco.2014.276.
39. Iwakawa M, Noda S, Ohta T, Oohira C, Tanaka H, Tsuji A, Ishikawa A, Imai T. Strain dependent differences in a histological study of CD44 and collagen fibers with an expression analysis of inflammatory response-related genes in irradiated murine lung. J Radiat Res. 2004; 45: 423–33.
40. Weigel C, Schmezer P, Plass C, Popanda O. Epigenetics in radiation-induced fibrosis. Oncogene. 2015; 34: 2145–55. https://doi.org/10.1038/onc.2014.145.
41. Edvardsen H, Landmark-Høyvik H, Reinertsen KV, Zhao X, Grenaker-Alnæs GI, Nebdal D, Syvänen AC, Rødningen O, Alsner J, Overgaard J, Borresen-Dale AL, Fosså SD, Kristensen VN. SNP in TXNRD2 associated with radiation-induced fibrosis: a study of genetic variation in reactive oxygen species metabolism and signaling. Int J Radiat Oncol Biol Phys. 2013; 86: 791–9. https://doi.org/10.1016/j.ijrobp.2013.02.025.
42. Delanian S, Depondt J, Lefaix JL. Major healing of refractory mandible osteoradionecrosis after treatment combining pentoxifylline and tocopherol: a phase II trial. Head Neck. 2005; 27: 114–23. https://doi.org/10.1002/hed.20121.
43. Delanian S, Lefaix JL. Complete healing of severe osteoradionecrosis with treatment combining pentoxifylline, tocopherol and clodronate. Br J Radiol. 2002; 75: 467–9. https://doi.org/10.1259/bjr.75.893.750467.
44. Gupta T, Agarwal J, Jain S, Phurailatpam R, Kannan S, Ghosh-Laskar S, Murthy V, Budrukkar A, Dinshaw K, Prabhash K, Chaturvedi P, D’Cruz A. Three-dimensional conformal radiotherapy (3D-CRT) versus intensity modulated radiation therapy (IMRT) in squamous cell carcinoma of the head and neck: a randomized controlled trial. Radiother Oncol. 2012; 104: 343–8. https://doi.org/10.1016/j.radonc.2012.07.001.
45. Jones HA, Hershock D, Machtay M, Chalian AA, Weber RS, Weinstein GS, Schumacher K, Kligerman MM, Berlin JA, Rosenthal DI. Preliminary investigation of symptom distress in the head and neck patient population: validation of a measurement instrument. Am J Clin Oncol. 2006; 29: 158–62. https://doi.org/10.1097/01.coc.0000207424.62275.9d.
46. Rosenthal DI, Lewin JS, Eisbruch A. Prevention and treatment of dysphagia and aspiration after chemoradiation for head and neck cancer. J Clin Oncol. 2006; 24: 2636–43. https://doi.org/10.1200/JCO.2006.06.0079.
47. Vainshtein JM, Griffith KA, Feng FY, Vineberg KA, Chepeha DB, Eisbruch A. Patient-reported voice and speech outcomes after whole-neck intensity modulated radiation therapy and chemotherapy for oropharyngeal cancer: prospective longitudinal study. Int J Radiat Oncol Biol Phys. 2014; 89: 973–80. https://doi.org/10.1016/j.ijrobp.2014.03.013.
48. Abratt RP, Morgan GW, Silvestri G, Willcox P. Pulmonary complications of radiation therapy. Clin Chest Med. 2004; 25: 167–77. https://doi.org/10.1016/S0272-5231(03)00126-6.
49. Delanian S, Martin M, Bravard A, Luccioni C, Lefaix JL. Abnormal phenotype of cultured fibroblasts in human skin with chronic radiotherapy damage. Radiother Oncol. 1998; 47: 255–61.
50. Coia LR, Myerson RJ, Tepper JE. Late effects of radiation therapy on the gastrointestinal tract. Int J Radiat Oncol Biol Phys. 1995; 31: 1213–36. https://doi.org/10.1016/0360-3016(94)00419-L.
51. Marks LB, Carroll PR, Dugan TC, Anscher MS. The response of the urinary bladder, urethra, and ureter to radiation and chemotherapy. Int J Radiat Oncol Biol Phys. 1995; 31: 1257–80. https://doi.org/10.1016/0360-3016(94)00431-J.
52. Pötter R, Knocke TH, Fellner C, Baldass M, Reinthaller A, Kucera H. Definitive radiotherapy based on HDR brachytherapy with iridium 192 in uterine cervix carcinoma: report on the Vienna University Hospital findings (1993-1997) compared to the preceding period in the context of ICRU 38 recommendations. Cancer Radiother. 2000; 4: 159–72. https://doi.org/10.1016/S1278-3218(00)88900-3.
53. Deng J, Ridner SH, Wells N, Dietrich MS, Murphy BA. Development and preliminary testing of head and neck cancer related external lymphedema and fibrosis assessment criteria. Eur J Oncol Nurs. 2015; 19: 75–80. https://doi.org/10.1016/j.ejon.2014.07.006.
54. Delanian S, Lefaix JL. Current management for late normal tissue injury: radiation-induced fibrosis and necrosis. Semin Radiat Oncol. 2007; 17: 99–107. https://doi.org/10.1016/j.semradonc.2006.11.006.
55. Travis EL. Organizational response of normal tissues to irradiation. Semin Radiat Oncol. 2001; 11: 184–96.
56. Cohen SR, Cohen EP. Chronic oxidative stress after irradiation: an unproven hypothesis. Med Hypotheses. 2013; 80: 172–5. https://doi.org/10.1016/j.mehy.2012.11.022.
57. Tak JK, Park JW. The use of ebselen for radioprotection in cultured cells and mice. Free Radic Biol Med. 2009; 46: 1177–85. https://doi.org/10.1016/j.freeradbiomed.2009.01.023.
58. Terasaki Y, Ohsawa I, Terasaki M, Takahashi M, Kunugi S, Dedong K, Urushiyama H, Amenomori S, Kaneko-Togashi M, Kuwahara N, Ishikawa A, Kamimura N, Ohta S, et al. Hydrogen therapy attenuates irradiation-induced lung damage by reducing oxidative stress. Am J Physiol Lung Cell Mol Physiol. 2011; 301: L415–26. https://doi.org/10.1152/ajplung.00008.2011.
59. Zhao W, Robbins ME. Inflammation and chronic oxidative stress in radiation-induced late normal tissue injury: therapeutic implications. Curr Med Chem. 2009; 16: 130–43.
60. Chittezhath M, Kuttan G. Radioprotective activity of naturally occurring organosulfur compounds. Tumori. 2006; 92: 163–9.
61. Greenberger JS, Epperly MW. Review. Antioxidant gene therapeutic approaches to normal tissue radioprotection and tumor radiosensitization. In Vivo. 2007; 21: 141–6.
62. Chaudière J, Ferrari-Iliou R. Intracellular antioxidants: from chemical to biochemical mechanisms. Food Chem Toxicol. 1999; 37: 949–62.
63. Darley-Usmar V, Halliwell B. Blood radicals: reactive nitrogen species, reactive oxygen species, transition metal ions, and the vascular system. Pharm Res. 1996; 13: 649–62.
64. Evans P, Halliwell B. Free radicals and hearing. Cause, consequence, and criteria. Ann N Y Acad Sci. 1999; 884: 19–40.
65. Denham JW, Hauer-Jensen M. The radiotherapeutic injury--a complex “wound.” Radiother Oncol. 2002; 63: 129–45.
66. Williams JP, Johnston CJ, Finkelstein JN. Treatment for radiation-induced pulmonary late effects: spoiled for choice or looking in the wrong direction? Curr Drug Targets. 2010; 11: 1386–94.
67. Boerma M, Hauer-Jensen M. Potential targets for intervention in radiation-induced heart disease. Curr Drug Targets. 2010; 11: 1405–12.
68. Lefaix JL, Daburon F. Diagnosis of acute localized irradiation lesions: review of the French experimental experience. Health Phys. 1998; 75: 375–84.
69. Abreu MT, Fukata M, Arditi M. TLR signaling in the gut in health and disease. J Immunol. 2005; 174: 4453–60.
70. Amadeu TP, Seabra AB, de Oliveira MG, Monte-Alto-Costa A. Nitric oxide donor improves healing if applied on inflammatory and proliferative phase. J Surg Res. 2008; 149: 84–93. https://doi.org/10.1016/j.jss.2007.10.015.
71. Calveley VL, Khan MA, Yeung IW, Vandyk J, Hill RP. Partial volume rat lung irradiation: temporal fluctuations of in-field and out-of-field DNA damage and inflammatory cytokines following irradiation. Int J Radiat Biol. 2005; 81: 887–99. https://doi.org/10.1080/09553000600568002.
72. Finkelstein JN, Johnston C, Barrett T, Oberdörster G. Particulate-cell interactions and pulmonary cytokine expression. Environ Health Perspect. 1997; 105 : 1179–82.
73. Olman MA, White KE, Ware LB, Cross MT, Zhu S, Matthay MA. Microarray analysis indicates that pulmonary edema fluid from patients with acute lung injury mediates inflammation, mitogen gene expression, and fibroblast proliferation through bioactive interleukin-1. Chest. 2002; 121: 69S–70S.
74. Porter DW, Ye J, Ma J, Barger M, Robinson VA, Ramsey D, McLaurin J, Khan A, Landsittel D, Teass A, Castranova V. Time course of pulmonary response of rats to inhalation of crystalline silica: NF-kappa B activation, inflammation, cytokine production, and damage. Inhal Toxicol. 2002; 14: 349–67. https://doi.org/10.1080/08958370252870998.
75. Johnston CJ, Piedboeuf B, Rubin P, Williams JP, Baggs R, Finkelstein JN. Early and persistent alterations in the expression of interleukin-1 alpha, interleukin-1 beta and tumor necrosis factor alpha mRNA levels in fibrosis-resistant and sensitive mice after thoracic irradiation. Radiat Res. 1996; 145: 762–7.
76. Kalash R, Berhane H, Goff J, Houghton F, Epperly MW, Dixon T, Zhang X, Sprachman MM, Wipf P, Franicola D, Wang H, Greenberger JS. Effects of thoracic irradiation on pulmonary endothelial compared to alveolar type-II cells in fibrosis-prone C57BL/6NTac mice. In Vivo. 2013; 27: 291–7.
77. Kalash R, Berhane H, Au J, Rhieu BH, Epperly MW, Goff J, Dixon T, Wang H, Zhang X, Franicola D, Shinde A, Greenberger JS. Differences in irradiated lung gene transcription between fibrosis-prone C57BL/6NHsd and fibrosis-resistant C3H/HeNHsd mice. In Vivo. 2014; 28: 147–71.
78. Rubin P, Johnston CJ, Williams JP, McDonald S, Finkelstein JN. A perpetual cascade of cytokines postirradiation leads to pulmonary fibrosis. Int J Radiat Oncol Biol Phys. 1995; 33: 99–109. https://doi.org/10.1016/0360-3016(95)00095-G.
79. Gordon S, Martinez FO. Alternative activation of macrophages: mechanism and functions. Immunity. 2010; 32: 593–604. https://doi.org/10.1016/j.immuni.2010.05.007.
80. Sica A, Mantovani A. Macrophage plasticity and polarization: in vivo veritas. J Clin Invest. 2012; 122: 787–95. https://doi.org/10.1172/JCI59643.
