
INTRODUCTION
Coronary heart disease (CHD) and ischemic stroke (IS) are the most prevalent geriatric diseases and the major determinant of mortality and morbidity worldwide [1-3]. More than 700,000 people die from CHD each year in China [4]. As a complex and multifactorial disorder, CHD and IS are resulted from lots of pathogenic factors, including genetic factors and environmental exposures [5]. The major pathological basis of these two diseases had proved to be atherosclerosis which the essential as an ambitious inflammatory disorder. Therefore, both of diseases would be participated in the same genetic and environmental backgrounds, including gender, time to life, hypercholesterol, hypertension, diabetes, cigarette smoking, and genetic factors [6-9]. A large number of genes and loci related to CHD [10] or IS [11] were reported in previous genome-wide association studies (GWASes). In addition, some genetic variants that were initially shown to have an effect on the risk of CHD were detected to be related to IS soon afterwards [12, 13].
Recent GWASes have found several novel loci at chromosome 12q24, including the mevalonate kinase (MVK) and methylmalonic aciduria (cobalamin deficiency) cblB type (MMAB) genes, both of which influence high-density lipoprotein cholesterol (HDL-C) levels [14, 15]. According to the relevant studies that this region can regulate serum lipid concentrations. [16-18]. Particularly, two head to head genes, MVK and MMAB took part in metabolic pathways may adjust to HDL metabolism [19]. MVK, encoded by MVK, play an important role in an initial stage in cholesterol biosynthesis. In humans, when MVK mutations in homozygosity can give rise to hyperimmunoglobulinemia D syndrome, which the basic symptoms were fever and high concentrations of immunoglobulins D and A in blood. When the patients suffered from hyperimmunoglobulinemia D syndrome, low HDL-C levels can be found, in accordance with the latest GWASes findings [14, 15]. However, when somebody lacked of cob (I) alamin adenosyltransferase, as an enzyme encoded by MMAB, may contribute to methylmalonic aciduria [20]. But, the exact metabolism by which MMAB influences cholesterol is still unknown. A relevant report about schizophrenia had revealed that urinary methylmalonic acid may negatively correlated with red blood cell membrane cholesterol levels in blood [21]. MVK, which is close to MMAB, gives rise to the increased sensibility of obesity, diabetes and atherosclerosis [22]. Therefore, MVK and MMAB may be appropriate candidates as genes to elevate HDL-C concentrations and then affect the risk of CHD and IS. Sun et al. had taken several SNPs into consideration, including MVK (rs3759387 and rs2287218) and MMAB (rs12817689, rs22411201, rs11067227, rs7134594, rs877710, rs11067233, rs9593, rs11831226 and rs8228), but only rs11067233 in MMAB may contribute to the susceptibility of CHD by decreasing plasma HDL-C levels in Han Chinese [23]. Junyent et al. suggested that the MMAB-3U3527G/C variant might contribute to the variation in HDL-C concentrations, particularly in subjects with high carbohydrate intakes [24]. In the current study, we aimed to detected whether four SNPs, rs3759387, rs7134594, rs877710 and rs9593 in the MVK and MMAB and their mutual effect between gene and environment, make an interaction with the risk of CHD and IS in Han populations where located in southern China.
RESULTS
General features of the demography
The general trait of the demography is summarized in Table 1. The levels of systolic blood pressure (SBP), pulse pressure, apolipoprotein (Apo) B, and the percentages of hypertension were higher but the concentrations of total cholesterol (TC), HDL-C, low-density lipoprotein cholesterol (LDL-C), the percentages of individuals who consumed alcohol and the ApoA1 to ApoB ratio were lower in persons who suffered from CHD than in normals (P < 0.05 for all). The same situation was also found in IS subjects compared to normals (P < 0.05 for all).
Table 1: Comparison of general characteristics and serum lipid levels between controls and patients
Parameter |
Control |
CHD |
IS |
P1 |
P2 |
|---|---|---|---|---|---|
Number |
847 |
846 |
869 |
- |
- |
Male/female |
607/240 |
625/221 |
628/241 |
0.224 |
0.267 |
Age, years1 |
61.71±11.82 |
62.18±10.58 |
62.72±12.35 |
0.423 |
0.676 |
Body mass index, kg/m2 |
22.58±3.16 |
23.89±3.27 |
23.21±3.39 |
0.384 |
0.221 |
Systolic blood pressure, mmHg |
127.96±19.96 |
133.40±23.27 |
147.05±21.16 |
0.000 |
0.013 |
Diastolic blood pressure, mmHg |
81.86±13.83 |
79.45±14.33 |
83.51±13.05 |
0.073 |
0.065 |
Pulse pressure, mmHg |
47.48±19.29 |
57.03±17.36 |
63.54±17.71 |
0.000 |
0.000 |
Cigarette smoking, n (%)2 |
330 (39.0) |
396 (46.8) |
379 (43.6) |
0.205 |
0.454 |
Alcohol consumption, n (%) |
354 (41.8) |
225 (27.0) |
262 (30.1) |
0.000 |
0.000 |
Total cholesterol, mmol/L |
4.91±0.99 |
4.54±1.26 |
4.51±1.12 |
0.000 |
0.001 |
Triglyceride, mmol/L3 |
1.36 (0.73) |
1.38 (0.95) |
1.47 (0.91) |
0.642 |
0.155 |
HDL-C, mmol/L |
1.87±0.48 |
1.16±0.39 |
1.23±0.38 |
0.000 |
0.000 |
LDL-C, mmol/L |
2.75±0.78 |
2.71±1.04 |
2.67±0.91 |
0.000 |
0.000 |
Apolipoprotein (Apo) A1, g/L |
1.40±0.26 |
1.05±0.60 |
1.01±0.22 |
0.070 |
0.061 |
ApoB, g/L |
0.90±0.20 |
0.91±0.27 |
0.92±0.25 |
0.000 |
0.000 |
ApoA1/ApoB |
1.63±0.53 |
1.36±2.48 |
1.26±0.65 |
0.019 |
0.000 |
Diabetes mellitus, n (%) |
153 (18.1) |
137 (16.2) |
159 (18.3) |
0.324 |
0.869 |
Hypertension, n (%) |
310 (36.6) |
354 (41.8) |
585 (67.3) |
0.000 |
0.000 |
CHD, coronary heart disease; IS, ischemic stroke; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol. 1 Mean ± SD determined by t-test. 2The rate or constituent ratio between the two groups was analyzed by the chi-square test. 3Median (interquartile range) tested by the Wilcoxon-Mann-Whitney test. P1, CHD vs. controls; P2, IS vs. controls.
Frequency of genotypes and alleles
The frequency of genotypes and alleles of the four MVK-MMAB SNPs is showed in Figure 1. Four of the detected SNPs were conformed to Hardy-Weinberg equilibrium (P > 0.05). The frequency of genotypes and alleles of rs3759387 and rs7134594, but not rs877710 and rs9593 SNPs, was distinguished normals from patients (CHD and IS, P < 0.05 for all). The rs3759387A allele and rs3759387AA genotype frequencies were higher in CHD (A, 14.70%; AA, 1.89%) and IS (A, 13.98%; AA, 1.73%) patients than in control subjects (A, 11.20%; AA, 0.95%; P < 0.01 for all). The rs7134594T allele and rs7134594CT genotype frequencies were higher in CHD (T, 42.96%; CT, 50.23%) and IS (T, 41.60%; CT, 47.3%) patients than in control subjects (T, 41.14%; CT, 45.45%; P < 0.05 for all).
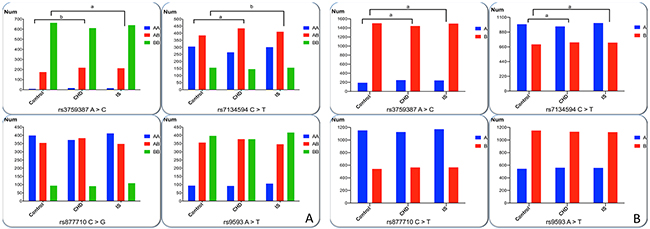
Figure 1: Genotypic and allelic frequencies of four SNPs in controls and patients. (A) AA genotypes represent as rs3759387AA, rs7134594CC, rs877710CC and rs9593AA. AB genotypes represent as rs3759387AC, rs7134594CT, rs877710CG and s9593AT. BB genotypes represent rs3759387CC, rs7134594TT, rs877710GG and rs9593TT. (B) The A allele represents rs3759387A, rs7134594C, rs877710C and rs9593A. The B allele represents rs3759387C, rs7134594T, rs877710G and rs9593T. All of the detected SNPs were in Hardy-Weinberg equilibrium (P > 0.05). The rs3759387 A >C and rs7134594 C > T allelic and genotypic frequencies were different among the three groups. aP< 0.0125; bP < 0.001.
