
INTRODUCTION
Osteoarthritis (OA) is a common degenerative disorder which affects the function of joints, causes pains and reduces quality of life in patients. In the pathogenesis of osteoarthritis, chondrocytes in the cartilage undergo hypertrophic change and the cartilage-specific extracellular matrix (ECM) gradually degrades. Blood vessels and nerves invade the cartilage thereafter, exacerbating the condition of affected joint [1]. Currently, there is no identified cause of blood vessel invasion and effective strategy for the treatment of osteoarthritis [2].
Increased angiogenic factors and/or decreased anti-angiogenic factors promote blood vessel formation [3]. Although there is no blood vessel in normal cartilage, there are angiogenic factors, such as fibroblast growth factor (FGF) and vascular endothelial growth factor (VEGF), arising from synovium and subchondral bone [4]. Therefore, cartilage possesses the potential of angiogenesis. However, the capability of angiogenesis in cartilage may be inhibited by anti-angiogenic factors in the microenvironment, which helps maintain the characteristics of healthy cartilage. Once the balance is broken, excessive angiogenic factors will promote blood vessel formation, as it was evidenced in inflammatory conditions [5]. As blood vessels play a devastating role in osteoarthritis, inhibition of angiogenesisis a strategy for the treatment. Therefore, elucidating the mechanisms of angiogenesis in osteoarthritis will be of great importance [6].
Transforming growth factor β1 (TGFβ1) has been investigated in osteoarthritis and chondrocytes for years [7, 8]. However, reports of its function varied in published data from different laboratories [9–11]. Previously, most studies reported that TGFβ1 was essential for cartilage integrity and ECM maintenance. Blockage of TGFβ1 signaling resulted in cartilage degeneration and osteoarthritis-like tissue formation [7]. More recent studies have demonstrated that TGFβ1 might accelerate the degeneration of cartilage, including hypertrophy, angiogenesis and ECM mineralization [12]. In patients with osteoarthritis, there was high level of TGFβ1 in subchondral bone and inhibition of the activity of TGFβ1 effectively attenuated the degradation of cartilage ECM, suggesting that high level of TGFβ1 in subchondral bone be a trigger of osteoarthritis [13, 14]. Further studies revealed that TGFβ1 could induce the aggregation of mesenchymal stem cells (MSC) and induce the blood vessel formation in osteoarthritis cartilage [13]. However, whether TGFβ1 induces morphological change and expression of angiogenic factors in chondrocytes cultured in vitro and the corresponding mechanisms were not identified [15].
In the present study, we investigated the effect of TGFβ1 on human chondrocytes cultured in vitro. DNA microarray was performed to screen the differentially expressed genes (DEGs) in chondrocytes culture. It was found that genes involved in chondrocyte hypertrophy and blood vessel development were significantly upregulated in TGFβ1 treated chondrocytes. Our results demonstrated that human chondrocytes treated with TGFβ1 in vitro undergo hypertrophic change and be induced to express angiogenic factors.
RESULTS
Isolation and identification of human articular chondrocytes
Collagen type II, type I and glycosaminoglycan (GAG) were the primary components in the cartilage ECM and were used as the criteria for the identification and evaluation of chondrocytes [9, 18]. Human articular chondrocytes were isolated and evaluated with toluidine blue staining, immunocytochemical staining and quantitative RT-PCR. Toluidine blue staining revealed production of GAGs decreased in -passage 1(P1) and passage 2(P2) cells compared with primary P0 cells (Figure 1A). In immunocytochemical staining, Col2A1 was detected only in P0 chondrocytes as the signals were very weak in chondrocytes of P1 and later passages (Figure 1B). Col1A1 was detected in all passages (Figure 1B). Quantitative RT-PCR demonstrated a decreased expression of Col2A1 and an increased expression of Col1A1 during cell passaging (Figure 1C). These results suggested that chondrocytes underwent dedifferentiation during passaging.
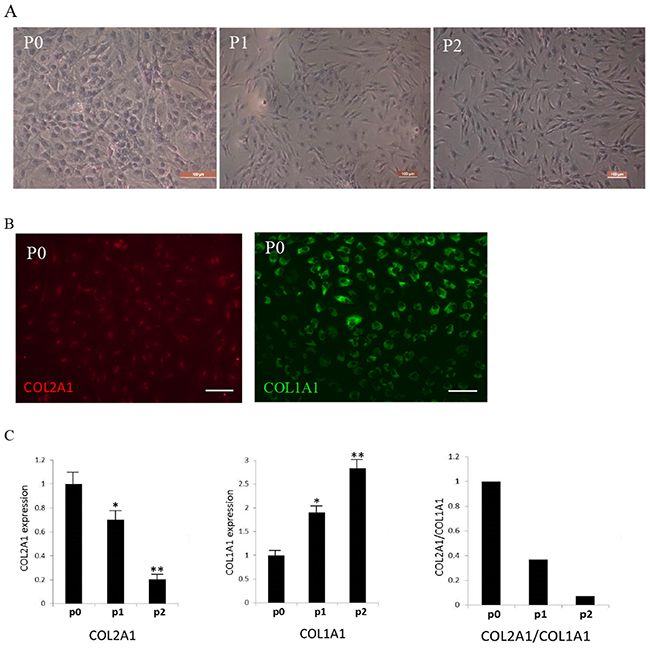
Figure 1: Identification of isolated human articular chondrocytes. (A) Toluidine blue staining (TB) of P0-P2 chondrocytes. (B) P0 chondrocytes were immunostained with type I (COL1A1) and type II collagen (COL2A1). (C) Examination of COL2A1 and COL1A1 expression by quantitative RT-PCR. * significant difference in gene expression (**P<0.01 and *P<0.05). Scale bar: 100μm.
