
INTRODUCTION
Polysaccharides belong to a class of natural polymers that are comprised of carbohydrate monomers linked by glycosidic linkages [1]. The influence of polysaccharides’ chemical composition, molecular weight and structure on physiological functions is variable depending on their sources. Polysaccharides also possess various physicochemical properties such as gelation, solubility, low osmotic effect and surface properties which depend on their composition and chemical structure [2, 3].
Lentinan, a (1-3)-beta-d-glucan extracted from the mushroom Lentinus edodes, is a potent anti-cancer drug that has been licensed in Japan for anti-tumor therapy since 1985 [4–6]. Polysaccharides play important roles in the rehabilitation of patients with numerous diseases. In addition to anti-diabetic, antibiotic, antioxidant, anti-mutant, and anticoagulant activity, polysaccharides and their derivatives have shown remarkable efficacy in combating certain cancers [7]. A large number of studies have suggested that polysaccharides exhibit an anti-cancer effect following some common mechanisms: (1) prevention of oncogenesis by oral consumption of active preparations [8]; (2) enhancement of immunity against tumors directly or indirectly through combination with chemotherapy [9]; (3) inhibiting tumors such as induction of tumor cell apoptosis or inhibition of tumor metastasis [10, 11]; (4) activation of humoral and cellular immunity to prevent cancer cell proliferation [12]. Activity of immunity-enhancement is important function of lentinan, which has become a hot topic in lentinan study [13]. The mechanisms for polysaccharides enhancing immune system involve both improvement of host defense against pathogens and modulation of adaptive immunity. Regulation of the levels of lymphocytes’ proliferative activity and macrophages’ phagocytic activity, cytokines, and antibodies may contribute to lentinan’s immunity-enhancement abilities [14, 15].
How to take advantage of polysaccharides such as lentinan appropriately has not been noted carefully, even if low toxicity or certain adverse effects have been reported [16]. In previous studies, the effects of cartilage polysaccharide on the apoptosis of human hepatoma BEL-7402 cells and immunoprophylaxis of murine H22 Hepatocarcinoma were reported. However, the survival status of KM mice immunized with cartilage polysaccharide was worse comparing with H22 hepatocarcinoma model mice [17].
In this study, we explored the effects of lentinan on cytotoxicity of HL7702 cells or H22 cells and mouse immunity against initial proliferation of H22 hepatocarcinoma. MTT assay was selected to test the cytotoxicity of lentinan on HL7702 cell and H22 cell in vitro. The cumulative survival assay, spleen lymphocyte proliferation tests and peritoneal macrophage phagocytosis assays were selected in vivo for the mice immunized with lentinan. Mass spectrometry (MS)-based proteomics was also used to profile the protein emerging of the H22 cell incubated with lentinan.
RESULTS
Cytotoxicity of lentinan in vitro
To confirm the general cytotoxicity of lentinan on HL7702 cells and H22 cells, cell proliferation condition was measured by MTT assay. For HL7702 cells, the RGR of the control cells was considered as 100%. After treatment with different concentrations of lentinan (0.02, 0.04, 0.08, 0.16, 0.32, 0.64, 1.28 and 2.56 mg/mL) for 24 h. The proliferation rate of HL7702 cells did not increased in a dose-dependent manner (Figure 1A), and there was no significantly increased compared to the control group. The viability of H22 cells after treatment with different concentrations of lentinan (0.02, 0.04, 0.08, 0.16, 0.32, 0.64, 1.28 and 2.56 mg/mL) was also measured and the average inhibition rate was 5.05%, 7.28%, 8.44%, 6.63%, 18.34%, 29.31%, 85.48 and 94.89% respectively. The inhibition rate of H22 cells increased in a dose-dependent manner (Figure 1B). And the inhibition rate of H22 cells significantly increased (P < 0.01) compared to the control group when they were exposed from 0.16 to 2.56 mg/mL of lentinan.
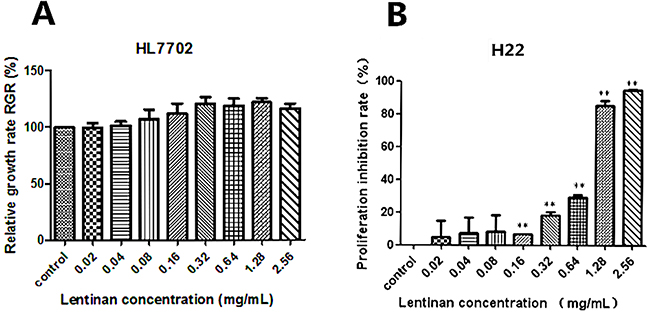
Figure 1: Effects of lentinan on cytotoxicity in vitro. Human normal liver cell line HL7702 and murine liver cancer cell line H22 were treated with different concentrations of lentinan (0.02, 0.04, 0.08, 0.16, 0.32, 0.64, 1.28 and 2.56 mg/mL) for 24 h. The relative growth rate (RGR, %) of HL7702 cells and proliferation inhibition rate of H22 cells were tested. (A) The relative growth rate (RGR, %) of HL7702 cells treated with different concentrations of lentinan. (B) The proliferation inhibition rate of H22 cells treated with different concentrations of lentinan. The asterisks represent statistical significance (*P < 0.05 and **P < 0.01) compared with the control value.