Genotypes and the risk of diseases
As presented in Table 2, in different genetic models, after Bonferroni correction only genotypes of rs3759387 were associated with the risk of CHD (P < 0.0125 for 0.05 adjusted for 4 variables was considered statistically significant). The dominant model: AA/AC vs. CC (OR = 1.44, 95% CI = 1.14–1.82, P = 0.0023) and log-additive model: A vs. C (OR = 1.42, 95% CI = 1.15–1.77, P = 0.0042) can be considered as a statistically meaning. Similarly, dominant model: AA/AC vs. CC (OR = 1.32, 95% CI = 1.04–1.69, P = 0.0018) and log-additive model: A vs. C (OR = 1.29, 95% CI = 1.03–1.61, P = 0.0025) in genotypes of the rs3759387 SNP were also increased the risk of IS.
Table 2: Genotypes of the four MVK-MMAB SNPs and the risk of CHD and IS
SNP/Model |
Ref. Genotype |
Effect Genotype |
CHD (OR 95% CI) |
P |
IS (OR 95% CI) |
P |
|---|---|---|---|---|---|---|
rs3759387 |
||||||
Codominant |
CC |
AC |
1.40 (1.10-1.78) |
0.053 |
1.32 (1.03-1.70) |
0.077 |
AA |
2.30 (0.95-5.58) |
1.30 (0.51-3.30) |
||||
Dominant |
CC |
AC/AA |
1.44 (1.14-1.82) |
0.0023 |
1.32 (1.04-1.69) |
0.0018 |
Recessive |
CC/AC |
AA |
2.12 (0.88-5.14) |
0.087 |
1.22 (0.48-3.08) |
0.68 |
Overdominant |
CC/AA |
AC |
1.38 (1.08-1.75) |
0.056 |
1.32 (1.03-1.69) |
0.028 |
Log-additive |
1.42 (1.15-1.77) |
0.0042 |
1.29 (1.03-1.61) |
0.0025 |
||
rs7134594 |
||||||
Codominant |
CC |
CT |
1.32 (1.05-1.65) |
0.051 |
1.05 (0.83-1.32) |
0.68 |
TT |
1.10 (0.83-1.49) |
0.92 (0.68-1.24) |
||||
Dominant |
CC |
CT/ TT |
1.26 (1.02-1.56) |
0.034 |
0.81 (1.01-1.26) |
0.92 |
Recessive |
CC/ CT |
TT |
0.94 (0.72-1.22) |
0.64 |
0.90 (0.69-1.17) |
0.43 |
Overdominant |
CC/ TT |
CT |
1.27 (1.04-1.55) |
0.019 |
1.08 (0.88-1.32) |
0.48 |
Log-additive |
1.09 (0.94-1.26) |
0.24 |
0.97 (0.84-1.13) |
0.72 |
||
rs877710 |
||||||
Codominant |
CC |
CG |
1.17 (0.95-1.45) |
0.33 |
0.90 (0.72-1.12) |
0.59 |
GG |
1.02 (0.73-1.44) |
1.02 (0.73-1.43) |
||||
Dominant |
CC |
CG/GG |
1.14 (0.93-1.39) |
0.21 |
0.93 (0.75-1.14) |
0.46 |
Recessive |
CC/CG |
GG |
0.95 (0.69-1.31) |
0.74 |
1.07 (0.78-1.48) |
0.66 |
Overdominant |
CC/GG |
CG |
1.16 (0.95-1.43) |
0.14 |
0.90 (0.73-1.11) |
0.30 |
Log-additive |
1.06 (0.91-1.24) |
0.43 |
0.98 (0.84-1.14) |
0.75 |
||
rs9593 |
||||||
Codominant |
TT |
AT |
1.13 (0.91-1.39) |
0.54 |
0.88 (0.70-1.09) |
0.48 |
AA |
1.02 (0.73-1.43) |
0.98 (0.71-1.37) |
||||
Dominant |
TT |
AT/AA |
1.00 (0.75-1.35) |
0.98 |
0.90 (0.73-1.11) |
0.65 |
Recessive |
TT/AT |
AA |
0.96 (0.70-1.33) |
0.82 |
1.04 (0.76-1.44) |
0.79 |
Overdominant |
TT/AA |
AT |
1.04 (0.77-1.40) |
0.82 |
0.88 (0.71-1.08) |
0.23 |
Log-additive |
0.98 (0.74-1.29) |
0.86 |
0.95 (0.82-1.11) |
0.54 |
SNP, single nucleotide polymorphism; CHD, coronary heart disease; IS, ischemic stroke.
Interactions of the rs3759387 SNP and environmental exposures on the risk of diseases
As presented in Figure 2, the rs3759387 AA/AC genotypes interacted with males resulted in increasing the risk of CHD (OR = 2.08, 95% CI = 1.55–2.77, P < 0.001) and alcohol drinking to reduce the risk of CHD (OR = 0.66, 95% CI = 0.38–1.03, P < 0.001). The rs3759387 AA/AC genotypes also interacted with males (OR = 1.76, 95% CI = 1.31–2.38) and age > 75 years to increase the risk of IS (OR = 4.99, 95% CI = 1.39–8.91, P < 0.001).
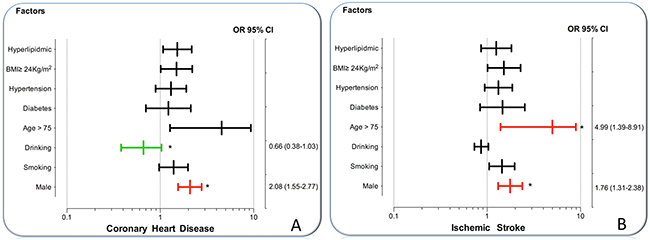
Figure 2: The interactions of the MVK rs3759387 SNP and drinking, smoking, BMI, age, hypertension, diabetes, hyperlipidemia and sex on the risk of CHD and IS. (A) The rs3759387 AA/AC genotypes interacted with male to increase the risk of CHD (OR = 2.08, 95% CI = 1.55 –2.77) and alcohol consumption to decrease the risk of CHD (OR = 0.66, 95% CI = 0.38–1.03). (B) The rs3759387 AA/AC genotypes interacted with male to increase the risk of IS (OR = 1.76, 95% CI = 1.31–2.38) and age > 75 years to increase the risk of IS (OR = 4.99, 95% CI = 1.39–8.91). *P < 0.001.
Haplotypes and the risk of diseases
Strong LD can be found among the rs3759387, rs7134594, rs877710 and rs9593 SNPs in normals and patients (D’ = 0.92–0.99) (Supplementary Figure 1). Besides, haplotype analyses which combined with the four SNPs, and the associations of their different haplotypes and the risk of diseases had also been carried out. The four main haplotypes are presented in Table 3. The A-T-G-A and C-T-C-T haplotypes were associated with an increased risk of CHD (adjusted OR = 1.43, 95% CI = 1.14–1.81, P = 0.023 and OR = 1.30, 95% CI = 1.01–1.68, P = 0.045, respectively). The A-T-G-A haplotype was associated with an increased risk of IS (adjusted OR = 1.28, 95% CI = 1.01–1.63, P = 0.041), but the C-T-G-A haplotype was associated with a decreased risk of IS (adjusted OR = 0.83, 95% CI = 0.69–0.99, P = 0.043).
Table 3: Haplotype frequencies of the four MVK-MMAB SNPs and the risk of CHD and IS
Haplotype |
Control Frequency |
CHD |
IS |
||||
|---|---|---|---|---|---|---|---|
Frequency |
OR (95% CI) |
P |
Frequency |
OR (95% CI) |
P |
||
C-C-C-T |
0.5840 |
0.5565 |
1.00 |
1.00 |
0.5822 |
1.00 |
1.00 |
C-T-G-A |
0.2103 |
0.1845 |
0.92 (0.77 - 1.11) |
0.39 |
0.1835 |
0.83 (0.69 – 0.99) |
0.043 |
A-T-G-A |
0.1045 |
0.1353 |
1.43 (1.14 - 1.81) |
0.023 |
0.1835 |
1.28 (1.01 - 1.63) |
0.041 |
C-T-C-T |
0.0980 |
0.1051 |
1.30 (1.01 - 1.68) |
0.045 |
0.0915 |
0.99 (0.760 - 1.29) |
0.96 |
CHD, coronary heart disease; IS, ischemic stroke. The haplotypes consist of four alleles in the order of rs3759387, rs7134594, rs877710 and rs9593 SNPs.
The mutual effect between the haplotypes and environmental exposures on the risk of diseases
As shown in Figure 3, the interactions of the MVK-MMAB haplotypes and several risk factors for CHD and IS were noted in this study. The interactions of rs3759387A-rs7134594T-rs877710G-rs9593A-smoking (adjusted OR = 1.87, 95% CI = 1.21–2.90, P = 0.00022), rs3759387A-rs7134594T-rs877710G-rs9593A-age > 75 years (OR = 5.56, 95% CI = 2.12–13.72, P < 0.001), rs3759387C-rs7134594T-rs877710C-rs9593T-smoking (OR = 2.32, 95% CI = 1.42–3.79, P = 0.00002), and rs3759387C-rs7134594T-rs877710C-rs9593T-age > 75 years (OR = 2.18, 95% CI = 1.44–5.25, P = 0.0016) increased the risk of CHD.
