
INTRODUCTION
Uveal melanoma is the most common primary intraocular tumor in adults, with about 1200–1500 new cases occurring per year in the United States [1, 2]. Although both uveal and cutaneous melanomas arise from melanocytes, uveal melanoma is biologically and genetically distinct from the more common cutaneous melanoma. Metastasis is a frequent occurrence in uveal melanoma, and outcomes are poor once distant spread occurs. It is estimated that 40–50% of uveal melanoma patients will die of metastatic disease, even with early diagnosis, proper treatment, and close follow-up [3]. By far the most common site of metastasis is the liver, reported in 87% of metastasis cases [4]. The management of localized uveal melanoma can be divided into globe-preserving therapy or enucleation. Globe-preserving therapies can broadly be classified into radiation, surgical, and laser therapy. The majority of primary uveal melanoma lesions in the United States are treated with plaque brachytherapy based upon results of the Collaborative Ocular Melanoma Study (COMS) trial, which randomized patients with medium-sized choroidal melanomas to primary therapy with 125I brachytherapy versus enucleation. No difference was observed in mortality between the two groups up to 15 years of follow-up [5]. Furthermore, no chemotherapeutic regimen or immunotherapy was demonstrated to be effective and at this stage they are not considered a clinically significant alternative to radiation or surgery. Bioinformatics as well as other methods are regression, classification or statistical methods used in the chemical and biological sciences helping in predict variables or in understanding patterns [6–8]. To this regard, a recent report using the L1000CDS2 web-based utility was able to predict small molecules and drugs targeting uveal melanoma gene signature [9]. In this bioinformatics study, cinnarizine (Figure 1), an anti-histaminic drug used for motion sickness, was proposed as a promising drug for the treatment of metastatic uveal melanoma [9]. Since cinnarizine significantly inhibited (+)-[3H]- (+)-pentazocine binding (IC50 = 162 ± 28 nM), and a QSAR study predicted a high Sigma-2 (σ2) receptor affinity, we hypothesized that Sigma (σ) receptors could play also a role in uveal melanoma progression [10–13].
Figure 1: Chemical structures of σ receptor ligands (+)-Pentazocine, Cinnarizine, Haloperidol, and Haloperidol metabolite II.
Sigma receptors, first introduced as subtypes of the opioid receptors, are now considered unique class of proteins. There exist two different σ receptors namely Sigma-1 (σ1) and σ2 receptors distinguished by structure, biological functions and ligands sensitivity. Indeed, σ1 binding sites display high affinity for dextro benzomorphan enantiomers like (+)-pentazocine, while opioid receptors bind levo isomers [14–16]. Small molecules that bind to the σ receptor (σ ligands or σ-modulating drugs) have many potential functions and are used in diagnostic tumor imaging [17–23]. Such compounds are also able to modify the growth rates of human cancer cell lines both in vitro and in vivo. Several lines of evidence suggest that both receptors are involved in different biological functions including cell proliferation and survival. Interestingly, previous reports showed that σ1 receptor is involved in apoptosis because of its location at the mitochondria-associated membranes [24–31]. Similarly, a previous study showed that both sigma receptors are also associated to autophagy in multiple myeloma cells [32].
Taken all together, these observations suggest that σ receptor biological significance is not fully elucidated and may be dependent on the experimental conditions. Therefore, the aim of the present study was to evaluate the pharmacological effects of σ receptor ligands with different pharmacological profile such as haloperidol, haloperidol metabolite II, and (+)-pentazocine (Figure 1, Table 1) on apoptosis and autophagy in uveal melanoma. Our results showed that all tested compounds promote apoptosis and autophagy whereas the σ1 receptor agonist (+)-pentazocine has opposite effects on cell proliferation with respect to haloperidol and haloperidol metabolite II, which possess σ1 antagonist/σ2 ligand profiles.
Table 1: Binding profile of haloperidol, haloperidol metabolite II, and (+)-pentazocine
Ki (nM) ± S.E.M. |
||||
|---|---|---|---|---|
σ1 |
σ2 |
D2 |
D3 |
|
Haloperidola |
2.7 ± 0.5 |
17.0 ± 1.5 |
2.5 ± 0.7 |
6.1 ± 1.5 |
Haloperidol metabolite IIa |
2.9 ± 0.8 |
2.4 ± 0.5 |
241 ± 38 |
1024 ± 217 |
(+)-Pentazocineb |
5.0 ± 1.0 |
1824 ± 36 |
− |
− |
aReference [28].
bReference [52].
