
INTRODUCTION
Pancreatic ductal adenocarcinoma (PDAC) is a highly lethal malignancy, as the 5-year survival rate after diagnosis is only 5% [1, 2]. PDAC cells are extremely aggressive, and more than 50% of patients develop local recurrence or distant metastasis after curative resection [3, 4]. Recently developed molecularly targeted therapeutics have not shown beneficial effects in patients with PDAC [5, 6]. The molecular pathogenesis of the aggressive phenotype in PDAC remains unclear. Therefore, in order to improve disease outcomes in PDAC patients, it is necessary to define the molecular pathogenesis underlying the aggressiveness of PDAC using advanced genomic approaches.
microRNAs (miRNAs) belong to a family of short, non-coding RNAs transcribed from the human genome. Their discovery suggested new directions for the study of human cancer pathogenesis [7]. A unique characteristic of miRNAs is the ability of a single miRNA to tune a large number of RNA transcripts in human cells [8]. Thus, dysregulated miRNA expression can disrupt tightly controlled RNA networks and increase the aggressiveness of cancer cells. Numerous studies have indicated that miRNAs are aberrantly expressed in several cancers, including PDAC [9–12].
We have been identified anti-tumour miRNAs and their mediated novel cancer networks based on the miRNA expression signatures in various cancers [13–20]. Construction of miRNA signatures using clinical specimens is a first step towards identification of dysregulated miRNAs and their resultant clinical characteristics. Current RNA sequencing methods offer superior analytical approaches for studying the transcriptome. We recently showed that the passenger strand of miR-145-3p acted as an anti-tumour miRNA through regulating of several oncogenic genes in bladder cancer and lung cancer [21, 22]. Moreover, we showed that both strands of pre-miR-139 (miR-139-5p and miR-139-3p) targeted matrix metalloprotease 11 in bladder cancer [23]. The involvement of passenger strand miRNAs in the regulation of cellular processes is a novel concept in RNA research.
In this study, we analysed the miRNA expression signature of PDAC clinical specimens by RNA sequencing. Using the PDAC signature, we focused on 4 clustered miRNAs, miR-216a-5p, miR-216a-3p, miR-216b-5p and miR-216b-3p on human chromosome 2p16.1 and investigated their functional significance in PDAC pathogenesis. Transcription of all members of the miR-216 cluster was significantly reduced in PDAC specimens, and ectopic expression of these miRNAs suppressed cancer cell aggressiveness. We further investigated the target genes of miR-216. The resultant data provide novel insights into the molecular pathogenesis of PDAC.
RESULTS
Construction of expression signature and identification of downregulated miRNAs in PDAC
Libraries of small RNAs (from seven PDAC tissues and four normal pancreatic tissues) were analysed by using RNA sequencing technology. The patients’ characteristics are shown in Table 1. Initially, 16,930,437 to 24,733,299 raw sequence reads were produced for the libraries. After filtering for low quality and adaptors sequence reads, from 10,615,842 to 23,407,926 clean small RNA reads were obtained (Table 2). Differentially expressed miRNAs by RNA sequence were summarized in Supplementary Tables 1 and 2.
Table 1: Clinical features of PDAC patients
No. |
Age |
Sex |
T |
N |
M |
Stage |
Recurrence |
Remarks |
|---|---|---|---|---|---|---|---|---|
T1 |
44 |
M |
3 |
1 |
0 |
IIB |
(+) |
deep sequencing |
T2 |
76 |
M |
4 |
1 |
0 |
III |
(+) |
deep sequencing |
T3 |
67 |
M |
3 |
1 |
0 |
IIB |
(+) |
deep sequencing |
T4 |
65 |
M |
3 |
1 |
0 |
IIB |
(+) |
deep sequencing |
T5 |
56 |
F |
3 |
1 |
1 |
IV |
(+) |
deep sequencing |
T6 |
83 |
F |
3 |
0 |
0 |
IIA |
(-) |
deep sequencing |
T7 |
66 |
F |
3 |
1 |
0 |
IIB |
(-) |
deep sequencing |
N1 |
77 |
F |
deep sequencing |
|||||
N2 |
65 |
F |
deep sequencing |
|||||
N3 |
71 |
F |
deep sequencing |
|||||
N4 |
83 |
F |
deep sequencing |
|||||
PCR_T1 |
67 |
M |
3 |
1 |
0 |
IIB |
(+) |
RT-PCR |
PCR_T2 |
78 |
F |
3 |
0 |
0 |
IIA |
(-) |
RT-PCR |
PCR_T3 |
58 |
F |
3 |
0 |
0 |
IIA |
(+) |
RT-PCR |
PCR_T4 |
42 |
F |
3 |
1 |
0 |
IIB |
(+) |
RT-PCR |
PCR_T5 |
56 |
F |
3 |
0 |
0 |
IIA |
(+) |
RT-PCR |
PCR_T6 |
79 |
F |
3 |
0 |
0 |
IIA |
(+) |
RT-PCR |
PCR_T7 |
63 |
F |
3 |
0 |
0 |
IIA |
(+) |
RT-PCR |
PCR_T8 |
52 |
M |
3 |
0 |
0 |
IIA |
(-) |
RT-PCR |
PCR_T9 |
78 |
F |
3 |
1 |
0 |
IIB |
(+) |
RT-PCR |
PCR_T10 |
66 |
F |
3 |
1 |
0 |
IIB |
(-) |
RT-PCR |
PCR_T11 |
66 |
F |
3 |
0 |
0 |
IIA |
(-) |
RT-PCR |
PCR_T12 |
67 |
M |
is |
0 |
0 |
0 |
(+) |
RT-PCR |
PCR_T13 |
74 |
M |
4 |
0 |
0 |
III |
(-) |
RT-PCR |
PCR_T14 |
74 |
F |
3 |
1 |
0 |
IIB |
(-) |
RT-PCR |
PCR_T15 |
65 |
F |
3 |
1 |
0 |
IIB |
(-) |
RT-PCR |
PCR_T16 |
78 |
F |
3 |
0 |
0 |
IIA |
(-) |
RT-PCR |
PCR_T17 |
42 |
M |
4 |
1 |
0 |
III |
(+) |
RT-PCR |
PCR_T18 |
44 |
M |
3 |
1 |
0 |
IIB |
(+) |
RT-PCR |
PCR_T19 |
76 |
M |
3 |
1 |
0 |
IIB |
(+) |
RT-PCR |
PCR_T20 |
70 |
M |
3 |
0 |
0 |
IIA |
(+) |
RT-PCR |
PCR_T21 |
78 |
F |
1 |
0 |
0 |
I |
(-) |
RT-PCR |
PCR_T22 |
56 |
M |
1 |
0 |
0 |
I |
(+) |
RT-PCR |
PCR_T23 |
65 |
M |
3 |
0 |
0 |
IIA |
(-) |
RT-PCR |
PCR_T24 |
70 |
F |
3 |
1 |
0 |
IIB |
(+) |
RT-PCR |
PCR_N1 |
76 |
M |
RT-PCR |
|||||
PCR_N2 |
58 |
F |
RT-PCR |
|||||
PCR_N3 |
67 |
M |
RT-PCR |
|||||
PCR_N4 |
67 |
M |
RT-PCR |
|||||
PCR_N5 |
42 |
F |
RT-PCR |
|||||
PCR_N6 |
71 |
F |
RT-PC |
|||||
PCR_N7 |
60 |
M |
RT-PCR |
|||||
PCR_N8 |
56 |
F |
RT-PCR |
|||||
PCR_N9 |
67 |
M |
RT-PCR |
|||||
PCR_N10 |
85 |
F |
RT-PCR |
|||||
PCR_N11 |
66 |
F |
RT-PCR |
|||||
PCR_N12 |
66 |
F |
RT-PCR |
|||||
PCR_N13 |
44 |
M |
RT-PCR |
|||||
PCR_N14 |
63 |
F |
RT-PCR |
|||||
IHC_1 |
72 |
M |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_2 |
80 |
M |
3 |
1 |
0 |
IIB |
(-) |
IHC |
IHC_3 |
75 |
F |
3 |
1 |
0 |
IIB |
(-) |
IHC |
IHC_4 |
60 |
M |
4 |
0 |
0 |
III |
(+) |
IHC |
IHC_5 |
52 |
M |
3 |
0 |
0 |
IIA |
(+) |
IHC |
IHC_6 |
50 |
M |
4 |
0 |
0 |
III |