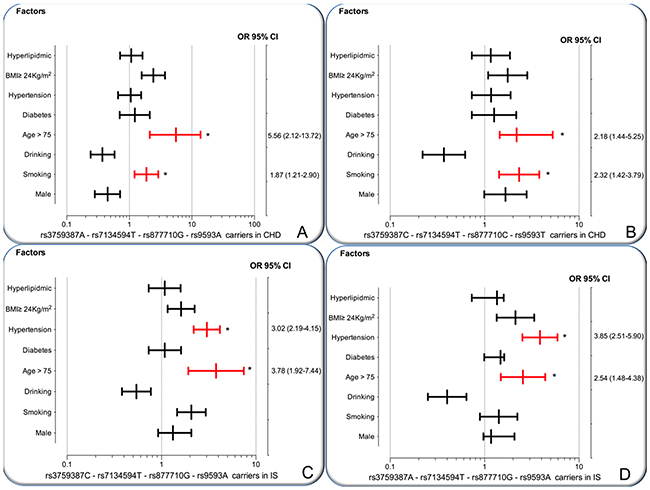
Figure 3: The interactions of the MVK-MMAB haplotype and the risk of CHD and IS. CHD, coronary heart disease; IS, ischemic stroke. (A and B) The rs3759387A-rs7134594T-rs877710G-rs9593A carriers and rs3759387C-rs7134594T-rs877710C-rs9593T carriers interacted with cigarette smoking and age > 75 years to increase the risk of CHD. (C and D) The rs3759387C-rs7134594T-rs877710G-rs9593A carriers and rs3759387A-rs7134594-rs877710G-rs9593A carriers interacted with hypertension and age > 75 years to increase the risk of IS. *P < 0.001.
The interactions of rs3759387C-rs7134594T-rs877710G-rs9593A-hypertension (adjusted OR = 3.02, 95% CI = 2.19–4.15, P = 0.00012), rs3759387C-rs7134594T-rs877710G-rs9593A-age > 75 years (OR = 3.78, 95% CI = 1.92–7.44, P = 0.0009), rs3759387A-rs7134594T-rs877710G-rs9593A-hypertension (OR = 3.85, 95% CI = 2.51–5.90, P = 0.0001), and rs3759387A-rs7134594T-rs877710G-rs9593A-age > 75 years (OR = 2.54, 95% CI = 1.48–4.38, P < 0.001) increased the risk of IS.
Genotypes and serum lipid concentrations
The association of the MVK-MMAB SNPs and serum lipid concentrations in normals is shown in Figure 4. There were distinct serum HDL-C concentrations among the three genotypes of the rs3759387 (P = 0.008); however, no positive findings were observed among the other three SNPs (P > 0.0125 for all). The subjects with the rs3759387AA genotype had lower HDL-C concentrations than did the individuals with rs3759387AC and rs3759387CC genotypes. There were no differences in serum TC, triglycerides (TG), LDL-C, ApoA1, ApoB concentrations, and the ratio of ApoA1 to ApoB among the three genotypes of the SNPs.
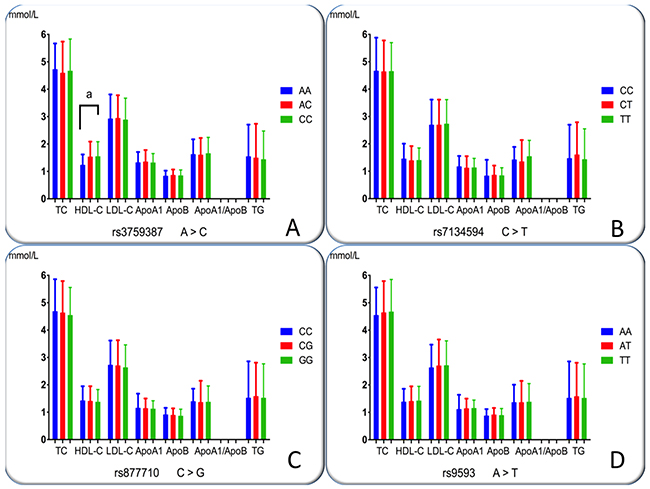
Figure 4: Genotypes of the four MVK-MMAB SNPs and serum lipid levels in controls. TC, total cholesterol; TG, triglyceride; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; ApoA1, apolipoprotein A1; ApoB, apolipoprotein B; ApoA1/ApoB, the ratio of apolipoprotein A1 to apolipoprotein B. The value of triglyceride is presented as the median (interquartile range), and the difference among the genotypes was determined by the Kruskal-Wallis test. aP< 0.0125 (after adjusting for 4 independent tests by the Bonferroni correction). (A), rs3759387; (B), rs7134594; (C), rs877710; and (D), rs9593 SNPs.
The interactions between the MVK-MMAB SNPs and several environmental exposures on serum lipid concentrations and the risk of diseases
The interactions between the MVK-MMAB SNPs and several environmental exposures, including alcohol consumption, smoking, BMI, age, and sex, on serum lipid levels and the risk of CHD and IS are revealed in Table 4. The mutual effect between some SNPs and alcohol consumption to add serum HDL-C (rs3759387, Figure 5B; rs877710, Figure 5H; and rs9593, Figure 5I) and ApoA1 (rs3759387, Figure 5C; rs7134594 Figure 5G) levels. The mutual effect between rs3759387 (Figure 5A) and rs7134594 (Figure 5F) and cigarette smoking to add serum HDL-C concentrations. The mutual effect between rs7134594 and BMI ≥ 24 kg/m2 to reduce serum HDL-C (Figure 5E) concentrations. The mutual effect between rs3759387 and BMI ≥ 24 kg/m2 to reduce serum ApoA1 (Figure 5D) concentrations.
Table 4: The PI values for the interactions of genotypes and drinking, smoking, and BMI on serum lipid levels and the risk of CHD and IS
SNP/Factor |
Lipid |
CHD |
IS |
||||||
|---|---|---|---|---|---|---|---|---|---|
TC |
TG |
HDL-C |
LDL-C |
ApoA1 |
ApoB |
ApoA1/ApoB |
|||
rs3759387 |
|||||||||
Smoking |
0.126 |
0.447 |
0.000 |
0.035 |
0.029 |
0.131 |
0.878 |
0.616 |
0.298 |
Drinking |
0.003 |
0.380 |
0.000 |
0.749 |
0.000 |
0.016 |
0.728 |
0.433 |
0.193 |
BMI |
0.005 |
0.293 |
0.000 |
0.004 |
0.003 |
0.046 |
0.044 |
0.276 |
0.888 |
Age |
0.007 |
0.013 |
0.016 |
0.066 |
0.005 |
0.016 |
0.340 |
0.245 |
0.960 |
Sex |
0.011 |
0.003 |
0.022 |
0.072 |
0.004 |
0.003 |
0.198 |
0.456 |
0.267 |
rs7134594 |
|||||||||
Smoking |
0.994 |
0.381 |
0.000 |
0.176 |
0.670 |
0.381 |
0.004 |
0.450 |
0.785 |
Drinking |
0.229 |
0.239 |
0.000 |
0.299 |
0.001 |
0.239 |
0.909 |
0.507 |
0.504 |
BMI |
0.040 |
0.004 |
0.001 |
0.003 |
0.727 |
0.059 |
0.081 |
0.587 |
0.059 |
Age |
0.785 |
0.590 |
0.883 |
0.702 |
0.375 |
0.727 |
0.112 |
0.329 |
0.976 |
Sex |
0.241 |
0.504 |
0.694 |
0.039 |
0.290 |
0.241 |
0.198 |
0.956 |
0.096 |
rs877710 |
|||||||||
Smoking |
0.727 |
0.375 |
0.991 |
0.035 |
0.072 |
0.975 |
0.909 |
0.507 |
0.504 |
Drinking |
0.700 |
0.618 |
0.001 |
0.383 |
0.404 |
0.639 |
0.355 |
0.906 |
0.864 |
BMI |
0.264 |
0.198 |
0.438 |
0.702 |
0.135 |
0.207 |
0.507 |
0.253 |
0.743 |
Age |
0.869 |
0.459 |
0.081 |
0.262 |
0.628 |
0.308 |
0.016 |
0.507 |
0.681 |
Sex |
0.131 |
0.811 |
0.142 |
0.653 |
0.016 |
0.011 |
0.431 |
0.842 |
0.502 |
rs9593 |
|||||||||
Smoking |
0.709 |
0.492 |
0.441 |
0.081 |
0.262 |
0.507 |
0.253 |
0.239 |
0.371 |
Drinking |
0.404 |
0.633 |
0.001 |
0.112 |
0.702 |
0.375 |
0.727 |
0.004 |
0.991 |
BMI |
0.135 |
0.753 |
0.264 |
0.198 |
0.438 |
0.089 |
0.752 |
0.590 |
0.672 |
Age |
0.628 |
0.782 |
0.066 |
0.348 |
0.653 |
0.565 |
0.290 |
0.565 |
0.587 |
Sex |
0.354 |
0.851 |
0.445 |
0.195 |
0.842 |
0.032 |
0.792 |
0.693 |
0.814 |
SNP, single nucleotide polymorphism; TC, total cholesterol; TG, triglyceride; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; ApoA1, apolipoprotein A1; ApoB, apolipoprotein B; CHD, coronary heart disease; IS, ischemic stroke; BMI, body mass index. PI ≤ 0.00125 was considered statistically significant after Bonferroni correction.