RESULTS
Effect of pharmacological treatments on cell proliferation and migration
Our results showed that haloperidol treatment resulted in a dose dependent effect on cell proliferation (Figure 2A). Interestingly, haloperidol treatment at 1 μM resulted in a significant increase (p < 0.01) of cell proliferation when compared to control over a 72 h period. Interestingly all the other concentrations (10, and 25 μM) resulted in a significant (p < 0.01) decrease of cell proliferation when compared to control. Conversely, haloperidol metabolite II (Figure 2B) showed a significant decrease of cell proliferation at all concentrations when compared to control (p < 0.01). By contrast, continuous cell proliferation analysis showed that (+)-pentazocine (Figure 2C) resulted in a dose dependent increase of cell proliferation when compared to control. In particular, the concentrations exhibiting higher proliferative effects were 1 and 25 μM (p < 0.01). These two concentrations exhibited their effect following 24 h treatment whereas 10 μM was able to induce proliferation at a lower extent following 48 h.
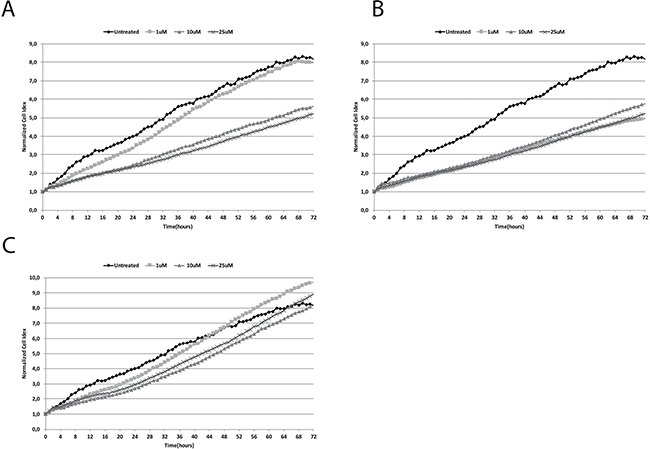
Figure 2: Real time cell proliferation monitoring by xCELLigence system following treatments with (A) (+)-Pentazocine, (B) Haloperidol and (C) Haloperidol metabolite II. Cell index values were normalized at the time of pharmacological treatments in order to obtain a normalized cell index. Each line is expressing the average of four different experiments.
Interestingly, all tested compounds showed significant decrease of cell migration as measured by wound healing assay (Figure 3A–3C).
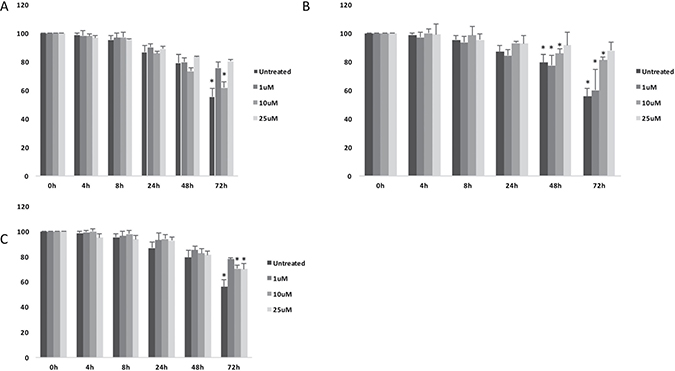
Figure 3: Cell migration analysis following treatments with (A) (+)-Pentazocine, (B) Haloperidol and (C) Haloperidol metabolite II. Values are presented as percentage of the open wound following 4, 8, 24, 48 and 72 hours (wound at time 0 was assumed as 100% and used as control). Values are expressed as the mean ± SD of four different experiments. (*p < 0.01 vs control).
Effect of pharmacological treatments on colony formation capacity
These results were further confirmed in part by clonogenic assay (Figure 4A and 4B) showing that haloperidol was able to induce colonies formation at 1 μM whereas had an opposite effect at 10 and 25 μM (p < 0.05). Similarly, all haloperidol metabolite II concentrations were able to decrease colonies formations when compared to control (Figure 4A and 4B). By contrast, (+)-pentazocine treatment resulted in a significant increase in colonies formation at 1 and 10 μM whereas it exhibited opposite results at 25 μM (Figure 4A and 4B).