(+) |
IHC |
IHC_7 |
68 |
F |
4 |
0 |
0 |
III |
(-) |
IHC |
IHC_8 |
77 |
F |
4 |
0 |
0 |
III |
(+) |
IHC |
IHC_9 |
57 |
M |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_10 |
44 |
M |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_11 |
68 |
M |
4 |
1 |
0 |
III |
(+) |
IHC |
IHC_12 |
76 |
M |
4 |
1 |
0 |
III |
(+) |
IHC |
IHC_13 |
54 |
M |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_14 |
74 |
F |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_15 |
67 |
M |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_16 |
76 |
F |
3 |
0 |
0 |
IIA |
(-) |
IHC |
IHC_17 |
54 |
M |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_18 |
75 |
F |
3 |
0 |
0 |
IIA |
(-) |
IHC |
IHC_19 |
77 |
F |
2 |
0 |
0 |
IB |
(-) |
IHC |
IHC_20 |
66 |
M |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_21 |
69 |
M |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_22 |
69 |
M |
2 |
0 |
0 |
IB |
(+) |
IHC |
IHC_23 |
50 |
F |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_24 |
67 |
F |
3 |
1 |
0 |
III |
(+) |
IHC |
IHC_25 |
78 |
M |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_26 |
70 |
M |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_27 |
59 |
M |
3 |
0 |
0 |
IIA |
(-) |
IHC |
IHC_28 |
74 |
F |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_29 |
75 |
M |
3 |
0 |
0 |
IIA |
(+) |
IHC |
IHC_30 |
58 |
F |
3 |
0 |
0 |
IIA |
(+) |
IHC |
IHC_31 |
74 |
M |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_32 |
59 |
F |
3 |
1 |
0 |
IIB |
(+) |
IHC |
IHC_33 |
69 |
F |
3 |
0 |
0 |
IIA |
(-) |
IHC |
IHC_34 |
65 |
F |
4 |
1 |
0 |
III |
(+) |
IHC |
IHC_35 |
85 |
F |
3 |
1 |
0 |
IIB |
(+) |
IHC |
Table 2: Annotation of reads aligned to small RNAs
PDAC samples |
#T1 |
#T2 |
#T3 |
#T4 |
#T5 |
#T6 |
#T7 |
|||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Count |
(%) |
Count |
(%) |
Count |
(%) |
Count |
(%) |
Count |
(%) |
Count |
(%) |
Count |
(%) |
|
Total |
17591803 |
100 |
15426965 |
100 |
16586380 |
100 |
10615842 |
100 |
18965893 |
100 |
23407926 |
100 |
12845961 |
100 |
exon |
1018127 |
5.79 |
809048 |
5.24 |
184851 |
1.11 |
155240 |
1.46 |
540260 |
2.85 |
459079 |
1.96 |
263018 |
2.05 |
exon_antisense |
9 |
0.00 |
4 |
0.00 |
5 |
0.00 |
6 |
0.00 |
3 |
0.00 |
4 |
0.00 |
10 |
0.00 |
miRNA |
3045486 |
17.31 |
4835786 |
31.35 |
3464002 |
20.88 |
1721101 |
16.21 |
2740681 |
14.45 |
2664925 |
11.38 |
3152583 |
24.54 |
rRNA |
489091 |
2.78 |
1588798 |
10.30 |
216971 |
1.31 |
440961 |
4.15 |
181910 |
0.96 |
583477 |
2.49 |
490623 |
3.82 |
tRNA |
829391 |
4.71 |
110334 |
0.72 |
1076000 |
6.49 |
141156 |
1.33 |
1067387 |
5.63 |
542804 |
2.32 |
263598 |
2.05 |
snRNA |
24109 |
0.14 |
31039 |
0.20 |
9153 |
0.06 |
14372 |
0.14 |
18154 |
0.10 |
28997 |
0.12 |
24883 |
0.19 |
snoRNA |
848025 |
4.82 |
478768 |
3.10 |
617055 |
3.72 |
260963 |
2.46 |
440008 |
2.32 |
1211925 |
5.18 |
691475 |
5.38 |
lcnRNA |
77 |
0.00 |
56 |
0.00 |
30 |
0.00 |
24 |
0.00 |
273 |
0.00 |
78 |
0.00 |
85 |
0.00 |
ribozyme |
18531 |
0.11 |
8449 |
0.05 |
7111 |
0.04 |
6490 |
0.06 |
7255 |
0.04 |
16746 |
0.07 |
10127 |
0.08 |
sRNA |
15 |
0.00 |
17 |
0.00 |
17 |
0.00 |
17 |
0.00 |
29 |
0.00 |
27 |
0.00 |
28 |
0.00 |
Unannotated |
1489059 |
8.46 |
2684735 |
17.40 |
717881 |
4.33 |
5430607 |
51.16 |
2993173 |
15.78 |
8071185 |
34.48 |
4184768 |
32.58 |
Unmapped |
9829883 |
55.88 |
4879931 |
31.63 |
10293304 |
62.06 |
2444905 |
23.03 |
10976760 |
57.88 |
9828679 |
41.99 |
3764763 |
29.31 |
Normal pancreas samples |
#N1 |
#N2 |
#N3 |
#N4 |
||||
|---|---|---|---|---|---|---|---|---|
Count |
(%) |
Count |
(%) |
Count |
(%) |
Count |
(%) |
|
Total |
16535900 |
100 |
20967863 |
100 |
17370365 |
100 |
18447846 |
100 |
exon |
281915 |
1.70 |
270434 |
1.29 |
576712 |
3.32 |
963571 |
5.22 |
exon_antisense |
2 |
0.00 |
9 |
0.00 |
39 |
0.00 |
24 |
0.00 |
miRNA |
4247206 |
25.68 |
4331251 |
20.66 |
3534149 |
20.35 |
5133067 |
27.82 |
rRNA |
659176 |
3.99 |
310278 |
1.48 |
1457154 |
8.39 |
810443 |
4.39 |
tRNA |
334461 |
2.02 |
1145451 |
5.46 |
418994 |
2.41 |
568066 |
3.08 |
snRNA |
19163 |
0.12 |
22558 |
0.11 |
28997 |
0.17 |
25124 |
0.14 |
snoRNA |
703693 |
4.26 |
880199 |
4.20 |
714087 |
4.11 |
641479 |
3.48 |
lcnRNA |
41 |
0.00 |
39 |
0.00 |
15 |
0.00 |
72 |
0.00 |
ribozyme |
3967 |
0.02 |
7398 |
0.04 |
11294 |
0.07 |
9258 |
0.05 |
sRNA |
23 |
0.00 |
31 |
0.00 |
37 |
0.00 |
32 |
0.00 |
Unannotated |
5295677 |
32.03 |
966037 |
4.61 |
2152804 |
12.39 |
1710926 |
9.27 |
Unmapped |
4990576 |
30.18 |
13034178 |
62.16 |
8476083 |
48.80 |
8585784 |
46.54 |
We focused on 23 downregulated miRNAs (Table 3), and these miRNAs were mapped on the human genome. We found that miR-217, miR-216a-5p/-3p and miR216b-5p/-3p were close together and were defined as clustered miRNAs. Because the role of the miR-216 cluster had not been reported in PDAC, we focused on it in further studies. As shown in Figure 1, miR-216a-5p/-3p and miR216b-5p/-3p are located within the same chromosomal region (2q16.1).
Table 3: Differentially expressed miRNAs in PDAC by RNA seqencing
Downregulated miRNAs in PDAC |
|||||
miRNA |
miRBase_accession |
Locus |
LogFC (tumor/normal) |
P value |
FDR |
hsa-miR-216a-5p |
MIMAT0000273 |
chr2:56216155-56216176 |
-3.22 |
0.0033 |
0.4602 |
hsa-miR-802 |
MIMAT0004185 |
chr21:37093030-37093052 |
-3.17 |
0.0001 |
0.0901 |
hsa-miR-3186-3p |
MIMAT0015068 |
chr17:79418142-79418162 |
-3.16 |
0.0072 |
0.5030 |
hsa-miR-217 |
MIMAT0000274 |
chr2:56210155-56210177 |
-3.13 |
0.0034 |
0.4602 |
hsa-miR-2114-5p |
MIMAT0011156 |
chrX:149396251-149396272 |
-3.08 |
0.0033 |
0.4602 |
hsa-miR-216b-5p |
MIMAT0004959 |
chr2:56227899-56227920 |
-2.77 |
0.0286 |
0.9691 |
hsa-miR-3186-5p |
MIMAT0015067 |
chr17:79418178-79418199 |
-2.70 |
0.0230 |
0.9200 |
hsa-miR-216b-3p |
MIMAT0026721 |
chr2:56227859-56227882 |
-2.58 |
0.0008 |
0.1722 |
hsa-miR-6510-3p |
MIMAT0025477 |
chr17:39673418-39673438 |
-2.57 |
0.0098 |
0.6010 |
hsa-miR-122-5p |
MIMAT0000421 |
chr18:56118320-56118341 |
-2.51 |
0.0207 |
0.8791 |
hsa-miR-412-3p |
MIMAT0002170 |