Cells aggregated upon TGFβ1 treatment
Generally, chondrocytes cultured in vitro were evenly distributed on the plate (Figure 2A). Upon TGFβ1 treatment, cells moved and aggregated, forming network-like structure after 3 days in culture (Figure 2B-2D). Different concentrations of TGFβ1 have been used in the experiment. A concentration of more than 2ng/ml was high enough to promote chondrocytes to aggregate. The number of junctions, total length and total segment length increased with increasing TGFβ1 concentrations. Additionally, chondrocytes of P0 to P2 passages formed network-like structure with TGFβ1 treatment and data of the P0 to P2 were very close, with P0 slightly higher. Representative images of chondrocytes from P0 to P2 treated with TGFβ1 were shown in Figure 3.
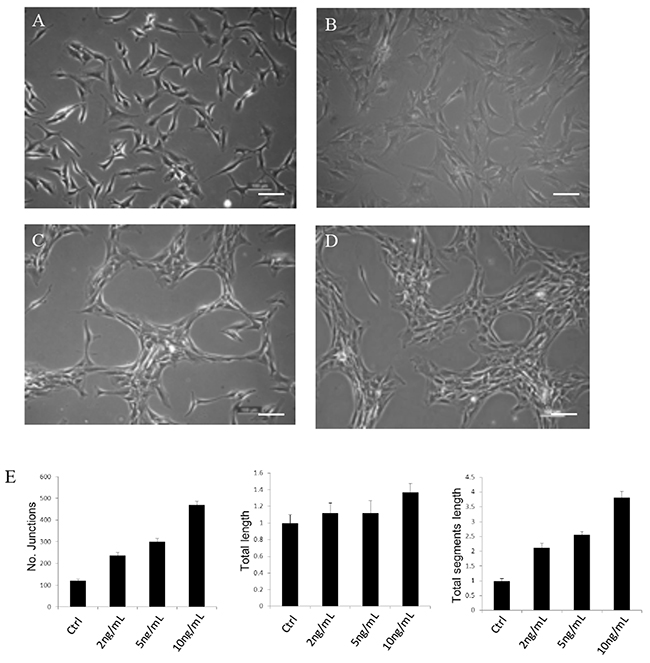
Figure 2: Representative cell images of chondrocytes taken at day 3 treated without TGFβ1 (A) and with 2ng/ml (B) 5ng/ml (C) 10ng/ml (D) TGFβ1 and quantitative analysis of network formation. Images shown are representative of at least three separate experiments. Cells started to aggregate after treatment with TGFβ1 for 1 day and form network-like structure in about 3 days. A concentration of more than 2ng/ml was high enough to induce network formation. Scale bar: 100μm.
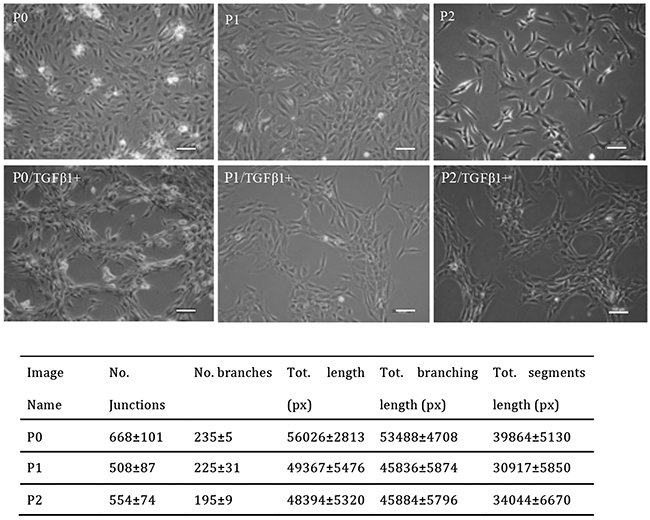
Figure 3: Representative images of P0 to P2 chondrocytes treated with 10ng/ml TGFβ1 and quantitative analysis of network formation. Images shown are representative of at least three separate experiments. Cells from P0 to P2 started to aggregate after treatment with TGFβ1 for 1 day and form network in 3 days. Scale bar: 100μm.
Differentially expressed genes screened by DNA microarray analysis
A total of 2416 differentially expressed genes (fold change ≥ 5 and fold change≤ 0.2) were found in 10ng/mL TGFβ1-treated chondrocytes. Of these genes, 956 genes were up-regulated and 1450 genes were down-regulated. The top 10 most significantly up- or down-regulated DEGs were listed in Table 1. These genes were involved in cell adhesion and movement (AMIGO2, CLEC18A, CDH2, SEPP1 and ITGB8), cell growth and differentiation (CDKN2B, IL11, TSPAN2 and NTRK2), cellular metabolism (ST6GAL2, STC1, SERPINE1 and CFD), oxidation-reduction process (NOX4,FMO2,FMO3 and AKR1C1) and development (COL10A1, MEST and FOXQ1). According to Gene Ontology (GO) analysis, the DEGs were enriched in system development, blood vessel development, regulation of signaling, response to external stimulus and functions listed in Table 2. KEGG analysis revealed the genes could be classified to gap junction, ECM-receptor interaction and MAPK signaling pathways in Table 3. As expected, TGFβ1 signaling pathway was also detected.