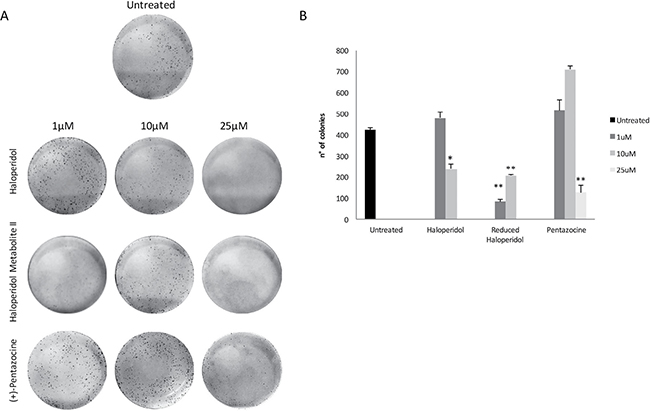
Figure 4: Colony formation capacity following treatments with (+)-Pentazocine, Haloperidol and Haloperidol metabolite II. (A) Images are representative of four separate experiments and (B) number of colonies were manually counted and presented as the mean ± SD of four independent experiments. (*p < 0.01 vs control).
Effect of pharmacological treatments on cell autophagy
Our results showed that all pharmacological treatments significantly increased cell autophagy. In particular, haloperidol treatment resulted in a significant dose and time dependent increase of autophagy peaking at 4 h and 8 h (Figure 5A, 5B and Figure 8). Haloperidol metabolite II exhibited also a pro-autophagic activity peaking at 4 h and 8 h, however we did not observe a dose and time dependent effects of various tested concentrations (Figure 6A, 6B and Figure 8). Finally, (+)-pentazocine treatment resulted in a significant dose and time dependent increase of autophagy peaking at 4 h and 8 h (Figure 7A, 7B and Figure 8).
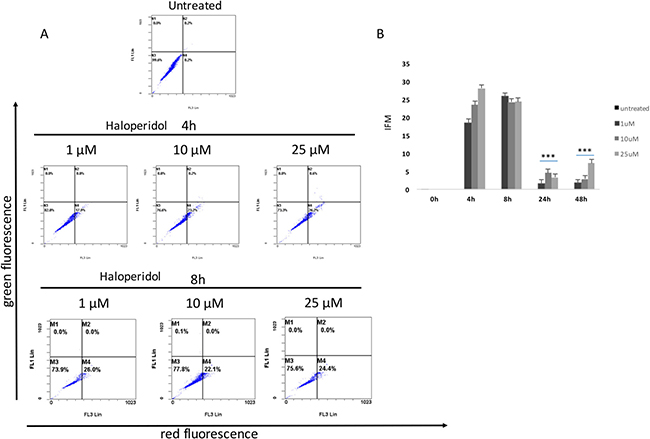
Figure 5: Cytofluorimetric analysis of cell autophagy following Haloperidol treatment at different concentrations and time points. (A) Blots are representative of three independent experiments; (B) percentage of autophagic cells are presented as the mean ± SD of three independent experiments. (*p < 0.01 vs control).
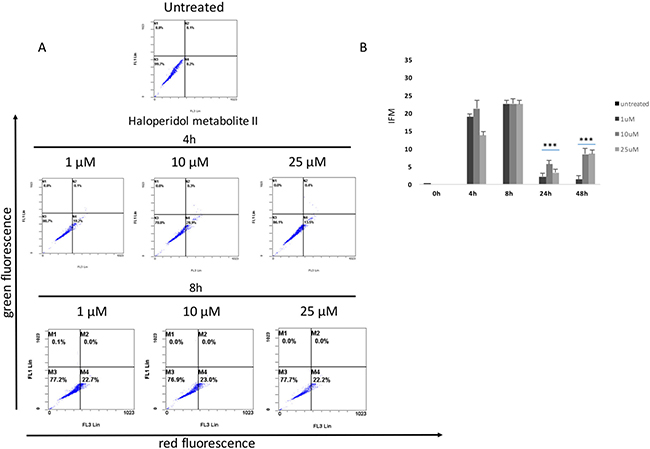
Figure 6: Cytofluorimetric analysis of cell autophagy following Haloperidol metabolite II treatment at different concentrations and time points. (A) Blots are representative of three independent experiments; (B) percentage of autophagic cells are presented as the mean ± SD of three independent experiments. (*p < 0.01 vs control).
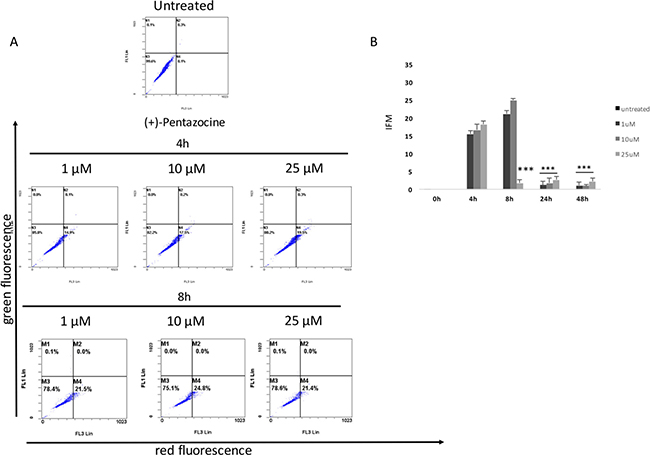
Figure 7: Cytofluorimetric analysis of cell autophagy following (+)-pentazocine treatment at different concentrations and time points. (A) Blots are representative of three independent experiments; (B) percentage of autophagic cells are presented as the mean ± SD of three independent experiments. (*p < 0.01 vs control).
