
INTRODUCTION
Ovarian cancer is one of the most common cancer in the world [1]. During the development of new therapies and treatments, the five-year survival rate of patients had been increased. However, because patients were always diagnosed at an advanced-stage, ovarian cancer still had a high mortality rate [2]. Understanding the detail mechanism of ovarian cancer is helpful for us to find the therapy target.
Forkhead box (FOX) family has been found to play important function in several cell process, including embryonic development [3] and organogenesis [4]. Moreover, there are many reports indicate that FOX family regulates physiological processes [3, 5], including cell cycle [6], cell signaling [7] and metabolic processes [8]. Therefore, abnormal expression of FOX family protein results in cancer development [9-11]. Forkhead box k1 (FOXK1) belongs to FOX family, as a transcription factor, it recognizes and binds DNA consensus sequence, WRTAAAAYA, to regulate transcription [12, 13]. Previous studies indicate that FOXK1 regulates c-myc, p21 and cdc2 gene in mice [14]. Moreover, there are also many reports demonstrate that FOXK1 takes part in tumorigenesis [15, 16]. In addition, FOXK1 inhibition suppresses cell proliferation in human osteosarcoma cancer cells [17]. However, the role of FOXK1 in ovarian cancer is still unknown.
Cell proliferation is a complex cell program, it is regulated by numerous cell process, including cell cycle, cell apoptosis and so on. p21, also known as p21WAF1/CIP1, is mediated by p53. p21 plays a key function in G1 growth arrest [18, 19]. p21 regulates cell cycle arrest in response to multiple stimuli and there by promoting DNA repair.
Here, our work demonstrates that FOXK1 is significantly high expression in ovarian cancer tissues and cell lines. Moreover, high expression of FOXK1 predicts poor prognosis in ovarian cancer patients. In addition, FOXK1 promotes the proliferation through transcriptionally regulating p21. FOXK1 also facilitates metastasis in ovarian cancer cells. Our work firstly find the effect of FOXK1 on cell proliferation and metastasis in the ovarian cancer, and explain the molecular mechanism of FOXK1 in cell proliferation.
RESULTS
FOXK1 is up-regulated in ovarian cancer and the expression of FOXK1 is corrected with poor prognosis of ovarian cancer
To analyze the function of FOXK1 in ovarian cancer, we first detected the expression of FOXK1 in different ovarian cancer cell lines, SKOV3 and OVCA429, human normal ovarian cell lines IOSE80 was used as a control group. The results of qRT-PCR and western blotting indicated FOXK1 was high expression in ovarian cancer cell lines, SKOV3 and OVCA429, compared with that in IOSE80 cells (Figure 1A and 1B). Subsequently, we collected 87 pairs’ ovarian cancer tissue samples and adjacent normal tissue samples from patients who were diagnosed as ovarian cancer in Affiliated Tumor Hospital of Guangxi Medical University during 2015-2016, then qRT-PCR and western blotting analysis were performed to determine the expression of FOXK1. Interestingly, we found that FOXK1 was up-regulated in ovarian cancer samples (Figure 1C and 1D). Moreover, we found high expression of FOXK1 was correlated with multiple clinic pathologic in ovarian cancer, including tumor size, pathological stage and metastasis. However, there were no obviously correlation with age and differentiation. Furthermore, we analyzed the relationship between FOXK1 and survival curve in ovarian cancer. The result suggested that the patients who had high expression of FOXK1 had shorter survival time than the patients who had low expression of FOXK1 (Figure 1E, P=0.037). In conclusion, we hypothesize that FOXK1 plays an important function in ovarian cancer.
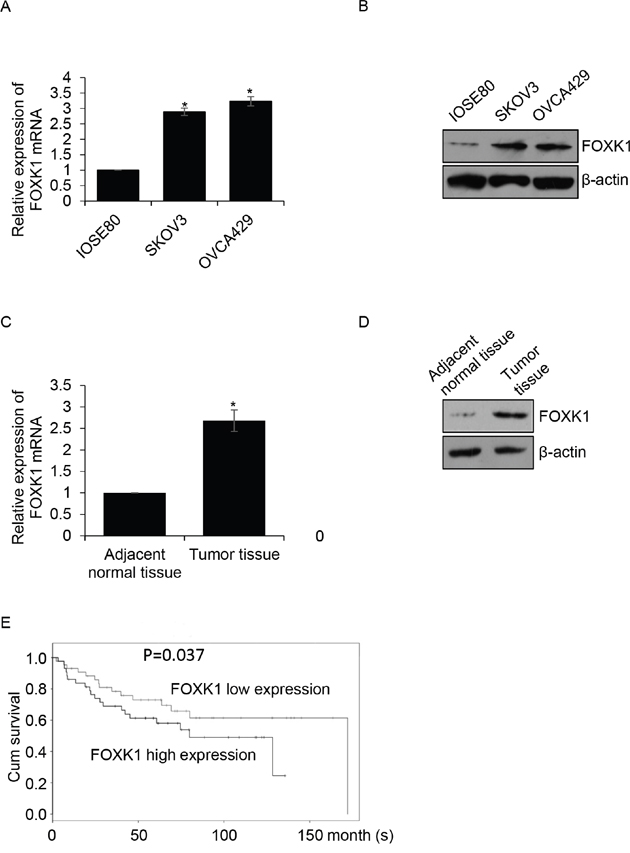
Figure 1: FOXK1 is up-regulated in ovarian cancer and the expression of FOXK1 is corrected with poor prognosis of ovarian cancer (A). Total RNA was prepared from IOSE80, SKOV3 and OVCA429 cells, qRT-PCR was performed to detect the expression of FOXK1. (B). Whole protein was prepared from IOSE80, SKOV3 and OVCA429 cells, western blotting was performed to detect the expression of FOXK1. (C). Total RNA was prepared from 87 pairs’ tumor tissue samples and adjacent normal tissue samples, qRT-PCR was performed to detect the expression of FOXK1. (D). Western blotting was used to detect the expression of FOXK1 in tumor tissue samples and adjacent normal tissue samples. (E). The relationship between FOXK1 expression and survival curve was analyzed by Kaplan Meier method.