Effects of lentinan on mice in vivo
The variation rates of the average body weight of the mice in L1 and L2 groups were between that of the control and model groups. The increase of the average mouse body weight in L3 group was higher than that of the model group (P<0.05). After four days, mouse body weight in L3 group showed significant increased compared to control group, while there was no significant difference in the other groups. The increase velocity of the average mouse body weight in other groups were model >L1 >L2 >control group from day 8 to 12 (Figure 2A). As shown in Figure 2B, the survival rate (20%) of the mice in L2 group was the highest among all the experimental groups, while the survival rates in other groups (L1 and L3 groups) were zero.
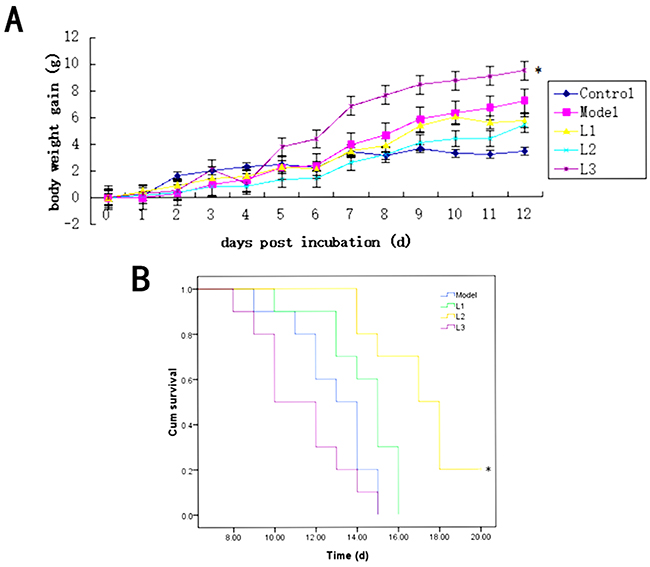
Figure 2: Effects of L1, L2, and L3 on mice body weight and survival. L1, the concentration of lentinan was 0.02mg/kg body weight; L2, the concentration of lentinan was 0.4mg/kg body weight; and L3, the concentration of lentinan was 1mg/kg body weight. (A) Average body weight gain of mice in each group after modeling. (B) The increase in life span (ILS) or survival rate of each group. *P < 0.05 compared to model.
Analysis of the immunoprophylaxis activity of lentinan on H22 hepatocarcinoma
Firstly, the spleens of mice in experimental (L1, L2 and L3) and model groups appeared visibly different from that of control group. The spleens’ khaki atrophy of mice in L1, L3 and model groups had been observed visibly. However, for spleens of mice in L2 group, morphological characters were purplish red color and plump which were close to the control group (Figure 3A). The diverification of immune organ index were listed in Figure 3B/3C. The spleen (si, 1.21) and thymus (ti, 0.49) indices in L3 group showed significantly lower values than those of control group (P<0.01).
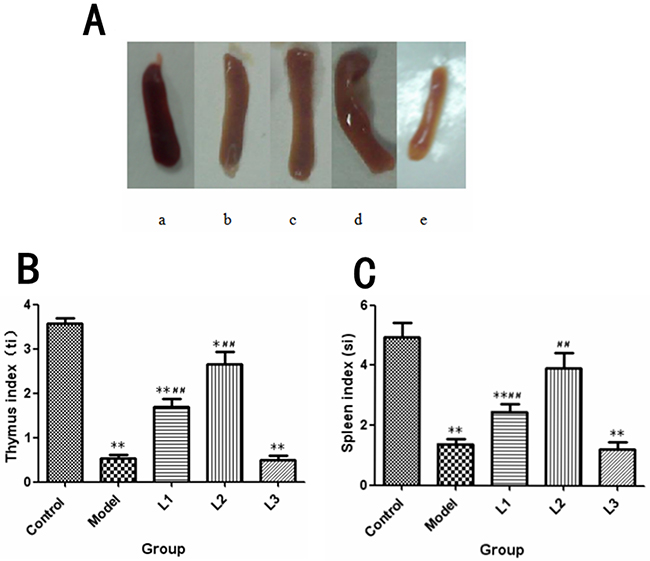
Figure 3: The changes in immune organs in each group. L1, the concentration of lentinan was 0.02mg/kg body weight; L2, the concentration of lentinan was 0.4mg/kg body weight; or L3, the concentration of lentinan was 1mg/kg body weight. (A) The appearance of the spleen in each group of mice was recorded. a: control group, b: model group, c: L1 group, d: L2 group, e: L3 group. (B) ti (thymus index) of each group was tested. (C) si (spleen index) of each group was tested. *P<0.05 vs control, **P<0.01 vs control, #P < 0.05 compared to model, ##P < 0.01 compared to model.
The SI (stimulation index) of each group was shown in Figure 4. It was showed that the proliferation rate of spleen lymphocytes in L3 group (SI-ConA, 1.8; SI-LPS, 1.7) was remarkably depressed compared to that of control group (P<0.01). While for the mice L2 group, which was treated with 0.4mg/kg body weight of soluble lentinan, showed a significant improvement in both T-cell (SI-ConA, 8.1) and B-cell (SI-ConA, 6.37) proliferation rate compared to that of model group (P<0.01).