chr14:101531837-101531859 |
-2.42 |
0.0393 |
1.0000 |
hsa-miR-129-1-3p |
MIMAT0004548 |
chr7:127847973-127847994 |
-2.40 |
0.0011 |
0.2082 |
hsa-miR-216a-3p |
MIMAT0022844 |
chr2:56216116-56216137 |
-2.38 |
0.0214 |
0.8791 |
hsa-miR-135a-3p |
MIMAT0004595 |
chr3:52328248-52328269 |
-2.37 |
0.0008 |
0.1722 |
hsa-miR-148a-5p |
MIMAT0004549 |
chr7:25989580-25989601 |
-2.30 |
0.0009 |
0.1722 |
hsa-miR-891a-5p |
MIMAT0004902 |
chrX:145109359-145109380 |
-2.25 |
0.0053 |
0.5030 |
hsa-miR-190b |
MIMAT0004929 |
chr1:154166189-154166209 |
-2.12 |
0.0003 |
0.1429 |
hsa-miR-148a-3p |
MIMAT0000243 |
chr7:25989542-25989563 |
-2.12 |
0.0001 |
0.0901 |
hsa-miR-211-5p |
MIMAT0000268 |
chr15:31357298-31357319 |
-2.10 |
0.0084 |
0.5545 |
hsa-miR-1224-5p |
MIMAT0005458 |
chr3:183959193-183959211 |
-2.05 |
0.0450 |
1.0000 |
hsa-miR-338-5p |
MIMAT0004701 |
chr17:79099723-79099744 |
-2.02 |
0.0002 |
0.0956 |
hsa-miR-129-2-3p |
MIMAT0004605 |
chr11:43603000-43603021 |
-2.01 |
0.0090 |
0.5650 |
hsa-miR-137 |
MIMAT0000429 |
chr1:98511647-98511669 |
-2.01 |
0.0874 |
1.0000 |
Upregulated miRNAs in PDAC |
|||||
miRNA |
miRBase_accession |
Locus |
LogFC (tumor/normal) |
Pvalue |
FDR |
hsa-miR-6887-3p |
MIMAT0027675 |
chr19:35613648-35613668 |
4.13 |
0.0007 |
0.1722 |
hsa-miR-4713-5p |
MIMAT0019820 |
chr15:51534397-51534418 |
4.04 |
0.0039 |
0.4750 |
hsa-miR-615-5p |
MIMAT0004804 |
chr12:54427751-54427772 |
3.61 |
0.0053 |
0.5030 |
hsa-miR-1248 |
MIMAT0005900 |
chr3:186504464-186504490 |
3.50 |
0.0002 |
0.0956 |
hsa-miR-490-3p |
MIMAT0002806 |
chr7:136587989-136588010 |
3.47 |
0.0386 |
1.0000 |
hsa-miR-184 |
MIMAT0000454 |
chr15:79502182-79502203 |
3.46 |
0.0467 |
1.0000 |
hsa-miR-196a-5p |
MIMAT0000226 |
chr17:46709894-46709915chr12:54385546-54385567 |
3.24 |
0.0103 |
0.6146 |
hsa-miR-187-3p |
MIMAT0000262 |
chr18:33484798-33484819 |
3.05 |
0.0195 |
0.8676 |
hsa-miR-6516-3p |
MIMAT0030418 |
chr17:75085551-75085572 |
3.03 |
0.0046 |
0.5030 |
hsa-miR-490-5p |
MIMAT0004764 |
chr7:136587952-136587971 |
2.95 |
0.0457 |
1.0000 |
hsa-miR-3131 |
MIMAT0014996 |
chr2:219923447-219923469 |
2.94 |
0.0112 |
0.6428 |
hsa-miR-4697-3p |
MIMAT0019792 |
chr11:133768401-133768424 |
2.90 |
0.0070 |
0.5030 |
hsa-miR-615-3p |
MIMAT0003283 |
chr12:54427794-54427815 |
2.86 |
0.0001 |
0.0901 |
hsa-miR-196b-5p |
MIMAT0001080 |
chr7:27209147-27209168 |
2.73 |
0.0009 |
0.1722 |
hsa-miR-187-5p |
MIMAT0004561 |
chr18:33484834-33484855 |
2.70 |
0.0610 |
1.0000 |
hsa-miR-6886-3p |
MIMAT0027673 |
chr19:11224187-11224207 |
2.68 |
0.0247 |
0.9200 |
hsa-miR-650 |
MIMAT0003320 |
chr22:23165285-23165305 |
2.67 |
0.0138 |
0.7105 |
hsa-miR-492 |
MIMAT0002812 |
chr12:95228203-95228225 |
2.66 |
0.0257 |
0.9200 |
hsa-miR-6744-5p |
MIMAT0027389 |
chr11:1277840-1277858 |
2.65 |
0.0169 |
0.8201 |
hsa-miR-6805-5p |
MIMAT0027510 |
chr19:55899554-55899575 |
2.55 |
0.0603 |
1.0000 |
hsa-miR-935 |
MIMAT0004978 |
chr19:54485616-54485638 |
2.53 |
0.0059 |
0.5030 |
hsa-miR-4485 |
MIMAT0019019 |
chr11:10529824-10529843 |
2.53 |
0.0192 |
0.8676 |
hsa-miR-196a-3p |
MIMAT0004562 |
chr12:54385583-54385604 |
2.52 |
0.0627 |
1.0000 |
hsa-miR-1 |
MIMAT0000416 |
chr18:19408976-19408997 chr20:61151558-61151579 |
2.48 |
0.0697 |
1.0000 |
hsa-miR-4768-5p |
MIMAT0019920 |
chrX:17444012-17444033 |
2.42 |
0.0567 |
1.0000 |
hsa-miR-6887-5p |
MIMAT0027674 |
chr19:35613609-35613631 |
2.39 |
0.0749 |
1.0000 |
hsa-miR-181b-3p |
MIMAT0022692 |
chr1:198828016-198828036 |
2.34 |
0.0827 |
1.0000 |
hsa-miR-4745-5p |
MIMAT0019878 |
chr19:804941-804963 |
2.32 |
0.0491 |
1.0000 |
hsa-miR-4724-5p |
MIMAT0019841 |
chr17:29861911-29861933 |
2.25 |
0.0950 |
1.0000 |
hsa-miR-6814-3p |
MIMAT0027529 |
chr21:43166932-43166953 |
2.15 |
0.0664 |
1.0000 |
hsa-miR-203b-3p |
MIMAT0019814 |
chr14:104583765-104583787 |
2.10 |
0.0518 |
1.0000 |
hsa-miR-143-5p |
MIMAT0004599 |
chr5:148808507-148808528 |
2.09 |
0.0019 |
0.3270 |
hsa-miR-6850-5p |
MIMAT0027600 |
chr8:146017355-146017376 |
2.09 |
0.1212 |
1.0000 |
hsa-miR-549a |
MIMAT0003333 |
chr15:81134334-81134354 |
2.06 |
0.1130 |
1.0000 |
hsa-miR-133a-3p |
MIMAT0000427 |
chr18:19405673-19405694 chr20:61162177-61162198 |
2.06 |
0.0275 |
0.9691 |
hsa-miR-3661 |
MIMAT0018082 |
chr5:133561468-133561489 |
2.03 |
0.0800 |
1.0000 |
hsa-miR-4649-5p |
MIMAT0019711 |
chr7:44150450-44150473 |
2.02 |
0.0661 |
1.0000 |
hsa-miR-31-3p |
MIMAT0004504 |
chr9:21512120-21512141 |
2.02 |
0.0195 |
0.8676 |
hsa-miR-6862-5p |
MIMAT0027625 |
chr16:28402346-28402367 chr16:28735578-28735599 |
2.01 |
0.0770 |
1.0000 |
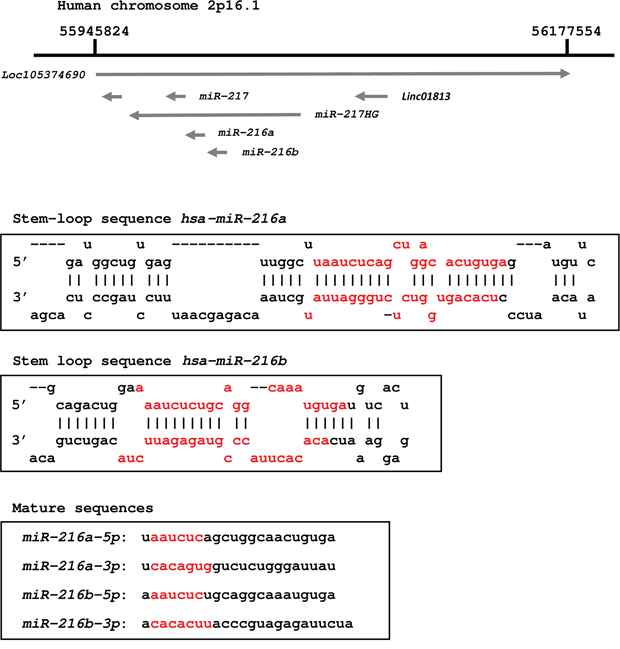
Figure 1: Schematic representation of the human miR-216 family in chromosomal location. The miR-216 family members and miR-217 are located on human chromosome 2q16.1. Mature miRNAs, miR-216a-5p (guide strand) and miR-216a-3p (passenger strand) are derived from pre-miR-216a. Likewise pre-miR-216a, miR-216b-5p (guide strand) and miR-216b-3p (passenger strand) are derived from pre-miR-216b. Seed sequences of miR-216a-5p and miR-216b-5p are identical. In contrast, seed sequences of miR-216a-3p and miR-216b-3p are independent sequences.
Expression levels of the miR-216 family in PDAC specimens and cell lines
We evaluated expression levels of the miR-216 family in PDAC tissues (n = 24), normal pancreatic tissues (n = 14) and two PDAC cell lines (PANC-1 and SW 1990). Patient clinicopathological features are summarized in Table 1. The expression levels of miR-216a-5p, miR-216a-3p, miR-216b-5p and miR-216b-3p were significantly lower in tumour tissues than in normal pancreatic tissues (Figure 2). It was not recognized the significant relationships between any of the clinical parameters (i.e., TNM stage, metastasis or survival rate) and expression levels of any member of the miR-216-family (data not shown).
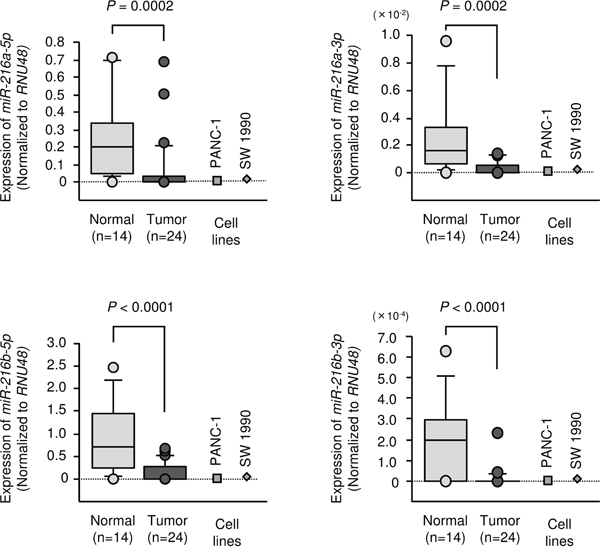
Figure 2: Expression levels of miR-216 family members. Expression levels of miR-216 family members in clinical specimens and PDAC cell lines were determined using qRT-PCR. Data were normalized to RNU48 expression.