Overexpression of FOXK1 facilitates growth of ovarian cancer cells
In order to investigate the function of FOXK1 in ovarian cancer, we overexpressed or knocked down FOXK1 in SKOV3 or OVCA429 cells, respectively. Next, western blotting and qRT-PCR analysis were performed to determine the expression of FOXK1 in SKOV3 or OVCA429 cells. The results revealed that the expression of FOXK1 was obviously increased while cells were transfected with FLAG-FOXK1, compared with that of control group. In addition, the expression of FOXK1 was significantly decreased while cells were transfected with FOXK1 siRNA, compared with that of scramble siRNA (SCR) group. Moreover, we found siFOXK1-1 was more efficiency than siFOXK1-2, so siFOXK1-1 was used for the further experiments (Figure 2A and 2B). Because of the correlation between the expression of FOXK1 and tumor size, we assumed whether FOXK1 promoted ovarian cancer cells proliferation. Next, CCK-8 assay was used to detect the effect of FOXK1 on cell growth. The results suggested that SKOV3 cells transfected with FOXK1 grew faster than those transfected with vector (Figure 2C). On the contrary, the FOXK1-depleted cells grew slower than those transfected with SCR. The similar results were observed in OVCA429 cells (Figure 2C). Next, colony formation analysis was performed to determine the effect of FOXK1 on cell proliferation. The results showed that FOXK1 remarkably increased the number of colonies, on the contrary, FOXK1 inhibition decreased the number of colonies (Figure 2D). In conclusion, FOXK1 facilitates ovarian cancer cell proliferation.
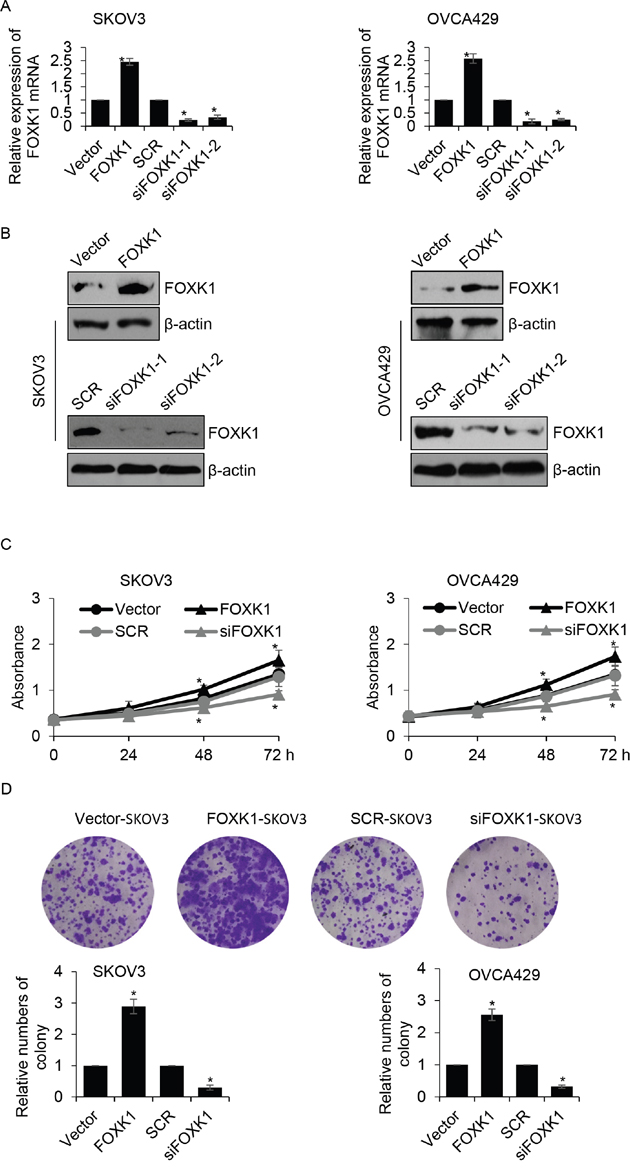
Figure 2: Overexpression of FOXK1 facilitates growth of ovarian caner cells. (A). SKOV3 and OVCA429 cells were transfected with empty plasmid (vector), FLAG-FOXK1, scramble siRNA (SCR), siFOXK1, respectively. The mRNA level of FOXK1 was determined by qRT-PCR. (B). FOXK1 was overexpressed or silenced in SKOV3 and OVCA429 cells, respectively. The protein level of FOXK1 was determined by western blotting. (C). FOXK1 was overexpressed or silenced in SKOV3 and OVCA429 cells, respectively. CCK-8 analysis was used to detect the effect of FOXK1 on cell proliferation. (D). FOXK1 was overexpressed or silenced in SKOV3 and OVCA429 cells, respectively. Colony formation analysis was used to detect the effect of FOXK1 on cell proliferation.