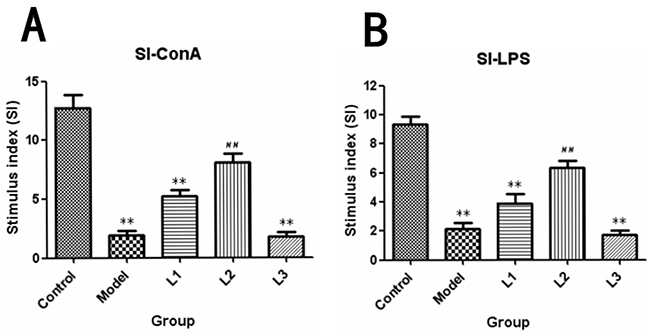
Figure 4: SI-stimulus index induced by ConA or LPS in lentinan groups of mice. L1, the concentration of lentinan was 0.02mg/kg body weight; L2, the concentration of lentinan was 0.4mg/kg body weight; and L3, the concentration of lentinan was 1mg/kg body weight. (A) Stimulus index (SI)-ConA. (B) Stimulus index (SI)-LPS. **P < 0.01 compared to control, ##P < 0.01 compared to model.
As shown in Figure 5, the macrophage phagocytic indexs in L1 (phagocytic indexs, 0.44) and L3 (phagocytic indexs, 0.41) groups were significantly lower than that of control group (P<0.05), while there was no significant difference in phagocytosis rate (P>0.05) between experimental (lentinan) groups and control or model groups.
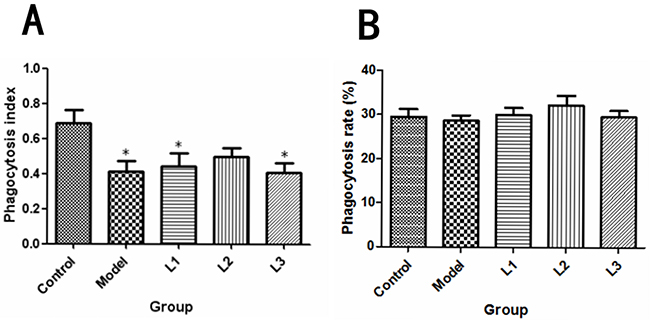
Figure 5: Effects of lentinan on mice peritoneal macrophage phagocytosis. L1, the concentration of lentinan was 0.02mg/kg body weight; L2, the concentration of lentinan was 0.4mg/kg body weight; and L3, the concentration of lentinan was 1mg/kg body weight. (A) Macrophage phagocytosis rate of each group (control, model, L1, L2, and L3) was calculated. (B) Macrophage phagocytosis index of each group (control, model, L1, L2, and L3) was tested. *P<0.05 vs control group.
MS analysis of emerging protein
MS/MS analysis and protein database searching were carried out to investigate the change of protein after the treatment of lentinan. Mass spectrometry analysis of H22 cells (incubated without lentinan) protein as a control versus H22 cells (incubated with lentinan) protein. In total 309 proteins were identified in H22 cells, whereas 96 new emerging proteins were detected in the H22 cells incubated with lentinan. According to the protein functional studies we focused on 6 proteins which could promote cancer occurrence or cancer cell proliferation. The 6 proteins were 60Sacidic ribosomal protein P2, Peroxiredoxin-2, Annexin A5, PDZ and LIM domain protein 1, Src substrate cortactin and Moesin as shown in Table 1.
Table 1: Liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis of the protein samples
Accession |
Protein name |
Sum PEP score |
Coverage |
# Peptides |
# PSMs |
# Unique peptides |
# AAs |
MW [kDa] |
Calc. pI |
Area |
|---|---|---|---|---|---|---|---|---|---|---|
P99027 |
60Sacidic ribosomal protein P2 |
6.465735225 |
21.73913043 |
2 |
2 |
2 |
115 |
11.644 |
4.54 |
5.30E+07 |
Q61171 |
Peroxiredoxin-2 |
5.603677658 |
22.72727273 |
3 |
3 |
3 |
198 |
21.765 |
5.41 |
8.70E+07 |
P48036 |
Annexin A5 |
5.518135923 |
7.836990596 |
2 |
2 |
2 |
319 |
35.73 |
4.96 |
1.50E+07 |
O70400 |
PDZ and LIM domain protein 1 |
5.306689611 |
6.727828746 |
2 |
2 |
2 |
327 |
35.752 |
6.84 |
2.70E+07 |
Q60598 |
Src substrate cortactin |
4.602854608 |
5.860805861 |
2 |
2 |
2 |
546 |
61.212 |
5.4 |
5.30E+07 |
P26041 |
Moesin |
4.69751607 |
4.679376083 |
3 |
3 |
1 |
577 |
67.725 |
6.6 |
1.50E+07 |
DISCUSSION
Searching for novel effective anticancer agents with less toxic effects is a hot topic for scientists to solve the problems in cancer therapy. Lentinus edodes, with a long history as a food source, have been believed to have health benefits including anti-tumour [18, 19], immuno-modulatory [20] and anti-microbial effects [21]. Lentinan, a polysaccharide extracted from lentinus edodes, is the best known and most potent mushroom-derived substance with anti-tumor and immuno-modulating properties [20, 22].
In the study, the cytotoxicity of lentinan over intake was tested by MTT assay in vitro with human normal liver cell line HL7702 and murine liver cancer cell line H22. The results revealed that lentinan was a relatively safe agent at concentration from 0.001mg/mL to 2mg/mL. For antitumor effect of lentinan in vitro, the proliferation rates of H22 cells treated with lentinan (0.16, 0.32, 0.64, 1.28 and 2.56 mg/mL) were inhibited obviously in a dose-dependent manner. Then the side-effect of lentinan on cancer immune especially on cancer cells clear in time should be tested in vivo.