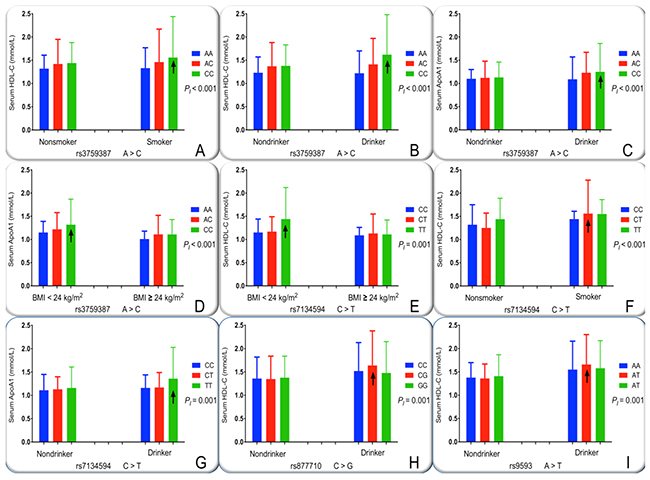
Figure 5: The interactions of the MVK-MMAB SNPs and drinking, smoking and BMI on serum lipid levels. HDL-C, high-density lipoprotein cholesterol; ApoA1, apolipoprotein A1; BMI, body mass index. The differences in serum HDL-C and ApoA1 levels among the genotypes were assessed using analysis of covariance. The the interactions of the genotypes and alcohol consumption, cigarette smoking or BMI ≥ 24 kg/m2 on serum lipid levels were detected by using a factorial regression analysis after controlling for potential confounders (PI). ↑, genotype and alcohol consumption or cigarette smoking or BMI ≥ 24 kg/m2 interaction increased serum lipid levels. PI ≤ 0.00125 was considered statistically significant after Bonferroni correction (corresponding to P < 0.05 after adjusting for five environment exposures multiply eight outcomes by the Bonferroni correction). (A), rs3759387-smoking on HDL-C; (B), rs3759387-drinking on HDL-C; (C), rs3759387-drinking on ApoA1; (D), rs3759387-BMI on ApoA1; (E), rs7134594-BMI on HDL-C; (F), rs7134594-smoking on HDL-C; (G), rs7134594-drinking on ApoA1; (H), rs877710-drinking on HDL-C; and (I), rs9593-drinking on HDL-C.
Relative factors for serum lipid parameters
As shown in Figure 6, Pearson correlation analysis showed that several environmental exposures just as time to life, sex, cigarette smoking, and alcohol consumption and traditional cardiovascular risk factors such as BMI and blood pressure levels, also correlated with serum lipid phenotypes of the patient groups.
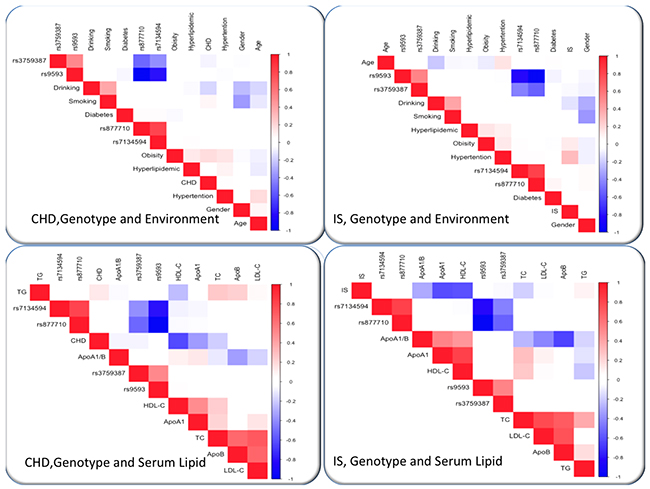
Figure 6: Correlation among environmental exposures and serum lipid variables, as well as the candidate loci in CHD or IS. CHD, coronary heart disease; IS, ischemic stroke. TC, total cholesterol; TG, triglyceride; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; ApoA1, apolipoprotein A1; ApoB, apolipoprotein B; ApoA1/B, the ratio of apolipoprotein A1 to apolipoprotein B; BMI, body mass index.
DISCUSSION
In the current study, we showed distinctions between the frequency of genotypes and alleles of the rs3759387 and rs7134594 SNPs between normals and CHD or IS patients. Only the rs3759387 genotypes or alleles were associated with the disease risk. In different genetic model, the mutual effect between dominant genotypes and males to add the risk of CHD, but they interacted with alcohol drinking to reduce the risk of CHD. Similarly, the dominant genotypes interacted with males and age > 75 years to increase the risk of IS. The SNPs of rs3759387, rs7134594, rs877710 and rs9593 were in strong LD in normals and patients. Four main haplotypes were detected among the four SNPs. The haplotypes of A-T-G-A and C-T-C-T were associated with an added risk of CHD, but the haplotypes of C-T-G-A and A-T-G-A were associated with an added risk of IS. The individuals with the rs3759387AA genotype in normals had lower HDL-C concentrations than did the individuals with rs3759387AC/CC genotypes. The mutual effect between several SNPs and alcohol drinking to affect serum HDL-C (rs3759387, rs877710 and rs9593) and ApoA1 (rs3759387 and rs7134594) levels. The mutual effect between rs3759387 and rs7134594 and cigarette smoking to influence serum HDL-C concentrations. Two SNPs interacted with BMI ≥ 24 kg/m2 to decrease serum HDL-C (rs7134594) and ApoA1 (rs3759387) levels. The interactions of haplotypes and several risk factors on CHD and IS were also observed. Several environmental exposures and traditional cardiovascular risk factors were also correlated with serum lipid phenotypes in both patient groups. As far as we had known that this is the first report to detect the interactions among the four MVK-MMAB SNPs and their haplotypes and environmental exposures on serum lipid concentrations and the risk of CHD and IS.
We revealed that the frequency of rs3759387 and rs7134594 genotypes and alleles were distinct from normals and CHD or IS patients. The patients with CHD or IS had higher frequencies of the rs3759387A allele than did the controls. At the same time, the patients with CHD or IS had higher frequencies of the rs7134594T allele than did the controls. The rs3759387 SNP was also associated with the risk of CHD and IS after Bonferroni correction in different genetic models and the mutual effect between dominant genotypes and environmental exposures to increase the risk of CHD and IS.
In a previous GWAS, the information in the International HapMap Project’s database showed that the rs3759387A allelic frequency was 23.3% in Europeans, 17.8% in Han Chinese in Beijing (CHB), 9.9% in Japanese, and 45.1% in Sub-Saharan African. Besides, we also discovered that the rs3759387A allelic frequency in our current research populations was lower than in CHB in comparison with the other populations, a reasonable explanation would be contributed to different sample numbers and because of CHB in Guangxi are a part of Han. Above results remind us that the prevalence of the rs3759387A allele variation may have racial/ethnic specificity. The prevalence of the rs3759387A allele is higher in Europeans than in Chinese. All of these findings would be a reasonable explanation for the distinct prevalence of CHD between European and Chinese.
The association of the rs3759387, rs7134594, rs877710 and rs9593 SNPs and the risk of CHD and IS has never been detected in previous studies. In the current study, we revealed that the rs3759387 SNP may have an association with the three SNPs and influence the risk of CHD and IS. Furthermore, high LD among the four SNPs was also found in controls and patients. Moreover, when we analyzed the haplotypes among the four SNPs, the haplotypes of A-T-G-A and C-T-C-T were associated with an increased risk of CHD, whereas the haplotypes of C-T-G-A and A-T-G-A were associated with an increased risk of IS. However, these results require further investigation in the different populations with bigger sample numbers.