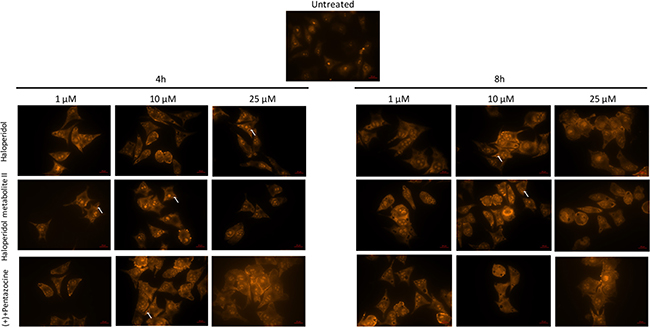
Figure 8: Microscopy analysis of cell autophagy following various pharmacological treatments and time points. The protonated form of acridine orange accumulates in acidic compartments and forms aggregates, which are characterized by red/orange fluorescence. Arrows indicate autophagic vacuoles. Images were obtained with a 40× magnification.
Effect of pharmacological treatments on cell apoptosis
Similarly to autophagy, all pharmacological treatments induced apoptosis at all tested concentrations even though such effect was evident at later time points when compared to autophagy (24 h, and 48 h). In particular, haloperidol was apoptotic at all tested concentrations peaking at 24 h (Figure 9A and 9B). In addition, haloperidol metabolite II induced apoptosis at all tested concentrations peaking at 24 h and exhibited increased late apoptotic cell death at 48 h compared to control and haloperidol (p < 0.01 for both) (Figure 10A and 10B). Finally, (+)-pentazocine exhibited an apoptotic profile peaking at 24 h and where it was significantly higher at 10 and 25 μM (p < 0.01 compared to control and 1 μM) (Figure 11A and 11B).
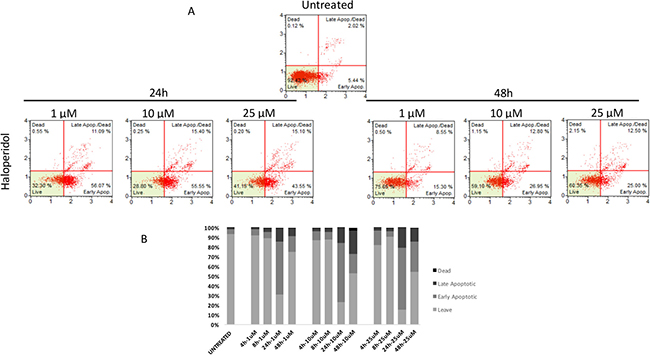
Figure 9: Cytofluorimetric analysis of cell apoptosis following Haloperidol treatment at different concentrations and time points. (A) Blots are representative of three independent experiments; (B) percentage of viable, early and late apoptotic cells and necrotic cells are presented as the mean ± SD of three independent experiments.
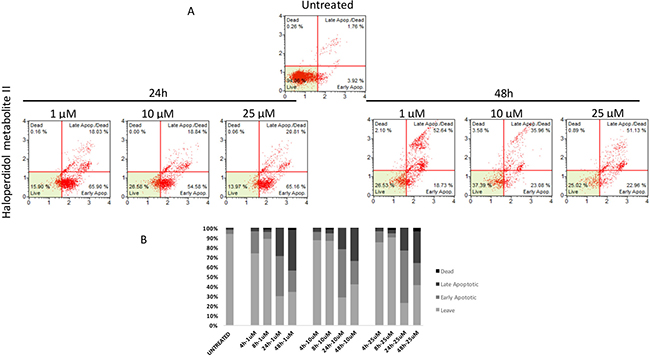
Figure 10: Cytofluorimetric analysis of cell apoptosis following Haloperidol metabolite II treatment at different concentrations and time points. (A) Blots are representative of three independent experiments; (B) percentage of viable, early and late apoptotic cells and necrotic cells are presented as the mean ± SD of three independent experiments.