Table 1: The top 10 most significantly up- or down-regulated DEGs
Gene ID |
Gene Symbol |
Gene Title |
Fold Change |
|---|---|---|---|
Up-regulated genes |
|||
AW004016 |
ST6GAL2 |
ST6 beta-galactosamide alpha-2,6-sialyltranferase 2 |
79.4 |
AI376003 |
COL10A1 |
collagen type X alpha 1 |
75.9 |
AC004010 |
AMIGO2 |
adhesion molecule with Ig-like domain 2 |
61.7 |
AW444761 |
CDKN2B |
cyclin-dependent kinase inhibitor 2B |
32.6 |
BF057185 |
CLEC18A |
C-type lectin domain family 18, member A |
31 |
NM_000641.1 |
IL11 |
interleukin 11 |
30.3 |
NM_016931.1 |
NOX4 |
NADPH oxidase 4 |
24.4 |
BF129969 |
TSPAN2 |
tetraspanin 2 |
16.7 |
M34064.1 |
CDH2 |
cadherin 2, type 1, N-cadherin |
12.3 |
BC020765.1 |
SERPINE1 |
serpin peptidase inhibitor, clade E |
9.9 |
Down-regulated genes |
|||
NM_005410 |
SEPP1 |
selenoprotein P, plasma, 1 |
0.013 |
AI758223 |
FMO2 |
flavin containing monooxygenase 2 |
0.021 |
BF513121 |
ITGB8 |
integrin, beta 8 |
0.024 |
AA707199 |
NTRK2 |
neurotrophic tyrosine kinase, receptor, type 2 |
0.025 |
NM_003155 |
STC1 |
stanniocalcin 1 |
0.033 |
NM_002402 |
MEST |
mesoderm specific transcript homolog |
0.037 |
AI676059 |
FOXQ1 |
forkhead box Q1 |
0.042 |
M83772 |
FMO3 |
flavin containing monooxygenase 3 |
0.044 |
BF508244 |
AKR1C1 |
secreted frizzled-related protein 1 |
0.049 |
NM_001928 |
CFD |
complement factor D |
0.05 |
Table 2: The enriched GO functions of DEGs
Names |
Genes_In_Term |
DEG |
Significant |
|---|---|---|---|
GO:0048731//system development |
6697 |
781 |
TRUE |
GO:0048523//negative regulation of cellular process |
6308 |
734 |
TRUE |
GO:0007275//multicellular organismal development |
7875 |
890 |
TRUE |
GO:0048519//negative regulation of biological process |
6805 |
783 |
TRUE |
GO:0009966//regulation of signal transduction |
4170 |
512 |
TRUE |
GO:0032502//developmental process |
8833 |
979 |
TRUE |
GO:0001568//blood vessel development |
891 |
146 |
TRUE |
GO:0048856//anatomical structure development |
7770 |
873 |
TRUE |
GO:0001944//vasculature development |
950 |
152 |
TRUE |
GO:0044699//single-organism process |
21723 |
2173 |
TRUE |
GO:0009605//response to external stimulus |
2033 |
276 |
TRUE |
GO:0010646//regulation of cell communication |
4670 |
556 |
TRUE |
GO:0023051//regulation of signaling |
4645 |
553 |
TRUE |
Guide: Names-pathways, Genes in Term- number of genes in the term, DEG- number of differentially expressed genes.
Table 3: The enriched KEGG pathways of DEGs
Names |
Genes_In_Term |
DEG |
Significant |
|
|---|---|---|---|---|
Total |
20594 |
1788 |
- |
|
ko04540 |
Gap junction |
315 |
55 |
TRUE |
ko04350 |
TGF-beta signaling pathway |
244 |
41 |
TRUE |
ko04512 |
ECM-receptor interaction |
338 |
52 |
TRUE |
ko04115 |
p53 signaling pathway |
250 |
41 |
TRUE |
ko00380 |
Tryptophan metabolism |
123 |
23 |
TRUE |
ko00910 |
Nitrogen metabolism |
72 |
15 |
TRUE |
ko03030 |
DNA replication |
87 |
17 |
TRUE |
ko04510 |
Focal adhesion |
898 |
104 |
TRUE |
ko04010 |
MAPK signaling pathway |
907 |
104 |
TRUE |
Note: Names-pathways, Genes inTerm-number of genes in the term, DEG-number of differentially expressed genes.
Gene expression validation
Genes associated with cellular hypertrophy and in TGFβ signaling pathway were examined by quantitative RT-PCR (Figure 4). Expression of COL2A1 was significantly decreased while COL10A1 drastically increased, suggesting the cells undergo hypertrophic change during the culture. As important molecules in TGFβ signaling pathway, SMAD3 was down-regulated while the inhibitory molecule SMAD7 was up-regulated, which was in consistent with studies by Narcisi R and Yang YH [15, 19].