To elucidate molecular mechanisms of low expression of the clustered miRNAs, miR-216a-5p, miR-216a-3p, miR-216b-5p and miR-216b-3p in PDAC cells, PANC-1 and SW1990 cells were treated with the demethylating agent [5-aza-2’-deoxycytidine (5-aza-dC)]. Expression levels of all clustered miRNAs in PDAC cells were significantly elevated by 5-aza-dc treatment (Supplementary Figure 1). These data suggested that DNA methylation might cause silencing of these clustered miRNAs in PDAC cells.
Expression of miR-216 family members and the effects on cell growth, migration and invasion in PDAC cell lines
The functional roles of the miR-216-family were performed by using restoration of mature miRNAs assays. Functional assays showed that proliferation, migration and invasion activities of cancer cells were significantly suppressed in miR-216a-5p, miR-216a-3p, miR-216b-5p and miR-216b-3p transfectants compared with mock or miR-control transfectants (Figure 3). Present results suggested that the miR-216 family could have anti-tumour functions in PDAC cells. Among the miR-216 family, miR-216b-3p markedly inhibited PDAC cell aggressiveness (Figure 3).
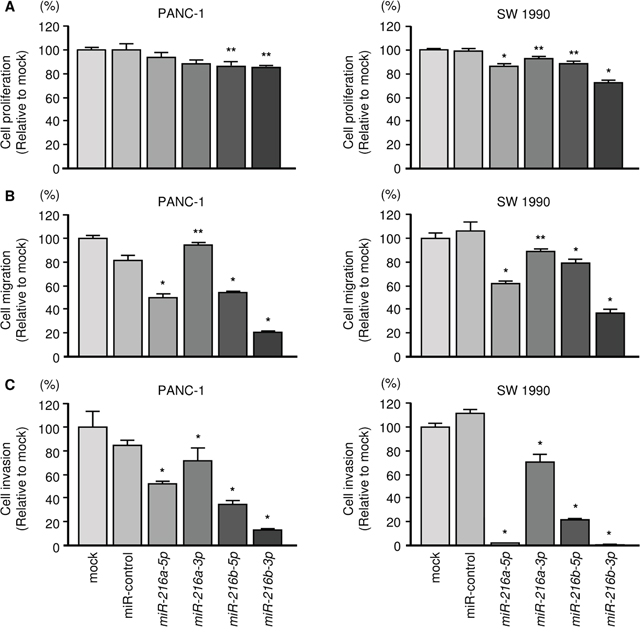
Figure 3: Effects of miR-216a-5p, miR-216a-3p, miR-216b-5p and miR-216b-3p transfection into PDAC cells. (A) Cell growth was determined using XTT assays 72 h after transfection with 10 nM miR-216 family members. *, P < 0.0001; **, P < 0.05. (B) Cell migration activity was determined using BD Falcon Cell Culture Inserts. *, P < 0.0001. (C) Cell invasion activity was determined using Matrigel invasion assays. *, P < 0.0001.
We hypothesized that passenger strand of miR-216-3p may be incorporated into and function as part of the RISC structure. To check this hypothesis, we performed immunoprecipitation with antibody targeting Ago2, which plays a pivotal role in the RISC. After transfection with miR-216-3p, Ago2-bound miRNAs were isolated, and RT-qPCR was carried out to determine whether miR-216-3p bound to Ago2. Expression of miR-216-3p levels were significantly higher than those of mock and miRNA control transfected cells (Supplementary Figure 2). Moreover, expression levels of miR-216a-5p, miR-216a-3p and miR-216b-5p were same levels of mock and miRNA control transfectants (Supplementary Figure 2).
Identification of genes regulated by miR-216b-3p in PDAC cells
To elucidate the molecular mechanisms and pathways regulated by anti-tumour miR-216b-3p in PDAC cells, we used a combination of gene expression and in silico analyses. Figure 4 presents the strategy for narrowing down the target genes of miR-216b-3p. In gene expression analyses, 439 and 806 genes were downregulated (log2 ratio <-1.0) in PANC-1 and SW 1990 miR-216b-3p transfectants, respectively, in comparison with control transfectants (GEO accession number, GSE82108). Next, we pared down the list of genes using the TargetscanHuman and GEO database (GEO accession number, GSE15471). We found that 20 genes were putatively targeted by miR-216b-3p in PANC-1 and SW 1990 miR-216b-3p transfectants.
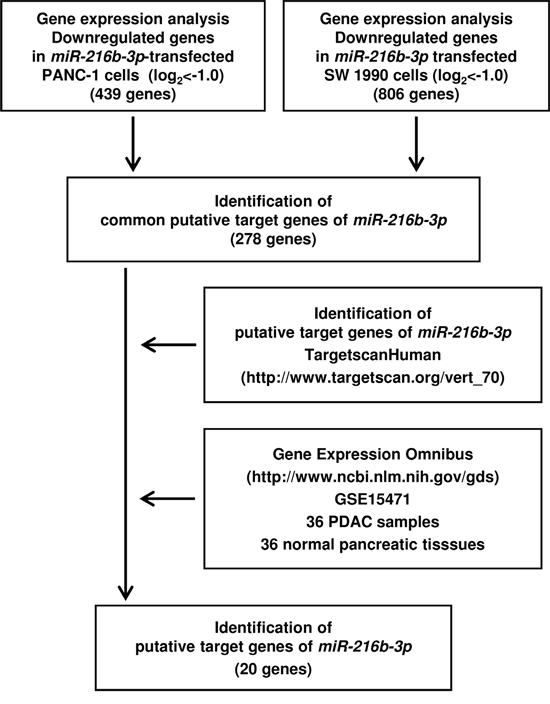
Figure 4: Flow chart illustrating the analytic strategy for miR-216b-3p target genes. Strategy for identification of putative candidate genes regulated by miR-216b-3p in PDAC cells. Approach to identifying miR-216b-3p target genes by in silico analysis of genome-wide gene expression, TargetScan and GEO database analysis of miR-216b-3p transfectants and PDAC clinical specimens.
Among these candidate genes, 6 genes (PLAU, FOXQ1, PGM2L1, MBOAT2, PERP and UHMK1) were significantly associated with poor prognosis of patients with PDAC (Table 4 and Supplementary Figure 3). Past studies showed that aberrant expressed FOX-family of transcription factors were involved in several types of cancers, however, functional significance of FOXQ1 in PDAC is still unclear.
We determined that FOXQ1 was upregulated in clinical PDAC samples using qRT-PCR (Figure 5A). Negative correlations between miR-216b-3p expression and FOXQ1 mRNA expression were found using Spearman’s rank test (R = -0.5160, P = 0.0006, Figure 5B). Kaplan–Meier survival curves constructed from PDAC patients in the TCGA dataset showed that patient groups with high expression of FOXQ1 had significantly shorter overall survival (P = 0.0070; Figure 5D). The TCGA dataset was analysed by OncoLnc database (http://www.oncolnc.org/)[24].
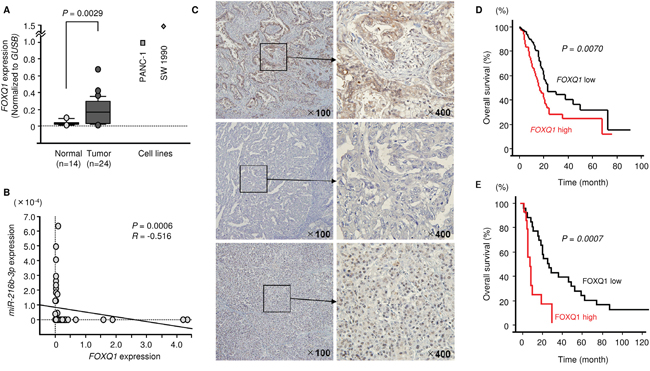
Figure 5: Expression of FOXQ1 (mRNA and protein) in PDAC and association of FOXQ1 with overall survival. (A) Expression levels of FOXQ1 in clinical specimens and PDAC cell lines were determined using qRT-PCR. Data were normalized to GUSB expression. (B) Correlation between miR-216-3p and FOXQ1 expression. (C) Immunohistochemical staining of FOXQ1 in PDAC clinical specimens. Overexpression of FOXQ1 was observed in cancer lesions (original magnification ×400). In contrast, negative staining of FOXQ1 was observed in normal tissues. (IHC_13) Positively stained cancer lesion (T3N1M0). (IHC_18) Negatively stained cancer lesion (T3N0M0). (IHC_29) Negatively stained normal pancreas tissue. (D) TCGA-based large cohort study of high expression of FOXQ1 in 174 PDAC patients. (E) Kaplan-Meier patient survival curves for overall survival rates based on FOXQ1 expression in 35 patients with PDAC. P-values were calculated using the log-rank test.