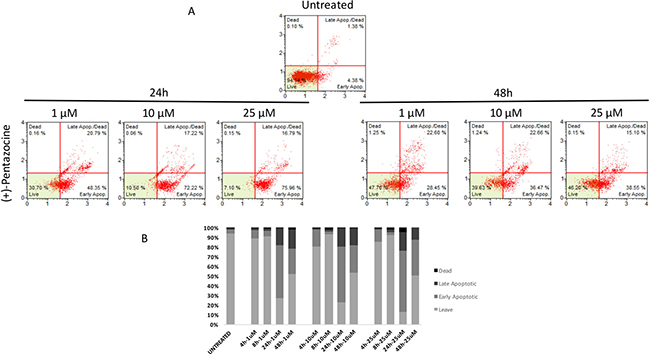
Figure 11: Cytofluorimetric analysis of cell apoptosis following (+)-Pentazocine treatment at different concentrations and time points. (A) Blots are representative of three independent experiments; (B) percentage of viable, early and late apoptotic cells and necrotic cells are presented as the mean ± SD of three independent experiments.
DISCUSSION
Uveal melanoma is a rare subset of melanoma, representing approximately 3–5% of all melanomas and with an incidence of approximately six per million per year in the United States. Although considered a rare tumor, uveal melanoma is the most common primary ocular malignancy in adults and accounts for 85–95% of all ocular melanoma cases. Despite excellent rates of local disease control with surgery or radiotherapy, nearly 50% of patients with uveal melanoma will develop metastatic disease within 15 years from initial diagnosis. Therefore the aim of the present study was to evaluate the “druggability” of σ receptors as possible target for uveal melanoma treatment.
In the first set of experiment we showed that prototypical σ ligands such as (+)-pentazocine (a putative σ1 agonist activity) or haloperidol and its reduced metabolite (with σ1 antagonist and σ2 agonist properties) exhibited different effect on cell proliferation compared to σ1 agonist. In particular, the σ1 agonist (+)-pentazocine, induced cancer cell proliferation whereas haloperidol and haloperidol metabolite II inhibited cancer growth. Noteworthy, lowest concentration of haloperidol (1 μM) treatment resulted in a significant increase of cell proliferation compared to the other tested concentrations or its metabolite. To this regard, different receptor affinity should be taken into due account for haloperidol and its metabolite. Indeed, haloperidol exhibits a high affinity for both σ receptor subtypes as well as for dopaminergic, adrenergic, serotoninergic and histaminergic receptors [33] whereas haloperidol metabolite II has significant higher affinity and selectivity for σ receptor. Therefore, it is conceivable that haloperidol at low concentrations may exhibit additional non-specific effects.
Our results are consistent with previous studies showing that σ2 receptor ligands are associated with proliferation inhibition of cancer cells [34, 35] whereas under our experimental conditions σ1 receptor ligand treatment resulted in an increased cell proliferation. These results were partially confirmed by clonogenic assay, which however showed a significant reduction of colony formation capacity following 25 μM (+)-pentazocine treatment. This discrepancy may be related to the different experimental conditions used for the two assays used. In particular, live continuous cell monitoring with xCELLingence analysis allows cell proliferation to be assessed over 72 h time frame, whereas for clonogenic assay, cell are grown at very low confluency and for longer time (10 days). Therefore, it is not possible to exclude that high concentrations of (+)-pentazocine may have different effects on cell proliferation following chronic treatment. Our results are of particular interest, since previous reports showed similar effect of σ1 and σ2 ligands on cell proliferation whereas we are showing that in uveal melanoma they may have opposite effects on cell proliferation. However, since this is the first report characterizing the σ receptor system in uveal melanoma, we cannot exclude that this might be a cell specific effect. Consistently with this hypothesis is the evidence that even the same receptor mediates opposite pharmacological effects under various experimental conditions. Futhermore, it should be taken into due account that haloperidol and its metabolite II, beside being σ2 ligands, share common σ1 antagonist activity. In particular, previous reports suggested that σ1 may serve a chaperone molecule, possesses various oligomerization states and different intracellular localization compared to σ2 receptor [36, 37]. As far as concern cell migration, our data are consistent with previous reports showing that both sigma receptor ligands significantly reduces cell migration under various experimental conditions [28, 38, 39].