FOXK1 promotes G1/S phase transition, but has no effect on cell apoptosis
Subsequently, we further decipher the function of FOXK1 on cell cycle. Flow cytometry analysis indicated that overexpression of FOXK1 markedly improved the percentage of cells in the S phase, but the number of cells in G0/G1 phase was decreased. On the contrary, inhibition of FOXK1 strongly decreased the percentage of cells in S phase, but the number of cells in G0/G1 phase was increased (Figure 3A). Cell apoptosis also decrease cell proliferation, so we next detected if FOXK1 influenced cell apoptosis. The results demonstrated that FOXK1 had no effect on cell apoptosis (Figure 3B). Caspase-3 and caspase-8 as cell apoptosis markers, next, we detect whether they were activated or inhibited by FOXK1. As shown in Figure 3C and 3D, the mRNA and protein levels of caspase-3 and caspase-8 were not activated or inhibited by FOXK1. Together, FOXK1 facilitates cell proliferation through promoting G1/S phase transition.
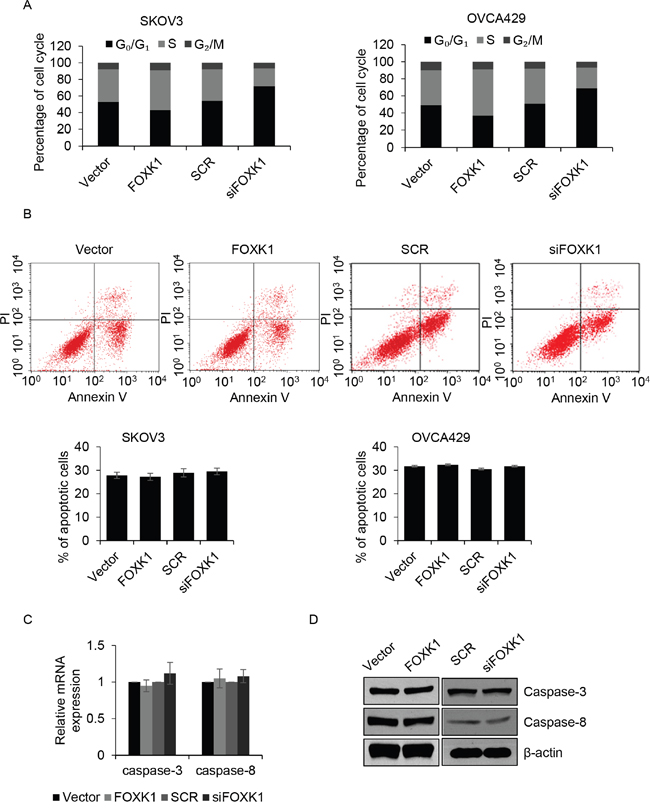
Figure 3: FOXK1 promotes G1/S transition, but has no effect on cell apoptosis. (A). SKOV3 and OVCA429 cells were transfected with empty plasmid (vector), FLAG-FOXK1, scramble siRNA (SCR), siFOXK1, respectively. Cell cycle was assessed by flow cytometer. (B). FOXK1 was overexpressed or silenced in SKOV3 and OVCA429 cells, respectively. After incubation with CPT for 6 h, numbers of apoptotic cell was detected by flow cytometer. (C). FOXK1 was overexpressed or silenced in SKOV3 cells, respectively. The mRNA level of caspase-3 and caspase-8 were detected by qRT-PCR. (D). FOXK1 was overexpressed or silenced in SKOV3 cells, respectively. The protein level of caspase-3 and caspase-8 were detected by western blotting.
FOXK1 improves the migration and invasion ability of ovarian cancer cells
We found that the expression of FOXK1 was correlated with metastasis, so we hypothesized that FOXK1 incresed cancer cell metastasis. To verify our hypothesis, we first performed transwell analysis, the result indicated that ectopic expression of FOXK1 obviously promoted cell invasion, the mean invaded cells were 63 ± 7 and 124 ± 8 for vector-or FOXK1-transfected OVCA429 cells, respectively (Figure 4A). However, FOXK1 inhibition dramatically increased cell invasion, the mean relative migration distance were 66 ± 4 and 23 ± 3 for SCR-or siFOXK-transfected OVCA429 cells, respectively (Figure 4A). Subsequently, wound healing analysis was performed to detect the effect of FOXK1 on cell migration. We found ectopic expression of FOXK1 enhanced the migration ability of SKOV3 and OVCA429 cells, the mean relative migration distance were increased when FOXK1 was overexpressed in SKOV3 cells (Figure 4B). Whereas, FOXK1 inhibition suppressed the migration ability of SKOV3 cells (Figure 4B). The similar results were observed in OVCA429 cells (Figure 4B). MMP-9, a marker of cell invasion, has been found to increase tumor growth and metastasis in ovarian cancer [20-22]. We next detected whether FOXK1 increased MMP-9 expression. Both in SKOV3 and OVCA429 cells, the expression of MMP-9 were up-regulated by FOXK1 (Figure 4C and 4D). Meanwhile, the expression of MMP-9 was decreased when FOXK1 was knocked down (Figure 4C and 4D). In order to investigate whether FOXK1 transcriptionally increased MMP-9, we performed ChIP assay and luciferase reporter assay, the results indicated that FOXK1 didn’t transcriptionally increased MMP-9. FOXK1 might increase MMP-9 expression through regulation of any other proteins.