Table 4: Candidate target genes regulated by miR-216b-3p in PDAC
Entrez gene ID |
Gene symbol |
Description |
Microarray (Log2 ratio)miR-216b-3p |
Target site (Targetscan) |
GEO |
TCGA-OncoLncKaplan plot P-value |
||
|---|---|---|---|---|---|---|---|---|
PANC-1 |
SW1990 |
Average |
miR-216b-3p |
FC |
||||
5328 |
PLAU |
plasminogen activator, urokinase |
-1.03 |
-1.10 |
-1.06 |
(+) |
5.07 |
0.0161 |
59345 |
GNB4 |
guanine nucleotide binding protein (G protein), beta polypeptide 4 |
-1.76 |
-2.27 |
2.01 |
(+) |
4.57 |
0.3330 |
94234 |
FOXQ1 |
forkhead box Q1 |
-1.50 |
-1.52 |
-1.51 |
(+) |
3.88 |
0.0089 |
9240 |
PNMA1 |
paraneoplastic Ma antigen 1 |
-1.24 |
-1.69 |
-1.47 |
(+) |
3.27 |
0.9910 |
283209 |
PGM2L1 |
phosphoglucomutase 2-like 1 |
-1.67 |
-1.91 |
-1.79 |
(+) |
3.23 |
0.0477 |
60481 |
ELOVL5 |
ELOVL fatty acid elongase 5 |
-1.50 |
-2.77 |
-2.13 |
(+) |
3.15 |
0.4970 |
129642 |
MBOAT2 |
membrane bound O-acyltransferase domain containing 2 |
-1.20 |
-1.40 |
-1.30 |
(+) |
2.92 |
0.0158 |
5912 |
RAP2B |
RAP2B, member of RAS oncogene family |
-1.31 |
-1.25 |
-1.28 |
(+) |
2.74 |
0.1040 |
5738 |
PTGFRN |
prostaglandin F2 receptor inhibitor |
-1.57 |
-2.00 |
-1.79 |
(+) |
2.64 |
0.4350 |
493 |
ATP2B4 |
ATPase, Ca++ transporting, plasma membrane 4 |
-1.45 |
-2.27 |
-1.86 |
(+) |
2.55 |
0.0966 |
54492 |
NEURL1B |
neuralized E3 ubiquitin protein ligase 1B |
-1.32 |
-1.95 |
-1.64 |
(+) |
2.36 |
0.0365 |
64065 |
PERP |
PERP, TP53 apoptosis effector |
-1.78 |
-1.98 |
-1.88 |
(+) |
2.33 |
<0.0001 |
160418 |
TMTC3 |
transmembrane and tetratricopeptide repeat containing 3 |
-1.85 |
-2.24 |
-2.05 |
(+) |
2.30 |
0.9080 |
55589 |
BMP2K |
BMP2 inducible kinase |
-1.04 |
-1.54 |
-1.29 |
(+) |
2.29 |
0.5220 |
3977 |
LIFR |
leukemia inhibitory factor receptor alpha |
-1.96 |
-2.09 |
-2.03 |
(+) |
2.26 |
0.0062* |
10920 |
COPS8 |
COP9 signalosome subunit 8 |
-1.43 |
-2.04 |
-1.73 |
(+) |
2.20 |
0.2740 |
9637 |
FEZ2 |
fasciculation and elongation protein zeta 2 (zygin II) |
-1.03 |
-1.28 |
-1.15 |
(+) |
2.18 |
0.0974 |
4286 |
MITF |
microphthalmia-associated transcription factor |
-1.02 |
-1.07 |
-1.05 |
(+) |
2.17 |
0.0963 |
29967 |
LRP12 |
low density lipoprotein receptor-related protein 12 |
-1.32 |
-2.22 |
-1.77 |
(+) |
2.09 |
0.6200 |
127933 |
UHMK1 |
U2AF homology motif (UHM) kinase 1 |
-1.20 |
-1.79 |
-1.50 |
(+) |
2.04 |
0.0329 |
* Poor prognosis with a low expression
Expression of FOXQ1 in PDAC clinical specimens
We validated the expression of FOXQ1 in PDAC clinical specimens using immunohistochemistry. A total of 35 specimens were evaluated, and 12 samples were classified as having high expression of FOXQ1. Clinicopathological characteristics are summarized in Table 1. Table 5 shows the correlation between FOXQ1 expression and various clinicopathological factors. High FOXQ1 expression was significantly associated with increased lymph node metastasis. Furthermore, patients with high FOXQ1 expression had significantly shorter OS than those with low FOXQ1 expression (P = 0.0007) (Figure 5E).
Table 5: Correlation between the expression of FOXQ1 and clinicopathological factors in PDAC (n =35)
Characteristic |
FOXQ1 |
P |
|
|---|---|---|---|
Low (n = 23) |
High (n = 12) |
||
Age (y)a |
bNS |
||
≥ 60 |
17 |
10 |
|
< 60 |
6 |
2 |
|
Gender (n) |
|||
Male (20) |
13 |
7 |
NS |
Female (15) |
10 |
5 |
|
Tumor size (n) |
|||
> 40 mm (5) |
2 |
3 |
NS |
≤ 40 mm (30) |
21 |
9 |
|
Lymph node metastasis (n) |
|||
No (11) |
10 |
1 |
0.0088 |
Yes (24) |
11 |
13 |
|
Distant metastasis (n) |
|||
No (35) |
23 |
12 |
NS |
TNM Stage (n) |
|||
I / II (27) |
18 |
9 |
NS |
III / IV (8) |
5 |
3 |
|
Recurrence (n) |
|||
No (8) |
6 |
2 |
NS |
Yes (27) |
17 |
10 |
|
aValues are mean ± SD.
bNS, not significant.
Moreover, we categorized patients into two groups, with adjuvant chemotherapy or without adjuvant chemotherapy. Regardless of the treatment, patients with high FOXQ1 expression were associated with poor prognosis (Supplementary Figure 4).
We confirmed the expression of FOXQ1 in PDAC clinical specimens using immunohistochemistry. Expression of FOXQ1 was detected in in ductal carcinoma, not in normal ductal cells, acinar cells and islets cells (Supplementary Figure 5).
FOXQ1 is a direct target of miR-216b-3p in PDAC cells
We performed qRT-PCR to validate miR-216b-3p-mediated reduction of FOXQ1 mRNA expression in PDAC cell lines. Our studies demonstrated that FOXQ1 mRNA was markedly reduced in miR-216b-3p transfectants compared to mock or miR-control transfectants (P < 0.0001, Figure 6A). FOXQ1 protein expression was also repressed in the miR-216b-3p transfectants (Figure 6B).
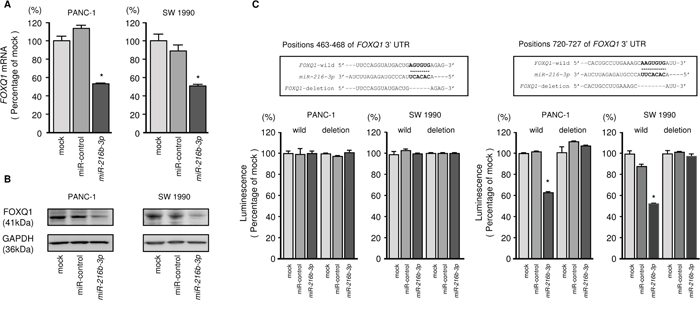
Figure 6: Direct regulation of FOXQ1 by miR-216b-3p in PDAC cells. (A) FOXQ1 mRNA expression was evaluated using qRT-PCR in PANC-1 and SW 1990 cells 72 h after transfection with miR-216b-3p. GUSB was used as an internal control. *, P < 0.0001. (B) FOXQ1 protein expression was evaluated by Western blotting in PANC-1 and SW 1990 cells 72 to 96 h after transfection with miR-216b-3p. GAPDH was used as a loading control. (C) miR-216b-3p binding sites in the 3’-UTR of FOXQ1 mRNA. Dual luciferase reporter assays using vectors encoding putative miR-216b-3p target sites of the FOXQ1 3’-UTR (positions 463-468 and 720-727) for both wild-type and deleted regions. Data were normalized by expressing ratios of Renilla/firefly luciferase activities. *, P < 0.0001.
Target prediction databases indicated the presence of 2 putative target sites in the 3'-UTR of FOXQ1 (Figure 6C). To determine whether FOXQ1 mRNA had a functional target site, we performed luciferase reporter assays. Compared with the miR-control, luminescence intensity was significantly reduced by transfection with miR-216b-3p at its target site, position 720-727 in the 3’-UTR of FOXQ1 (Figure 6C).
Effects of silencing FOXQ1 on PDAC cell lines
To investigate the oncogenic functions of FOXQ1 in PDAC cells, we carried out knockdown assays by using si-FOXQ1. First, we evaluated the knockdown efficiency achieved by si-FOXQ1 transfection of PDAC cell lines. We used 2 types of si-FOXQ1 (si-FOXQ1-1 and si-FOXQ1-2) in this study. Both siRNAs effectively downregulated FOXQ1 expression in both cell lines (Figure 7A and 7B). Functional assays showed that si-FOXQ1 transfection significantly reduced cell proliferation, migration and invasion compared with mock- or siRNA-control-transfected cells (Figure 7C-7E).