In order to study the effect of lentinan on murine H22 hepatocarcinoma immunoprophylaxis, we immunized mice with lentinan. The dose conversion coefficients of per kilogram body weight from mice (20g) to human (70kg) was 9.01. The lentinan at concentration of 0.02mg/kg, 0.4mg/kg and 1mg/kg body weight were selected respectively to immunized the mice which were establish a mice model bearing with H22 hepatocarcinoma. Then We examined the physiological mechanisms activated by polysaccharides for the immunized mice.
The increase of mice body weight indicated the proliferation of ascitic tumor cells. In this study, the increase of mice body weight in L1 and L2 groups was less than that of model group, which suggested that the proliferation of H22 hepatocarcinoma cells could be inhibited to some extent treated by 0.02mg/kg or 0.4mg/kg body weight of lentinan. However, we were also puzzled by the phenomenon that the body weight increase in L3 group was significantly higher than that of control group from the fourth day to the twelfth day, which may indicate the proliferation of cancer cells could be promoted by 1mg/kg body weight of lentinan (L3 group). The survival rate of the mice in L2 group reached 20% which was higher than that of model group significantly (P < 0.05).
Thymus and spleen are important immune organs for vertebrates. To a certain extent, the immune condition of the body can be checked by organ appearance or an organ weight index. The growth index of the spleen (spleen weight normalized to body weight, SI), which indicates the immune function changes of the spleen [23]. The appearance or organ weight index of the spleen in L2 group was different from that of model group. The spleen and thymus indexes in L3 group were significantly lower than that of model group, indicating that treatment by high dose of lentinan (L3) before immunization could accelerate the proliferative rates of cancer cells by decrease of the immune organ functions.
The results showed that immunization with lentinan could contribute to the proliferation of both T and B lymphocytes of the mice in L2 group, nevertheless this effect was dose-independent (Figure 3). Furthermore, the proliferation of T and B lymphocytes in L3 group was lower than that of control group (P<0.01). The proliferation of T and B lymphocytes is an immune parameter in the investigations of lymphocyte responsiveness considering its considerable sensitivity. Cellular proliferation induced by ConA is commonly used to detect T lymphocyte immunity, and LPS-induced activation of B cells indicates B lymphocyte immunity [24, 25]. Previous reports also indicated that lentinan could promote both humoral and cellular immune responses against ovarian cancer, gastric cancer, colorectal cancer, and liver cancer [22, 24, 26, 27].
In the present study, the phagocytic index of the immunized mice in L1 or L3 group was decreased compared with control group but not model group. There were no significant difference in the phagocytic rate among control group, model group and immunized groups. Peritoneal macrophages are important for cancer immunity, because they can kill cancer cells directly or indirectly [28]. Therefore, we speculated that stimulation of macrophages may be one of the most critical mechanisms for the cancer inhibitory activity exerted by polysaccharides.
Using MS analysis, six proteins existed in H22 cells incubated with lentinan were the targets we focus on. These 6 proteins 60Sacidic ribosomal protein P2, Peroxiredoxin-2, Annexin A5, PDZ and LIM domain protein 1, Src substrate cortactin and Moesin which had relationship with cancer proliferation in vitro or in vivo according to the previous reports. 1) The phosphorylation level of P2 protein is one of the regulatory mechanisms for the overall rate of protein elongation. Thus, increased expression of P2 protein can increasing proliferation rate of cancer cells [29–31]. 2) Peroxiredoxin 2 is a member of the peroxiredoxin family, which has been found to be elevated in several human cancer cells and tissues, including colorectal cancer, pancreatic cancer and breast cancer, and it influences cells’ survival, proliferation, and apoptosis processes, which suggests a possible role for Peroxiredoxin 2 in the maintenance of cancer cell[32–34]. 3) Annexin A5 a calcium-binding protein which is involved in membrane organization and dynamics has been implicated in the carcinogenesis of several carcinomas. The biological function of endogenous annexin A5 was its possible influence on proliferation and invasion capacity [35, 36]. 4) PDZ and LIM domain protein 1 (PDLIM1) also known as CLP36, Elfin or CLIM1 is critical for promoting breast cancercell migration and invasion in vitro and metastasis in vivo, whereas it is dispensable for breast cell proliferation and anchorage-independent growth in vitro and tumor growth in vivo [37]. 5) Cortactin frequently over expressed in cancer was first identified as one of the major substrates for src kinase [38]. Promotion of tumor invasion and metastasis was the role of cortactin over expression in vivo. Many other studies suggest that cortactin promotes cell motility and invasion, including a critical role in invadopodia, actin rich-subcellular protrusions associated with degradation of the extracellular matrix by cancer cells [39]. 6) Moesin which plays a key role in the control of cell morphology, motility, adhesion and other processes of tumourigenesis is a linker between the actin cytoskeleton and the plasma membrane [40]. The study of Wu M, et al. has showed the expression of moesin was strongly negatively correlated with the patient progression-free survival and overall survival. And the moesin protein involved in the genesis and progression of astrocytomas and might be regarded as an independent predictor of poor prognosis [41].
MATERIALS AND METHODS
Animals
SPF female KM mice weighted 18-22 g were purchased from the animal center of Hebei Medical University (SCXK (JI) 2013-1-003, 1410014, Shijiazhuang, China). All experiments were conducted following the Guide for the Care and Use of Laboratory Animals of National Institutes of Health. This study was approved by the animal ethics committee of Hebei University of Science and Technology(Permission number: 2014-A02-01). The animals were maintained in an environment with controlled temperature (20 to 25 °C) and humidity (50% ± 5%), with food and water available at any time and a natural light. The health of the mice was monitored every day. Before blood collection or immune organ index test, mice were anesthetized with ether, and the other mice were euthanized by cervical dislocation under anesthesia with isoflurane.