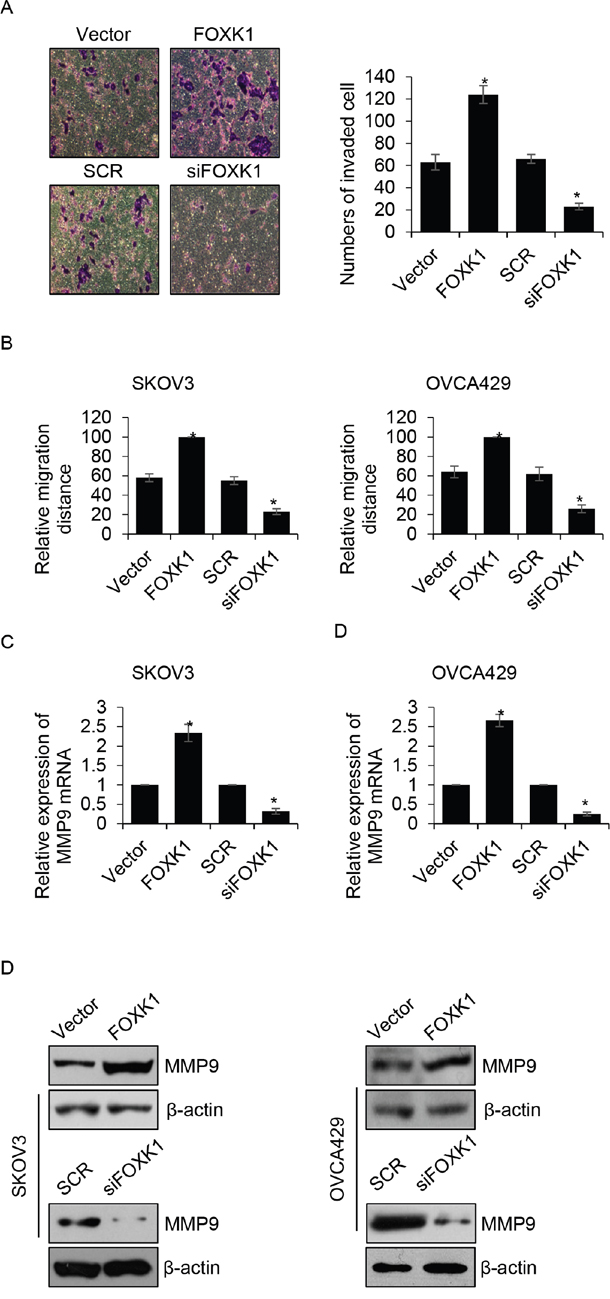
Figure 4: FOXK1 improves the migration and invasion ability of ovarian cancer cells. (A). OVCA429 cells were transfected with empty vector, FLAG-FOXK1, scramble siRNA (SCR), siFOXK1, respectively. Transwell invasion assay was used to investigate the effect of FOXK1 on cancer cell invasion. (B). FOXK1 was overexpressed or silenced in SKOV3 and OVCA429 cells, respectively. After scratch by peptide, relative migration distance was measured at different time points. (C). FOXK1 was overexpressed or silenced in SKOV3 and OVCA429 cells, respectively. The mRNA level of MMP-9 was detected by qRT-PCR. (D). FOXK1 was overexpressed or silenced in SKOV3 and OVCA429 cells, respectively. The protein level of MMP-9 was detected by western blotting.
FOXK1 transcriptionally inhibits p21 in ovarian cancer cells
Cell cycle was regulated by numerous protein, including promoting cell cycle protein, such as cyclin D1 and cyclin E1, and inhibiting cell cycle protein, such as p16 and p21. To determine whether the effect of FOXK1 on G1/S phase transition through regulating the expression of these protein, we overexpressed or knocked down FOXK1 in SKOV3 cells, and performed Western blotting to detect the expression of those protein. The result revealed the expression of p21 was obviously suppressed when FOXK1 was overexpressed; however, FOXK1 inhibition promoted the expression of p21. But there were little effect on the expression of cyclin D1, cyclin E1 and p16 (Figure 5A). Furthermore, qRT-PCR suggested the mRNA level of p21 was also increased by FOXK1 (Figure 5B). Above results indicated that p21 might be transcriptionally increased by FOXK1. Subsequently, we performed ChIP analysis, the results showed FOXK1 interacted with the promotor region of p21, but not the promotor region of CCND1, CCNE1 and p16 (Figure 5C). Next, we sub-cloned the promotor region of p16, p21, CCND1 and CCNE1 into pGL4 plasmid. Luciferase reporter assay was performed to determine whether FOXK1 transcriptionally increased p16, p21, CCND1 and CCNE1. Consisted with our previous work, we found only p21 was transcriptionally inhibited by FOXK1 (Figure 5D), but FOXK1 had no effect on p21, CCND1 and CCNE1 (data not shown). In conclusion, FOXK1 promotes cell proliferation through transcriptionally inhibits p21.