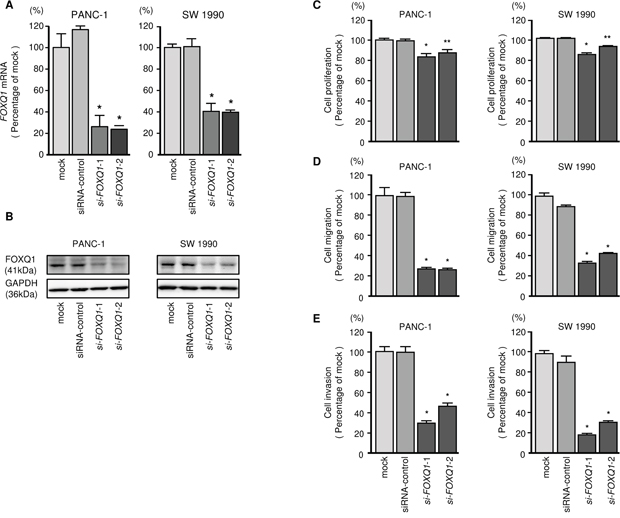
Figure 7: Effects of FOXQ1 silencing on PDAC cell lines. (A) FOXQ1 mRNA expression was evaluated using qRT-PCR analysis of PANC-1 and SW 1990 cells 72 h after transfection with si-FOXQ1-1 or si-FOXQ1-2. GUSB was used as an internal control. (B) FOXQ1 protein expression was evaluated using Western blotting analysis of PANC-1 and SW 1990 cells 72 to 96 h after transfection with miR-216b-3p. GAPDH was used as a loading control. (C) Cell proliferation was determined using XTT assays 72 h after transfection with 10 nM si-FOXQ1-1 or si-FOXQ1-2. *, P < 0.0001; **, P < 0.05. (D) Cell migration activity was determined using BD Falcon Cell Culture Inserts. *, P < 0.0001. (E) Cell invasion activity was determined using Matrigel invasion assays. *, P < 0.0001.
In contrast to loss-of-function assays, overexpression of FOXQ1 enhanced cancer cell migration and invasion in PANC-1 cells (Supplementary Figure 6).
Investigation of FOXQ1-mediated downstream genes in PDAC cells
Genome-wide gene expression (GEO accession number, GSE77790) and in silico analyses were performed to investigate the downstream genes mediated by FOXQ1 in a PDAC cell line (PANC-1) transfected with si-FOXQ1. A total of 18,277 genes were commonly downregulated (log2 ratio < 0). Further GEO analyses showed that 1,003 genes upregulated expression (log2 ratio > 1.0) in GEO accession number GSE15471. Next, we assigned genes to KEGG pathways using the GENECODIS program and identified 261 pathways. In the top 3 pathways, 41 genes were significantly enriched (Supplementary Tables 3A and 3B).
TCGA database analyses demonstrated that high expression of 13 genes were significantly associated with poor survival in patients with PDAC (Supplementary Figure 7).
DISCUSSION
miRNA fine-tunes the expression control of protein coding/noncoding RNAs by repressing translation or cleaving RNA transcripts. Dysregulated miRNAs can disrupt tightly controlled RNA networks and thereby enhance the aggressiveness of cancer cells. Identification of dysregulated miRNAs and their targeted genes can improve our understanding of the molecular pathogenesis of human cancers. In that regard, we have identified novel RNA networks in several cancers [13–15].
In the present study, we newly constructed a miRNA expression signature of PDAC by RNA sequencing methods. Several miRNA signatures of PDAC were reported in which formalin-fixed paraffin-embedded tissue or fine-needle aspiration biopsies specimens were used for PCR or microarray-based applications [25–27]. Recently, we summarized dysregulated miRNAs in PDAC using information in public databases [12]. Downregulation of miR-217, miR-141, miR-148a and miR-375 in PDAC tissues was commonly observed in PDAC tissues [12]. More recently, we demonstrated the anti-tumour roles of miR-375 through its targeting of ZFP36L2 in PDAC cells [28]. Our present signature included several anti-tumour miRNAs that had been previously identified in PDAC, suggesting that our RNA-based signature was effective for identification of novel oncogenes and anti-tumour miRNAs in PDAC cells. These data may provide a benchmark for future studies of PDAC.
Our signature revealed that novel miRNAs were aberrantly expressed in PDAC cells. Among them, we focused on 4 clustered miRNAs, miR-216a-5p, miR-216a-3p, miR-216b-5p and miR-216b-3p located on human chromosome 2p16.1. Here, we showed that the clustered miR-216 family has anti-tumour activity in PDAC. Those results suggest that the miR-216 family is important in PDAC oncogenesis. Past studies showed that guide strands of miR-216a-5p and miR-216b-5p acted as anti-tumour miRNAs in several cancers [29, 30]. On the other hand, the functional significance of passenger strands miR-216a-3p and miR-216b-3p has not been adequately analysed.
In miRNA biogenesis, the RNA guide strand from the duplex is recruited to the RNA-induced silencing complex (RISC) to target messenger RNAs, whereas the passenger strand is degraded and no functional activity [31, 32]. Nevertheless, our present data showed that passenger strands miR-216a-3p and miR-216b-3p have anti-tumour functions in PDAC cells. Recently, we demonstrated that both strands of pre-miR-145 (miR-145-5p, guide strand, and miR-145-3p, passenger strand) were incorporated into RISC and function anti-tumour miRNAs by targeting several oncogenes in bladder and lung cancer cells [21, 22]. The involvement of passenger strand miRNAs in the regulation of cellular processes is a novel concept in RNA research.
The KrasG12D oncogene is a key oncogenic mutation that is associated with development of PDAC [33, 34]. A recent study evaluated miRNA expression in the KrasG12D transgenic mouse model of PDAC [35]. In that study, 3 miRNAs (miR-216a, miR-216b, and miR-217) were reduced in expression in the KrasG12D pancreas. Other study showed that significantly downregulation of miR-216a and miR-217 was detected in KrasG12d;Pdx1-Cre mouse at 25 weeks [36]. Due to the lack of human clinical specimens from early stages of PDAC, the investigation is important to analyze expression patterns of miRNAs in early stages of PDAC by using mouse model. The molecular mechanisms of downregulation of these clustered miRNAs are still obscure.
To better understand and identify the anti-tumour targets of miR-216b-3p in PDAC cells, we have applied in silico database and genome-wide gene expression analyses. In this study, FOXQ1 (a member of the forkhead transcription factor family) was identified as a direct target of miR-216b-3p regulation in PDAC cells. Moreover, our functional studies revealed that expression of FOXQ1 promoted cancer cell aggressiveness, and overexpression of FOXQ1 was involved in PDAC pathogenesis. The FOX-family consists of a large number of transcription factors that have been categorized in 19 subfamilies, FOXA-FOXS [37, 38]. FOX transcription factors play pivotal roles in regulating the expression of genes involved in several biological processes [37, 38]. Numerous studies have demonstrated that FOXQ1 (as well as other FOX-family member) is associated with the progression, metastasis and drug resistance of human cancers [39].
The epithelial-mesenchymal transition (EMT) is pivotal in cancer cell invasion, metastasis and chemoresistance [40, 41]. Many EMT-promoting transcription factors have been implicated in these events in cancer cells [40, 41]. In lung cancer cells, elimination of FOXQ1 affected the re-expression of epithelial markers and decreases mesenchymal markers in vitro and in vivo[42]. Similar studies reported that upregulation of FOXQ1 triggered EMT and contributed to metastasis and chemoresistance in several cancers [43–45]. The existence of cancer stem cells (CSCs) is an important concept in cancer cell aggressiveness, metastasis and chemoresistance [46]. In PDAC, CSCs were isolated based on expression of CD44, CD133 and EpCAM. Overexpression of FOXQ1 was detected in CSCs and CSCs behaver have more aggressiveness for cell growth, compared with parental MiaPaCa-2 cells [47].
A large number of cohort analyses of the TCGA dataset showed that high expression of FOXQ1 predicted shortened survival of patients with PDAC. Likewise, there are many reports that expression of FOXQ1 indicates poor prognosis [44, 45, 48]. In this study, we identified “ECM-receptor interaction”, “regulation of actin cytoskeleton” and “focal adhesion pathways” as FOXQ1-mediated downstream pathways in PDAC cells. Interestingly, among 41 downstream genes of FOXQ1, 13 genes were associated with poor prognosis of patients with PDAC by TCGA analyses. These findings suggest that expression of FOXQ1 and FOXQ1-mediated pathways contribute significantly to PDAC aggressiveness.
Our miRNA signature showed that miR-217 was also markedly reduced in PDAC tissues. This miRNA is located in the same genomic region as the miR-216 cluster. Our data showed that ectopic expression of miR-217 significantly inhibited cancer cell aggressiveness in PDAC cells by targeting focal adhesion pathways (data not shown). Past studies pointed to the anti-tumour function of miR-217 in several cancers, including the targeting of the KRAS oncogene in PDAC [49].
In conclusion, downregulation of 4 clustered miRNAs (miR-216a-5p, miR-216a-3p, miR-216b-5p and miR-216b-3p) was detected by RNA sequencing-based miRNA signature analysis. The tumour-suppressor functions of these miRNAs were shown in PDAC cells. The expression of FOXQ1 was directly regulated by miR-216b-3p and its overexpression was involved in PDAC pathogenesis. Identification of novel cancer networks mediated by aberrantly expressed miRNAs and tumour-suppressor miRNAs may improve our understanding of PDAC molecular pathogenesis. Our newly created RNA sequencing-based miRNA signature provides a basis for further PDAC research.
MATERIALS AND METHODS
Clinical specimens and cell lines
We used fresh (n = 24) and preserved (formalin-fixed, paraffin-embedded blocks) (n = 35) specimens for the current study. Tissues were obtained from PDAC patients who underwent surgery at Kagoshima University Hospital between 1991 and 2014. Normal pancreatic tissue specimens (n = 14) were collected from noncancerous tumour-adjacent tissue. Each surgical specimen was histologically classified according to the TNM classification system [50]. All patients in this study provided informed consent and the study protocol was approved by the Institutional Review Board of Kagoshima University. Patient information and clinicopathological features are shown in Table 1.