Cell lines
The human normal liver cell line HL7702 cells and murine liver cancer cell line H22 cells (China PLA General Hospital, Beijing, China) were cultured in RPMI 1640 medium supplemented with 10% heat-inactivated fetal bovine serum (FBS) and antibiotics (100U/ml penicillin and 100 g/ml streptomycin) at 37°C in an atmosphere of 5% CO2.
Lentinan solution
Lentinan used in this study was gifted from Kang Yuan (Jiangsu, China). Different concentrations of lentinan saline solution were prepared, 0.02 mg/mL, 0.04 mg/mL, 0.08 mg/mL, 0.16 mg/mL, 0.32 mg/mL, 0.64 mg/mL, 1.28 mg/mL, 2.56 mg/mL and 0.02mg/kg body weight (L1group), 0.4mg/kg body weight (L2 group), 1mg/kg body weight (L3 group) were used for the tests in vitro and in vivo respectively.
Cytotoxicity assay in vitro
The cytotoxicity of lentinan on HL7702 and H22 cells in vitro was tested by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay. HL7702 and H22 cells at logarithmic phase were seeded in 96-well micro-plates (Corning, NY, USA) at a density of 5×103 and 1×106 cells per well per 100 μL of medium respectively. After incubation for 24 h, the cells were treated with various concentrations of lentinan solution (0.02-2.56 mg/mL) and continued to be incubated at 37°C for another 24h. Then the cells seeded in 96-well plates were harvested and pre-incubated with 0.5 mg/ml MTT (Solarbio, Beijing, China) for 4 h at 37 °C. When MTT incubation was completed, supernatant was discarded. Then, 150 μL of dimethyl sulfoxide (DMSO, Solarbio, Beijing, China) was added to each well and incubated for 10 min with shaking to dissolve the crystallization. Finally, the absorbance of each well was measured at 570 nm using a microplate reader (Model 680, Bio-Rad, Hercules, Calif., U.S.A.). RPMI-1640 medium was used to determine the background (blank) OD, which was subtracted from the OD of the samples. The relative growth rate (RGR, %) (n=3) of HL7702 cell was: ODtreatment / ODcontrol×100%. The inhibition rate for H22 cell proliferation (%) (n=3) was : (ODcontrol–ODtreatment) / ODcontrol×100%.
Establishment of mouse model bearing H22 tumors
A total of 100 KM mice were randomly divided into a control group, model group, lentinan 1(L1) group, lentinan 2 (L2) group, and lentinan 3 (L3) group,with 20 mice in each group. The lentinan groups were immunized (intraperitonially [i.p.]) with lentinan saline solution (L1: 0.02mg/kg body weight ; L2: 0.4mg/kg body weight; 1mg/kg body weight) once a week for 3 weeks. The control group was treated with 0.9% normal saline. The model group was used to be as positive group and treated with 0.9% normal saline. One week after the third immunization, all of the mice except the control group were injected i.p. with H22 hepatocarcinoma cells (1.5×106 cells/mouse) to establish the H22 hepatocarcinoma mouse model. After establishment of the H22 mouse model, the body weight (BW) of the mice in each group was measured and recorded every day for 12 days. Ten mice in each group were selected randomly to calculate the immune organ (thymus and spleen) index or cellular immunity (T-cell proliferation test and peritoneal macrophage phagocytosis) on the twelfth day after establishment of the H22 mouse model. The spleens or thymus were collected from each mouse, and the growth index for the thymus and spleen were calculated and recorded.
Growth index of spleen = spleen weights (mg) / BW (g)
Growth index of thymus = thymus weights (mg) / BW (g)
Twenty days after establishment of the H22 mouse model, the cum survival of the mice in each group were tested with Kaplan-Meier method (n=10).
Spleen lymphocyte proliferation tests
For the lymphocyte proliferation assay, spleen lymphocytecells from each group were collected and suspended at concentration of 2×106 viable cells/mL in RPMI 1640 (containing 10% FBS) medium. Then Concanavalin A (ConA) or Lipopolysaccharide (LPS) (10 μg/mL) were mixed with spleen cell suspension at a ratio of 3:1 and added in 96-well plates, 100 μL/well. The cell cultures were incubated at 37 °C for 72 h. Four h before the cells were harvested, 0.5 mg/mL of MTT solution was added. Then 150 μL of DMSO was added to each well and incubated for 10 min with shaking to dissolve the crystallization. Finally, the absorbance of each well was read at 570 nm using a microplate reader.
Stimulus index (SI)= ODexperimental / ODcontrol
Peritoneal macrophage phagocytosis assays
In order to test the phagocytosis activity of peritoneal macrophages, mice in each group was injected (i.p.) with bouillon culture medium (Amidulin, 6%) once a day. Three days later, Chick erythrocytes (1% suspension) was injected (i.p.) into mice and the specimens of peritoneal fluid was harvested 0.5 h later. Then the peritoneal fluid of mice in each group was smeared on glass slides and stained with Wright Giemsa. 100 macrophages of each sample were observed. When chick erythrocytes appeared in the cytoplasm, the macrophages cells were considered to be actively phagocytic. The number of the activated macrophages and the number of chick erythrocytes in macrophages were recorded. The phagocytosis rate and phagocytosis index were calculated according to the reference [12].