Previous epidemiological researches have given strong evidence that high serum HDL-C concentrations would be inversed CHD morbidity and mortality [25-27]. Each 1% decrease in LDL-C concentration has been estimated to decrease the risk of CHD by 1% [28], and each 1% increase in HDL-C level decreases the risk of CHD by 2% [29]. The exact mechanism why HDL-C can protection against atherosclerosis includes the below three main points: (1), reverse cholesterol transport from peripheral tissues to the liver [30]; (2), inhibition of LDL-C oxidation; and (3), stabilization of the production of prostacyclin [31]. In humans, MVK and MMAB are head-to-head orientation and located on chromosome 12. In addition, MVK and MMAB, which share the same promoter, are both modified by sterol-responsive element-binding protein 2 (SREBP2), which is a transcription factor that controls cholesterol homeostasis. What is more, the way in which these two neighboring genes take part in metabolic pathways may have an effect on HDL-C metabolism had been found. MVK encodes MVK, play an important role in an initial stage in cholesterol biosynthesis (Figure 7) [32]. In contrast, when lacked of cob (I) alamin adenosyltransferase, as an enzyme encoded by MMAB, someone may result to methylmalonic aciduria. The exact reason for MMAB adjusted cholesterol metabolism has not been identified, maybe cholesterol synthesis through SREBP2 can explain our findings [33]. The precise mechanism of MVK and MMAB on serum lipid metabolism remains to be investigated, which may provide a promising target for medical therapy. Similarly, little is known about the potential mutual effect between the MVK-MMAB SNPs and their haplotypes with environmental exposures on serum lipid levels and the risk of CHD and IS. According to the current study, we first reported the mutual effect between some SNPs and alcohol consumption to add serum HDL-C (rs3759387, rs877710 and rs9593) and ApoA1 (rs3759387 and rs7134594) levels. The mutual effect between rs3759387 and rs7134594 and cigarette smoking to add serum HDL-C levels. The mutual effect between rs3759387 and rs7134594 and alcohol consumption to add serum ApoA1 levels. The mutual effect between rs7134594 and BMI ≥ 24 kg/m2 to reduce serum HDL-C concentrations.
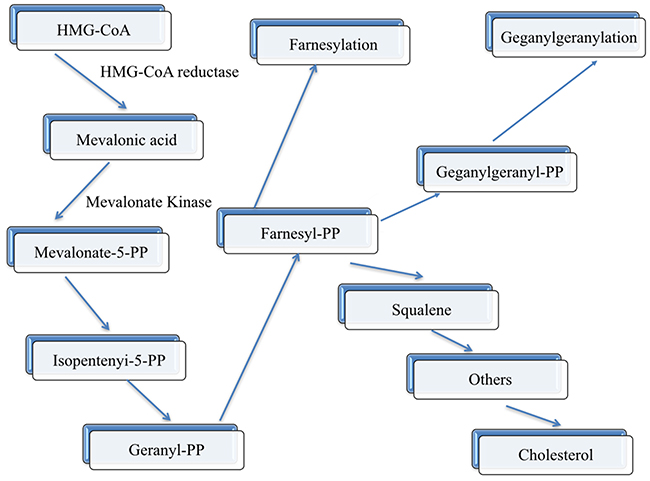
Figure 7: Schematic representation of the mevalonate pathway. The enzymes (HMG-CoA reductase and mevalonate kinase) are indicated along the pathway in bold characters.
The rs3759387A-rs7134594T-rs877710G-rs9593A and rs3759387C-rs7134594T-rs877710C-rs9593T haplo-type carriers interacted with cigarette smoking and age > 75 years to increase the risk of CHD. The rs3759387C-rs7134594T-rs877710G-rs9593A and rs3759387A-rs7134594T-rs877710G-rs9593A haplotype carriers interacted with hypertension and age > 75 years to increase the risk of IS.
These findings may reveal a disadvantageous effect of excessive alcohol consumption and cigarette smoking on serum lipid information. The present study has revealed that the mutual effect between rs7134594 CT genotypes and cigarette smoking to add serum HDL-C levels. Additionally, the rs3759387 CC, rs877710 CG and rs9593 AT genotypes interacted with alcohol consumption to increase serum HDL-C levels. Although this is a contradictory finding and the exactly mechanism is unknown, most smokers also get into habit of alcohol consumption, which may be a proper explanation in our study populations, and the potential interactions still need to be further confirmed.
A few potential limitations cannot be ignored. First, compared to many GWASes and replication studies, our sample numbers were relatively small. With these situations, larger sample numbers are needed to determine the consequences in future studies. Significant distinctions from demography were observed between the normal and patient groups. For the sake of statistical analysis accuracy, we adjusted for several environmental exposures, including time to life, sex, BMI, cigarette smoking, and alcohol drinking, but the potential influence of these factors on serum lipid concentrations and the risk of CHD and IS could not be completely eliminated. In addition, because many subjects were taking lipid-lowering drugs treatments, it was not proper to analyze the association of the four SNPs and serum lipid levels in the CHD and IS groups.
Last but not least, an individual’s genetic background and various environmental exposures can give rise to both CHD and IS, which are complex multifactorial disorders. The association between four MVK-MMAB SNPs and their haplotypes and the risk of CHD and IS had been probed, but many aspects remain to be explored, such as environmental exposures and genetic backgrounds and their mutual effects.
In conclusion, the consequences of the current study showed that the frequency of genotypes and alleles of the rs3759387 and rs7134594 SNPs was distinct from normals and patients. The rs3759387 genotypes were associated with the risk of CHD and IS in different genetic models. The rs3759387 SNP interacted with males to increase the risk of CHD and alcohol consumption to decrease the risk of CHD. The same SNP interacted with males and age > 75 years to increase the risk of IS. Four main haplotypes among the rs3759387, rs7134594, rs877710, and rs9593 SNPs were detected. The A-T-G-A and C-T-C-T haplotypes were associated with an increased risk of CHD. The A-T-G-A haplotype was associated with an increased risk of IS, whereas the C-T-G-A haplotype was associated with a decreased risk of IS. The A-T-G-A and C-T-C-T haplotypes interacted with cigarette smoking and age > 75 years to increase the risk of CHD. The C-T-G-A and A-T-G-A haplotypes interacted with hypertension and age > 75 years to increase the risk of IS. The individuals with the rs3759387AA genotype in normals took effect on reducing HDL-C concentrations than did the subjects with rs3759387AC/CC genotypes. The mutual effect between several SNPs and environmental exposures including alcohol consumption, cigarette smoking, and BMI ≥ 24 kg/m2 to adjust serum HDL-C and ApoA1 levels.
MATERIALS AND METHODS
Study patients
Totally, 1715 unrelated patients were recruited from the hospitalized patients who were treated in the First Affiliated Hospital, Guangxi Medical University. Among them, 846 subjects suffered from CHD, and another 869 patients were diagnosed with IS. CHD can be defined as including typical ischemic symptoms, plus one or more electrocardiographic changes (ST-segment depression or elevation of ≥ 0.5 mm, T-wave inversion of ≥ 3 mm in ≥ 3 leads, or left bundle branch block), in addition to increases in cardiac markers, such as creatinine kinase-MB and troponin T. Coronary angiography was carried out in patients with CHD. For the independent angiographers, two were blinded to the results of the genotypes. When coronary angiograms were performed, they were observed carefully. A vessel was chosen to be scored, and stenosis ≥ 50% had to be noted in an epicardial coronary vessel of interest or in one of its major branches. In the event of discordance of the number of vessels scored between the two reviewers, angiograms were scored by a third independent reviewer. The CHD subjects could be chosen for to our study when significant coronary stenosis (≥ 50%) was observed in at least one of the three main coronary arteries or their major branches (branch diameter ≥ 2 mm). In addition, the angiographic severity of disease was classified according to the number of coronary vessels with significant stenosis (luminal narrowing ≥ 50%) as one-, two-, or three-vessel disease in the three major coronary arteries [34, 35]. The definition of IS was ensured in accordance with the TOAST (Trial of Org 10172 in Acute Stroke Treatment) criteria [36] after rigorous examination, including neurological test, computed tomography, and/or magnetic resonance imaging (MRI). The IS patients entered in the study included individuals who were eligible for one of the two subtypes of TOAST criteria: large-artery atherosclerosis and small-vessel occlusion. However, if the subjects had a confirmed diagnosis of the below diseases, he/she must be excluded from our study: a history of hematologic or brain MRI revealing cerebral hemorrhage, cardioembolic stroke or unspecified stroke, neoplastic or intracranial space-occupying lesion, infection, other types of intracranial lesions, type 1 diabetes, and renal, liver, thyroid, and autoimmune diseases. IS patients who had a past history of CHD were excluded, as were CHD patients who had a past history of IS.
Control subjects
When CHD and IS patients were enrolled in our study, 847 control subjects, who were matched by age, gender, and ethnic group, were randomly selected from healthy adults who underwent periodical medical check-ups at the Physical Examination Center of the First Affiliated Hospital, Guangxi Medical University. The controls were healthy, without any CHD and IS details by questionnaires, history-taking, and clinical examination. The examination must be covered lots of items, just as physical examination, blood sampling, electrocardiography, chest X-ray, and Doppler echocardiography. All enrolled individuals were Han Chinese from Guangxi, the People’s Republic of China. The relevant information was gathered by trained research staff with standardized questionnaires for all participants, including demography, socioeconomic status, medical history and lifestyle exposures.