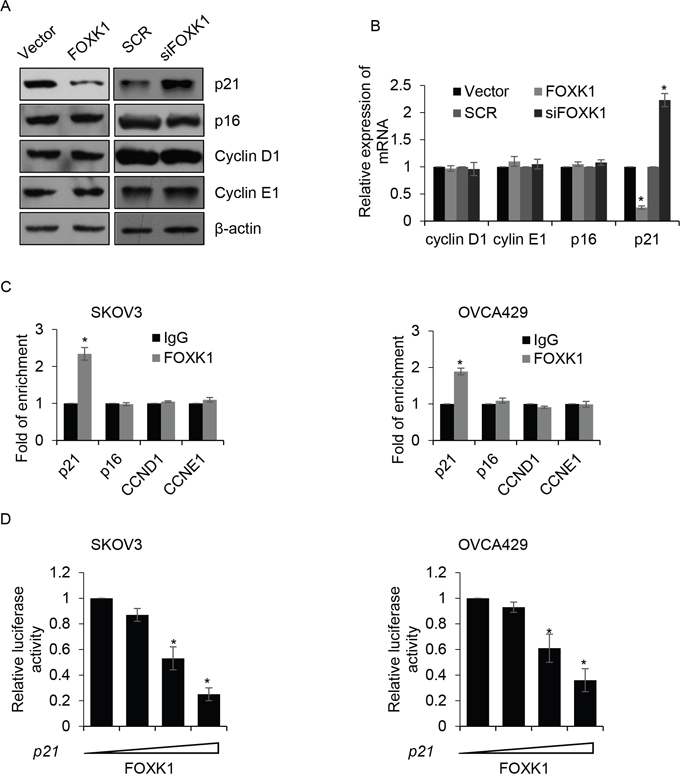
Figure 5: FOXK1 transcriptionally inhibits p21 in ovarian cancer cells. (A). SKOV3 cells were transfected with empty plasmid (vector), FLAG-FOXK1, scramble siRNA (SCR), siFOXK1, respectively. The protein levels of cyclin D1, cyclin E1, p16 and p21 were detected by western blotting. (B). FOXK1 was overexpressed or silenced in SKOV3 cells, respectively. The mRNA levels of cyclin D1, cyclin E1, p16 and p21 were detected by qRT-PCR. (C). ChIP assay was performed to detect whether FOXK1 bound the promoter region of indicated gene in SKOV3 and OVCA429 cells. (D). SKOV3 and OVCA429 cells were co-transfected with FOXK1, Renilla, pGL4-p21. 24 h after transfection, luciferase reporter assay was performed.
DISCUSSION
The members of FOX transcription factor family have different function during embryogenesis [23-25]. There are several reports indicate that FOX protein plays essential role in established cancers [9-11]. For instance, FOXA1 is high expression in multiple cancers, including pancreatic cancer [26], glioma cancer [27], breast cancer [28], prostate cancer [29] and bladder cancer [30]. Moreover, FOXK1 has been found to high express in multiple cancers, such as colorectal carcinoma [31].
In this study, we used ovarian tissue samples to explore FOXK1 expression profiles in adjacent normal tissues and cancer tissues. We found FOXK1 was high expression in cancer tissues, meanwhile, FOXK1 was up-regulated in ovarian cancer cell lines, so FOXK1 might act as an oncogene. In addition, survival curve analysis demonstrated a dramatically worse overall survival for ovarian cancer patients who had high expression of FOXK1, indicating that expression of FOXK1 was correlated with poor prognosis in ovarian cancer.
Recently, there were several researches revealed that FOXK1 facilitated proliferation, invasion and metastasis in different cancer [31-33]. Wang et al. found that FOXK1 and FOXK2 promote DVL translocating into nucleus, thereby regulation of the Wnt/β-catenin signaling [16]. However, the function of FOXK1 in ovarian cancer remains unknown. Consistently, our works have shown that FOXK1 promoted cell proliferation and invasion in ovarian cancer. These findings revealed that abnormal expression of FOXK1 might be an essential mechanism underlying cancer proliferation and metastasis.
FOXK1, a transcription factor, was identified as a crucial regulator for myogenic stem cell proliferation [12, 34, 35]. We identified that p21 as a downstream target of FOXK1, FOXK1 promoted G1/S phase transition through transcriptionally inhibiting p21. However, FOXK1 had no effect on cell apoptosis.
The detail mechanism of FOXK1 in metastasis remains unclear. EMT is a complex process which is widespread in malignant tumor. The main character of EMT is gain of mesenchymal character and loss of epithelial character, resulting epithelial cells transformed into mesenchymal cells. Recently, several members of the FOX family have been found to regulate EMT. FOXQ1 facilitated metastasis in colorectal carcinoma cells which had undergone TGF-β-induced EMT [36]. Moreover, FOXC2 was high expression in invasive ovarian cancer tissues and cell lines [37]. Furthermore, FOXC2 maintained TGF-β1-induced EMT. Recently report indicates that FOXK1 regulates EMT in gastric cancer (GC) [32]. So we assumed that FOXK1 might promote metastasis through regulation of EMT in ovarian cancer.