81. Varin A, Gordon S. Alternative activation of macrophages: immune function and cellular biology. Immunobiology. 2009; 214: 630–41. https://doi.org/10.1016/j.imbio.2008.11.009.
82. Li M, Jendrossek V, Belka C. The role of PDGF in radiation oncology. Radiat Oncol. 2007; 2: 5. https://doi.org/10.1186/1748-717X-2-5.
83. Li MO, Wan YY, Sanjabi S, Robertson AK, Flavell RA. Transforming growth factor-beta regulation of immune responses. Annu Rev Immunol. 2006; 24: 99–146. https://doi.org/10.1146/annurev.immunol.24.021605.090737.
84. Dadrich M, Nicolay NH, Flechsig P, Bickelhaupt S, Hoeltgen L, Roeder F, Hauser K, Tietz A, Jenne J, Lopez R, Roehrich M, Wirkner U, Lahn M, Huber PE. Combined inhibition of TGFβ and PDGF signaling attenuates radiation-induced pulmonary fibrosis. Oncoimmunology. 2016; 5: e1123366. https://doi.org/10.1080/2162402X.2015.1123366.
85. Rube CE, Uthe D, Schmid KW, Richter KD, Wessel J, Schuck A, Willich N, Rube C. Dose-dependent induction of transforming growth factor beta (TGF-beta) in the lung tissue of fibrosis-prone mice after thoracic irradiation. Int J Radiat Oncol Biol Phys. 2000; 47: 1033–42.
86. Thornton SC, Walsh BJ, Bennett S, Robbins JM, Foulcher E, Morgan GW, Penny R, Breit SN. Both in vitro and in vivo irradiation are associated with induction of macrophage-derived fibroblast growth factors. Clin Exp Immunol. 1996; 103: 67–73.
87. Yi ES, Bedoya A, Lee H, Chin E, Saunders W, Kim SJ, Danielpour D, Remick DG, Yin S, Ulich TR. Radiation-induced lung injury in vivo: expression of transforming growth factor-beta precedes fibrosis. Inflammation. 1996; 20: 339–52.
88. Campana F, Zervoudis S, Perdereau B, Gez E, Fourquet A, Badiu C, Tsakiris G, Koulaloglou S. Topical superoxide dismutase reduces post-irradiation breast cancer fibrosis. J Cell Mol Med. 2004; 8: 109–16.
89. Rodemann HP, Bamberg M. Cellular basis of radiation-induced fibrosis. Radiother Oncol. 1995; 35: 83–90.
90. Tomasek JJ, Gabbiani G, Hinz B, Chaponnier C, Brown RA. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat Rev Mol Cell Biol. 2002; 3: 349–63. https://doi.org/10.1038/nrm809.
91. Darby IA, Hewitson TD. Fibroblast differentiation in wound healing and fibrosis. Int Rev Cytol. 2007; 257: 143–79. https://doi.org/10.1016/S0074-7696(07)57004-X.
92. Chithra P, Sajithlal GB, Chandrakasan G. Influence of Aloe vera on the glycosaminoglycans in the matrix of healing dermal wounds in rats. J Ethnopharmacol. 1998; 59: 179–86.
93. Martin M, Lefaix J, Delanian S. TGF-beta1 and radiation fibrosis: a master switch and a specific therapeutic target? Int J Radiat Oncol Biol Phys. 2000; 47: 277–90.
94. Pardo A, Selman M. Matrix metalloproteases in aberrant fibrotic tissue remodeling. Proc Am Thorac Soc. 2006; 3: 383–8. https://doi.org/10.1513/pats.200601-012TK.
95. Nuutila K, Katayama S, Vuola J, Kankuri E. Human wound-healing research: issues and perspectives for studies using wide-scale analytic platforms. Adv Wound Care. 2014; 3: 264–71. https://doi.org/10.1089/wound.2013.0502.
96. Ran X, Cheng T, Shi C, Xu H, Qu J, Yan G, Su Y, Wang W, Xu R. The effects of total-body irradiation on the survival and skin wound healing of rats with combined radiation-wound injury. J Trauma. 2004; 57: 1087–93.
97. Finlay GA, Thannickal VJ, Fanburg BL, Paulson KE. Transforming growth factor-beta 1-induced activation of the ERK pathway/activator protein-1 in human lung fibroblasts requires the autocrine induction of basic fibroblast growth factor. J Biol Chem. 2000; 275: 27650–6. https://doi.org/10.1074/jbc.M000893200.
98. Burger A, Löffler H, Bamberg M, Rodemann HP. Molecular and cellular basis of radiation fibrosis. Int J Radiat Biol. 1998; 73: 401–8.
99. Rudolph R, Vande Berg J, Schneider JA, Fisher JC, Poolman WL. Slowed growth of cultured fibroblasts from human radiation wounds. Plast Reconstr Surg. 1988; 82: 669–77.
100. Toussaint O, Remacle J, Dierick JF, Pascal T, Frippiat C, Royer V, Chainiaux F. Approach of evolutionary theories of ageing, stress, senescence-like phenotypes, calorie restriction and hormesis from the view point of far-from-equilibrium thermodynamics. Mech Ageing Dev. 2002; 123: 937–46.
101. Bentzen SM, Overgaard M, Overgaard J. Clinical correlations between late normal tissue endpoints after radiotherapy: implications for predictive assays of radiosensitivity. Eur J Cancer. 1993; 29A: 1373–6.
102. Bentzen SM, Overgaard M. Relationship between early and late normal-tissue injury after postmastectomy radiotherapy. Radiother Oncol. 1991; 20: 159–65.
103. Bourhis J, Lapeyre M, Tortochaux J, Rives M, Aghili M, Bourdin S, Lesaunier F, Benassi T, Lemanski C, Geoffrois L, Lusinchi A, Verrelle P, Bardet E, et al. Phase III randomized trial of very accelerated radiation therapy compared with conventional radiation therapy in squamous cell head and neck cancer: a GORTEC trial. J Clin Oncol. 2006; 24: 2873–8. https://doi.org/10.1200/JCO.2006.08.057.
104. Barcellos-Hoff MH. How do tissues respond to damage at the cellular level? The role of cytokines in irradiated tissues. Radiat Res. 1998; 150: S109–20.
105. Beach TA, Johnston CJ, Groves AM, Williams JP, Finkelstein JN. Radiation induced pulmonary fibrosis as a model of progressive fibrosis: contributions of DNA damage, inflammatory response and cellular senescence genes. Exp Lung Res. 2017; 1–16. https://doi.org/10.1080/01902148.2017.1318975.
106. Ding NH, Li JJ, Sun LQ. Molecular mechanisms and treatment of radiation-induced lung fibrosis. Curr Drug Targets. 2013; 14: 1347–56.
107. Barcellos-Hoff MH, Derynck R, Tsang ML, Weatherbee JA. Transforming growth factor-beta activation in irradiated murine mammary gland. J Clin Invest. 1994; 93: 892–9. https://doi.org/10.1172/JCI117045.
108. Biswas S, Guix M, Rinehart C, Dugger TC, Chytil A, Moses HL, Freeman ML, Arteaga CL. Inhibition of TGF-beta with neutralizing antibodies prevents radiation-induced acceleration of metastatic cancer progression. J Clin Invest. 2007; 117: 1305–13. https://doi.org/10.1172/JCI30740.
109. Biswas S, Guix M, Rinehart C, Dugger TC, Chytil A, Moses HL, Freeman ML, Arteaga CL. Inhibition of TGF-β with neutralizing antibodies prevents radiation-induced acceleration of metastatic cancer progression. J Clin Invest. 2017; 127: 1116. https://doi.org/10.1172/JCI93333.
110. Travis EL, Rachakonda G, Zhou X, Korhonen K, Sekhar KR, Biswas S, Freeman ML. NRF2 deficiency reduces life span of mice administered thoracic irradiation. Free Radic Biol Med. 2011; 51: 1175–83. https://doi.org/10.1016/j.freeradbiomed.2011.05.038.
111. Bhowmick NA, Ghiassi M, Bakin A, Aakre M, Lundquist CA, Engel ME, Arteaga CL, Moses HL. Transforming growth factor-beta1 mediates epithelial to mesenchymal transdifferentiation through a RhoA-dependent mechanism. Mol Biol Cell. 2001; 12: 27–36.
112. Janda E, Lehmann K, Killisch I, Jechlinger M, Herzig M, Downward J, Beug H, Grünert S. Ras and TGF[beta] cooperatively regulate epithelial cell plasticity and metastasis: dissection of Ras signaling pathways. J Cell Biol. 2002; 156: 299–313. https://doi.org/10.1083/jcb.200109037.
113. Zavadil J, Böttinger EP. TGF-beta and epithelial-to-mesenchymal transitions. Oncogene. 2005; 24: 5764–74. https://doi.org/10.1038/sj.onc.1208927.
114. Nagarajan D, Melo T, Deng Z, Almeida C, Zhao W. ERK/GSK3β/Snail signaling mediates radiation-induced alveolar epithelial-to-mesenchymal transition. Free Radic Biol Med. 2012; 52: 983–92. https://doi.org/10.1016/j.freeradbiomed.2011.11.024.
115. Hinz B, Phan SH, Thannickal VJ, Galli A, Bochaton-Piallat ML, Gabbiani G. The myofibroblast: one function, multiple origins. Am J Pathol. 2007; 170: 1807–16. https://doi.org/10.2353/ajpath.2007.070112.
116. Hinz B, Phan SH, Thannickal VJ, Prunotto M, Desmoulière A, Varga J, De Wever O, Mareel M, Gabbiani G. Recent developments in myofibroblast biology: paradigms for connective tissue remodeling. Am J Pathol. 2012; 180: 1340–55. https://doi.org/10.1016/j.ajpath.2012.02.004.
117. Geraci JP, Mariano MS. Radiation hepatology of the rat: parenchymal and nonparenchymal cell injury. Radiat Res. 1993; 136: 205–13.
118. Arora H, Qureshi R, Park AK, Park WY. Coordinated regulation of ATF2 by miR-26b in γ-irradiated lung cancer cells. PLoS One. 2011; 6: e23802. https://doi.org/10.1371/journal.pone.0023802.
119. Seong J, Kim SH, Chung EJ, Lee WJ, Suh CO. Early alteration in TGF-beta mRNA expression in irradiated rat liver. Int J Radiat Oncol Biol Phys. 2000; 46: 639–43.
120. Chang XM, Chang Y, Jia A. Effects of interferon-alpha on expression of hepatic stellate cell and transforming growth factor-beta1 and alpha-smooth muscle actin in rats with hepatic fibrosis. World J Gastroenterol. 2005; 11: 2634–6.
121. Nouchi T, Tanaka Y, Tsukada T, Sato C, Marumo F. Appearance of alpha-smooth-muscle-actin-positive cells in hepatic fibrosis. Liver. 1991; 11: 100–5.
122. Imaeda M, Ishikawa H, Yoshida Y, Takahashi T, Ohkubo Y, Musha A, Komachi M, Nakazato Y, Nakano T. Long-term pathological and immunohistochemical features in the liver after intraoperative whole-liver irradiation in rats. J Radiat Res. 2014; 55: 665–73. https://doi.org/10.1093/jrr/rru005.
123. Gabbiani G, Chaponnier C, Hüttner I. Cytoplasmic filaments and gap junctions in epithelial cells and myofibroblasts during wound healing. J Cell Biol. 1978; 76: 561–8.