Phagocytosis index= number of the phagocytic chick erythrocyte/100
Phagocytosis rate (%) = (number of the activated macrophages/100) × 100%
LC-MS/MS analysis and protein database searching for emerging protein
H22 cells at logarithmic phase were incubated with or without lentinan (1.28mg/mL) at 37°C for 24h. The co-culture was frozen, thawed 5 times to inactivate living H22 hepatocarcinoma cells with PMSF and the supernatant was selected as samples. The protein sample was further concentrated using Amicon Ultra-15 centrifugal filter MWCO3000 (Millipore). Protein concentration of the samples was measured by Bradford Assay (Solarbio, Bradford Protein Assay Kit, Beijing, China). An aliquot of the concentrated preparation containing 100 μg protein was diluted with 50 mM NH4HCO3 to 70μL and added DTT with final concentration 10mM at 56°C for 1h. The sample was then incubated with iodoacetamide (10mM final concentration) at room temperature for 40 minutes before digestion with trypsin (trypsin:protein 1:50 w/w) at 37 °C for 14 h. After speed-vac, the sample peptides were reconstituted in 50 mM NH4HCO3 for LC-MS analysis.
Liquid chromatograohy/mass spectrometry was performed on Thermo Scientific™ Q Exactive equipped with a Nanospray Floex Ion-Soure. Peptides suspension dissolved in 0.1% formic acid were separated by nano-high performance liquid chromatography (Eksigent Technologies) on a secondary reversed-phase analytical column (Eksigent, C18, 3μm, 150 mm*75μm). Buffer A for the pump consisted of 0.1% formic acid in LC/MS-grade water; buffer B for the pump consisted of 0.1% formic acid in LC/MS-grade acetonitrile. Gradient conditions for pump B were as follows: 0% to 35% B from 0 to 90 minutes. A total of 5 mL of the prepared peptides was injected onto the trap column for concentration/purification. Flow rates were 300 nL/min. Some parameters in Orbitrap were as follows: spray voltage, 2.0 kV; capillary temperature, 250°C; m/z (mass to charge ratio) range (ms), 350 to 1800. AGC ion injection targets for each FTMS scan were 70,000 (100 ms max injection time). AGC ion injection targets for each MS2 scan were 17,500 (50 ms max ion injection time). Full MS/dd-MS2 (Top10) was used in this analysis, with a dynamic exclusion time of 25 s. Both identification and quantification were done by Proteome Discoverer (version 2.1demo, Thermo Fischer Scientific). Uniprot-mouse database was used for data mining. Peptides were generated from a tryptic digestion with up to 2 missed cleavages, dynamic modifications of methionine oxidation, and a static modification of cysteine carbamidomethylation. Precursor mass tolerance was 10 ppm, and product ions were searched at 0.02-Da tolerances. Peptide spectral matches were validated using percolator based on q values at a 1% false discovery rate.
Statistical analysis
The data were analyzed using SPSS software (version 11.5, SPSS, New York, NY, USA) and P < 0.05 was considered as statistically significant. All the data were presented as mean±SD.
CONCLUSION
In conclusion, the safety of lentinan was relative. Lentinan (from 0.16 mg/mL to 2.56 mg/mL) was non-toxic for normal liver HL7702 cells, and the H22 mice liver cancer cells proliferation were inhibited by lentinan dramatic in vitro. On the other hand, lentinan could promote the cancer cell proliferation at concentration of greater than 1mg/kg body weight in vivo. The cellular immune functions of H22 model mice were improved in L2 group (0.4 mg/kg body weight lentinan), mainly reflected in inhibition of cancer cell proliferation and protection of immune organs. Therefore, L2 may assist in liver cancer immunoprophylaxis. Conversely, L3 (1mg/kg body weight lentinan) not only did not inhibit cancer cell proliferation, but it promoted the proliferation of cancer cells. 60Sacidic ribosomal protein P2, Peroxiredoxin-2, Annexin A5, PDZ and LIM domain protein 1, Src substrate cortactin and Moesin were detected by MS assay. Based on the previous studies, these six proteins may play an important role in the promotion of cancer cell proliferation in this study. Thus, we need to be attentive to the dose of lentinan used in clinics based on these study results. As a double-edged sword, lentinan may make important contributions to cancer therapy or immunoprophylaxis when we used properly.
Author contributions
Ying Wang, Xue Han and Dong Jie Zhang designed the study and prepared the manuscript.
Ying Wang, Xue Han conducted both in vitro and in vivo studies.
Yan Dong Li conducted cell studies and MS assay.
Yabing Wang performed spleen lymphocyte proliferation tests and peritoneal macrophage phagocytosis assays.
Shi Yang Zhao carried out animal studies.
Yu Lu completed statistical analysis
ACKNOWLEDGMENTS
The authors thank Dr. Lian-feng Ai of Inspection and Quarantine Technology Center at Hebei entry-exit inspection and Quarantine Bureau for assistance with the data analysis of the MS experiment.
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest.
FUNDING
The present work was supported by grants from the National Science and Technology Supporting Plan of China (No. 2012BAD33B00); the Natural Science Foundation of Hebei Provence (No. C2014208064); the Science and Technology Project of Hebei Provincial Bureau of Animal Husbandry and Veterinary (No. 2014-1-05).