In conclusion, our work firstly find that FOXK1 is up-regulated in ovarian cancer tissues and cells. Moreover, we observe that FOXK1 has a relationship with poor prognosis. In addition, we find that FOXK1 promotes G1/S phase transition through transcriptionally inhibiting p21. Furthermore, FOXK1 facilitates invasion in ovarian cancer cell. Together, our study explain the function of FOXK1 in ovarian cancer and indicated that FOXK1 plays an essential function in mediating ovarian caner progression, and serves as a therapeutic target for ovarian cancer.
MATERIALS AND METHODS
Cell culture and ovarian tissue
Human ovarian cancer cell lines, SKOV3 and OVCA429 and human normal ovarian cell lines IOSE80 were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). Cells were maintained in DMEM (HyClone, Logan, UT, USA) supplemented with 1% Penicillin-Streptomycin solution (HyClone, Logan, UT, USA) and 10% FBS (HyClone, Logan, UT, USA) at 37°C with 5% CO2. For ovarian tissue sample experiments, the patients had known our experiments before we collected their tissue samples. Our tissue sample experiments were approved by Ethics Committee of the Affiliated Tumor Hospital of Guangxi Medical University.
Cell transfection
Cells were transfected with scramble siRNA (SCR) or FOXK1 siRNA (siFOXK1) at density of 30%-40% by Lipofectamine RNAiMax reagent (Thermo Fisher Scientific, Waltham, MA, USA). 48 h after transfection, cells were collected and used for the further experiments.
Western blotting
Cell lysate was prepared by RIPA lysis buffer (Beyotime, Jiangsu, People’s Republic of China) supplemented with cocktail, a protease inhibitor (Thermo Fisher Scientific, Waltham, MA, USA). Equal amount of proteins (45 μg) were separated on 10% SDS-PAGE, then proteins were transferred onto nitrocellulose filter membranes. 5% non-fat milk solution was used to block nonspecific antigen on the membranes, and the membranes were washed by PBST solution for three times. Subsequently, specific antibodies were used to incubate membranes at 4°C overnight. Next, the membranes were washed with PBST for three times and incubated with second antibody (1:5000; Sigma, USA) at room temperature for 1 h. The blot of specific protein was visualized by ECL (Millipore, Bedford, MA, USA).
qRT-PCR
Total RNA was prepared from cells by TRIzol reagent (Invitrogen, Carlsbad, CA, USA), and then RNA were reverse transcribed into cDNA by PrimeScript® 1st strand cDNA synthesis kit (TaKaRa, Tokyo, Japan). SYBR® Green Real-time PCR Master Mix (Sigma, USA) was used to determine the mRNA level of indicated gene by 7900HT qRT-PCR system. GAPDH was served as an internal control. Relative mRNA levels of indicated gene were analyzed using the 2-ΔΔCtmethod.
Cell cycle assay
Cell cycle assay was used to detect whether FOXK1 regulated cell cycle. In brief, FOXK1 was overexpressed or silenced in SKOV3 and OVCA429 cells, next, cells were collected, follow by fixed with ice-cold 70% ethanol for 20 min and stained with propidium iodide (PI, Transgene, Beijing, People’s Republic of China) at room temperature for 20 min. Cell cycle was determined through flow cytometry (FACS Calibur, BD, San Diego, CA, USA).
Cell apoptosis assay
Cell apoptosis assay was performed to assess the effect of FOXK1 on cell apoptosis. Briefly, FOXK1 was overexpressed or silenced in SKOV3 and OVCA429 cells, subsequently, cells were collected and resuspended in 100 μl binding buffer. Next, 5 μl annexin V-FITC and 5 μl PI (Jingmei Biotech, Shenzhen, People’s Republic of China) were added in cell solution, the cells were incubated at room temperature for 15 min and then cell apoptosis was analyzed by flow cytometry (FACS Calibur, BD, San Diego, CA, USA).
Wound healing assay
Wound healing assay was performed to assess the effect of FOXK1 on cell migration. Briefly, SKOV3 and OVCA429 cells were transfected with vector, FOXK1, SCR, FOXK1 siRNA, respecticly. When the cells density reached 90%-100%, a 20 μl pipette was used to create a scratch. Meanwhile, cells were incubated with serum-free DMEM. Scratches were observed under a microscope. The distance of scratch was measured at different time points (0 h and 24 h) by Image Pro-Plus 6.0 software (Media Cybernetics, USA).
Transwell invasion assay
Transwell invasion assay was performed to assess the effect of FOXK1 on cell invasion. Transwell chamber (8 μm pore size, Corning Costar, Corning, NY, USA) was used in this experiments. After transfection, 10,000 cells were resuspended in serum-free DMEM and placed into the upper well which coated with 100 μl Matrigel (BD, San Diego, CA, USA). Simultaneously, the lower well of the chamber was filled with DMEM containing 10% FBS. Cells were maintained at 37°C for 24 h. Subsequently, cells on the upper surface of the chambers were removed by cotton swab, the cells on the lower surface of the chambers were stained with 0.1% crystal violet and counted under a microscope.