124. Serini G, Bochaton-Piallat ML, Ropraz P, Geinoz A, Borsi L, Zardi L, Gabbiani G. The fibronectin domain ED-A is crucial for myofibroblastic phenotype induction by transforming growth factor-beta1. J Cell Biol. 1998; 142: 873–81.
125. Desmoulière A, Redard M, Darby I, Gabbiani G. Apoptosis mediates the decrease in cellularity during the transition between granulation tissue and scar. Am J Pathol. 1995; 146: 56–66.
126. Van De Water L, Varney S, Tomasek JJ. Mechanoregulation of the myofibroblast in wound contraction, scarring, and fibrosis: opportunities for new therapeutic intervention. Adv Wound Care. 2013; 2: 122–41. https://doi.org/10.1089/wound.2012.0393.
127. Hao H, Gabbiani G, Camenzind E, Bacchetta M, Virmani R, Bochaton-Piallat ML. Phenotypic modulation of intima and media smooth muscle cells in fatal cases of coronary artery lesion. Arterioscler Thromb Vasc Biol. 2006; 26: 326–32. https://doi.org/10.1161/01.ATV.0000199393.74656.4c.
128. Friedman SL. Evolving challenges in hepatic fibrosis. Nat Rev Gastroenterol Hepatol. 2010; 7: 425–36. https://doi.org/10.1038/nrgastro.2010.97.
129. Duffield JS. Cellular and molecular mechanisms in kidney fibrosis. J Clin Invest. 2014; 124: 2299–306. https://doi.org/10.1172/JCI72267.
130. Galligan CL, Fish EN. The role of circulating fibrocytes in inflammation and autoimmunity. J Leukoc Biol. 2013; 93: 45–50. https://doi.org/10.1189/jlb.0712365.
131. Chiavegato A, Bochaton-Piallat ML, D’Amore E, Sartore S, Gabbiani G. Expression of myosin heavy chain isoforms in mammary epithelial cells and in myofibroblasts from different fibrotic settings during neoplasia. Virchows Arch Int J Pathol. 1995; 426: 77–86.
132. Kendall RT, Feghali-Bostwick CA. Fibroblasts in fibrosis: novel roles and mediators. Front Pharmacol. 2014; 5: 123. https://doi.org/10.3389/fphar.2014.00123.
133. Varga J, Pasche B. Transforming growth factor beta as a therapeutic target in systemic sclerosis. Nat Rev Rheumatol. 2009; 5: 200–6. https://doi.org/10.1038/nrrheum.2009.26.
134. Wei J, Bhattacharyya S, Tourtellotte WG, Varga J. Fibrosis in systemic sclerosis: emerging concepts and implications for targeted therapy. Autoimmun Rev. 2011; 10: 267–75. https://doi.org/10.1016/j.autrev.2010.09.015.
135. Kim KK, Kugler MC, Wolters PJ, Robillard L, Galvez MG, Brumwell AN, Sheppard D, Chapman HA. Alveolar epithelial cell mesenchymal transition develops in vivo during pulmonary fibrosis and is regulated by the extracellular matrix. Proc Natl Acad Sci U S A. 2006; 103: 13180–5. https://doi.org/10.1073/pnas.0605669103.
136. Goffin JM, Pittet P, Csucs G, Lussi JW, Meister JJ, Hinz B. Focal adhesion size controls tension-dependent recruitment of alpha-smooth muscle actin to stress fibers. J Cell Biol. 2006; 172: 259–68. https://doi.org/10.1083/jcb.200506179.
137. Desmoulière A, Geinoz A, Gabbiani F, Gabbiani G. Transforming growth factor-beta 1 induces alpha-smooth muscle actin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. J Cell Biol. 1993; 122: 103–11.
138. Hinz B, Celetta G, Tomasek JJ, Gabbiani G, Chaponnier C. Alpha-smooth muscle actin expression upregulates fibroblast contractile activity. Mol Biol Cell. 2001; 12: 2730–41.
139. Dugina V, Fontao L, Chaponnier C, Vasiliev J, Gabbiani G. Focal adhesion features during myofibroblastic differentiation are controlled by intracellular and extracellular factors. J Cell Sci. 2001; 114: 3285–96.
140. Conti MA, Adelstein RS. Nonmuscle myosin II moves in new directions. J Cell Sci. 2008; 121: 11–8. https://doi.org/10.1242/jcs.007112.
141. Lecarpentier E, Claes V, Timbely O, Hébert JL, Arsalane A, Moumen A, Guerin C, Guizard M, Michel F, Lecarpentier Y. Role of both actin-myosin cross bridges and NO-cGMP pathway modulators in the contraction and relaxation of human placental stem villi. Placenta. 2013; 34: 1163–9. https://doi.org/10.1016/j.placenta.2013.10.007.
142. Matsumura S, Sakurai K, Shinomiya T, Fujitani N, Key K, Ohashi M. Biochemical and immunohistochemical characterization of the isoforms of myosin and actin in human placenta. Placenta. 2011; 32: 347–55. https://doi.org/10.1016/j.placenta.2011.02.008.
143. Parizi M, Howard EW, Tomasek JJ. Regulation of LPA-promoted myofibroblast contraction: role of Rho, myosin light chain kinase, and myosin light chain phosphatase. Exp Cell Res. 2000; 254: 210–20. https://doi.org/10.1006/excr.1999.4754.
144. Tomasek JJ, Vaughan MB, Kropp BP, Gabbiani G, Martin MD, Haaksma CJ, Hinz B. Contraction of myofibroblasts in granulation tissue is dependent on Rho/Rho kinase/myosin light chain phosphatase activity. Wound Repair Regen. 2006; 14: 313–20. https://doi.org/10.1111/j.1743-6109.2006.00126.x.
145. Kovács M, Wang F, Hu A, Zhang Y, Sellers JR. Functional divergence of human cytoplasmic myosin II: kinetic characterization of the non-muscle IIA isoform. J Biol Chem. 2003; 278: 38132–40. https://doi.org/10.1074/jbc.M305453200.
146. Lecarpentier Y, Claes V, Lecarpentier E, Guerin C, Hébert JL, Arsalane A, Moumen A, Krokidis X, Michel F, Timbely O. Ultraslow myosin molecular motors of placental contractile stem villi in humans. PLoS One. 2014; 9: e108814. https://doi.org/10.1371/journal.pone.0108814.
147. Lecarpentier Y, Claes V, Hébert JL, Krokidis X, Blanc FX, Michel F, Timbely O. Statistical mechanics of the human placenta: a stationary state of a near-equilibrium system in a linear regime. PLoS One. 2015; 10: e0142471. https://doi.org/10.1371/journal.pone.0142471.
148. Farley AE, Graham CH, Smith GN. Contractile properties of human placental anchoring villi. Am J Physiol Regul Integr Comp Physiol. 2004; 287: R680-685. https://doi.org/10.1152/ajpregu.00222.2004.
149. Huszar G, Bailey P. Isolation and characterization of myosin in the human term placenta. Am J Obstet Gynecol. 1979; 135: 707–12.
150. Michael C. Actomyosin content of the human placenta. J Obstet Gynaecol Br Commonw. 1974; 81: 307–10.
151. van Amerongen R, Nusse R. Towards an integrated view of Wnt signaling in development. Development. 2009; 136: 3205–14. https://doi.org/10.1242/dev.033910.
152. Al-Harthi L. Wnt/β-catenin and its diverse physiological cell signaling pathways in neurodegenerative and neuropsychiatric disorders. J Neuroimmune Pharmacol. 2012; 7: 725–30. https://doi.org/10.1007/s11481-012-9412-x.
153. Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol. 2004; 20: 781–810. https://doi.org/10.1146/annurev.cellbio.20.010403.113126.
154. Angers S, Moon RT. Proximal events in Wnt signal transduction. Nat Rev Mol Cell Biol. 2009; 10: 468–77. https://doi.org/10.1038/nrm2717.
155. Clevers H, Nusse R. Wnt/β-catenin signaling and disease. Cell. 2012; 149: 1192–205. https://doi.org/10.1016/j.cell.2012.05.012.
156. Aberle H, Bauer A, Stappert J, Kispert A, Kemler R. β-catenin is a target for the ubiquitin–proteasome pathway. EMBO J. 1997; 16: 3797–804. https://doi.org/10.1093/emboj/16.13.3797.
157. Gurung A, Uddin F, Hill RP, Ferguson PC, Alman BA. Beta-catenin is a mediator of the response of fibroblasts to irradiation. Am J Pathol. 2009; 174: 248–55. https://doi.org/10.2353/ajpath.2009.080576.
158. Chang HW, Roh JL, Jeong EJ, Lee S, Kim SW, Choi SH, Park SK, Kim SY. Wnt signaling controls radiosensitivity via cyclooxygenase-2-mediated Ku expression in head and neck cancer. Int J Cancer. 2008; 122: 100–7. https://doi.org/10.1002/ijc.23069.
159. Chen MS, Woodward WA, Behbod F, Peddibhotla S, Alfaro MP, Buchholz TA, Rosen JM. Wnt/beta-catenin mediates radiation resistance of Sca1+ progenitors in an immortalized mammary gland cell line. J Cell Sci. 2007; 120: 468–77. https://doi.org/10.1242/jcs.03348.
160. Kwon O, Kim KA, He L, Kim SO, Kim MS, Cha EY, Yoon BD, Sok DE, Jung M, Ahn JS, Kim BY. Ionizing radiation can induce GSK-3beta phosphorylation and NF-kappaB transcriptional transactivation in ATM-deficient fibroblasts. Cell Signal. 2008; 20: 602–12. https://doi.org/10.1016/j.cellsig.2007.10.022.
161. Rødningen OK, Overgaard J, Alsner J, Hastie T, Børresen-Dale AL. Microarray analysis of the transcriptional response to single or multiple doses of ionizing radiation in human subcutaneous fibroblasts. Radiother Oncol. 2005; 77: 231–40. https://doi.org/10.1016/j.radonc.2005.09.020.
162. Aumiller V, Balsara N, Wilhelm J, Günther A, Königshoff M. WNT/β-catenin signaling induces IL-1β expression by alveolar epithelial cells in pulmonary fibrosis. Am J Respir Cell Mol Biol. 2013; 49: 96–104. https://doi.org/10.1165/rcmb.2012-0524OC.
163. Baarsma HA, Spanjer AI, Haitsma G, Engelbertink LH, Meurs H, Jonker MR, Timens W, Postma DS, Kerstjens HA, Gosens R. Activation of WNT/β-catenin signaling in pulmonary fibroblasts by TGF-β1 is increased in chronic obstructive pulmonary disease. PLoS One. 2011; 6: e25450. https://doi.org/10.1371/journal.pone.0025450.
164. Dees C, Distler JH. Canonical Wnt signalling as a key regulator of fibrogenesis - implications for targeted therapies? Exp Dermatol. 2013; 22: 710–3. https://doi.org/10.1111/exd.12255.
165. Königshoff M, Kneidinger N, Eickelberg O. TGF-beta signaling in COPD: deciphering genetic and cellular susceptibilities for future therapeutic regimen. Swiss Med Wkly. 2009; 139: 554–63. https://doi.org/smw-12528.
166. Königshoff M, Eickelberg O. WNT signaling in lung disease: a failure or a regeneration signal? Am J Respir Cell Mol Biol. 2010; 42: 21–31. https://doi.org/10.1165/rcmb.2008-0485TR.