Two human PDAC cell lines were used in this study. PANC-1 cells were obtained from the RIKEN Cell Bank (Tsukuba, Ibaraki, Japan) and SW 1990 cells were obtained from the ATCC (Manassas, Virginia, USA).
Construction of the miRNA expression signature in PDAC
Total RNAs from 7 PDAC tissues and 4 normal pancreatic tissues were subjected to RNA sequencing. The construction procedure of miRNA signature included the following steps: small RNA isolation, cDNA library preparation and sequencing as described previously [14]
Quantitative real-time RT-PCR
Quantification of miRNA was performed using qRT-PCR as previously described [20, 28, 51, 52]. We used the following probes and primers: miR-216a-5p (product ID: 2220; Thermo Fisher Scientific, Waltham, MA, USA), miR-216a-3p (product ID: 475580_mat), miR-216b-5p (product ID: 2326), miR-216b-3p (product ID: 467177_mat), PLAU (product ID: Hs01547054_m1), GNB4 (product ID: Hs00222147_m1) and FOXQ1 (product ID: Hs00536425_s1. Human GUSB (product ID: Hs99999908_m1) and RNU48 (product ID: 001006) were used as internal controls.
Transfection of miRNA mimic and small interfering RNA (siRNA) into PDAC cell lines
PDAC cell lines were transfected with a miRNA mimic for gain-of-function experiments and siRNA for loss-of-function experiments. Pre-miR™ miRNA precursors for miR-216a-5p (product ID: PM10545; Thermo Fisher Scientific), miR-216a-3p (product ID: PM24316), miR-216b-5p (product ID: PM12302), miR-216b-3p (product ID: PM28358), negative control miRNA (product ID: AM 17111), two FOXQ1 siRNAs (product IDs: s41290 and s41291) and control siRNA (product ID: D-001810-10) were purchased from Thermo Fisher Scientific. The transfection efficiencies of miRNA in PANC-1 and SW 1990 cells were calculated as described in previous studies [20, 28, 51, 52].
Cell proliferation, migration and invasion assays
Cell proliferation, migration and invasion assays were described previously [20, 28, 51, 52].
Western blot analyses
Mouse anti-FOXQ1 antibodies (product ID: sc-166266; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) were diluted 1:500 for immunoblotting. Anti-GAPDH antibodies at a 1:1000 dilution (product ID: 010-25521; Wako, Osaka, Japan) were used as an internal loading control.
Immunohistochemistry
Tissue sections were incubated overnight at 4°C with anti-FOXQ1 antibodies diluted 1:300 (product ID: ab51340; Abcam, Cambridge, UK). Cytoplasmic staining of FOXQ1 in at least 1% of cancer cells was classified as high. If no cancer cells were stained, specimens were classified as negative for FOXQ1 staining. The expression of FOXQ1 was evaluated in 10 fields of 100 cells each using high-power microscopy (400×).
Genome-wide gene expression and in silico analyses
To identify miR-216b-3p target genes, a combination of genome-wide gene expression and in silico analyses was conducted as described previously [20, 28, 51, 52]. The microarray data were deposited into the Gene Omnibus database (GEO) repository under the accession number GSE82108. Next, we selected putative miRNA target genes using the TargetScanHuman (August, 2015 release, http://www.targetscan.org/vert_70) database.
Construction of plasmid vectors and dual luciferase reporter assays
Wild-type sequences of the 3׳-UTR of FOXQ1 containing the 2 miR-216b-3p target sites (positions 463-468 or 720-727) sequences or lacking a miR-216b-3p target site were inserted in the psiCHECK-2 vector (product ID: C8021; Promega, Madison, WI, USA). The procedure was described previously [20, 28, 51, 52].
Identification of downstream targets regulated by FOXQ1 in PDAC
Gene expression analyses of si-FOXQ1-transfected PANC-1 cells revealed molecular pathways and targets regulated by FOXQ1 in PDAC cells. This method was described in detail in previous studies [15, 20]. Microarray results were deposited in the GEO database (accession number GSE77790).
Statistical analysis
Relationships between expression values in 2 conditions or variables were analysed using the Mann-Whitney U test, and relationships between expression values in 3 conditions or variables were analysed using Bonferroni-adjusted Mann-Whitney U test. The correlation between the expression levels of miR-216b-3p and FOXQ1 was evaluated using Spearman's rank test. Associations between different categories were assessed using Fisher’s exact test and the chi-squared test. Overall survival (OS) after surgery was gauged using Kaplan–Meier curves. Patients were divided into two groups based on FOXQ1 expression, and differences in survival were estimated using the log-rank test. We used Expert StatView software (version 5.0 SAS Institute Inc., Cary, NC, USA) for these analyses.
ACKNOWLEDGMENTS
We wish to thank the Joint Research Laboratory, Kagoshima University Graduate School of Medical and Dental Sciences, for the use of their facilities. The present study was supported by KAKENHI(C) grant 15K10801, 26462067.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
1. Polireddy K, Chen Q. Cancer of the pancreas: molecular pathways and current advancement in treatment. J Cancer. 2016; 7:1497-1514.
2. Ilic M, Ilic I. Epidemiology of pancreatic cancer. World J Gastroenterol. 2016; 22:9694-9705.
3. Das S, Batra SK. Pancreatic cancer metastasis: are we being pre-EMTed? Curr Pharm Des. 2015; 21:1249-1255.
4. Castellanos JA, Merchant NB. Intensity of follow-up after pancreatic cancer resection. Ann Surg Oncol. 2014; 21:747-751.
5. Chiorean EG, Coveler AL. Pancreatic cancer: optimizing treatment options, new, and emerging targeted therapies. Drug Des Devel Ther. 2015; 9:3529-3545.
6. Ahn DH, Williams TM, Goldstein DA, El-Rayes B, Bekaii-Saab T. Adjuvant therapy for pancreas cancer in an era of value based cancer care. Cancer Treat Rev. 2016; 42:10-17.
7. Brunetti O, Russo A, Scarpa A, Santini D, Reni M, Bittoni A, Azzariti A, Aprile G, Delcuratolo S, Signorile M, Gnoni A, Palermo L, Lorusso V, Cascinu S, Silvestris N. MicroRNA in pancreatic adenocarcinoma: predictive/prognostic biomarkers or therapeutic targets? Oncotarget. 2015; 6:23323-23341.
8. Salmanidis M, Pillman K, Goodall G, Bracken C. Direct transcriptional regulation by nuclear microRNAs. Int J Biochem Cell Biol. 2014; 54:304-311.
9. Kurozumi A, Goto Y, Okato A, Ichikawa T, Seki N. Aberrantly expressed microRNAs in bladder cancer and renal cell carcinoma. J Hum Genet. 2017; 62:49-56.
10. Mizuno K, Mataki H, Seki N, Kumamoto T, Kamikawaji K, Inoue H. MicroRNAs in non-small cell lung cancer and idiopathic pulmonary fibrosis. J Hum Genet. 2017; 62:57-65.
11. Koshizuka K, Hanazawa T, Fukumoto I, Kikkawa N, Okamoto Y, Seki N. The microRNA signatures: aberrantly expressed microRNAs in head and neck squamous cell carcinoma. J Hum Genet. 2017; 62:3-13.
12. Yonemori K, Kurahara H, Maemura K, Natsugoe S. MicroRNA in pancreatic cancer. J Hum Genet. 2017; 62:33-40.
13. Fukumoto I, Kinoshita T, Hanazawa T, Kikkawa N, Chiyomaru T, Enokida H, Yamamoto N, Goto Y, Nishikawa R, Nakagawa M, Okamoto Y, Seki N. Identification of tumour suppressive microRNA-451a in hypopharyngeal squamous cell carcinoma based on microRNA expression signature. Br J Cancer. 2014; 111:386-394.
14. Itesako T, Seki N, Yoshino H, Chiyomaru T, Yamasaki T, Hidaka H, Yonezawa T, Nohata N, Kinoshita T, Nakagawa M, Enokida H. The microRNA expression signature of bladder cancer by deep sequencing: the functional significance of the miR-195/497 cluster. PLoS One. 2014; 9:e84311.
15. Fukumoto I, Hanazawa T, Kinoshita T, Kikkawa N, Koshizuka K, Goto Y, Nishikawa R, Chiyomaru T, Enokida H, Nakagawa M, Okamoto Y, Seki N. MicroRNA expression signature of oral squamous cell carcinoma: functional role of microRNA-26a/b in the modulation of novel cancer pathways. Br J Cancer. 2015; 112:891-900.
16. Goto Y, Kojima S, Nishikawa R, Kurozumi A, Kato M, Enokida H, Matsushita R, Yamazaki K, Ishida Y, Nakagawa M, Naya Y, Ichikawa T, Seki N. MicroRNA expression signature of castration-resistant prostate cancer: the microRNA-221/222 cluster functions as a tumour suppressor and disease progression marker. Br J Cancer. 2015; 113:1055-1065.
17. Fuse M, Kojima S, Enokida H, Chiyomaru T, Yoshino H, Nohata N, Kinoshita T, Sakamoto S, Naya Y, Nakagawa M, Ichikawa T, Seki N. Tumor suppressive microRNAs (miR-222 and miR-31) regulate molecular pathways based on microRNA expression signature in prostate cancer. J Hum Genet. 2012; 57:691-699.