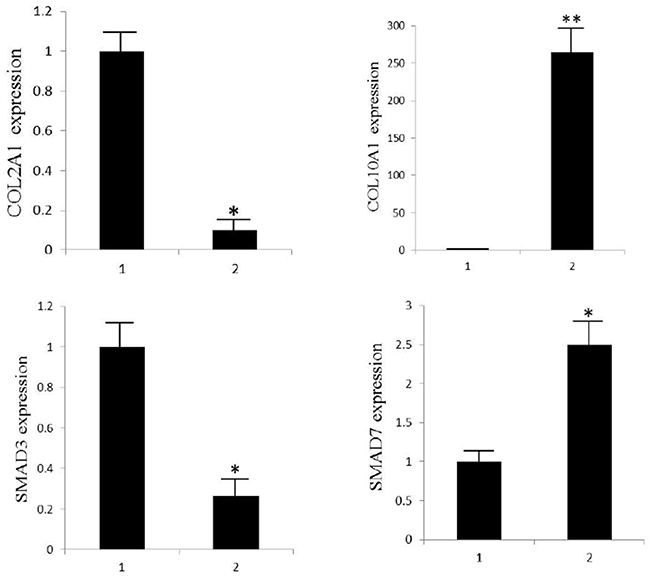
Figure 4: Quantitative RT-PCR analysis of COL2A1, COL10A1, SMAD3 and SMAD7 in TGFβ1 treated and control groups. Transcript levels were assessed on three different cultures; histograms depict average levels; error bars depict standard deviation values. *significant difference in gene expression (**P<0.01 and *P<0.05).
Microarray-screened DEGs involved in blood vessel formation were also examined. Endothelial cell-specific molecule 1 (ESM1), vascular endothelial growth factor receptor 2 (KDR/VEGFR2) and VEGF were selected and validated. In consistent with microarray analysis, expression of all the three genes increased significantly (Figure 5A). ESM1 was further validated by Western blot and Flow Cytometric analysis (Figure 5B–5C), which demonstrated significant increase at the protein level, with 1.53 fold by Western blot analysis.
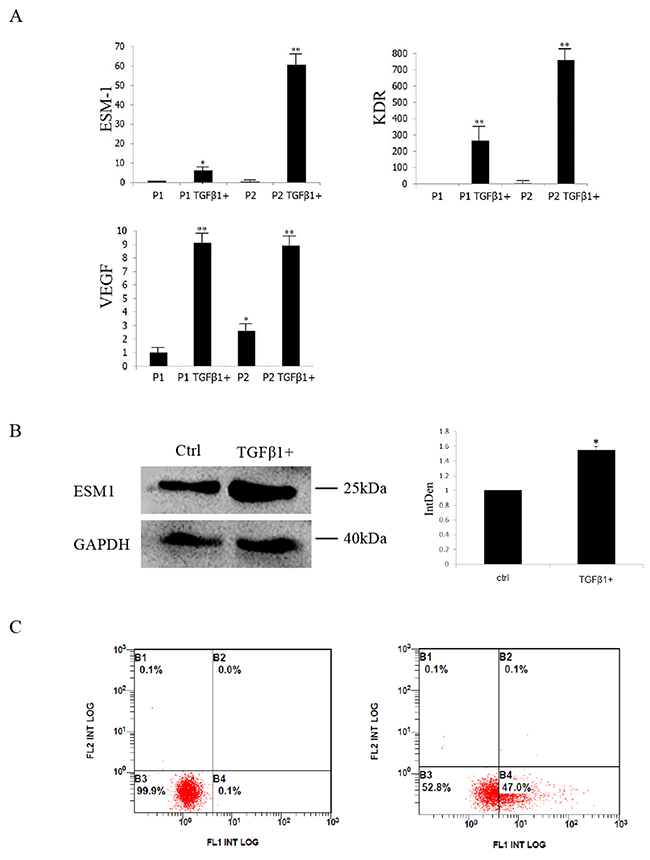
Figure 5: Differentially expressed genes studied in microarray experiments were validated by qRT-PCR, Western blot and Flow Cytometric analysis. (A) Quantitative RT-PCR analysis of ESM1, KDR and VEGF in 10ng/ml TGFβ1 treated and control groups. (B) ESM1 validation by Western blot; GAPDH was used as the internal control. (C) ESM1 validation by Flow Cytometric analysis. ESM1 positive cells increased after TGFβ1 treatment. * significant difference in gene expression (**P<0.01 and *P<0.05).
Network formation of chondrocytes in matrigel
As genes enriched in blood vessel formation were validated and chondrocytes had the capability to form networks in monolayer culture, the cells were then used for a matrigel assay to observe whether the cells could form networks in the gel. As the results showed, cell networks were observed from 2 out of 7 samples (Figure 6). No network formed in the control group.
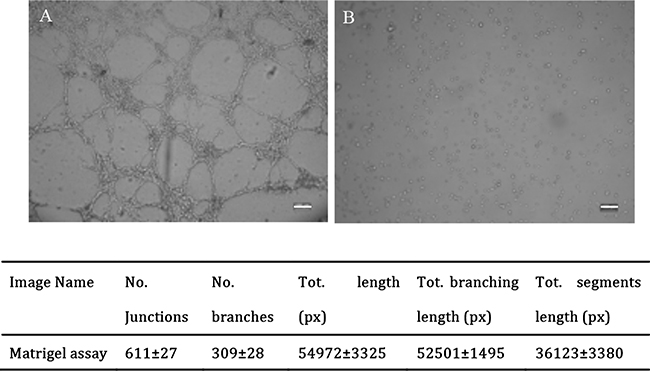
Figure 6: Representative images and quantitative analysis of network formation by chondrocytes in Matrigel assay. P1 chondrocytes seeded in the matrigel formed networks after 5 hours in culture (A) No network was observed in the wells with media plus angiogenesis inhibitor (B) Scale bar: 100μm
DISCUSSION
The TGFβ signaling pathway has been investigated in osteoarthritis and cartilage, but its effects on chondrocyte proliferation and cartilage ECM production were controversial [9, 20, 21]. TGFβ1 can promote upregulation of collagen and prevent loss of proteoglycan in articular cartilage [22, 23]. However, exogenous TGFβ administration induced osteoarthritis-like changes of the cartilage [24]. In the present study, the role of TGFβ1 in human chondrocytes cultured in vitro was investigated.