167. Wei J, Fang F, Lam AP, Sargent JL, Hamburg E, Hinchcliff ME, Gottardi CJ, Atit R, Whitfield ML, Varga J. Wnt/β-catenin signaling is hyperactivated in systemic sclerosis and induces Smad-dependent fibrotic responses in mesenchymal cells. Arthritis Rheum. 2012; 64: 2734–45. https://doi.org/10.1002/art.34424.
168. Brazil DP, Yang ZZ, Hemmings BA. Advances in protein kinase B signalling: AKTion on multiple fronts. Trends Biochem Sci. 2004; 29: 233–42. https://doi.org/10.1016/j.tibs.2004.03.006.
169. Ciuffreda L, Di Sanza C, Incani UC, Milella M. The mTOR pathway: a new target in cancer therapy. Curr Cancer Drug Targets. 2010; 10: 484–95.
170. Heras-Sandoval D, Pérez-Rojas JM, Hernández-Damián J, Pedraza-Chaverri J. The role of PI3K/AKT/mTOR pathway in the modulation of autophagy and the clearance of protein aggregates in neurodegeneration. Cell Signal. 2014; 26: 2694–701. https://doi.org/10.1016/j.cellsig.2014.08.019.
171. Yu JS, Cui W. Proliferation, survival and metabolism: the role of PI3K/AKT/mTOR signalling in pluripotency and cell fate determination. Development. 2016; 143: 3050–60. https://doi.org/10.1242/dev.137075.
172. Chen J, Alberts I, Li X. Dysregulation of the IGF-I/PI3K/AKT/mTOR signaling pathway in autism spectrum disorders. Int J Dev Neurosci. 2014; 35: 35–41. https://doi.org/10.1016/j.ijdevneu.2014.03.006.
173. Huang J, Nguyen-McCarty M, Hexner EO, Danet-Desnoyers G, Klein PS. Maintenance of hematopoietic stem cells through regulation of Wnt and mTOR pathways. Nat Med. 2012; 18: 1778–85. https://doi.org/10.1038/nm.2984.
174. Park KS, Lee RD, Kang SK, Han SY, Park KL, Yang KH, Song YS, Park HJ, Lee YM, Yun YP, Oh KW, Kim DJ, Yun YW, et al. Neuronal differentiation of embryonic midbrain cells by upregulation of peroxisome proliferator-activated receptor-gamma via the JNK-dependent pathway. Exp Cell Res. 2004; 297: 424–33. https://doi.org/10.1016/j.yexcr.2004.03.034.
175. Yue X, Lan F, Yang W, Yang Y, Han L, Zhang A, Liu J, Zeng H, Jiang T, Pu P, Kang C. Interruption of β-catenin suppresses the EGFR pathway by blocking multiple oncogenic targets in human glioma cells. Brain Res. 2010; 1366: 27–37. https://doi.org/10.1016/j.brainres.2010.10.032.
176. Ross SE, Erickson RL, Hemati N, MacDougald OA. Glycogen synthase kinase 3 is an insulin-regulated C/EBPalpha kinase. Mol Cell Biol. 1999; 19: 8433–41.
177. Tang QQ, Grønborg M, Huang H, Kim JW, Otto TC, Pandey A, Lane MD. Sequential phosphorylation of CCAAT enhancer-binding protein beta by MAPK and glycogen synthase kinase 3beta is required for adipogenesis. Proc Natl Acad Sci U S A. 2005; 102: 9766–71. https://doi.org/10.1073/pnas.0503891102.
178. Karin M. NF-kappaB as a critical link between inflammation and cancer. Cold Spring Harb Perspect Biol. 2009; 1: a000141. https://doi.org/10.1101/cshperspect.a000141.
179. Pasparakis M. Regulation of tissue homeostasis by NF-kappaB signalling: implications for inflammatory diseases. Nat Rev Immunol. 2009; 9: 778–88. https://doi.org/10.1038/nri2655.
180. Ben-Neriah Y, Karin M. Inflammation meets cancer, with NF-κB as the matchmaker. Nat Immunol. 2011; 12: 715–23. https://doi.org/10.1038/ni.2060.
181. Tak PP, Firestein GS. NF-kappaB: a key role in inflammatory diseases. J Clin Invest. 2001; 107: 7–11. https://doi.org/10.1172/JCI11830.
182. Ma B, Hottiger MO. Crosstalk between Wnt/β-catenin and NF-κB signaling pathway during inflammation. Front Immunol. 2016; 7: 378. https://doi.org/10.3389/fimmu.2016.00378.
183. Oguma K, Oshima H, Aoki M, Uchio R, Naka K, Nakamura S, Hirao A, Saya H, Taketo MM, Oshima M. Activated macrophages promote Wnt signalling through tumour necrosis factor-alpha in gastric tumour cells. EMBO J. 2008; 27: 1671–81. https://doi.org/10.1038/emboj.2008.105.
184. Zhang Y, Tomann P, Andl T, Gallant NM, Huelsken J, Jerchow B, Birchmeier W, Paus R, Piccolo S, Mikkola ML, Morrisey EE, Overbeek PA, Scheidereit C, et al. Reciprocal requirements for EDA/EDAR/NF-kappaB and Wnt/beta-catenin signaling pathways in hair follicle induction. Dev Cell. 2009; 17: 49–61. https://doi.org/10.1016/j.devcel.2009.05.011.
185. Nejak-Bowen K, Kikuchi A, Monga SPS. Beta-catenin-NF-κB interactions in murine hepatocytes: a complex to die for. Hepatology. 2013; 57: 763–74. https://doi.org/10.1002/hep.26042.
186. Spiegelman VS, Slaga TJ, Pagano M, Minamoto T, Ronai Z, Fuchs SY. Wnt/beta-catenin signaling induces the expression and activity of betaTrCP ubiquitin ligase receptor. Mol Cell. 2000; 5: 877–82.
187. Noubissi FK, Elcheva I, Bhatia N, Shakoori A, Ougolkov A, Liu J, Minamoto T, Ross J, Fuchs SY, Spiegelman VS. CRD-BP mediates stabilization of betaTrCP1 and c-myc mRNA in response to beta-catenin signalling. Nature. 2006; 441: 898–901. https://doi.org/10.1038/nature04839.
188. Kuphal S, Poser I, Jobin C, Hellerbrand C, Bosserhoff AK. Loss of E-cadherin leads to upregulation of NFkappaB activity in malignant melanoma. Oncogene. 2004; 23: 8509–19. https://doi.org/10.1038/sj.onc.1207831.
189. Anson M, Crain-Denoyelle AM, Baud V, Chereau F, Gougelet A, Terris B, Yamagoe S, Colnot S, Viguier M, Perret C, Couty JP. Oncogenic β-catenin triggers an inflammatory response that determines the aggressiveness of hepatocellular carcinoma in mice. J Clin Invest. 2012; 122: 586–99. https://doi.org/10.1172/JCI43937.
190. Wang X, Adhikari N, Li Q, Guan Z, Hall JL. The role of [beta]-transducin repeat-containing protein ([beta]-TrCP) in the regulation of NF-[kappa]B in vascular smooth muscle cells. Arterioscler Thromb Vasc Biol. 2004; 24: 85–90. https://doi.org/10.1161/01.ATV.0000104012.40720.c4.
191. Schön S, Flierman I, Ofner A, Stahringer A, Holdt LM, Kolligs FT, Herbst A. β-catenin regulates NF-κB activity via TNFRSF19 in colorectal cancer cells. Int J Cancer. 2014; 135: 1800–11. https://doi.org/10.1002/ijc.28839.
192. Zhao W, Sun Z, Wang S, Li Z, Zheng L. Wnt1 participates in inflammation induced by lipopolysaccharide through upregulating scavenger receptor A and NF-kB. Inflammation. 2015; 38: 1700–6. https://doi.org/10.1007/s10753-015-0147-8.
193. Choi YS, Hur J, Jeong S. Beta-catenin binds to the downstream region and regulates the expression C-reactive protein gene. Nucleic Acids Res. 2007; 35: 5511–9. https://doi.org/10.1093/nar/gkm547.
194. Yun K, So JS, Jash A, Im SH. Lymphoid enhancer binding factor 1 regulates transcription through gene looping. J Immunol. 2009; 183: 5129–37. https://doi.org/10.4049/jimmunol.0802744.
195. Ma B, Zhong L, van Blitterswijk CA, Post JN, Karperien M. T cell factor 4 is a pro-catabolic and apoptotic factor in human articular chondrocytes by potentiating nuclear factor κB signaling. J Biol Chem. 2013; 288: 17552–8. https://doi.org/10.1074/jbc.M113.453985.
196. Jia D, Yang W, Li L, Liu H, Tan Y, Ooi S, Chi L, Filion LG, Figeys D, Wang L. β-Catenin and NF-κB co-activation triggered by TLR3 stimulation facilitates stem cell-like phenotypes in breast cancer. Cell Death Differ. 2015; 22: 298–310. https://doi.org/10.1038/cdd.2014.145.
197. Cummins EP, Berra E, Comerford KM, Ginouves A, Fitzgerald KT, Seeballuck F, Godson C, Nielsen JE, Moynagh P, Pouyssegur J, Taylor CT. Prolyl hydroxylase-1 negatively regulates IkappaB kinase-beta, giving insight into hypoxia-induced NFkappaB activity. Proc Natl Acad Sci U S A. 2006; 103: 18154–9. https://doi.org/10.1073/pnas.0602235103.
198. Scholz CC, Cavadas MA, Tambuwala MM, Hams E, Rodríguez J, von Kriegsheim A, Cotter P, Bruning U, Fallon PG, Cheong A, Cummins EP, Taylor CT. Regulation of IL-1β-induced NF-κB by hydroxylases links key hypoxic and inflammatory signaling pathways. Proc Natl Acad Sci U S A. 2013; 110: 18490–5. https://doi.org/10.1073/pnas.1309718110.
199. Buss H, Dörrie A, Schmitz ML, Frank R, Livingstone M, Resch K, Kracht M. Phosphorylation of serine 468 by GSK-3beta negatively regulates basal p65 NF-kappaB activity. J Biol Chem. 2004; 279: 49571–4. https://doi.org/10.1074/jbc.C400442200.
200. Saegusa M, Hashimura M, Kuwata T, Hamano M, Okayasu I. Crosstalk between NF-kappaB/p65 and beta-catenin/TCF4/p300 signalling pathways through alterations in GSK-3beta expression during trans-differentiation of endometrial carcinoma cells. J Pathol. 2007; 213: 35–45. https://doi.org/10.1002/path.2198.
201. Lamberti C, Lin KM, Yamamoto Y, Verma U, Verma IM, Byers S, Gaynor RB. Regulation of beta-catenin function by the IkappaB kinases. J Biol Chem. 2001; 276: 42276–86. https://doi.org/10.1074/jbc.M104227200.
202. Yun K, Choi YD, Nam JH, Park Z, Im SH. NF-kappaB regulates Lef1 gene expression in chondrocytes. Biochem Biophys Res Commun. 2007; 357: 589–95. https://doi.org/10.1016/j.bbrc.2007.03.170.
203. Cho HH, Joo HJ, Song JS, Bae YC, Jung JS. Crossregulation of beta-catenin/Tcf pathway by NF-kappaB is mediated by lzts2 in human adipose tissue-derived mesenchymal stem cells. Biochim Biophys Acta. 2008; 1783: 419–28. https://doi.org/10.1016/j.bbamcr.2007.08.005.