Furthermore, we showed that both σ1 ligand and mixed σ1 antagonist/σ2 agonists share the same effects on autophagy and apoptosis. Several chemotherapeutic agents have been shown to induce autophagy [40]. However, in many cases it remains unclear whether cell death occurs by autophagy, whether cell death is associated with autophagy, or whether autophagy is a survival response to cytotoxic chemotherapy [41–44]. Emerging data suggest that autophagy participates in integrated responses to cellular stress that determine cell death versus survival. Interestingly, we showed that all tested σ receptor ligands induce autophagy following 4 h and 8 h treatments as measured by cytofluorimetric and microscopy analysis. Our results are consistent with previous reports showing that both σ receptors mediate autophagy under various experimental conditions. In particular, under our experimental conditions, autophagy may represent a mechanism related to cell toxicity rather than a resistance mechanism since apoptosis occurs at later time points (24 h, and 48 h). Our data are consistent with a previous study showing that both sigma receptors are also associated to autophagy in multiple myeloma cells [32]. Consistently with a previous report [45], our data demonstrate that σ receptor ligands induce cell death by multiple signaling pathways and that both autophagy and apoptosis coexist, even though at different time points, following σ receptor ligands treatment. Finally, both cell death mechanisms seem to be cell specific since under different experimental conditions σ receptor ligands do not induce either apoptosis or autophagy [46].
Taken all together, our data suggest that both σ receptors are able to induce apoptosis and autophagy in uveal melanoma but they exert different effects on cell proliferation. In conclusion, σ receptors represent a valuable target for uveal melanoma treatment and additional studies are now warranted in order to fully elucidate the molecular mechanisms downstream receptor activation in order to elucidate their possible translation into a clinical setting.
MATERIALS AND METHODS
Cell culture and pharmacological treatments
Human uveal melanoma cells (92.1) were purchased from ATCC Company (Milan, Italy). Cells were suspended in RPMI1640 culture medium containing 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 U/mL streptomycin). At 80% confluency, cells were passaged using trypsin-EDTA solution (0.05% trypsin and 0.02% EDTA) [47]. (+)-Pentazocine (Sigma–Aldrich, Milan, Italy), haloperidol, and haloperidol metabolite II were added separately to cell culture of all experiments at different final concentrations of 1, 10, and 25 μM.
Clonogenic assay
Colony assays performed by seeding cells in 6 wells plates at low density (3000 cells/well) and allowing growth for 10 days. Colonies were fixed, stained with crystal violet and counted.
Annexin V and dead cell evaluation by cytofluorimetric analysis
Cell apoptosis was evaluated by Muse™ Annexin V & Dead cell kit (Catalog No. MCH100105, Millipore, Milan, Italy) according to the manufacture’s guidelines. Briefly, 100 μl of the Muse™ Annexin V & Dead Cell Reagent to 100 μl of cell suspension. Such preparation was mixed thoroughly by vortexing at a medium speed for 3–5 s and samples were allowed to stain for 20 min at room temperature in the dark. Samples were read by Muse™ Cell Analyzer (Millipore).
Effects of pharmacological treatments on cell migration
Cell migration was studied by employing the “wound healing” assay. Briefly, cells were seeded in 24 wells dishes and cultured until confluence. Cells were treated with vehicle, (+)-pentazocine, haloperidol or haloperidol metabolite II and were then scraped with a 200 μl micropipette tip and monitored at 0 h, 4 h, 8 h, 24, 48 h, and 72 h. The uncovered wound area was measured and quantified at different intervals with ImageJ 1.37v (NIH).
Real time cell proliferation monitoring by xCELLigence system
xCELLigence experiments were performed using the RTCA (Real-Time Cell Analyzer) DP (Dual Plate) instrument according to manufacturers’ instructions (Roche Applied Science, Mannheim, Germany and ACEA Biosciences, San Diego, CA). The RTCA DP Instrument includes three main components: (i) RTCA DP Analyzer, which is placed inside a humidified incubator maintained at 37°C and 5% CO2, (ii) RTCA Control Unit with RTCA Software preinstalled, and (iii) E-Plate 16 for proliferation assay. First, the optimal seeding number was determined by cell titration and growth experiments. After seeding the optimal cell number (5000 cells/well), cells were automatically monitored every 15 min [48, 49]. Optimal cell number was determined in a preliminary set of experiments (data not shown) in order to obtain a significant cell index value and a constant cell growth during the entire duration of the experiment. Cells were treated with the compounds about 8h after seeding, when the cells were in the log growth phase.
Cytofluorimetric and microscopy analysis of autophagy
Human uveal melanoma cell line was incubated with 3 μL of acridine orange (Sigma, St. Louis, MO, USA) at a final concentration of 1 μM for 15 min at 37°C at room temperature in the dark. Following washing with phosphate buffer solution (PBS), the acidic vacuoles were detected using a fluorescence microscope. Autophagic cells contained bright red cytoplasmic particles. Formation of acidic vesicular organelles was quantified also by flow cytometry following acridine orange staining. Briefly, cells were fixed with methanol for 3 minutes and rinsed with ware before acridine orange (1 μM in acetic acid) staining for 15 minutes. Finally, cell were rinsed with water andcells were analyzed by cytometry. In particular, the appropriate isotopic control was also included and labeled cells were acquired using a Beckman Coulter FC-500 flow cytometer [50, 51].