From our observations, chondrocytes treated with TGFβ1 aggregated with each other, forming network-like structure in the culture plate. Elevated expression of COL10A1 and decreased level of COL2A1 suggested that chondrocytes underwent hypertrophic change, which was in consistent with the research by R.NARCISI [15]. Upon microarray analysis, increased expression of angiogenic factors were detected from the chondrocytes treated with TGFβ1. Surprisingly, cells from 2 out of 7 samples formed network-like structure in matrigel assay. The formation of cell networks might be due to angiogenic factors, endothelial cells or precursor cells in the sample. Another explanation might be the chondrocytes underwent trans-differentiation to endothelial cells after TGFβ1 treatment as it was shown endothelial cells could trans-differentiate to chondrocyte-like cells under high-glucose circumstances [25]. Identification of MSC markers or chondrocyte markers would determine whether the cells underwent trans-differentiation [25, 26]. As cells from 5 out of 7 samples did not form networks in the gel, more investigations are needed. Hypertrophy and angiogenesis are closely integrated processes in cartilage and hypertrophy might contribute to angiogenesis [27, 28]. Therefore, induced hypertrophy and angiogenesis by TGFβ1 suggested TGFβ1 did not play a role in cartilage regeneration.
In osteoarthritis, chondrocytes undergo hypertrophic change and the ECM of the cartilage degenerated, followed by blood vessel invasion. Bone formation is often coupled with chondrocyte hypertrophy and angiogenesis in development. Of the top 10 most up- or down-regulated genes detected, COL10A1 and STC1 were related with chondrocyte hypertrophy and bone formation [29, 30]. SEPP1, which was closely associated with endothelial cells and were able to impair angiogenesis [31], was detected as one of the most down-regulated genes in this study. As TGFβ signaling pathway in endothelial progenitor cells would promote angiogenesis [32], it is possible that the signaling pathway played the same role in chondrocytes. Additionally, molecules related to blood vessel formation were up-regulated after TGFβ1 treatment. VEGF has possibly been involved in the pathogenesis of osteoarthritis [33] and our results provided evidence for its potential role in chondrocytes. Endothelial cell biomarkers, ESM1 and KDR were significantly up-regulated. Matrigel assay further showed that networks were formed by chondrocytes in the matrigel. According to our study, it could be speculated that in vivo, besides synovium and subchondral bone, angiogenic factors in the cartilage could also come from chondrocytes with TGFβ1 stimulation, which would promote the progression of OA through the assistance for blood vessel formation or invasion. However, as in vitro study cannot reflect the real situation in vivo, the functions and mechanisms of TGFβ1 on cartilage and chondrocytes need more explorations in the future.
In summary, the present study investigated the effect of TGFβ1 on human chondrocyte in vitro and found that angiogenic factors could be induced from chondrocytes upon TGFβ1 treatment. Therefore, chondrocytes with TGFβ1 stimulation might also play a role in angiogenesis in osteoarthritis. Although TGFβ1 are widely used to induce chondrogenesis in stem cells [34, 35], it seems it is not appropriate to use it for chondrocyte culture in vitro for practical use, especially for therapeutic purpose such as autologous chondrocyte implantation (ACI). It is speculated that high level of TGFβ1 might play a negative role in cartilage regeneration and chondrocyte culture. Therefore, usage of TGFβ1 for chondrocyte culture in clinic should be considered prudentially. Instead, targeting TGFβ1 or relevant receptors might be a strategy to prevent or alleviate progression of osteoarthritis.
MATERIALS AND METHODS
Isolation and culture of human articular chondrocytes
This study was approved by the Institutional Review Board of Shenzhen Second People’s Hospital and all experiments were performed in accordance with the relevant guidelines and regulations. Patients received detailed explanation of the study and submitted written informed consents. Human articular cartilage tissues were collected from knee joints of 7 patients who had undergone total knee replacement surgery. The cartilage samples were rinsed with phosphate buffer saline (PBS) plus penicillin and streptomycin (Gibco) for 3 times. The tissues were then cut to about 1 cubic millimeter and placed in 1mg/ml collagenase II (Sigma) for digestion at 37°C for 6 hours with shaking. After filtration and centrifugation, cells were resuspended with DMEM/F12 (Thermo Scientific Hyclone) plus 10% FBS (Gibco) and 1% non-essential amino acid (Sigma). The cells were seeded at the density of 4× 103 cell/cm2 in 6-well plates. Media were refreshed every 3 days and the cells were passaged when the confluence reached 80% or above. TGFβ1 was added to the wells of plate when chondrocytes adhered to the bottom of the plate at different concentrations (2ng/ml, 5ng/ml and 10ng/ml). Cells with the confluence of 80% or above were collected and treated with Trizol reagent (Invitrogen) for total RNA isolation or fixed with 4% paraformaldehyde (PFA) for toluidine blue staining and immunocytochemical staining. Quantitative analysis of network formation was performed with Image J software and the data was presented as the means ± standard deviation from triplicate experiments.
Cytochemistry and immunocytochemistry
The fixed cells were rinsed with PBS for 3 times and stained with 1% toluidine blue for 4h with shaking. The cells were then washed with water and observed. The images were captured using a Leica inverted microscope (Leica DM16000B).