204. Cho HH, Song JS, Yu JM, Yu SS, Choi SJ, Kim DH, Jung JS. Differential effect of NF-kappaB activity on beta-catenin/Tcf pathway in various cancer cells. FEBS Lett. 2008; 582: 616–22. https://doi.org/10.1016/j.febslet.2008.01.029.
205. Albanese C, Wu K, D’Amico M, Jarrett C, Joyce D, Hughes J, Hulit J, Sakamaki T, Fu M, Ben-Ze’ev A, Bromberg JF, Lamberti C, Verma U, et al. IKKalpha regulates mitogenic signaling through transcriptional induction of cyclin D1 via Tcf. Mol Biol Cell. 2003; 14: 585–99. https://doi.org/10.1091/mbc.02-06-0101.
206. Li D, Beisswenger C, Herr C, Hellberg J, Han G, Zakharkina T, Voss M, Wiewrodt R, Bohle RM, Menger MD, Schmid RM, Stöckel D, Lenhof HP, et al. Myeloid cell RelA/p65 promotes lung cancer proliferation through Wnt/β-catenin signaling in murine and human tumor cells. Oncogene. 2014; 33: 1239–48. https://doi.org/10.1038/onc.2013.75.
207. Ahmed KM, Li JJ. NF-kappa B-mediated adaptive resistance to ionizing radiation. Free Radic Biol Med. 2008; 44: 1–13. https://doi.org/10.1016/j.freeradbiomed.2007.09.022.
208. Chou CH, Chen SU, Cheng JC. Radiation-induced interleukin-6 expression through MAPK/p38/NF-kappaB signaling pathway and the resultant antiapoptotic effect on endothelial cells through Mcl-1 expression with sIL6-Ralpha. Int J Radiat Oncol Biol Phys. 2009; 75: 1553–61. https://doi.org/10.1016/j.ijrobp.2009.08.034.
209. Yang H, Zhao LF, Zhao ZF, Wang Y, Zhao JJ, Zhang L. Heme oxygenase-1 prevents liver fibrosis in rats by regulating the expression of PPARγ and NF-κB. World J Gastroenterol. 2012; 18: 1680–8. https://doi.org/10.3748/wjg.v18.i14.1680.
210. Sinha S, Ghildiyal R, Mehta VS, Sen E. ATM-NFκB axis-driven TIGAR regulates sensitivity of glioma cells to radiomimetics in the presence of TNFα. Cell Death Dis. 2013; 4: e615. https://doi.org/10.1038/cddis.2013.128.
211. Colston JT, de la Rosa SD, Koehler M, Gonzales K, Mestril R, Freeman GL, Bailey SR, Chandrasekar B. Wnt-induced secreted protein-1 is a prohypertrophic and profibrotic growth factor. Am J Physiol Heart Circ Physiol. 2007; 293: H1839–46. https://doi.org/10.1152/ajpheart.00428.2007.
212. Königshoff M, Kramer M, Balsara N, Wilhelm J, Amarie OV, Jahn A, Rose F, Fink L, Seeger W, Schaefer L, Günther A, Eickelberg O. WNT1-inducible signaling protein-1 mediates pulmonary fibrosis in mice and is upregulated in humans with idiopathic pulmonary fibrosis. J Clin Invest. 2009; 119: 772–87. https://doi.org/10.1172/JCI33950.
213. Desnoyers L, Arnott D, Pennica D. WISP-1 binds to decorin and biglycan. J Biol Chem. 2001; 276: 47599–607. https://doi.org/10.1074/jbc.M108339200.
214. Li X, Chen Y, Ye W, Tao X, Zhu J, Wu S, Lou L. Blockade of CCN4 attenuates CCl4-induced liver fibrosis. Arch Med Sci. 2015; 11: 647–53. https://doi.org/10.5114/aoms.2015.52371.
215. Blobe GC, Schiemann WP, Lodish HF. Role of transforming growth factor beta in human disease. N Engl J Med. 2000; 342: 1350–8. https://doi.org/10.1056/NEJM200005043421807.
216. Carthy JM, Garmaroudi FS, Luo Z, McManus BM. Wnt3a induces myofibroblast differentiation by upregulating TGF-β signaling through SMAD2 in a β-catenin-dependent manner. PLoS One. 2011; 6: e19809. https://doi.org/10.1371/journal.pone.0019809.
217. Bhattacharyya S, Wei J, Tourtellotte WG, Hinchcliff M, Gottardi CG, Varga J. Fibrosis in systemic sclerosis: common and unique pathobiology. Fibrogenesis Tissue Repair. 2012; 5: S18. https://doi.org/10.1186/1755-1536-5-S1-S18.
218. Georgescu MM. PTEN tumor suppressor network in PI3K-Akt pathway control. Genes Cancer. 2010; 1: 1170–7. https://doi.org/10.1177/1947601911407325.
219. White ES, Atrasz RG, Hu B, Phan SH, Stambolic V, Mak TW, Hogaboam CM, Flaherty KR, Martinez FJ, Kontos CD, Toews GB. Negative regulation of myofibroblast differentiation by PTEN (Phosphatase and Tensin Homolog Deleted on chromosome 10). Am J Respir Crit Care Med. 2006; 173: 112–21. https://doi.org/10.1164/rccm.200507-1058OC.
220. Lam AP, Herazo-Maya JD, Sennello JA, Flozak AS, Russell S, Mutlu GM, Budinger GR, DasGupta R, Varga J, Kaminski N, Gottardi CJ. Wnt coreceptor Lrp5 is a driver of idiopathic pulmonary fibrosis. Am J Respir Crit Care Med. 2014; 190: 185–95. https://doi.org/10.1164/rccm.201401-0079OC.
221. Alarcón C, Zaromytidou AI, Xi Q, Gao S, Yu J, Fujisawa S, Barlas A, Miller AN, Manova-Todorova K, Macias MJ, Sapkota G, Pan D, Massagué J. Nuclear CDKs drive Smad transcriptional activation and turnover in BMP and TGF-beta pathways. Cell. 2009; 139: 757–69. https://doi.org/10.1016/j.cell.2009.09.035.
222. Akhmetshina A, Palumbo K, Dees C, Bergmann C, Venalis P, Zerr P, Horn A, Kireva T, Beyer C, Zwerina J, Schneider H, Sadowski A, Riener MO, et al. Activation of canonical Wnt signalling is required for TGF-β-mediated fibrosis. Nat Commun. 2012; 3: 735. https://doi.org/10.1038/ncomms1734.
223. Furuhashi M, Yagi K, Yamamoto H, Furukawa Y, Shimada S, Nakamura Y, Kikuchi A, Miyazono K, Kato M. Axin facilitates Smad3 activation in the transforming growth factor β signaling pathway. Mol Cell Biol. 2001; 21: 5132–41. https://doi.org/10.1128/MCB.21.15.5132-5141.2001.
224. Liu W, Rui H, Wang J, Lin S, He Y, Chen M, Li Q, Ye Z, Zhang S, Chan SC, Chen YG, Han J, Lin SC. Axin is a scaffold protein in TGF-β signaling that promotes degradation of Smad7 by Arkadia. EMBO J. 2006; 25: 1646–58. https://doi.org/10.1038/sj.emboj.7601057.
225. Hayashi H, Abdollah S, Qiu Y, Cai J, Xu YY, Grinnell BW, Richardson MA, Topper JN, Gimbrone MA, Wrana JL, Falb D. The MAD-related protein Smad7 associates with the TGFbeta receptor and functions as an antagonist of TGFbeta signaling. Cell. 1997; 89: 1165–73.
226. Ferrigno O, Lallemand F, Verrecchia F, L’Hoste S, Camonis J, Atfi A, Mauviel A. Yes-associated protein (YAP65) interacts with Smad7 and potentiates its inhibitory activity against TGF-beta/Smad signaling. Oncogene. 2002; 21: 4879–84. https://doi.org/10.1038/sj.onc.1205623.
227. Han G, Li AG, Liang YY, Owens P, He W, Lu S, Yoshimatsu Y, Wang D, Ten Dijke P, Lin X, Wang XJ. Smad7-induced beta-catenin degradation alters epidermal appendage development. Dev Cell. 2006; 11: 301–12. https://doi.org/10.1016/j.devcel.2006.06.014.
228. Liu F, Lagares D, Choi KM, Stopfer L, Marinković A, Vrbanac V, Probst CK, Hiemer SE, Sisson TH, Horowitz JC, Rosas IO, Fredenburgh LE, Feghali-Bostwick C, et al. Mechanosignaling through YAP and TAZ drives fibroblast activation and fibrosis. Am J Physiol Lung Cell Mol Physiol. 2015; 308: L344–57. https://doi.org/10.1152/ajplung.00300.2014.
229. Mannaerts I, Leite SB, Verhulst S, Claerhout S, Eysackers N, Thoen LF, Hoorens A, Reynaert H, Halder G, van Grunsven LA. The Hippo pathway effector YAP controls mouse hepatic stellate cell activation. J Hepatol. 2015; 63: 679–88. https://doi.org/10.1016/j.jhep.2015.04.011.
230. Piersma B, Bank RA, Boersema M. Signaling in fibrosis: TGF-β, WNT, and YAP/TAZ converge. Front Med (Lausanne). 2015; 2: 59. https://doi.org/10.3389/fmed.2015.00059.
231. Varelas X, Samavarchi-Tehrani P, Narimatsu M, Weiss A, Cockburn K, Larsen BG, Rossant J, Wrana JL. The Crumbs complex couples cell density sensing to Hippo-dependent control of the TGF-β-SMAD pathway. Dev Cell. 2010; 19: 831–44. https://doi.org/10.1016/j.devcel.2010.11.012.
232. Imajo M, Miyatake K, Iimura A, Miyamoto A, Nishida E. A molecular mechanism that links Hippo signalling to the inhibition of Wnt/β-catenin signalling. EMBO J. 2012; 31: 1109–22. https://doi.org/10.1038/emboj.2011.487.
233. Varelas X, Sakuma R, Samavarchi-Tehrani P, Peerani R, Rao BM, Dembowy J, Yaffe MB, Zandstra PW, Wrana JL. TAZ controls Smad nucleocytoplasmic shuttling and regulates human embryonic stem-cell self-renewal. Nat Cell Biol. 2008; 10: 837–48. https://doi.org/10.1038/ncb1748.
234. Ahmadian M, Suh JM, Hah N, Liddle C, Atkins AR, Downes M, Evans RM. PPARγ signaling and metabolism: the good, the bad and the future. Nat Med. 2013; 19: 557–66. https://doi.org/10.1038/nm.3159.
235. Elbrecht A, Chen Y, Cullinan CA, Hayes N, Leibowitz MD, Moller DE, Berger J. Molecular cloning, expression and characterization of human peroxisome proliferator activated receptors gamma 1 and gamma 2. Biochem Biophys Res Commun. 1996; 224: 431–7.
236. Fajas L, Auboeuf D, Raspé E, Schoonjans K, Lefebvre AM, Saladin R, Najib J, Laville M, Fruchart JC, Deeb S, Vidal-Puig A, Flier J, Briggs MR, et al. The organization, promoter analysis, and expression of the human PPARgamma gene. J Biol Chem. 1997; 272: 18779–89.
237. Ricote M, Glass CK. PPARs and molecular mechanisms of transrepression. Biochim Biophys Acta. 2007; 1771: 926–35. https://doi.org/10.1016/j.bbalip.2007.02.013.