REFERENCES
1. Ferreira SS, Passos CP, Madureira P, Vilanova M, Coimbra MA. Structure-function relationships of immunostimulatory polysaccharides: a review. Carbohydr Polym. 2015; 5: 378-396.
2. Marguerite R. Biomaterials based on a natural polysaccharide: alginate. TIP. 2014; 17: 92-96.
3. Zhang L, Tu ZC, Wang H, Kou Y, Wen QH, Fu ZF, Chang HX. Response surface optimization and physicochemical properties of polysaccharides from Nelumbo nucifera leaves. Int J Biol Macromol. 2015; 74: 103-110.
4. Chen YW, Hu DJ, Cheong KL, Li J, Xie J, Zhao J, Li SP. Quality evaluation of lentinan injection produced in China. J Pharm Biomed Anal. 2013; 78-79: 176-182.
5. Chihara G, Hamuro J, Maeda YY, Shiio T, Suga T, Takasuka N, Sasaki T. Antitumor and metastasis-inhibitory activities of lentinan as an immunomodulator: an overview. Cancer Detect Prev Suppl. 1987; 1: 423-443.
6. Ren L, Perera C, Hemar Y. Antitumor activity of mushroom polysaccharides: a review. Food Funct. 2012; 3: 1118-1130.
7. Rachel S, Gordon AM. The anti-diabetic potential of polysaccharides extracted from members of the cucurbit family: a review. Bioact Carbohydr Dietary Fibre. 2014; 3: 106-114.
8. Aizhen Z, Hongzhi C, Fengshan W. Anticancer polysaccharides from natural resources: a review of recent research. Carbohydr Polym. 2012; 90: 1395-410.
9. Yangyang Z, Sheng L, Xiaohua W, Lina Z, Peter CK. Advances in lentinan: isolation, structure, chain conformation and bioactivities. Food Hydrocoll. 2011; 25: 196-206.
10. Higashi D, Seki K, Ishibashi Y, Egawa Y, Koga M, Sasaki T, Hirano K, Mikami K, Futami K, Maekawa T, Sudo M. The effect of lentinan combination therapy for unresectable advanced gastric cancer. Anticancer Res. 2012; 32: 2365-2368.
11. Hazama S, Watanabe S, Ohashi M, Yagi M, Suzuki M, Matsuda K, Yamamoto T, Suga Y, Suga T, Nakazawa S, Oka M. Efficacy of orally administered superfine dispersed lentinan (β-1,3-glucan) for the treatment of advanced colorectal cancer. Anticancer Res. 2009; 29: 2611-2617.
12. Han X, Liu AJ, Zhao XH, Li YD, Zheng GQ, Zhang GR. Immunizing effects of co-cultures of H22 hepatocarcinoma cells and cartilage polysaccharide (CHCP) on murine H22 hepatocarcinoma. J Food Sci. 2010; 75: H265-H273.
13. Kupfahl C, Geginat G, Hof H. Lentinan has a stimulatory effect on innate and adaptive immunity against murine Listeria monocytogenes infection. Int Immunopharmacol. 2006; 6: 686-696.
14. Xiaofei X, Huidan Y, Jian T, Jian C, Xuewu Z. Polysaccharides in Lentinus edodes: isolation, structure, immunomodulating activity and future prospective. Crit Rev Food Sci. 2014; 54: 474-487.
15. Guo Z, Hu Y, Wang D, Ma X, Zhao X, Zhao B, Wang J, Liu P. Sulfated modification can enhance the adjuvanticity of lentinan and improve the immune effect of ND vaccine. Vaccine. 2009; 27: 660-665.
16. Pan H, Han Y, Huang J, Yu X, Jiao C, Yang X, Dhaliwal P, Xie Y, Yang BB. Purification and identification of a polysaccharide from medicinal mushroom Amauroderma rude with immunomodulatory activity and inhibitory effect on tumor growth. Oncotarget. 2015; 6: 17777-17791. http://dx.doi.org/10.18632/oncotarget.4397.
17. Anjun L, Xue H, Guorong Z, Xihua Z. Effects of cartilage polysaccharide on apoptosis and immune of human hepatoma BEL-7402 cells and murine H22 hepatocarcinoma. Int J Food Sci Nutr. 2009; 60: 47-58.
18. Wang J, Dong S, Liu C, Wang W, Sun S, Gu J, Wang Y, Boraschi D, Qu D. β-Glucan oligosaccharide enhances CD8(+) T cells immune response induced by a DNA vaccine encoding hepatitis B virus core antigen. J Biomed Biotechnol. 2010; 1: 645213.
19. Zhu Y, Li X, Chen C, Wang F, Li J, Hu C, Li Y, Miao L. Effects of aluminum trichloride on the trace elements and cytokines in the spleen of rats. Food Chem Toxicol. 2012; 50: 2911-2915.
20. Jia X, Liu Q, Zou S, Xu X, Zhang L. Construction of selenium nanoparticles/β-glucan composites for enhancement of the antitumor activity. Carbohydr Polym. 2015; 117: 434-442.
21. Li Pira G, Starc N, Conforti A, Bertaina A, Rutella S, Locatelli F, Manca F. Lymphocyte proliferation specific for recall, CMV and HIV antigens in miniaturized and automated format. J Immunol Methods. 2012; 384: 135-142.
22. Katsunada F, Masamichi T, Shigenori G. A case of recurrent ovarian cancer successfully treated with adoptive immunotherapy and lentinan. Anticancer Res. 2006; 26: 4015-4018.