CCK-8 analysis
CCK-8 analysis was used to determine the function of FOXK1 on cell proliferation. SKOV3 and OVCA429 cells were transfected with vector, FOXK1, SCR, siFOXK1 using Lipo 2000 reagent (Thermo Fisher Scientific, Waltham, MA, USA), respectively. 48 h after transfection, approximately 4,000 cells were placed into each well of 96-well plates. CCK-8 solution was added at 0 h, 24 h, 48 h and 72 h after placing. After adding 10 μl CCK-8 solution, cells were incubated for 1 h at 37 °C. The absorbance was measured at 450 nm.
Statistical analysis
The data were analyzed by Statistical Product and Service Solutions 19.0 (SPSS, IBM, USA). The comparison between two group was analyzed by Student’s t-test. The data was represented as mean ± SD. The p < 0.05 was considered as statistical significant (*p < 0.05).
CONFLICTS OF INTEREST
The authors declare that they have no competing interests.
REFERENCES
1. Lengyel E. Ovarian cancer development and metastasis. Am J Pathol. 2010; 177:1053-1064.
2. Banks E. The epidemiology of ovarian cancer. Methods Mol Med. 2001; 39:3-11.
3. Foucher I, Volovitch M, Frain M, Kim JJ, Souberbielle JC, Gan L, Unterman TG, Prochiantz A, Trembleau A. Hoxa5 overexpression correlates with IGFBP1 upregulation and postnatal dwarfism: evidence for an interaction between Hoxa5 and Forkhead box transcription factors. Development. 2002; 129:4065-4074.
4. Gaudet J, Mango SE. Regulation of organogenesis by the Caenorhabditis elegans FoxA protein PHA-4. Science. 2002; 295:821-825.
5. Wang X, Quail E, Hung NJ, Tan Y, Ye H, Costa RH. Increased levels of forkhead box M1B transcription factor in transgenic mouse hepatocytes prevent age-related proliferation defects in regenerating liver. Proc Natl Acad Sci U S A. 2001; 98:11468-11473.
6. Williamson EA, Wolf I, O'Kelly J, Bose S, Tanosaki S, Koeffler HP. BRCA1 and FOXA1 proteins coregulate the expression of the cell cycle-dependent kinase inhibitor p27(Kip1). Oncogene. 2006; 25:1391-1399.
7. Gao N, Ishii K, Mirosevich J, Kuwajima S, Oppenheimer SR, Roberts RL, Jiang M, Yu X, Shappell SB, Caprioli RM, Stoffel M, Hayward SW, Matusik RJ. Forkhead box A1 regulates prostate ductal morphogenesis and promotes epithelial cell maturation. Development. 2005; 132:3431-3443.
8. Clevidence DE, Overdier DG, Tao W, Qian X, Pani L, Lai E, Costa RH. Identification of nine tissue-specific transcription factors of the hepatocyte nuclear factor 3/forkhead DNA-binding-domain family. Proc Natl Acad Sci U S A. 1993; 90:3948-3952.
9. Teh MT, Wong ST, Neill GW, Ghali LR, Philpott MP, Quinn AG. FOXM1 is a downstream target of Gli1 in basal cell carcinomas. Cancer Res. 2002; 62:4773-4780.
10. Coffer PJ, Burgering BM. Forkhead-box transcription factors and their role in the immune system. Nat Rev Immunol. 2004; 4:889-899.
11. Muller SM, Terszowski G, Blum C, Haller C, Anquez V, Kuschert S, Carmeliet P, Augustin HG, Rodewald HR. Gene targeting of VEGF-A in thymus epithelium disrupts thymus blood vessel architecture. Proc Natl Acad Sci U S A. 2005; 102:10587-10592.
12. Hawke TJ, Jiang N, Garry DJ. Absence of p21CIP rescues myogenic progenitor cell proliferative and regenerative capacity in Foxk1 null mice. J Biol Chem. 2003; 278:4015-4020.
13. Shi X, Seldin DC, Garry DJ. Foxk1 recruits the Sds3 complex and represses gene expression in myogenic progenitors. Biochem J. 2012; 446:349-357.
14. Garry DJ, Meeson A, Elterman J, Zhao Y, Yang P, Bassel-Duby R, Williams RS. Myogenic stem cell function is impaired in mice lacking the forkhead/winged helix protein MNF. Proc Natl Acad Sci U S A. 2000; 97:5416-5421.
15. Huang JT, Lee V. Identification and characterization of a novel human FOXK1 gene in silico. Int J Oncol. 2004; 25:751-757.
16. Wang W, Li X, Lee M, Jun S, Aziz KE, Feng L, Tran MK, Li N, McCrea PD, Park JI, Chen J. FOXKs promote Wnt/beta-catenin signaling by translocating DVL into the nucleus. Dev Cell. 2015;32:707-718.
17. Yang Q, Kong Y, Rothermel B, Garry DJ, Bassel-Duby R, Williams RS. The winged-helix/forkhead protein myocyte nuclear factor beta (MNF-beta) forms a co-repressor complex with mammalian sin3B. Biochem J. 2000; 345 Pt 2:335-343.
18. Brugarolas J, Chandrasekaran C, Gordon JI, Beach D, Jacks T, Hannon GJ. Radiation-induced cell cycle arrest compromised by p21 deficiency. Nature. 1995; 377:552-557.