238. Lakatos HF, Thatcher TH, Kottmann RM, Garcia TM, Phipps RP, Sime PJ. The role of PPARs in lung fibrosis. PPAR Res. 2007; 2007: 71323. https://doi.org/10.1155/2007/71323.
239. Park EJ, Park SY, Joe E, Jou I. 15d-PGJ2 and rosiglitazone suppress Janus kinase-STAT inflammatory signaling through induction of suppressor of cytokine signaling 1 (SOCS1) and SOCS3 in glia. J Biol Chem. 2003; 278: 14747–52. https://doi.org/10.1074/jbc.M210819200.
240. Simon MF, Daviaud D, Pradère JP, Grès S, Guigné C, Wabitsch M, Chun J, Valet P, Saulnier-Blache JS. Lysophosphatidic acid inhibits adipocyte differentiation via lysophosphatidic acid 1 receptor-dependent down-regulation of peroxisome proliferator-activated receptor gamma2. J Biol Chem. 2005; 280: 14656–62. https://doi.org/10.1074/jbc.M412585200.
241. Tan JT, McLennan SV, Song WW, Lo LW, Bonner JG, Williams PF, Twigg SM. Connective tissue growth factor inhibits adipocyte differentiation. Am J Physiol Cell Physiol. 2008; 295: C740–51. https://doi.org/10.1152/ajpcell.00333.2007.
242. Yamasaki S, Nakashima T, Kawakami A, Miyashita T, Tanaka F, Ida H, Migita K, Origuchi T, Eguchi K. Cytokines regulate fibroblast-like synovial cell differentiation to adipocyte-like cells. Rheumatology (Oxford). 2004; 43: 448–52. https://doi.org/10.1093/rheumatology/keh092.
243. Okamura M, Kudo H, Wakabayashi K, Tanaka T, Nonaka A, Uchida A, Tsutsumi S, Sakakibara I, Naito M, Osborne TF, Hamakubo T, Ito S, Aburatani H, et al. COUP-TFII acts downstream of Wnt/beta-catenin signal to silence PPARgamma gene expression and repress adipogenesis. Proc Natl Acad Sci U S A. 2009; 106: 5819–24. https://doi.org/10.1073/pnas.0901676106.
244. Ajuwon KM, Spurlock ME. Adiponectin inhibits LPS-induced NF-kappaB activation and IL-6 production and increases PPARgamma2 expression in adipocytes. Am J Physiol Regul Integr Comp Physiol. 2005; 288: R1220–5. https://doi.org/10.1152/ajpregu.00397.2004.
245. Wei J, Ghosh AK, Sargent JL, Komura K, Wu M, Huang QQ, Jain M, Whitfield ML, Feghali-Bostwick C, Varga J. PPARγ downregulation by TGFß in fibroblast and impaired expression and function in systemic sclerosis: a novel mechanism for progressive fibrogenesis. PLoS One. 2010; 5: e13778. https://doi.org/10.1371/journal.pone.0013778.
246. Culver DA, Barna BP, Raychaudhuri B, Bonfield TL, Abraham S, Malur A, Farver CF, Kavuru MS, Thomassen MJ. Peroxisome proliferator-activated receptor gamma activity is deficient in alveolar macrophages in pulmonary sarcoidosis. Am J Respir Cell Mol Biol. 2004; 30: 1–5. https://doi.org/10.1165/rcmb.2003-0304RC.
247. Miyahara T, Schrum L, Rippe R, Xiong S, Yee HF, Motomura K, Anania FA, Willson TM, Tsukamoto H. Peroxisome proliferator-activated receptors and hepatic stellate cell activation. J Biol Chem. 2000; 275: 35715–22. https://doi.org/10.1074/jbc.M006577200.
248. Zheng F, Fornoni A, Elliot SJ, Guan Y, Breyer MD, Striker LJ, Striker GE. Upregulation of type I collagen by TGF-beta in mesangial cells is blocked by PPARgamma activation. Am J Physiol Renal Physiol. 2002; 282: F639–48. https://doi.org/10.1152/ajprenal.00189.2001.
249. Karnik P, Tekeste Z, McCormick TS, Gilliam AC, Price VH, Cooper KD, Mirmirani P. Hair follicle stem cell-specific PPARgamma deletion causes scarring alopecia. J Invest Dermatol. 2009; 129: 1243–57. https://doi.org/10.1038/jid.2008.369.
250. Dantas AT, Pereira MC, de Melo Rego MJ, da Rocha LF Jr, da Rocha Pitta I, Lopes Marques CD, Branco Pinto Duarte AL, Galdino da Rocha Pitta M. The role of PPAR gamma in systemic sclerosis. PPAR Res. 2015; 2015: 124624. https://doi.org/10.1155/2015/124624.
251. Ghosh AK, Wei J, Wu M, Varga J. Constitutive Smad signaling and Smad-dependent collagen gene expression in mouse embryonic fibroblasts lacking peroxisome proliferator-activated receptor-gamma. Biochem Biophys Res Commun. 2008; 374: 231–6. https://doi.org/10.1016/j.bbrc.2008.07.014.
252. Kapoor M, McCann M, Liu S, Huh K, Denton CP, Abraham DJ, Leask A. Loss of peroxisome proliferator-activated receptor gamma in mouse fibroblasts results in increased susceptibility to bleomycin-induced skin fibrosis. Arthritis Rheum. 2009; 60: 2822–9. https://doi.org/10.1002/art.24761.
253. Ghosh AK, Bhattacharyya S, Lakos G, Chen SJ, Mori Y, Varga J. Disruption of transforming growth factor beta signaling and profibrotic responses in normal skin fibroblasts by peroxisome proliferator-activated receptor gamma. Arthritis Rheum. 2004; 50: 1305–18. https://doi.org/10.1002/art.20104.
254. Ghosh AK, Bhattacharyya S, Wei J, Kim S, Barak Y, Mori Y, Varga J. Peroxisome proliferator-activated receptor-gamma abrogates Smad-dependent collagen stimulation by targeting the p300 transcriptional coactivator. FASEB J. 2009; 23: 2968–77. https://doi.org/10.1096/fj.08-128736.
255. Lakota K, Wei J, Carns M, Hinchcliff M, Lee J, Whitfield ML, Sodin-Semrl S, Varga J. Levels of adiponectin, a marker for PPAR-gamma activity, correlate with skin fibrosis in systemic sclerosis: potential utility as biomarker? Arthritis Res Ther. 2012; 14: R102. https://doi.org/10.1186/ar3827.
256. Ramirez A, Ballard EN, Roman J. TGFβ1 controls PPARγ expression, transcriptional potential, and activity, in part, through Smad3 signaling in murine lung fibroblasts. PPAR Res. 2012; 2012: 375876. https://doi.org/10.1155/2012/375876.
257. Wei J, Zhu H, Komura K, Lord G, Tomcik M, Wang W, Doniparthi S, Tamaki Z, Hinchcliff M, Distler JH, Varga J. A synthetic PPAR-γ agonist triterpenoid ameliorates experimental fibrosis: PPAR-γ-independent suppression of fibrotic responses. Ann Rheum Dis. 2014; 73: 446–54. https://doi.org/10.1136/annrheumdis-2012-202716.
258. Burgess HA, Daugherty LE, Thatcher TH, Lakatos HF, Ray DM, Redonnet M, Phipps RP, Sime PJ. PPARgamma agonists inhibit TGF-beta induced pulmonary myofibroblast differentiation and collagen production: implications for therapy of lung fibrosis. Am J Physiol Lung Cell Mol Physiol. 2005; 288: L1146–53. https://doi.org/10.1152/ajplung.00383.2004.
259. Marra F, Efsen E, Romanelli RG, Caligiuri A, Pastacaldi S, Batignani G, Bonacchi A, Caporale R, Laffi G, Pinzani M, Gentilini P. Ligands of peroxisome proliferator-activated receptor gamma modulate profibrogenic and proinflammatory actions in hepatic stellate cells. Gastroenterology. 2000; 119: 466–78.
260. Genovese T, Cuzzocrea S, Di Paola R, Mazzon E, Mastruzzo C, Catalano P, Sortino M, Crimi N, Caputi AP, Thiemermann C, Vancheri C. Effect of rosiglitazone and 15-deoxy-Delta12,14-prostaglandin J2 on bleomycin-induced lung injury. Eur Respir J. 2005; 25: 225–34. https://doi.org/10.1183/09031936.05.00049704.
261. Iglarz M, Touyz RM, Viel EC, Paradis P, Amiri F, Diep QN, Schiffrin EL. Peroxisome proliferator-activated receptor-alpha and receptor-gamma activators prevent cardiac fibrosis in mineralocorticoid-dependent hypertension. Hypertension. 2003; 42: 737–43. https://doi.org/10.1161/01.HYP.0000083511.91817.B1.
262. Maejima Y, Okada H, Haraguchi G, Onai Y, Kosuge H, Suzuki JI, Isobe M. Telmisartan, a unique ARB, improves left ventricular remodeling of infarcted heart by activating PPAR gamma. Lab Invest. 2011; 91: 932–44. https://doi.org/10.1038/labinvest.2011.45.
263. Aoki Y, Maeno T, Aoyagi K, Ueno M, Aoki F, Aoki N, Nakagawa J, Sando Y, Shimizu Y, Suga T, Arai M, Kurabayashi M. Pioglitazone, a peroxisome proliferator-activated receptor gamma ligand, suppresses bleomycin-induced acute lung injury and fibrosis. Respir Int Rev Thorac Dis. 2009; 77: 311–9. https://doi.org/10.1159/000168676.
264. Milam JE, Keshamouni VG, Phan SH, Hu B, Gangireddy SR, Hogaboam CM, Standiford TJ, Thannickal VJ, Reddy RC. PPAR-gamma agonists inhibit profibrotic phenotypes in human lung fibroblasts and bleomycin-induced pulmonary fibrosis. Am J Physiol Lung Cell Mol Physiol. 2008; 294: L891–901. https://doi.org/10.1152/ajplung.00333.2007.
265. Kawaguchi K, Sakaida I, Tsuchiya M, Omori K, Takami T, Okita K. Pioglitazone prevents hepatic steatosis, fibrosis, and enzyme-altered lesions in rat liver cirrhosis induced by a choline-deficient L-amino acid-defined diet. Biochem Biophys Res Commun. 2004; 315: 187–95. https://doi.org/10.1016/j.bbrc.2004.01.038.
266. Uto H, Nakanishi C, Ido A, Hasuike S, Kusumoto K, Abe H, Numata M, Nagata K, Hayashi K, Tsubouchi H. The peroxisome proliferator-activated receptor-gamma agonist, pioglitazone, inhibits fat accumulation and fibrosis in the livers of rats fed a choline-deficient, l-amino acid-defined diet. Hepatol Res. 2005; 32: 235–42. https://doi.org/10.1016/j.hepres.2005.05.008.
267. Calkin AC, Giunti S, Jandeleit-Dahm KA, Allen TJ, Cooper ME, Thomas MC. PPAR-alpha and -gamma agonists attenuate diabetic kidney disease in the apolipoprotein E knockout mouse. Nephrol Dial Transplant. 2006; 21: 2399–405. https://doi.org/10.1093/ndt/gfl212.
268. Kiss E, Popovic ZV, Bedke J, Adams J, Bonrouhi M, Babelova A, Schmidt C, Edenhofer F, Zschiedrich I, Domhan S, Abdollahi A, Schäfer L, Gretz N, et al. Peroxisome proliferator-activated receptor (PPAR)gamma can inhibit chronic renal allograft damage. Am J Pathol. 2010; 176: 2150–62. https://doi.org/10.2353/ajpath.2010.090370.