23. Chernysheva AD, Kirou KA, Crow MK. T cell proliferation induced by autologous non-T cells is a response to apoptotic cells processed by dendritic cells. J Immunol. 2002; 169: 1241-1250.
24. Oba K, Kobayashi M, Matusi T, Kodera Y, Sakamoto J. Individual patient data meta-analysis of the immunochemotherapy using lentinan in patients with advanced gastric cancer. Anticancer Res. 2009; 29: 2739-2745.
25. Renato O, Franz K, Peter JM, Gioacchino N. Macrophages and cancer: from mechanisms to therapeutic implications. Trend Immunol. 2015; 36: 229-239.
26. Isoda N, Eguchi Y, Nukaya H, Hosho K, Suga Y, Suga T, Nakazawa S, Sugano K. Clinical efficacy of superfine dispersed lentinan (beta-1,3-glucan) in patients with hepatocellular carcinoma. Hepatogastroenterology. 2009; 56: 437-441.
27. Nishitani Y, Zhang L, Yoshida M, Azuma T, Kanazawa K, Hashimoto T, Mizuno M. Intestinal anti-inflammatory activity of lentinan: influence on IL-8 and TNFR1 expression in intestinal epithelial cells. PLoS One. 2013; 8: e62441.
28. Lili J, Debin W, Xiaoyu Z, Bo L, Huanxi Z, Shuying L. Characterization and immuno stimulating effects on murine peritoneal macrophages of oligosaccharide isolated from Panax ginseng C.A. Meyer. J Ethnopharmacol. 2012; 144: 490-496.
29. Chen A, Kaganovsky E, Rahimipour S, Ben-Aroya N, Okon E, Koch Y. Two forms of gonadotropin-releasing hormone (GnRH) are expressed in human breast tissue and overexpressed in breast cancer: a putative mechanism for the antiproliferative effect of GnRH by down-regulation of acidic ribosomal phosphoproteins P1 and P2. Cancer Res. 2002; 62: 1036-1044.
30. Sengelaub CA, Navrazhina K, Ross JB, Halberg N, Tavazoie SF. PTPRN2 and PLCβ1 promote metastatic breast cancer cell migration through PI(4,5)P2-dependent actin remodeling. EMBO J. 2016; 35: 62-76.
31. Takai E, Tsukimoto M, Harada H, Sawada K, Moriyama Y, Kojima S. Autocrine regulation of TGF-β1-induced cell migration by exocytosis of ATP and activation of P2 receptors in human lung cancer cells. J Cell Sci. 2012; 125: 5051-5060.
32. Suenaga S, Kuramitsu Y, Wang Y, Baron B, Kitagawa T, Akada J, Tokuda K, Kaino S, Maehara S, Maehara Y, Sakaida I, Nakamura K. Human pancreatic cancer cells with acquired gemcitabine resistance exhibit significant up-regulation of peroxiredoxin-2 compared to sensitive parental cells. Anticancer Res. 2013; 33: 4821-4826.
33. Stresing V, Baltziskueta E, Rubio N, Blanco J, Arriba MC, Valls J, Janier M, Clézardin P, Sanz-Pamplona R, Nieva C, Marro M, Petrov D, Sierra A. Peroxiredoxin 2 specifically regulates the oxidative and metabolic stress response of human metastatic breast cancer cells in lungs. Oncogene. 2013; 32: 724-735.
34. Lu W, Fu Z, Wang H, Feng J, Wei J, Guo J. Peroxiredoxin 2 is upregulated in colorectal cancer and contributes to colorectal cancer cells’ survival by protecting cells from oxidative stress. Mol Cell Biochem. 2014; 387: 261-270.
35. Wehder L, Arndt S, Murzik U, Bosserhoff AK, Kob R, von Eggeling F, Melle C. Annexin A5 is involved in migration and invasion of oral carcinoma. Cell Cycle. 2009; 8: 1552-1558.
36. Xue G, Hao LQ, Ding FX, Mei Q, Huang JJ, Fu CG, Yan HL, Sun SH. Expression of annexin a5 is associated with higher tumor stage and poor prognosis in colorectal adenocarcinomas. J Clin Gastroenterol. 2009; 43: 831-837.
37. Liu Z, Zhan Y, Tu Y, Chen K, Liu Z, Wu C. PDZ and LIM domain protein 1(PDLIM1)/CLP36 promotes breast cancer cell migration, invasion and metastasis through interaction with α-actinin. Oncogene. 2015; 34: 1300-1311.
38. Wu H, Reynolds AB, Kanner SB, Vines RR, Parsons JT. Identification and characterization of a novel cytoskeleton-associated pp60src substrate. Mol Cell Biol. 1991; 11: 5113-5124.
39. Weaver AM. Cortactin in tumor invasiveness. Cancer Lett. 2008; 265: 157-166.
40. Pei XJ, Xue XF, Zhu YL, Liu SJ, Han AJ, Yang QX. Expression and clinical significance of moesin and E-cadherin in invasive carcinoma of breast, no specific type. Zhonghua Bing Li Xue Za Zhi. [Article in Chinese]. 2016; 45: 550-5.
41. Wu M, Liu DY, Yuan XR, Liu Q, Jiang XJ, Yuan D, Huang J, Li XJ, Yang ZQ. The expression of moesin in astrocytoma: correlation with pathologic grade and poor clinical outcome. Med Oncol. 2013; 30: 372.