The samples for immunocytochemical staining were first blocked with 10% normal horse serum for 1 hour at room temperature and incubated with primary antibody at 4°C overnight. Primary antibodies used in this study were anti-collagen type I (RD) and anti-collagen type II (Sigma). Secondary antibodies (Invitrogen) were applied on the second day after 3 washes of the samples with PBS. All the samples were mounted with the mounting solution Fluoroshield with DAPI (Sigma).
Quantitative RT-PCR analysis
Total RNA was extracted from cells using Trizol reagent (Invitrogen) according to the manufacturer’s instructions and then was reversely transcribed using the Revert Aid First Strand cDNA Sythesis Kit (Fermentas). Quantitative RT-PCR was conducted with SYBR Green Supermix (BioRad) using aViiA 7™ Real-Time PCR Sytsem (Applied Biosystems). All samples were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Each assay was performed in triplicate. Primers used were listed in Table 4. The fold change in the expression was analyzed using the ΔΔCt method. Student’s t-test was used in independent data for statistical analysis. The differences were considered significant when the P value was<0.05.
Table 4: Primer sequences for quantitative PCR
Primers |
Sequence 5′---- 3′ |
Information |
|---|---|---|
hGAPDH-F |
AAGGTGAAGGTCGGAGTCAA |
GAPDH expression |
hGAPDH-R |
AATGAAGGGGTCATTGATGG |
|
hCol2A1-F |
CTGTCCTTCGGTGTCAGGG |
Col2A1 expression |
hCol2A1-R |
CGGCTTCCACACATCCTTAT |
|
hSMAD3-F |
TCAACACCAAGTGCATCACC |
SMAD3 expression |
hSMAD3-R |
CGGCAGTAGATGACATGAGG |
|
hSMAD7-F |
CCAACTGCAGACTGTCCAGA |
SMAD7 expression |
hSMAD7-R |
CCAGGCTCCAGAAGAAGTTG |
|
hCOL10A1-F |
CATAAAAGGCCCACTACCCA |
COL10A1 expression |
hCOL10A1-R |
GTGGACCAGGAGTACCTTGC |
|
hVEGFA-F |
CAAGACAAGAAAATCCCTGTGG |
VEGF expression |
hVEGFA-R |
GCTTGTCACATCTGCAAGTACG |
|
hESM1-F |
TGTCAGCCTTCTAATGGGGA |
ESM1 expression |
hESM1-R |
ACTGGCAGTTGCAGGTCTCT |
|
hKDR-F |
CCTGTATGGAGGAGGAGGAA |
KDR expression |
hKDR-R |
CGGCTCTTTCGCTTACTGTT |
Western blot analysis
The proteins from different groups of cells were extracted by RIPA buffer (Cell Signaling) and denatured using Laemmli SDS sample buffer. The denatured samples were loaded onto a 12% gel, running at 90 V for 30min and 160V for 90min. The membranes carrying proteins were blocked with 5% non-fat dry milk (2.5g milk powder in 50ml TBST) for 1 hour at room temperature. Primary antibodies diluted in 5% non-fat dry milk (mouse anti-GAPDH (Sigma), 1:2000; goat anti-ESM1 (Santa Cruz), 1:1000) were applied to the membranes separately overnight with shaking at 4°C. Secondary antibodies (anti-mouse HRP, 1:3000, (Invitrogen); anti-goat HRP 1:2000, (Invitrogen)) were applied and incubated for 1 hour at room temperature the second day after washing the membrane with TBST for 3 times. ECL Western Blotting Detection Reagent (Pierce) and UVITEC Cambridge (UK) was used for the development. Densitometric analysis of Western blot was performed using ImageJ software. The integrated density was measured for 3 times and then used for statistical analysis.
Flow cytometric analysis
Indirect immunofluorescence analysis was performed for chondrocytes with anti-ESM1 antibody (Santa Cruz) at 4 °C for 30 min. After two washes with wash buffer (PBS supplemented with 2% FBS), the cells were incubated with the secondary mouse anti-goat IgG-FITC antibody (Invitrogen) for 30 min in the dark. Following two more washes, the cells were resuspended in PBS and analyzed by flow cytometry (Beckman Coulter Navios). Cells incubated with secondary antibody only were used as the negative control.
DNA microarray analysis
Affymetrix (Santa Clara, USA) Gene Chip Human Genome U133 Plus 2.0 Array was used in the study. This Array is widely used for human gene expression profile and allowed analysis of 47,000 transcripts. Samples from 3 patients were sent to CapitalBio Corporation (Beijing, China) for the experiment and the results were analyzed by Affymetrix Microarray Suite (MAS 5.0)[16, 17]. All the experiments of gene expression profiling were authorized to be performed by CapitalBio Corporation (Beijing, China). Briefly, cells treated or untreated (control) with 10ng/ml of TGFβ1 were harvested and total RNA was isolated with Trizol reagent (Invitrogen). Qualified RNA was used to synthesize a double-stranded cDNA that was used for the following biotin-tagged cDNA synthesis. The biotin-tagged cDNA was fragmented to strands with 35-200bp in length and then proceed to the hybridization process with Affymetrix Gene Chip Human Genome U133 Plus 2.0 Array. All the DEGs were analyzed with a free web-based Molecular Annotation System3.0 (MAS 3.0, www.capitalbio.com). Gene Ontology (GO) analysis and pathway analysis using Kyoto Encyclopedia of Genes and Genomes (KEGG) database were performed.