269. Kippenberger S, Loitsch SM, Grundmann-Kollmann M, Simon S, Dang TA, Hardt-Weinelt K, Kaufmann R, Bernd A. Activators of peroxisome proliferator-activated receptors protect human skin from ultraviolet-B-light-induced inflammation. J Invest Dermatol. 2001; 117: 1430–6. https://doi.org/10.1046/j.0022-202x.2001.01537.x.
270. Linard C, Grémy O, Benderitter M. Reduction of peroxisome proliferation-activated receptor gamma expression by gamma-irradiation as a mechanism contributing to inflammatory response in rat colon: modulation by the 5-aminosalicylic acid agonist. J Pharmacol Exp Ther. 2008; 324: 911–20. https://doi.org/10.1124/jpet.107.129122.
271. Robbins ME, Zhao W. Chronic oxidative stress and radiation-induced late normal tissue injury: a review. Int J Radiat Biol. 2004; 80: 251–9. https://doi.org/10.1080/09553000410001692726.
272. Spitz DR, Azzam EI, Li JJ, Gius D. Metabolic oxidation/reduction reactions and cellular responses to ionizing radiation: a unifying concept in stress response biology. Cancer Metastasis Rev. 2004; 23: 311–22. https://doi.org/10.1023/B:CANC.0000031769.14728.bc.
273. Zhao W, Payne V, Tommasi E, Diz DI, Hsu FC, Robbins ME. Administration of the peroxisomal proliferator-activated receptor gamma agonist pioglitazone during fractionated brain irradiation prevents radiation-induced cognitive impairment. Int J Radiat Oncol Biol Phys. 2007; 67: 6–9. https://doi.org/10.1016/j.ijrobp.2006.09.036.
274. Ramanan S, Kooshki M, Zhao W, Hsu FC, Robbins ME. PPARalpha ligands inhibit radiation-induced microglial inflammatory responses by negatively regulating NF-kappaB and AP-1 pathways. Free Radic Biol Med. 2008; 45: 1695–704. https://doi.org/10.1016/j.freeradbiomed.2008.09.002.
275. Crosby MB, Zhang J, Nowling TM, Svenson JL, Nicol CJ, Gonzalez FJ, Gilkeson GS. Inflammatory modulation of PPAR gamma expression and activity. Clin Immunol. 2006; 118: 276–83. https://doi.org/10.1016/j.clim.2005.09.018.
276. Han EJ, Im CN, Park SH, Moon EY, Hong SH. Combined treatment with peroxisome proliferator-activated receptor (PPAR) gamma ligands and gamma radiation induces apoptosis by PPARγ-independent up-regulation of reactive oxygen species-induced deoxyribonucleic acid damage signals in non-small cell lung cancer cells. Int J Radiat Oncol Biol Phys. 2013; 85: e239–48. https://doi.org/10.1016/j.ijrobp.2012.11.040.
277. Zand H, Rahimipour A, Salimi S, Shafiee SM. Docosahexaenoic acid sensitizes Ramos cells to gamma-irradiation-induced apoptosis through involvement of PPAR-gamma activation and NF-kappaB suppression. Mol Cell Biochem. 2008; 317: 113–20. https://doi.org/10.1007/s11010-008-9838-x.
278. Rehman MU, Tahir M, Quaiyoom Khan A, Khan R, Lateef A, Hamiza OO, Ali F, Sultana S. Diosmin protects against trichloroethylene-induced renal injury in Wistar rats: plausible role of p53, Bax and caspases. Br J Nutr. 2013; 110: 699–710. https://doi.org/10.1017/S0007114512005752.
279. Lecarpentier Y, Claes V, Vallée A, Hébert JL. Interactions between PPAR gamma and the canonical Wnt/beta-catenin pathway in type 2 diabetes and colon cancer. PPAR Res. 2017; 2017: 1–9. https://doi.org/10.1155/2017/5879090.
280. Lecarpentier Y, Claes V, Vallée A, Hébert JL. Thermodynamics in cancers: opposing interactions between PPAR gamma and the canonical WNT/beta-catenin pathway. Clin Transl Med. 2017; 6: 14. https://doi.org/10.1186/s40169-017-0144-7.
281. Lecarpentier Y, Vallée A. Opposite interplay between PPAR gamma and canonical Wnt/beta-catenin pathway in amyotrophic lateral sclerosis. Front Neurol. 2016; 7: 100. https://doi.org/10.3389/fneur.2016.00100.
282. Vallée A, Lecarpentier Y, Guillevin R, Vallée JN. Thermodynamics in gliomas: interactions between the canonical WNT/beta-catenin pathway and PPAR gamma. Front Physiol. 2017; 8: 352. https://doi.org/10.3389/fphys.2017.00352.
283. Vallée A, Lecarpentier Y, Guillevin R, Vallée JN. Aerobic glycolysis hypothesis through WNT/beta-catenin pathway in exudative age-related macular degeneration. J Mol Neurosci. 2017. https://doi.org/10.1007/s12031-017-0947-4.
284. Vallée A, Lecarpentier Y. Alzheimer disease: crosstalk between the canonical Wnt/beta-catenin pathway and PPARs alpha and gamma. Front Neurosci. 2016; 10: 459. https://doi.org/10.3389/fnins.2016.00459.
285. Sharma C, Pradeep A, Wong L, Rana A, Rana B. Peroxisome proliferator-activated receptor gamma activation can regulate beta-catenin levels via a proteasome-mediated and adenomatous polyposis coli-independent pathway. J Biol Chem. 2004; 279: 35583–94. https://doi.org/10.1074/jbc.M403143200.
286. Liu J, Wang H, Zuo Y, Farmer SR. Functional interaction between peroxisome proliferator-activated receptor gamma and beta-catenin. Mol Cell Biol. 2006; 26: 5827–37. https://doi.org/10.1128/MCB.00441-06.
287. Takada I, Kouzmenko AP, Kato S. Wnt and PPARgamma signaling in osteoblastogenesis and adipogenesis. Nat Rev Rheumatol. 2009; 5: 442–7. https://doi.org/10.1038/nrrheum.2009.137.
288. Lu D, Carson DA. Repression of beta-catenin signaling by PPAR gamma ligands. Eur J Pharmacol. 2010; 636: 198–202. https://doi.org/10.1016/j.ejphar.2010.03.010.
289. Moldes M, Zuo Y, Morrison RF, Silva D, Park BH, Liu J, Farmer SR. Peroxisome-proliferator-activated receptor gamma suppresses Wnt/beta-catenin signalling during adipogenesis. Biochem J. 2003; 376: 607–13. https://doi.org/10.1042/BJ20030426.
290. Farshbaf MJ, Ghaedi K, Shirani M, Nasr-Esfahani MH. Peroxisome proliferator activated receptor gamma (PPARγ) as a therapeutic target for improvement of cognitive performance in Fragile-X. Med Hypotheses. 2014; 82: 291–4. https://doi.org/10.1016/j.mehy.2013.12.012.
291. Xu C, Wang J, Zhu T, Shen Y, Tang X, Fang L, Xu Y. Cross-talking between PPAR and WNT signaling and its regulation in mesenchymal stem cell differentiation. Curr Stem Cell Res Ther. 2016; 11: 247–54.
292. Drygiannakis I, Valatas V, Sfakianaki O, Bourikas L, Manousou P, Kambas K, Ritis K, Kolios G, Kouroumalis E. Proinflammatory cytokines induce crosstalk between colonic epithelial cells and subepithelial myofibroblasts: implication in intestinal fibrosis. J Crohns Colitis. 2013; 7: 286–300. https://doi.org/10.1016/j.crohns.2012.04.008.
293. Farmer SR. Regulation of PPARgamma activity during adipogenesis. Int J Obes (Lond). 2005; 29: S13–6. https://doi.org/10.1038/sj.ijo.0802907.
294. Jeon KI, Kulkarni A, Woeller CF, Phipps RP, Sime PJ, Hindman HB, Huxlin KR. Inhibitory effects of PPARγ ligands on TGF-β1-induced corneal myofibroblast transformation. Am J Pathol. 2014; 184: 1429–45. https://doi.org/10.1016/j.ajpath.2014.01.026.
295. Kumar V, Mundra V, Mahato RI. Nanomedicines of Hedgehog inhibitor and PPAR-γ agonist for treating liver fibrosis. Pharm Res. 2014; 31: 1158–69. https://doi.org/10.1007/s11095-013-1239-5.
296. Lee Y, Kim SH, Lee YJ, Kang ES, Lee BW, Cha BS, Kim JW, Song DH, Lee HC. Transcription factor Snail is a novel regulator of adipocyte differentiation via inhibiting the expression of peroxisome proliferator-activated receptor γ. Cell Mol Life Sci. 2013; 70: 3959–71. https://doi.org/10.1007/s00018-013-1363-8.
297. Li Q, Yan Z, Li F, Lu W, Wang J, Guo C. The improving effects on hepatic fibrosis of interferon-γ liposomes targeted to hepatic stellate cells. Nanotechnology. 2012; 23: 265101. https://doi.org/10.1088/0957-4484/23/26/265101.
298. Liu J, Farmer SR. Regulating the balance between peroxisome proliferator-activated receptor gamma and beta-catenin signaling during adipogenesis. A glycogen synthase kinase 3beta phosphorylation-defective mutant of beta-catenin inhibits expression of a subset of adipogenic genes. J Biol Chem. 2004; 279: 45020–7. https://doi.org/10.1074/jbc.M407050200.
299. Qian J, Niu M, Zhai X, Zhou Q, Zhou Y. β-Catenin pathway is required for TGF-β1 inhibition of PPARγ expression in cultured hepatic stellate cells. Pharmacol Res. 2012; 66: 219–25. https://doi.org/10.1016/j.phrs.2012.06.003.
300. Segel MJ, Izbicki G, Cohen PY, Or R, Christensen TG, Wallach-Dayan SB, Breuer R. Role of interferon-gamma in the evolution of murine bleomycin lung fibrosis. Am J Physiol Lung Cell Mol Physiol. 2003; 285: L1255–62. https://doi.org/10.1152/ajplung.00303.2002.
301. Shim CY, Song BW, Cha MJ, Hwang KC, Park S, Hong GR, Kang SM, Lee JE, Ha JW, Chung N. Combination of a peroxisome proliferator-activated receptor-gamma agonist and an angiotensin II receptor blocker attenuates myocardial fibrosis and dysfunction in type 2 diabetic rats. J Diabetes Investig. 2014; 5: 362–71. https://doi.org/10.1111/jdi.12153.
302. Sabatino L, Pancione M, Votino C, Colangelo T, Lupo A, Novellino E, Lavecchia A, Colantuoni V. Emerging role of the β-catenin-PPARγ axis in the pathogenesis of colorectal cancer. World J Gastroenterol. 2014; 20: 7137–51. https://doi.org/10.3748/wjg.v20.i23.7137.
303. Akinyeke TO, Stewart LV. Troglitazone suppresses c-Myc levels in human prostate cancer cells via a PPARγ-independent mechanism. Cancer Biol Ther. 2011; 11: 1046–58.