Statistical analysis
The data were expressed as the means ± SD. Statistical analysis was performed via one-way analysis of variance (ANOVA) using SPSS11.0 software. p < 0.05 was considered to be significant.
Abbreviations
Collaborative Ocular Melanoma Study (COMS); Fetal Bovine Serum (FBS); Non-small cell lung carcinoma (NSCLC); Phosphate Buffer Solution (PBS); Real-Time Cell Analyzer (RTCA).
Authors’ contributions
LL, CCC, DT, GR, NLP and GLV performed and designed the experiments, analyzed the data and contributed to the manuscript writing. RA, MV, MR, AL, AR designed the experiments, provided the clinical background and contributed to the manuscript writing. OP, AM and EA provided sigma ligands, designed the experiments, analyzed the data and contributed to the manuscript writing.
CONFLICTS OF INTEREST
All the Authors declare to have no conflicts of interest
FUNDING
This work was supported by Fondazione “Mani Amiche” Onlus (Catania, Italy) and University of Catania (FIR2014) (Catania, Italy).
REFERENCES
1. Chang AE, Karnell LH, Menck HR. The National Cancer Data Base report on cutaneous and noncutaneous melanoma: a summary of 84,836 cases from the past decade. The American College of Surgeons Commission on Cancer and the American Cancer Society. Cancer. 1998; 83:1664–1678.
2. Singh AD, Topham A. Incidence of uveal melanoma in the United States: 1973–1997. Ophthalmology. 2003; 110:956–961.
3. Kujala E, Makitie T, Kivela T. Very long-term prognosis of patients with malignant uveal melanoma. Invest Ophthalmol Vis Sci. 2003; 44:4651–4659.
4. Kath R, Hayungs J, Bornfeld N, Sauerwein W, Hoffken K, Seeber S. Prognosis and treatment of disseminated uveal melanoma. Cancer. 1993; 72:2219–2223.
5. Collaborative Ocular Melanoma Study G. The COMS randomized trial of iodine 125 brachytherapy for choroidal melanoma: V. Twelve-year mortality rates and prognostic factors: COMS report No. 28. Arch Ophthalmol. 2006; 124:1684–1693.
6. Amata E, Xi H, Colmenarejo G, Gonzalez-Diaz R, Cordon-Obras C, Berlanga M, Manzano P, Erath J, Roncal NE, Lee PJ, Leed SE, Rodriguez A, Sciotti RJ, et al. Identification of “Preferred” Human Kinase Inhibitors for Sleeping Sickness Lead Discovery. Are Some Kinases Better than Others for Inhibitor Repurposing? ACS infectious diseases. 2016; 2:180–186.
7. Diaz R, Luengo-Arratta SA, Seixas JD, Amata E, Devine W, Cordon-Obras C, Rojas-Barros DI, Jimenez E, Ortega F, Crouch S, Colmenarejo G, Fiandor JM, Martin JJ, et al. Identification and characterization of hundreds of potent and selective inhibitors of Trypanosoma brucei growth from a kinase-targeted library screening campaign. PLoS Negl Trop Dis. 2014; 8:e3253.
8. Bao QX, Zhao MY, Chen L, Wang Y, Wu SY, Wu WC, Liu XJ. MicroRNA-297 promotes cardiomyocyte hypertrophy via targeting sigma-1 receptor. Life Sci. 2017; 175:1–10.
9. Fagone P, Caltabiano R, Russo A, Lupo G, Anfuso CD, Basile MS, Longo A, Nicoletti F, De Pasquale R, Libra M, Reibaldi M. Identification of novel chemotherapeutic strategies for metastatic uveal melanoma. Sci Rep. 2017; 7:44564.
10. Basile AS, Paul IA, Mirchevich A, Kuijpers G, De Costa B. Modulation of (+)-[3H]pentazocine binding to guinea pig cerebellum by divalent cations. Mol Pharmacol. 1992; 42:882–889.
11. Rescifina A, Floresta G, Marrazzo A, Parenti C, Prezzavento O, Nastasi G, Dichiara M, Amata E. Development of a Sigma-2 Receptor affinity filter through a Monte Carlo based QSAR analysis. Eur J Pharm Sci. 2017; 106:94–101.
12. Rescifina A, Floresta G, Marrazzo A, Parenti C, Prezzavento O, Nastasi G, Dichiara M, Amata E. Sigma-2 receptor ligands QSAR model dataset. Data in Brief. 2017; 13:514–535.