Matrigel assay
Matrigel matrix (BD) was used for the tube formation assay. The matrix was melted at 4°C for at least 2 hours before the experiment and 50μL matrix was added in each well of the 96-well plate on ice. The plate was transferred to the incubator for about 30min to allow the matrix to gel. The cell density was adjusted to 1.2×105 cell/mL and 100μL per well of the single cell suspensions was added into wells with gelled matrix. Each sample was examined in triplicate. The plate was then put into the incubator. Images were captured with an inverted microscope (Leica DMIL LED) 5 hours after the incubation. Angiogenesis inhibitor SFN 1.5uM (Sigma) was added to the control group. Three random areas for each sample were photographed. The total length of the capillary-like structures, the number of junctions and branches, and the total segments length were measured using ImageJ software and the data was presented as the means ± standard deviation from triplicate images.
Author contributions
Jie-Lin Chen and Chang Zou conceived the study and drafted the manuscript; Jie-Lin Chen, Yunfang Chen, Weimin Zhu, Jianghong Huang and Cui Jiaming participated in the experiments and interpreted the data obtained; Wei Liu, Qisong, Daming Wang and Jia Zhaofeng performed bioinformatic analysis; Li Duan and Jianyi Xiong revised the manuscript critically with important intellectual inputs; Daping Wang obtained funding, conceived the study and revised the draft.
ACKNOWLEDGMENTS AND FUNDING
The authors thank Dr. Zhang Shiquan for his help in clinic and Dr. Lai Wen for his valuable comments on this manuscript.
This study was supported by Grants from The National Natural Science Foundation of China (no.81572198, 81672234, no.21602137, 81402370), The Natural Science Foundation of Guangdong Province, China (no.2015A030313772 and 2015A030313829), The Guangdong Science and Technology Project (no.2015A020212001, 2015B090904007 and 2016A050503009) and The Shenzhen Science and Technology Project (project nos. JSGG20151030140325149, JCYJ20140414170821200, JCYJ20160301111338144, JCYJ20160425104312430).
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest to this work.
REFERENCES
1. Man GS, Mologhianu G. Osteoarthritis pathogenesis - a complex process that involves the entire joint. J Med Life. 2014; 7: 37-41.
2. Sofat N, Kuttapitiya A. Future directions for the management of pain in osteoarthritis. Int J Clin Rheumtol. 2014; 9: 197-276.
3. Distler JH, Hirth A, Kurowska-Stolarska M, Gay RE, Gay S, Distler O. Angiogenic and angiostatic factors in the molecular control of angiogenesis. Q J Nucl Med. 2003; 47: 149-161.
4. Carmeliet P, Jain RK. Molecular mechanisms and clinical applications of angiogenesis. Nature. 2011; 473: 298-307.
5. Bonnet CS, Walsh DA. Osteoarthritis, angiogenesis and inflammation. Rheumatology (Oxford). 2005; 44: 7-16.
6. Pesesse L, Sanchez C, Henrotin Y. Osteochondral plate angiogenesis: a new treatment target in osteoarthritis. Joint Bone Spine. 2011; 78: 144-149.
7. Blaney Davidson EN, van der Kraan PM, van den Berg WB. TGF-beta and osteoarthritis. Osteoarthritis Cartilage. 2007; 15: 597-604.
8. Shen J, Li S, Chen D. TGF-beta signaling and the development of osteoarthritis. Bone Res. 2014; 2.
9. Chen JL, Duan L, Zhu W, Xiong J, Wang D. Extracellular matrix production in vitro in cartilage tissue engineering. J Transl Med. 2014; 12: 88.
10. Chadjichristos C, Ghayor C, Herrouin JF, Ala-Kokko L, Suske G, Pujol JP, Galera P. Down-regulation of human type II collagen gene expression by transforming growth factor-beta 1 (TGF-beta 1) in articular chondrocytes involves SP3/SP1 ratio. J Biol Chem. 2002; 277: 43903-43917.
11. Galera P, Redini F, Vivien D, Bonaventure J, Penfornis H, Loyau G, Pujol JP. Effect of transforming growth factor-beta 1 (TGF-beta 1) on matrix synthesis by monolayer cultures of rabbit articular chondrocytes during the dedifferentiation process. Exp Cell Res. 1992; 200: 379-392.
12. Blaney Davidson EN, Vitters EL, van der Kraan PM, van den Berg WB. Expression of transforming growth factor-beta (TGFbeta) and the TGFbeta signalling molecule SMAD-2P in spontaneous and instability-induced osteoarthritis: role in cartilage degradation, chondrogenesis and osteophyte formation. Ann Rheum Dis. 2006; 65: 1414-1421.
13. Zhen G, Wen C, Jia X, Li Y, Crane JL, Mears SC, Askin FB, Frassica FJ, Chang W, Yao J, Carrino JA, Cosgarea A, Artemov D, et al. Inhibition of TGF-beta signaling in mesenchymal stem cells of subchondral bone attenuates osteoarthritis. Nat Med. 2013; 19: 704-712.
14. Verdier MP, Seite S, Guntzer K, Pujol JP, Boumediene K. Immunohistochemical analysis of transforming growth factor beta isoforms and their receptors in human cartilage from normal and osteoarthritic femoral heads. Rheumatol Int. 2005; 25: 118-124.
15. Narcisi R, Quarto R, Ulivi V, Muraglia A, Molfetta L, Giannoni P. TGF beta-1 administration during ex vivo expansion of human articular chondrocytes in a serum-free medium redirects the cell phenotype toward hypertrophy. J Cell Physiol. 2012; 227: 3282-3290.