304. Di Gregorio J, Sferra R, Speca S, Vetuschi A, Dubuquoy C, Desreumaux P, Pompili S, Cristiano L, Gaudio E, Flati V, Latella G. Role of glycogen synthase kinase-3β and PPAR-γ on epithelial-to-mesenchymal transition in DSS-induced colorectal fibrosis. PLoS One. 2017; 12: e0171093. https://doi.org/10.1371/journal.pone.0171093.
305. Gustafson B, Eliasson B, Smith U. Thiazolidinediones increase the wingless-type MMTV integration site family (WNT) inhibitor Dickkopf-1 in adipocytes: a link with osteogenesis. Diabetologia. 2010; 53: 536–40. https://doi.org/10.1007/s00125-009-1615-1.
306. Prusty D, Park BH, Davis KE, Farmer SR. Activation of MEK/ERK signaling promotes adipogenesis by enhancing peroxisome proliferator-activated receptor gamma (PPARgamma) and C/EBPalpha gene expression during the differentiation of 3T3-L1 preadipocytes. J Biol Chem. 2002; 277: 46226–32. https://doi.org/10.1074/jbc.M207776200.
307. Kim S, Ha JM, Yun SJ, Kim EK, Chung SW, Hong KW, Kim CD, Bae SS. Transcriptional activation of peroxisome proliferator-activated receptor-gamma requires activation of both protein kinase A and Akt during adipocyte differentiation. Biochem Biophys Res Commun. 2010; 399: 55–9. https://doi.org/10.1016/j.bbrc.2010.07.038.
308. Peng XD, Xu PZ, Chen ML, Hahn-Windgassen A, Skeen J, Jacobs J, Sundararajan D, Chen WS, Crawford SE, Coleman KG, Hay N. Dwarfism, impaired skin development, skeletal muscle atrophy, delayed bone development, and impeded adipogenesis in mice lacking Akt1 and Akt2. Genes Dev. 2003; 17: 1352–65. https://doi.org/10.1101/gad.1089403.
309. Zhang HH, Huang J, Düvel K, Boback B, Wu S, Squillace RM, Wu CL, Manning BD. Insulin stimulates adipogenesis through the Akt-TSC2-mTORC1 pathway. PLoS One. 2009; 4: e6189. https://doi.org/10.1371/journal.pone.0006189.
310. Grimes CA, Jope RS. The multifaceted roles of glycogen synthase kinase 3beta in cellular signaling. Prog Neurobiol. 2001; 65: 391–426.
311. Huelsken J, Behrens J. The Wnt signalling pathway. J Cell Sci. 2002; 115: 3977–8.
312. Park HJ, Yun J, Jang SH, Kang SN, Jeon BS, Ko YG, Kim HD, Won CK, Kim GS, Cho JH. Coprinus comatus cap inhibits adipocyte differentiation via regulation of PPARγ and Akt signaling pathway. PLoS One. 2014; 9: e105809. https://doi.org/10.1371/journal.pone.0105809.
313. Aljada A, O’Connor L, Fu YY, Mousa SA. PPAR gamma ligands, rosiglitazone and pioglitazone, inhibit bFGF- and VEGF-mediated angiogenesis. Angiogenesis. 2008; 11: 361–7. https://doi.org/10.1007/s10456-008-9118-0.
314. Goetze S, Eilers F, Bungenstock A, Kintscher U, Stawowy P, Blaschke F, Graf K, Law RE, Fleck E, Gräfe M. PPAR activators inhibit endothelial cell migration by targeting Akt. Biochem Biophys Res Commun. 2002; 293: 1431–7. https://doi.org/10.1016/S0006-291X(02)00385-6.
315. Ma ZG, Yuan YP, Zhang X, Xu SC, Wang SS, Tang QZ. Piperine attenuates pathological cardiac fibrosis via PPAR-γ/AKT pathways. EBioMedicine. 2017; 18: 179–87. https://doi.org/10.1016/j.ebiom.2017.03.021.
316. Zheng S, Chen A. Disruption of transforming growth factor-beta signaling by curcumin induces gene expression of peroxisome proliferator-activated receptor-gamma in rat hepatic stellate cells. Am J Physiol Gastrointest Liver Physiol. 2007; 292: G113–23. https://doi.org/10.1152/ajpgi.00200.2006.
317. Wu M, Melichian DS, Chang E, Warner-Blankenship M, Ghosh AK, Varga J. Rosiglitazone abrogates bleomycin-induced scleroderma and blocks profibrotic responses through peroxisome proliferator-activated receptor-gamma. Am J Pathol. 2009; 174: 519–33. https://doi.org/10.2353/ajpath.2009.080574.
318. Nan YM, Fu N, Wu WJ, Liang BL, Wang RQ, Zhao SX, Zhao JM, Yu J. Rosiglitazone prevents nutritional fibrosis and steatohepatitis in mice. Scand J Gastroenterol. 2009; 44: 358–65. https://doi.org/10.1080/00365520802530861.
319. Zhao C, Chen W, Yang L, Chen L, Stimpson SA, Diehl AM. PPARgamma agonists prevent TGFbeta1/Smad3-signaling in human hepatic stellate cells. Biochem Biophys Res Commun. 2006; 350: 385–91. https://doi.org/10.1016/j.bbrc.2006.09.069.
320. Li Y, Wen X, Spataro BC, Hu K, Dai C, Liu Y. hepatocyte growth factor is a downstream effector that mediates the antifibrotic action of peroxisome proliferator-activated receptor-gamma agonists. J Am Soc Nephrol. 2006; 17: 54–65. https://doi.org/10.1681/ASN.2005030257.
321. Zhao Y, Huang Y, He J, Li C, Deng W, Ran X, Wang D. Rosiglitazone, a peroxisome proliferator-activated receptor-γ agonist, attenuates airway inflammation by inhibiting the proliferation of effector T cells in a murine model of neutrophilic asthma. Immunol Lett. 2014; 157: 9–15. https://doi.org/10.1016/j.imlet.2013.11.004.
322. De Backer O, Elinck E, Priem E, Leybaert L, Lefebvre RA. Peroxisome proliferator-activated receptor gamma activation alleviates postoperative ileus in mice by inhibition of Egr-1 expression and its downstream target genes. J Pharmacol Exp Ther. 2009; 331: 496–503. https://doi.org/10.1124/jpet.109.155135.
323. Handy JA, Fu PP, Kumar P, Mells JE, Sharma S, Saxena NK, Anania FA. Adiponectin inhibits leptin signalling via multiple mechanisms to exert protective effects against hepatic fibrosis. Biochem J. 2011; 440: 385–95. https://doi.org/10.1042/BJ20102148.
324. Shafiei MS, Shetty S, Scherer PE, Rockey DC. Adiponectin regulation of stellate cell activation via PPARγ-dependent and -independent mechanisms. Am J Pathol. 2011; 178: 2690–9. https://doi.org/10.1016/j.ajpath.2011.02.035.
325. Yao Q, Ayala ER, Qian JQ, Stenvinkel P, Axelsson J, Lindholm B. A combination of a PPAR-gamma agonist and an angiotensin II receptor blocker attenuates proinflammatory signaling and stimulates expression of Smad7 in human peritoneal mesothelial cells. Clin Nephrol. 2007; 68: 295–301.
326. Zhou D, Wang J, He LN, Li BH, Ding YN, Chen YW, Fan JG. Prolyl oligopeptidase attenuates hepatic stellate cell activation through induction of Smad7 and PPAR-γ. Exp Ther Med. 2017; 13: 780–6. https://doi.org/10.3892/etm.2017.4033.
327. Reka AK, Kurapati H, Narala VR, Bommer G, Chen J, Standiford TJ, Keshamouni VG. Peroxisome proliferator-activated receptor-gamma activation inhibits tumor metastasis by antagonizing Smad3-mediated epithelial-mesenchymal transition. Mol Cancer Ther. 2010; 9: 3221–32. https://doi.org/10.1158/1535-7163.MCT-10-0570.
328. Tan X, Dagher H, Hutton CA, Bourke JE. Effects of PPAR gamma ligands on TGF-beta1-induced epithelial-mesenchymal transition in alveolar epithelial cells. Respir Res. 2010; 11: 21. https://doi.org/10.1186/1465-9921-11-21.
329. Lee SJ, Yang EK, Kim SG. Peroxisome proliferator-activated receptor-gamma and retinoic acid X receptor alpha represses the TGFbeta1 gene via PTEN-mediated p70 ribosomal S6 kinase-1 inhibition: role for Zf9 dephosphorylation. Mol Pharmacol. 2006; 70: 415–25. https://doi.org/10.1124/mol.106.022954.
330. Kulkarni AA, Thatcher TH, Olsen KC, Maggirwar SB, Phipps RP, Sime PJ. PPAR-γ ligands repress TGFβ-induced myofibroblast differentiation by targeting the PI3K/Akt pathway: implications for therapy of fibrosis. PLoS One. 2011; 6: e15909. https://doi.org/10.1371/journal.pone.0015909.
331. Bu S, Asano Y, Bujor A, Highland K, Hant F, Trojanowska M. Dihydrosphingosine 1-phosphate has a potent antifibrotic effect in scleroderma fibroblasts via normalization of phosphatase and tensin homolog levels. Arthritis Rheum. 2010; 62: 2117–26. https://doi.org/10.1002/art.27463.
332. Kuwano K. PTEN as a new agent in the fight against fibrogenesis. Am J Respir Crit Care Med. 2006; 173: 5–6. https://doi.org/10.1164/rccm.2510001.
333. Deng YL, Xiong XZ, Cheng NS. Organ fibrosis inhibited by blocking transforming growth factor-β signaling via peroxisome proliferator-activated receptor γ agonists. Hepatobiliary Pancreat Dis Int. 2012; 11: 467–78.
334. Kattla JJ, Carew RM, Heljic M, Godson C, Brazil DP. Protein kinase B/Akt activity is involved in renal TGF-beta1-driven epithelial-mesenchymal transition in vitro and in vivo. Am J Physiol Renal Physiol. 2008; 295: F215–25. https://doi.org/10.1152/ajprenal.00548.2007.
335. Zhou B, Buckley ST, Patel V, Liu Y, Luo J, Krishnaveni MS, Ivan M, DeMaio L, Kim KJ, Ehrhardt C, Crandall ED, Borok Z. Troglitazone attenuates TGF-β1-induced EMT in alveolar epithelial cells via a PPARγ-independent mechanism. PLoS One. 2012; 7: e38827. https://doi.org/10.1371/journal.pone.0038827.
336. Zhou B, Liu Y, Kahn M, Ann DK, Han A, Wang H, Nguyen C, Flodby P, Zhong Q, Krishnaveni MS, Liebler JM, Minoo P, Crandall ED, et al. Interactions between β-catenin and transforming growth factor-β signaling pathways mediate epithelial-mesenchymal transition and are dependent on the transcriptional co-activator cAMP-response element-binding protein (CREB)-binding protein (CBP). J Biol Chem. 2012; 287: 7026–38. https://doi.org/10.1074/jbc.M111.276311.
337. Rosenbloom J, Castro SV, Jimenez SA. Narrative review: fibrotic diseases: cellular and molecular mechanisms and novel therapies. Ann Intern Med. 2010; 152: 159–66. https://doi.org/10.7326/0003-4819-152-3-201002020-00007.
338. Wei J, Bhattacharyya S, Varga J. Peroxisome proliferator-activated receptor γ: innate protection from excessive fibrogenesis and potential therapeutic target in systemic sclerosis. Curr Opin Rheumatol. 2010; 22: 671–6. https://doi.org/10.1097/BOR.0b013e32833de1a7.