19. Deng C, Zhang P, Harper JW, Elledge SJ, Leder P. Mice lacking p21CIP1/WAF1 undergo normal development, but are defective in G1 checkpoint control. Cell. 1995; 82:675-684.
20. Desmeules P, Trudel D, Turcotte S, Sirois J, Plante M, Grégoire J, Renaud MC, Orain M, Têtu B, Bairati I. Prognostic significance of TIMP-2, MMP-2, and MMP-9 on high-grade serous ovarian carcinoma using digital image analysis. Hum Pathol. 2015; 46:739-745.
21. Guo F, Tian J, Cui M, Fang M, Yang L. Downregulation of matrix metalloproteinase 9 by small interfering RNA inhibits the tumor growth of ovarian epithelial carcinoma in vitro and in vivo. Mol MedRep. 2015; 12:753-759.
22. Zou M, Zhang X, Xu C. IL6-induced metastasis modulators p-STAT3, MMP-2 and MMP-9 are targets of 3,3'-diindolylmethane in ovarian cancer cells. Cell Oncol (Dordr). 2016; 39:47-57.
23. Katoh M, Katoh M. Human FOX gene family (Review). Int J Oncol. 2004; 25:1495-1500.
24. Yang Y, Hou H, Haller EM, Nicosia SV, Bai W. Suppression of FOXO1 activity by FHL2 through SIRT1-mediated deacetylation. EMBO J. 2005; 24:1021-1032.
25. Murakami H, Aiba H, Nakanishi M, Murakami-Tonami Y. Regulation of yeast forkhead transcription factors and FoxM1 by cyclin-dependent and polo-like kinases. Cell Cycle. 2010; 9:3233-3242.
26. Mirosevich J, Gao N, Gupta A, Shappell SB, Jove R, Matusik RJ. Expression and role of Foxa proteins in prostate cancer. Prostate. 2006; 66:1013-1028.
27. Reddy OL, Cates JM, Gellert LL, Crist HS, Yang Z, Yamashita H, Taylor JA 3rd, Smith JA Jr, Chang SS, Cookson MS, You C, Barocas DA, Grabowska MM, et al. Loss of FOXA1 drives sexually dimorphic changes in urothelial differentiation and is an independent predictor of poor prognosis in bladder cancer. Am J pathol. 2015; 185:1385-1395.
28. Jungert K, Buck A, von Wichert G, Adler G, König A, Buchholz M, Gress TM, Ellenrieder V. Sp1 is required for transforming growth factor-beta-induced mesenchymal transition and migration in pancreatic cancer cells. Cancer Res. 2007; 67:1563-1570.
29. Fan DM, Feng XS, Qi PW, Chen YW. Forkhead factor FOXQ1 promotes TGF-beta1 expression and induces epithelial-mesenchymal transition. Mol Cell Biochem. 2014; 397:179-186.
30. Badve S, Turbin D, Thorat MA, Morimiya A, Nielsen TO, Perou CM, Dunn S, Huntsman DG, Nakshatri H. FOXA1 expression in breast cancer--correlation with luminal subtype A and survival. Clin Cancer Res. 2007; 13:4415-4421.
31. Wu Y, Peng Y, Wu M, Zhang W, Zhang M, Xie R, Zhang P, Bai Y, Zhao J, Li A, Nan Q, Chen Y, Ren Y, et al. Oncogene FOXK1 enhances invasion of colorectal carcinoma by inducing epithelial-mesenchymal transition. Oncotarget. 2016; 7:51150-51162. https://doi.org/10.18632/oncotarget.9457.
32. Peng Y, Zhang P, Huang X, Yan Q, Wu M, Xie R, Wu Y, Zhang M, Nan Q, Zhao J, Li A, Xiong J, Ren Y, et al. Direct regulation of FOXK1 by C-jun promotes proliferation, invasion and metastasis in gastric cancer cells. Cell Death Dis. 2016; 7:e2480.
33. Wu M, Wang J, Tang W, Zhan X, Li Y, Peng Y, Huang X, Bai Y, Zhao J, Li A, Chen C, Chen Y, Peng H, et al. FOXK1 interaction with FHL2 promotes proliferation, invasion and metastasis in colorectal cancer. Oncogenesis. 2016; 5:e271.
34. Meeson AP, Shi X, Alexander MS, Williams RS, Allen RE, Jiang N, Adham IM, Goetsch SC, Hammer RE, Garry DJ. Sox15 and Fhl3 transcriptionally coactivate Foxk1 and regulate myogenic progenitor cells. EMBO J. 2007; 26:1902-1912.
35. Shi X, Garry DJ. Sin3 interacts with Foxk1 and regulates myogenic progenitors. MolCell Biochem. 2012; 366:251-258.
36. Zhang H, Meng F, Liu G, Zhang B, Zhu J, Wu F, Ethier SP, Miller F, Wu G. Forkhead transcription factor foxq1 promotes epithelial-mesenchymal transition and breast cancer metastasis. Cancer Res. 2011; 71:1292-1301.
37. Zhou Z, Zhang L, Xie B, Wang X, Yang X, Ding N, Zhang J, Liu Q, Tan G, Feng D, Sun LQ. FOXC2 promotes chemoresistance in nasopharyngeal carcinomas via induction of epithelial mesenchymal transition. Cancer Lett. 2015; 363:137-145.