This study was carried out following the rules of the Declaration of Helsinki of 1975 (http://www.wma.net/en/30publications/10policies/b3/), revised in 2008. The study design was approved by the Ethics Committee of the First Affiliated Hospital, Guangxi Medical University (No: Lunshen-2011-KY-Guoji-001; Mar. 7, 2011). All procedures were performed in accordance with ethical standards. Informed consent was obtained from all participants before data collection.
Biochemical measurements
We received venous blood samples from all subjects after at least 12 h of fasting. The enzymatic methods with commercially available kits, including Tcho-1, TG-LH (RANDOX Laboratories Ltd., Ardmore, Diamond Road, Crumlin Co., Antrim, UK, BT29 4QY), Cholestest N HDL, and Cholestest LDL (Daiichi Pure Chemicals Co., Ltd., Tokyo, Japan) were used for detecting levels of serum TC, TG, HDL-C, and LDL-C, respectively, in samples. However, the immunoturbidimetric immunoassay (RANDOX Laboratories Ltd.) was needed to explore serum ApoA1 and ApoB concentrations. All determinations were performed with an autoanalyzer (Type 7170A; Hitachi Ltd., Tokyo, Japan) in the Clinical Science Experiment Center of the First Affiliated Hospital, Guangxi Medical University [37-48].
Diagnostic criteria
The levels were 3.10–5.17 (TC), 0.56–1.70 (TG), 0.91–1.81 (HDL-C), 2.70–3.20 (LDL-C) mmol/L, 1.00–1.78 (ApoA1), 0.63–1.14 g/L (ApoB), and 1.00–2.50 (ApoA1/B), defined as the normal values [37-49]. According to WHO diagnostic criteria, Type 2 diabetes should be defined as (1) fasting glucose (FPG) ≥ 7.0 mmol/L, (2) 2 h postprandial glucose ≥ 11.1 mmol/L, or (3) self-reported diagnosis of diabetes or use of anti-diabetic medications [50, 51]. Individuals with TC > 5.17 mmol/L and/or TG >1.70 mmol/L were diagnosed as hyperlipidemic [52-54]. The 1999 World Health Organization-International Society of Hypertension Guidelines were used to define the management of hypertension [55-57]. Normal weight, overweight, and obesity were defined as a BMI < 24, 24-28, and > 28 kg/m2, respectively [58, 59].
SNP selection and genotyping
We selected four SNPs in MVK/MMAB with the following steps. (1) MVK gene clusters, were selected from a previous GWAS associated with lipid-metabolism. MMAB gene clusters are found to be close to MVK gene clusters and associated with serum lipid levels, especially HDL-C. (2) Tagging SNPs were performed by Haploview (Broad Institute of MIT and Harvard, USA, version 4.2), and functional SNPs were predicted to lead to serum lipid changes from the current version of the online resource (1000 Genome Project Database). (3) SNPs information was obtained from NCBI dbSNP Build 132 (http://www.ncbi.nlm.nih.gov/SNP/). (4) SNPs were restricted to a minor allele frequency (MAF) > 1%. (5) SNPs might be associated with the serum lipid levels or cardiovascular disease in recent studies. (6) MVK rs3759587 and rs7134594 and MMAB rs9593 and rs877710 were selected by the block-based approach. This strategy was enabled by the correlations between tagging SNPs manifested as LD. Although classic tagging is not the goal of SNP selection, with innovative tagging SNPs selection bias is inevitable. [60-66]. Genomic DNA of the samples was isolated from peripheral blood leukocytes according to the phenol-chloroform method [37-49]. Genotyping of 4 mutations was performed by PCR-RFLP and determined by Sanger sequencing. The characteristics of each mutation and the details of each primer pair, annealing temperature, and length of the PCR products are summarized in Supplementary Table 1 and Supplementary Figures 2 and 3. The PCR products of the samples were sequenced with a sequencer ABI Prism 3100 Genetic Analyzer (Applied Biosystems, International Equipment Trading Ltd., Vernon Hills, IL, USA) at Shanghai Sangon Biological Engineering Technology & Services Co. Ltd., Shanghai China (Supplementary Figure 4).
Statistical analyses
We employed the statistical software package SPSS 22.0 (SPSS Inc., Chicago, IL, USA) to analyze the data. Quantitative variables are expressed as the means ± standard deviation (because serum TG was not a normal distribution, the levels are presented as medians and interquartile ranges and were analyzed by Wilcoxon-Mann-Whitney test). Qualitative variables are presented as percentages. Allele frequency was determined via direct counting, and the standard goodness-of-fit test was used to test Hardy-Weinberg equilibrium. When analyzing the difference in genotype distribution and sex ratio between the groups, we used a chi-square analysis. The general characteristics between patient and control groups were tested by the Student’s unpaired t-test. The association of genotypes and serum lipid parameters was tested by analysis of covariance (ANCOVA). Any variants associated with the serum lipid parameter at a value of P < 0.0125 (corresponding to P < 0.05 after adjusting for four independent tests by the Bonferroni correction) were considered statistically significant. After adjusting for the age, gender, BMI, smoking, and alcohol consumption, we employed unconditional logistic regression to evaluate the correlation between the risk of CHD and IS and genotypes. The same methods were used to calculate the odds ratio (OR) and 95% confidence interval (95% CI). When considering the interactions of four SNPs with environment exposures, including alcohol consumption, cigarette smoking, BMI ≥ 24 kg/m2, age, and sex, on serum lipid levels and the risk of CHD and IS, we employed a factorial regression to analysis [67-71] after controlling for potential confounders. A PI ≤ 0.00125 was considered statistically significant after Bonferroni correction (corresponding to P < 0.05 after adjusting for five environment exposures multiplied by eight outcomes by the Bonferroni correction). Haploview (Broad Institute of MIT and Harvard, USA, version 4.2) analyzed the haplotype frequencies and pairwise LD among the detected SNPs. The heart-map of the inter-locus models was measured by R software (version 3.3.0).
ACKNOWLEDGMENTS
L.M. conceived the study, participated in the design, undertook the genotyping, performed the statistical analyses, and drafted the manuscript. R.-X.Y. conceived the study, participated in the design, carried out the epidemiological survey, collected the samples, and helped draft the manuscript. F.H. collaborated with genotyping. W.-X.C., X.-L.C. and J.-Z.W. carried out the epidemiological survey and collected the samples. All authors read and approved the final manuscript.
CONFLICTS OF INTEREST
The authors have no potential conflicts of interest to report.
FUNDING
The authors acknowledge the essential role of the funding of the National Natural Science Foundation of China (No: 81460169) and the Innovation Project of Guangxi Graduate Education in this work.
REFERENCES
1. Ruff CT, Braunwald E. The evolving epidemiology of acute coronary syndromes. Nat Rev Cardiol. 2011; 8:140-147.
2. Strong K, Mathers C, Bonita R. Preventing stroke: saving lives around the world. Lancet Neurol. 2007; 6:182-187.
3. Murray CJ, Vos T, Lozano R, Naghavi M, Flaxman AD, Michaud C, Ezzati M, Shibuya K, Salomon JA, Abdalla S, Aboyans V, Abraham J, Ackerman I, et al. Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990-2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet. 2012; 380:2197-2223.
4. Deloukas P, Kanoni S, Willenborg C, Farrall M, Assimes TL, Thompson JR, Ingelsson E, Saleheen D, Erdmann J, Goldstein BA, Stirrups K, König IR, Cazier JB, et al, and CARDIoGRAMplusC4D Consortium, and DIAGRAM Consortium, and CARDIOGENICS Consortium, and MuTHER Consortium, and Wellcome Trust Case Control Consortium. Large-scale association analysis identifies new risk loci for coronary artery disease. Nat Genet. 2013; 45:25-33.
5. Wang F, Xu CQ, He Q, Cai JP, Li XC, Wang D, Xiong X, Liao YH, Zeng QT, Yang YZ, Cheng X, Li C, Yang R, et al. Genome-wide association identifies a susceptibility locus for coronary artery disease in the Chinese Han population. Nat Genet. 2011; 43:345-349.
6. Souiden Y, Mallouli H, Meskhi S, Chaabouni Y, Rebai A, Chéour F, Mahdouani K. MnSOD and GPx1 polymorphism relationship with coronary heart disease risk and severity. Biol Res. 2016; 49:22.
7. Yamada Y, Matsui K, Takeuchi I, Fujimaki T. Association of genetic variants with coronary artery disease and ischemic stroke in a longitudinal population-based genetic epidemiological study. Biomed Rep. 2015; 3:413-419.
8. Ding H, Xu Y, Wang X, Wang Q, Zhang L, Tu Y, Yan J, Wang W, Hui R, Wang CY, Wang DW. 9p21 is a shared susceptibility locus strongly for coronary artery disease and weakly for ischemic stroke in Chinese Han population. Circ Cardiovasc Genet. 2009; 2:338-346.
9. Banerjee A, Lim CC, Silver LE, Welch SJ, Banning AP, Rothwell PM. Familial history of stroke is associated with acute coronary syndromes in women. Circ Cardiovasc Genet. 2011; 4:9-15.