16. Peng S, Yan L, Zhang J, Wang Z, Tian M, Shen H. An integrated metabonomics and transcriptomics approach to understanding metabolic pathway disturbance induced by perfluorooctanoic acid. J Pharm Biomed Anal. 2013; 86: 56-64.
17. Huang X, Hao C, Shen X, Liu X, Shan Y, Zhang Y, Chen L. Differences in the transcriptional profiles of human cumulus cells isolated from MI, MII oocytes of patients with polycystic ovary syndrome. Reproduction. 2013; 145: 597-608.
18. Mahmoudifar N, Doran PM. Chondrogenesis and cartilage tissue engineering: the longer road to technology development. Trends Biotechnol. 2012; 30: 166-176.
19. Yang YH, Barabino GA. Differential morphology and homogeneity of tissue-engineered cartilage in hydrodynamic cultivation with transient exposure to insulin-like growth factor-1 and transforming growth factor-beta1. Tissue Eng Part A. 2013; 19: 2349-2360.
20. Shi S, Mercer S, Eckert GJ, Trippel SB. Growth factor transgenes interactively regulate articular chondrocytes. J Cell Biochem. 2013; 114: 908-919.
21. Li TF, Chen D, Wu Q, Chen M, Sheu TJ, Schwarz EM, Drissi H, Zuscik M, O’Keefe RJ. Transforming growth factor-beta stimulates cyclin D1 expression through activation of beta-catenin signaling in chondrocytes. J Biol Chem. 2006; 281: 21296-21304.
22. van Beuningen HM, Glansbeek HL, van der Kraan PM, van den Berg WB. Differential effects of local application of BMP-2 or TGF-beta 1 on both articular cartilage composition and osteophyte formation. Osteoarthritis Cartilage. 1998; 6: 306-317.
23. Scharstuhl A, Glansbeek HL, van Beuningen HM, Vitters EL, van der Kraan PM, van den Berg WB. Inhibition of endogenous TGF-beta during experimental osteoarthritis prevents osteophyte formation and impairs cartilage repair. J Immunol. 2002; 169: 507-514.
24. van Beuningen HM, Glansbeek HL, van der Kraan PM, van den Berg WB. Osteoarthritis-like changes in the murine knee joint resulting from intra-articular transforming growth factor-beta injections. Osteoarthritis Cartilage. 2000; 8: 25-33.
25. Tang R, Gao M, Wu M, Liu H, Zhang X, Liu B. High glucose mediates endothelial-to-chondrocyte transition in human aortic endothelial cells. Cardiovasc Diabetol. 2012; 11: 113.
26. Barzilay R, Melamed E, Offen D. Introducing transcription factors to multipotent mesenchymal stem cells: making transdifferentiation possible. Stem cells. 2009; 27: 2509-2515.
27. Sanchez C, Deberg MA, Bellahcene A, Castronovo V, Msika P, Delcour JP, Crielaard JM, Henrotin YE. Phenotypic characterization of osteoblasts from the sclerotic zones of osteoarthritic subchondral bone. Arthritis Rheum. 2008; 58: 442-455.
28. Pesesse L, Sanchez C, Delcour JP, Bellahcene A, Baudouin C, Msika P, Henrotin Y. Consequences of chondrocyte hypertrophy on osteoarthritic cartilage: potential effect on angiogenesis. Osteoarthritis Cartilage. 2013; 21: 1913-1923.
29. Johnston J, Ramos-Valdes Y, Stanton LA, Ladhani S, Beier F, Dimattia GE. Human stanniocalcin-1 or -2 expressed in mice reduces bone size and severely inhibits cranial intramembranous bone growth. Transgenic Res. 2010; 19: 1017-1039.
30. Yoshiko Y, Aubin JE, Maeda N. Stanniocalcin 1 (STC1) protein and mRNA are developmentally regulated during embryonic mouse osteogenesis: the potential of stc1 as an autocrine/paracrine factor for osteoblast development and bone formation. J Histochem Cytochem. 2002; 50: 483-492.
31. Ishikura K, Misu H, Kumazaki M, Takayama H, Matsuzawa-Nagata N, Tajima N, Chikamoto K, Lan F, Ando H, Ota T, Sakurai M, Takeshita Y, Kato K, et al. Selenoprotein P as a diabetes-associated hepatokine that impairs angiogenesis by inducing VEGF resistance in vascular endothelial cells. Diabetologia. 2014; 57: 1968-1976.
32. Patel-Hett S, D’Amore PA. Signal transduction in vasculogenesis and developmental angiogenesis. Int J Dev Biol. 2011; 55: 353-363.
33. Murata M, Yudoh K, Masuko K. The potential role of vascular endothelial growth factor (VEGF) in cartilage: how the angiogenic factor could be involved in the pathogenesis of osteoarthritis? Osteoarthritis Cartilage. 2008; 16: 279-286.
34. Nakagawa T, Lee SY, Reddi AH. Induction of chondrogenesis from human embryonic stem cells without embryoid body formation by bone morphogenetic protein 7 and transforming growth factor beta1. Arthritis Rheum. 2009; 60: 3686-3692.
35. Steinert AF, Palmer GD, Pilapil C, Noth U, Evans CH, Ghivizzani SC. Enhanced in vitro chondrogenesis of primary mesenchymal stem cells by combined gene transfer. Tissue Eng Part A. 2009; 15: 1127-1139.