18. Nohata N, Hanazawa T, Kikkawa N, Sakurai D, Fujimura L, Chiyomaru T, Kawakami K, Yoshino H, Enokida H, Nakagawa M, Katayama A, Harabuchi Y, Okamoto Y, Seki N. Tumour suppressive microRNA-874 regulates novel cancer networks in maxillary sinus squamous cell carcinoma. Br J Cancer. 2011; 105:833-841.
19. Kikkawa N, Hanazawa T, Fujimura L, Nohata N, Suzuki H, Chazono H, Sakurai D, Horiguchi S, Okamoto Y, Seki N. miR-489 is a tumour-suppressive miRNA target PTPN11 in hypopharyngeal squamous cell carcinoma (HSCC). Br J Cancer. 2010; 103:877-884.
20. Goto Y, Kurozumi A, Nohata N, Kojima S, Matsushita R, Yoshino H, Yamazaki K, Ishida Y, Ichikawa T, Naya Y, Seki N. The microRNA signature of patients with sunitinib failure: regulation of UHRF1 pathways by microRNA-101 in renal cell carcinoma. Oncotarget. 2016; 7:59070-59086. https://doi.org/10.18632/oncotarget.10887.
21. Matsushita R, Yoshino H, Enokida H, Goto Y, Miyamoto K, Yonemori M, Inoguchi S, Nakagawa M, Seki N. Regulation of UHRF1 by dual-strand tumor-suppressor microRNA-145 (miR-145-5p and miR-145-3p): inhibition of bladder cancer cell aggressiveness. Oncotarget. 2016; 7:28460-28487. https://doi.org/10.18632/oncotarget.8668.
22. Mataki H, Seki N, Mizuno K, Nohata N, Kamikawaji K, Kumamoto T, Koshizuka K, Goto Y, Inoue H. Dual-strand tumor-suppressor microRNA-145 (miR-145-5p and miR-145-3p) coordinately targeted MTDH in lung squamous cell carcinoma. Oncotarget. 2016; 7:72084-72098. https://doi.org/10.18632/oncotarget.12290.
23. Yonemori M, Seki N, Yoshino H, Matsushita R, Miyamoto K, Nakagawa M, Enokida H. Dual tumor-suppressors miR-139-5p and miR-139-3p targeting matrix metalloprotease 11 in bladder cancer. Cancer Sci. 2016; 107:1233-1242.
24. Anaya J. OncoLnc: linking TCGA survival data to mRNAs, miRNAs, and lncRNAs. PeerJ CompSci. 2016; 2:e67.
25. Ali S, Saleh H, Sethi S, Sarkar FH, Philip PA. MicroRNA profiling of diagnostic needle aspirates from patients with pancreatic cancer. Br J Cancer. 2012; 107:1354-1360.
26. Schultz NA, Werner J, Willenbrock H, Roslind A, Giese N, Horn T, Wojdemann M, Johansen JS. MicroRNA expression profiles associated with pancreatic adenocarcinoma and ampullary adenocarcinoma. Mod Pathol. 2012; 25:1609-1622.
27. Hong TH, Park IY. MicroRNA expression profiling of diagnostic needle aspirates from surgical pancreatic cancer specimens. Ann Surg Treat Res. 2014; 87:290-297.
28. Yonemori K, Seki N, Kurahara H, Osako Y, Idichi T, Arai T, Koshizuka K, Kita Y, Maemura K, Natsugoe S. ZFP36L2 promotes cancer cell aggressiveness and is regulated by antitumor microRNA-375 in pancreatic ductal adenocarcinoma. Cancer Sci. 2017; 108:124-135.
29. Hou BH, Jian ZX, Cui P, Li SJ, Tian RQ, Ou JR. miR-216a may inhibit pancreatic tumor growth by targeting JAK2. FEBS Lett. 2015; 589:2224-2232.
30. Deng M, Tang H, Zhou Y, Zhou M, Xiong W, Zheng Y, Ye Q, Zeng X, Liao Q, Guo X, Li X, Ma J, Li G. miR-216b suppresses tumor growth and invasion by targeting KRAS in nasopharyngeal carcinoma. J Cell Sci. 2011; 124:2997-3005.
31. Diederichs S, Haber DA. Dual role for argonautes in microRNA processing and posttranscriptional regulation of microRNA expression. Cell. 2007; 131:1097-1108.
32. Matranga C, Tomari Y, Shin C, Bartel DP, Zamore PD. Passenger-strand cleavage facilitates assembly of siRNA into Ago2-containing RNAi enzyme complexes. Cell. 2005; 123:607-620.
33. di Magliano MP, Logsdon CD. Roles for KRAS in pancreatic tumor development and progression. Gastroenterology. 2013; 144:1220-1229.
34. Eser S, Schnieke A, Schneider G, Saur D. Oncogenic KRAS signalling in pancreatic cancer. Br J Cancer. 2014; 111:817-822.
35. Azevedo-Pouly AC, Sutaria DS, Jiang J, Elgamal OA, Amari F, Allard D, Grippo PJ, Coppola V, Schmittgen TD. miR-216 and miR-217 expression is reduced in transgenic mouse models of pancreatic adenocarcinoma, knockout of miR-216/miR-217 host gene is embryonic lethal. Funct Integr Genomics. 2017; 17:203-212.
36. Rachagani S, Macha MA, Menning MS, Dey P, Pai P, Smith LM, Mo YY, Batra SK. Changes in microRNA (miRNA) expression during pancreatic cancer development and progression in a genetically engineered KrasG12D;Pdx1-Cre mouse (KC) model. Oncotarget. 2015; 6:40295-40309. https://doi.org/10.18632/oncotarget.5641.
37. Jackson BC, Carpenter C, Nebert DW, Vasiliou V. Update of human and mouse forkhead box (FOX) gene families. Hum Genomics. 2010; 4:345-352.
38. Uhlenhaut NH, Treier M. Forkhead transcription factors in ovarian function. Reproduction. 2011; 142:489-495.
39. Li Y, Zhang Y, Yao Z, Li S, Yin Z, Xu M. Forkhead box Q1: a key player in the pathogenesis of tumors (Review). Int J Oncol. 2016; 49:51-58.
40. Gonzalez DM, Medici D. Signaling mechanisms of the epithelial-mesenchymal transition. Sci Signal. 2014; 7:re8.
41. Diaz VM, Vinas-Castells R, Garcia de Herreros A. Regulation of the protein stability of EMT transcription factors. Cell Adh Migr. 2014; 8:418-428.
42. Feng J, Xu L, Ni S, Gu J, Zhu H, Wang H, Zhang S, Zhang W, Huang J. Involvement of FoxQ1 in NSCLC through regulating EMT and increasing chemosensitivity. Oncotarget. 2014; 5:9689-9702. https://doi.org/10.18632/oncotarget.2103.
43. Zhu Z, Zhu Z, Pang Z, Xing Y, Wan F, Lan D, Wang H. Short hairpin RNA targeting FOXQ1 inhibits invasion and metastasis via the reversal of epithelial-mesenchymal transition in bladder cancer. Int J Oncol. 2013; 42:1271-1278.
44. Xia L, Huang W, Tian D, Zhang L, Qi X, Chen Z, Shang X, Nie Y, Wu K. Forkhead box Q1 promotes hepatocellular carcinoma metastasis by transactivating ZEB2 and VersicanV1 expression. Hepatology. 2014; 59:958-973.
45. Zhang J, Liu Y, Zhang J, Cui X, Li G, Wang J, Ren H, Zhang Y. FOXQ1 promotes gastric cancer metastasis through upregulation of Snail. Oncol Rep. 2016; 35:3607-3613.
46. Geng SQ, Alexandrou AT, Li JJ. Breast cancer stem cells: multiple capacities in tumor metastasis. Cancer Lett. 2014; 349:1-7.
47. Bao B, Azmi AS, Aboukameel A, Ahmad A, Bolling-Fischer A, Sethi S, Ali S, Li Y, Kong D, Banerjee S, Back J, Sarkar FH. Pancreatic cancer stem-like cells display aggressive behavior mediated via activation of FoxQ1. J Biol Chem. 2014; 289:14520-14533.
48. Feng J, Zhang X, Zhu H, Wang X, Ni S, Huang J. FoxQ1 overexpression influences poor prognosis in non-small cell lung cancer, associates with the phenomenon of EMT. PLoS One. 2012; 7:e39937.
49. Zhao WG, Yu SN, Lu ZH, Ma YH, Gu YM, Chen J. The miR-217 microRNA functions as a potential tumor suppressor in pancreatic ductal adenocarcinoma by targeting KRAS. Carcinogenesis. 2010; 31:1726-1733.
50. Sobin LH, Gospodarowicz MK, Wittekind C. (2009). International Union Against Cancer (UICC) TNM Classification of Malignant Tumors. 7th ed. (Oxford, UK: Wiley-Blackwell).
51. Koshizuka K, Hanazawa T, Fukumoto I, Kikkawa N, Matsushita R, Mataki H, Mizuno K, Okamoto Y, Seki N. Dual-receptor (EGFR and c-MET) inhibition by tumor-suppressive miR-1 and miR-206 in head and neck squamous cell carcinoma. J Hum Genet. 2017; 62:113-121.
52. Osako Y, Seki N, Kita Y, Yonemori K, Koshizuka K, Kurozumi A, Omoto I, Sasaki K, Uchikado Y, Kurahara H, Maemura K, Natsugoe S. Regulation of MMP13 by antitumor microRNA-375 markedly inhibits cancer cell migration and invasion in esophageal squamous cell carcinoma. Int J Oncol. 2016; 49: 2255-2264.