10. Sabater-Lleal M, Huang J, Chasman D, Naitza S, Dehghan A, Johnson AD, Teumer A, Reiner AP, Folkersen L, Basu S, Rudnicka AR, Trompet S, Mälarstig A, et al, and VTE Consortium, and STROKE Consortium, and Wellcome Trust Case Control Consortium 2 (WTCCC2), and C4D Consortium, and CARDIoGRAM Consortium. Multiethnic meta-analysis of genome-wide association studies in >100 000 subjects identifies 23 fibrinogen-associated Loci but no strong evidence of a causal association between circulating fibrinogen and cardiovascular disease. Circulation. 2013; 128:1310-1324.
11. Bevan S, Traylor M, Adib-Samii P, Malik R, Paul NL, Jackson C, Farrall M, Rothwell PM, Sudlow C, Dichgans M, Markus HS. Genetic heritability of ischemic stroke and the contribution of previously reported candidate gene and genomewide associations. Stroke. 2012; 43:3161-3167.
12. Williams FM, Carter AM, Hysi PG, Surdulescu G, Hodgkiss D, Soranzo N, Traylor M, Bevan S, Dichgans M, Rothwell PM, Sudlow C, Farrall M, Silander K, et al, and EuroCLOT Investigators, and Wellcome Trust Case Control Consortium 2, and MOnica Risk, Genetics, Archiving and Monograph, and MetaStroke, and International Stroke Genetics Consortium. Ischemic stroke is associated with the ABO locus: the EuroCLOT study. Ann Neurol. 2013; 73:16-31.
13. Dichgans M, Malik R, König IR, Rosand J, Clarke R, Gretarsdottir S, Thorleifsson G, Mitchell BD, Assimes TL, Levi C, O’Donnell CJ, Fornage M, Thorsteinsdottir U, et al, and METASTROKE Consortium, and CARDIoGRAM Consortium, and C4D Consortium, and International Stroke Genetics Consortium. Shared genetic susceptibility to ischemic stroke and coronary artery disease: a genome-wide analysis of common variants. Stroke. 2014; 45:24-36.
14. Willer CJ, Sanna S, Jackson AU, Scuteri A, Bonnycastle LL, Clarke R, Heath SC, Timpson NJ, Najjar SS, Stringham HM, Strait J, Duren WL, Maschio A, et al. Newly identified loci that influence lipid concentrations and risk of coronary artery disease. Nat Genet. 2008; 40:161-169.
15. Deodato F, Boenzi S, Santorelli FM, Dionisi-Vici C. Methylmalonic and propionic aciduria. Am J Med Genet C Semin Med Genet. 2006; 142C:104-112.
16. Feitosa MF, Rice T, North KE, Kraja A, Rankinen T, Leon AS, Skinner JS, Blangero J, Bouchard C, Rao DC. Pleiotropic QTL on chromosome 19q13 for triglycerides and adiposity: the HERITAGE Family Study. Atherosclerosis. 2006; 185:426-432.
17. Bossé Y, Chagnon YC, Després JP, Rice T, Rao DC, Bouchard C, Pérusse L, Vohl MC. Genome-wide linkage scan reveals multiple susceptibility loci influencing lipid and lipoprotein levels in the Quebec Family Study. J Lipid Res. 2004; 45:419-426.
18. Welch CL, Xia YR, Shechter I, Farese R, Mehrabian M, Mehdizadeh S, Warden CH, Lusis AJ. Genetic regulation of cholesterol homeostasis: chromosomal organization of candidate genes. J Lipid Res. 1996; 37:1406-1421.
19. Holleboom AG, Vergeer M, Hovingh GK, Kastelein JJ, Kuivenhoven JA. The value of HDL genetics. Curr Opin Lipidol. 2008; 19:385-394.
20. Dobson CM, Wai T, Leclerc D, Kadir H, Narang M, Lerner-Ellis JP, Hudson TJ, Rosenblatt DS, Gravel RA. Identification of the gene responsible for the cblB complementation group of vitamin B12-dependent methylmalonic aciduria. Hum Mol Genet. 2002; 11:3361-3369.
21. Ozcan O, Ipçioğlu OM, Gültepe M, Başoğglu C. Altered red cell membrane compositions related to functional vitamin B(12) deficiency manifested by elevated urine methylmalonic acid concentrations in patients with schizophrenia. Ann Clin Biochem. 2008; 45:44-49.
22. Chen Y, Zhu J, Lum PY, Yang X, Pinto S, MacNeil DJ, Zhang C, Lamb J, Edwards S, Sieberts SK, Leonardson A, Castellini LW, Wang S, et al. Variations in DNA elucidate molecular networks that cause disease. Nature. 2008; 452:429-435.
23. Sun J, Qian Y, Jiang Y, Chen J, Dai J, Jin G, Wang J, Hu Z, Liu S, Shen C, Shen H. Association of KCTD10, MVK, and MMAB polymorphisms with dyslipidemia and coronary heart disease in Han Chinese population. Lipids Health Dis. 2016; 15:171.
24. Junyent M, Parnell LD, Lai CQ, Lee YC, Smith CE, Arnett DK, Tsai MY, Kabagambe EK, Straka RJ, Province M, An P, Borecki I, Ordovás JM. Novel variants at KCTD10, MVK, and MMAB genes interact with dietary carbohydrates to modulate HDL-cholesterol concentrations in the Genetics of Lipid Lowering Drugs and Diet Network Study. Am J Clin Nutr. 2009; 90:686-694.
25. Gordon DJ, Probstfield JL, Garrison RJ, Neaton JD, Castelli WP, Knoke JD, Jacobs DR Jr, Bangdiwala S, Tyroler HA. High-density lipoprotein cholesterol and cardiovascular disease. Four prospective American studies. Circulation. 1989; 79:8-15.
26. Wilson PW, D’Agostino RB, Levy D, Belanger AM, Silbershatz H, Kannel WB. Prediction of coronary heart disease using risk factor categories. Circulation. 1998; 97:1837-1847.
27. Abbott RD, Donahue RP, Kannel WB, Wilson PW. The impact of diabetes on survival following myocardial infarction in men vs women. The Framingham Study. JAMA. 1988; 260:3456-3460.
28. Grundy SM, Cleeman JI, Merz CN, Brewer HB Jr, Clark LT, Hunninghake DB, Pasternak RC, Smith SC Jr, Stone NJ, and National Heart, Lung, and Blood Institute, and American College of Cardiology Foundation, and American Heart Association. Implications of recent clinical trials for the National Cholesterol Education Program Adult Treatment Panel III guidelines. Circulation. 2004; 110:227-239.
29. Gotto AM Jr, Brinton EA. Assessing low levels of high-density lipoprotein cholesterol as a risk factor in coronary heart disease - A working group report and update. J Am Coll Cardiol. 2004; 43:717-724.
30. Ng DS, Wong NC, Hegele RA. HDL-is it too big to fail? Nat Rev Endocrinol. 2013; 9:308-312.
31. Mackness B, Mackness M. The antioxidant properties of high-density lipoproteins in atherosclerosis. Panminerva Med. 2012; 54:83-90.
32. Kuijk LM, Beekman JM, Koster J, Waterham HR, Frenkel J, Coffer PJ. HMG-CoA reductase inhibition induces IL-1beta release through Rac1/PI3K/PKB-dependent caspase-1 activation. Blood. 2008; 112:3563-3573.
33. Murphy C, Murray AM, Meaney S, Gåfvels M. Regulation by SREBP-2 defines a potential link between isoprenoid and adenosylcobalamin metabolism. Biochem Biophys Res Commun. 2007; 355:359–64.
34. Chen Q, Reis SE, Kammerer CM, McNamara DM, Holubkov R, Sharaf BL, Sopko G, Pauly DF, Merz CN, Kamboh MI, and WISE Study Group. Association between the severity of angiographic coronary artery disease and paraoxonase gene polymorphisms in the National Heart, Lung, and Blood Institute-sponsored Women’s Ischemia Syndrome Evaluation (WISE) study. Am J Hum Genet. 2003; 72:13-22.
35. Xu Y, Wang W, Zhang L, Qi LP, Li LY, Chen LF, Fang Q, Dang AM, Yan XW. A polymorphism in the ABCG1 promoter is functionally associated with coronary artery disease in a Chinese Han population. Atherosclerosis. 2011; 219:648-654.
36. Adams HP Jr, Bendixen BH, Kappelle LJ, Biller J, Love BB, Gordon DL, Marsh EE 3rd. Classification of subtype of acute ischemic stroke. Definitions for use in a multicenter clinical trial. TOAST. Trial of Org 10172 in Acute Stroke Treatment. Stroke. 1993; 24:35-41.
37. Wu DF, Yin RX, Cao XL, Chen WX, Aung LH, Wang W, Huang KK, Huang P, Zeng XN, Wu J. Scavenger receptor class B type 1 gene rs5888 single nucleotide polymorphism and the risk of coronary artery disease and ischemic stroke: a case-control study. Int J Med Sci. 2013; 10:1771-1777.
38. Yin RX, Yang Q, Zhou YJ. Polymorphisms in the FADS1/FADS2 gene cluster are associated with the risk of coronary artery disease and ischemic stroke. J Am Coll Cardiol. 2014; 64:C117-C118.
39. Yin RX, Zhou YJ, Hong SC. Polymorphisms in the glucokinase regulator gene are associated with serum lipid levels and the risk of coronary artery disease and ischemic stroke. J Am Coll Cardiol. 2014; 64:C119.
40. Wu DF, Yin RX, Cao XL, Chen WX. Association between single nucleotide polymorphism rs1044925 and the risk of coronary artery disease and ischemic stroke. Int J Mol Sci. 2014; 15:3546-3559.
41. Nie RJ, Guo T, Cao XL, Chen WX, Wu JZ, Yin RX. Polymorphisms in the DOCK7 gene and the risks of coronary artery disease and ischemic stroke. J Am Coll Cardiol. 2015; 66:C268.
42. Zhou YJ, Yang Q, Yin RX. Polymorphisms in the CELSR2-PSRC1-SORT1 are associated with serum lipid traits, the risk of coronary artery disease and ischemic stroke. J Am Coll Cardiol. 2015; 66:C104-C105.
43. Zhou YJ, Hong SC, Yang Q, Yin RX, Cao XL, Chen WX. Association of variants in CELSR2-PSRC1-SORT1 with risk of serum lipid traits, coronary artery disease and ischemic stroke. Int J Clin Exp Pathol. 2015; 8:9543-9551.
44. Yang Q, Yin RX, Cao XL, Wu DF, Chen WX, Zhou YJ. Association of two polymorphisms in the FADS1/FADS2 gene cluster and the risk of coronary artery disease and ischemic stroke. Int J Clin Exp Pathol. 2015; 8:7318-7331.
45. Zhou YJ, Hong SC, Yin RX, Yang Q, Cao XL, Chen WX. Polymorphisms in the GCKR are associated with serum lipid traits, the risk of coronary artery disease and ischemic stroke. Int J Clin Exp Med. 2015; 8:10678-10686.
46. Nie RJ, Yin RX, Huang F, Cao XL, Wu JZ, Chen WX, Li ZM. Two DOCK7 polymorphisms and their haplotypes are associated with the risk of coronary artery disease and ischemic stroke. Int J Clin Exp Pathol. 2016; 9:2169-2180.
47. Zhang QH, Yin RX, Gao H, Huang F, Wu JZ, Pan SL, Lin WX, Yang DZ. Association of the SPTLC3 rs364585 polymorphism and serum lipid profiles in two Chinese ethnic groups. Lipids Health Dis. 2017; 16:1.
48. Yang Q, Zhou YJ, Yin RX. Polymorphisms in the MAFB gene are associated with the risk of coronary artery disease and ischemic stroke. J Am Coll Cardiol. 2015; 66:C139–140.
49. Yang Q, Yin RX, Zhou YJ, Cao XL, Guo T, Chen WX. Association of polymorphisms in the MAFB gene and the risk of coronary artery disease and ischemic stroke: a case-control study. Lipids Health Dis. 2015; 14:79.
50. Wang H, Qiu Q, Tan LL, Liu T, Deng XQ, Chen YM, Chen W, Yu XQ, Hu BJ, Chen WQ. Prevalence and determinants of diabetes and impaired fasting glucose among urban community-dwelling adults in Guangzhou, China. Diabetes Metab. 2009; 35:378-384.
51. Yang M, Chen P, Jin H, Xie X, Gao T, Yang L, Yu X. Circulating levels of irisin in middle-aged first-degree relatives of type 2 diabetes mellitus - correlation with pancreatic β-cell function. Diabetol Metab Syndr. 2014; 6:133.
52. Aung LH, Yin RX, Wu DF, Wang W, Liu CW, Pan SL. Association of the variants in the BUD13-ZNF259 genes and the risk of hyperlipidaemia. J Cell Mol Med. 2014; 18:1417-1428.
53. Guo T, Yin RX, Huang F, Yao LM, Lin WX, Pan SL. Association between the DOCK7, PCSK9 and GALNT2 gene polymorphisms and serum lipid levels. Sci Rep. 2016; 6: 19079.
54. Liao PJ, Xie RB, Yin RX, Wei DX, Huang J, Huang F, Li H, Lin WX, Wu JZ, Pan SL. Serum lipid profiles, the prevalence of dyslipidemia and the risk factors in two isolated Chinese minorities. Int J Clin Exp Med. 2015; 8:19200-19211.
55. Yin RX, Wu DF, Wu JZ, Cao XL, Aung LH, Miao L, Long XJ, Liu WY, Zhang L, Li M. Interactions of several lipid-related gene polymorphisms and cigarette smoking on blood pressure levels. Int J Biol Sci. 2012; 8:685-696.
56. Yin RX, Aung LH, Long XJ, Yan TT, Cao XL, Huang F. Interactions of several single nucleotide polymorphisms and alcohol consumption on blood pressure levels. J Am Coll Cardiol. 2015; 66:C197.
57. Yin RX, Aung LH, Long XJ, Yan TT, Cao XL, Huang F, Wu JZ, Yang DZ, Lin WX, Pan SL. Interactions of several genetic polymorphisms and alcohol consumption on blood pressure levels. Biofactors. 2015; 41:339-351.
58. Zhou BF, and Cooperative Meta-Analysis Group of the Working Group on Obesity in China. Predictive values of body mass index and waist circumference for risk factors of certain related diseases in Chinese adults - Study on optimal cut-off points of body mass index and waist circumference in Chinese adults. Biomed Environ Sci. 2002; 15:83-96.
59. Wildman RP, Gu D, Reynolds K, Duan X, He J. Appropriate body mass index and waist circumference cutoffs for categorization of overweight and central adiposity among Chinese adults. Am J Clin Nutr. 2004; 80:1129-1136.
60. Aulchenko YS, Ripatti S, Lindqvist I, Boomsma D, Heid IM, Pramstaller PP, Penninx BW, Janssens AC, Wilson JF, Spector T, Martin NG, Pedersen NL, Kyvik KO, et al, and ENGAGE Consortium. Loci influencing lipid levels and coronary heart disease risk in 16 European population cohorts. Nat Genet. 2009; 41:47-55.
61. Huang KK, Yin RX, Zeng XN, Huang P, Lin QZ, Wu J, Guo T, Wang W, Yang DZ, Lin WX. Association of the rs7395662 SNP in the MADD-FOLH1 and several environmental factors with serum lipid levels in the Mulao and Han populations. Int J Med Sci. 2013; 10:1537-1546.
62. Wang XB, Han YD, Cui NH, Gao JJ, Yang J, Huang ZL, Zhu Q, Zheng F. Associations of lipid levels susceptibility loci with coronary artery disease in Chinese population. Lipids Health Dis. 2015; 14:80.
63. Fedele F, Mancone M, Chilian WM, Severino P, Canali E, Logan S, De Marchis ML, Volterrani M, Palmirotta R, Guadagni F. Role of genetic polymorphisms of ion channels in the pathophysiology of coronary microvascular dysfunction and ischemic heart disease. Basic Res Cardiol. 2013; 108:387.
64. Fedele F, Severino P, Bruno N, Stio R, Caira C, D’Ambrosi A, Brasolin B, Ohanyan V, Mancone M. Role of ion channels in coronary microcirculation: a review of the literature. Future Cardiol. 2013; 9:897-905.
65. Carty CL, Buzková P, Fornage M, Franceschini N, Cole S, Heiss G, Hindorff LA, Howard BV, Mann S, Martin LW, Zhang Y, Matise TC, Prentice R, et al. Associations between incident ischemic stroke events and stroke and cardiovascular disease-related genome-wide association studies single nucleotide polymorphisms in the Population Architecture Using Genomics and Epidemiology study. Circ Cardiovasc Genet. 2012; 5:210–16.
66. Franceschini N, Carty C, Bůzková P, Reiner AP, Garrett T, Lin Y, Vöckler JS, Hindorff LA, Cole SA, Boerwinkle E, Lin DY, Bookman E, Best LG, et al. Association of genetic variants and incident coronary heart disease in multiethnic cohorts: the PAGE study. Circ Cardiovasc Genet. 2011; 4:661–72.
67. Finlay KW, Wilkinson GN. The analysis of adaptation in a plant-breeding program. Aust J Agric Res. 1963; 14: 742-754.
68. Wood JT. The use of environmental variables in the interpretation of genotype-environment interaction. Heredity. 1976; 37: 1-7.
69. Denis JB. Analyse de regression factorielle. Biom Praxim. 1980; 20: 1-34.
70. Denis JB, Vincourt P. Panorama des methodes statistiques d’analyse des interactions genotype X milieu. Agronomie. 1982; 2:219–30.
71. Denis JB. Two-way analysis using covariates. Statistics. 1988; 19: 123-132.