13. Nastasi G, Miceli C, Pittala V, Modica MN, Prezzavento O, Romeo G, Rescifina A, Marrazzo A, Amata E. S2RSLDB: a comprehensive manually curated, internet-accessible database of the sigma-2 receptor selective ligands. J Cheminform. 2017; 9:3.
14. Pasquinucci L, Prezzavento O, Marrazzo A, Amata E, Ronsisvalle S, Georgoussi Z, Fourla DD, Scoto GM, Parenti C, Arico G, Ronsisvalle G. Evaluation of N-substitution in 6,7-benzomorphan compounds. Bioorg Med Chem. 2010; 18:4975–4982.
15. Parenti C, Turnaturi R, Arico G, Marrazzo A, Prezzavento O, Ronsisvalle S, Scoto GM, Ronsisvalle G, Pasquinucci L. Antinociceptive profile of LP1, a non-peptide multitarget opioid ligand. Life Sci. 2012; 90:957–961.
16. Prezzavento O, Campisi A, Parenti C, Ronsisvalle S, Arico G, Arena E, Pistolozzi M, Scoto GM, Bertucci C, Vanella A, Ronsisvalle G. Synthesis and resolution of cis-(+/–)-methyl (1R,2S/1S,2R)-2-[(4-hydroxy-4-phenylpiperidin-1-yl)methyl]-1-(4-methylphenyl)cycl opropanecarboxylate [(+/–)-PPCC)]: new sigma receptor ligands with neuroprotective effect. J Med Chem. 2010; 53:5881–5885.
17. Schinina B, Martorana A, Colabufo NA, Contino M, Niso M, Perrone MG, De Guidi G, Catalfo A, Rappazzo G, Zuccarello E, Prezzavento O, Amata E, Rescifina A, Marrazzo A. 4-Nitro-2,1,3-benzoxadiazole derivatives as potential fluorescent sigma receptor probes. RSC Advances. 2015; 5:47108–47116.
18. Mach RH, Zeng C, Hawkins WG. The sigma2 receptor: a novel protein for the imaging and treatment of cancer. J Med Chem. 2013; 56:7137–7160.
19. ClinicalTrials.gov. [18F]ISO-1 PET/CT in Breast Cancer. 2014.
20. ClinicalTrials.gov. Imaging of in Vivo Sigma-2 Receptor Expression With 18F-ISO-1 Positron Emission Tomography in Metastatic Breast Cancer. 2016.
21. ClinicalTrials.gov. Assessment of Cellular Proliferation in Tumors by Positron Emission Tomography (PET) Using 18F ISO-1. 2009.
22. Ronsisvalle S, Arico G, Cova AM, Blanco P, Amata E, Pappalardo M, Pasquinucci L, Spadaro A, Ronsisvalle N. Caspase-3 activation in human melanoma A375 cell line by a novel selective sigma-2 agonist. Pharmazie. 2016; 71:146–151.
23. Huang YS, Lu HL, Zhang LJ, Wu ZW. Sigma-2 Receptor Ligands and Their Perspectives in Cancer Diagnosis and Therapy. Med Res Rev. 2014; 34:532–566.
24. Su TP, Hayashi T, Maurice T, Buch S, Ruoho AE. The sigma-1 receptor chaperone as an inter-organelle signaling modulator. Trends Pharmacol Sci. 2010; 31:557–566.
25. Maurice T. Improving Alzheimer’s disease-related cognitive deficits with sigma(1) receptor Agonists. Drug News Perspect. 2002; 15:617–625.
26. Chu UB, Ruoho AE. Biochemical Pharmacology of the Sigma-1 Receptor. Mol Pharmacol. 2016; 89:142–153.
27. Prezzavento O, Arena E, Sanchez-Fernandez C, Turnaturi R, Parenti C, Marrazzo A, Catalano R, Amata E, Pasquinucci L, Cobos EJ. (+)-and (–)-Phenazocine enantiomers: Evaluation of their dual opioid agonist/sigma1 antagonist properties and antinociceptive effects. Eur J Med Chem. 2017; 125:603–610.
28. Olivieri M, Amata E, Vinciguerra S, Fiorito J, Giurdanella G, Drago F, Caporarello N, Prezzavento O, Arena E, Salerno L, Rescifina A, Lupo G, Anfuso CD, Marrazzo A. Antiangiogenic Effect of (+/–)-Haloperidol Metabolite II Valproate Ester [(+/–)-MRJF22] in Human Microvascular Retinal Endothelial Cells. J Med Chem. 2016; 59:9960–9966.
29. Mesangeau C, Amata E, Alsharif W, Seminerio MJ, Robson MJ, Matsumoto RR, Poupaert JH, McCurdy CR. Synthesis and pharmacological evaluation of indole-based sigma receptor ligands. Eur J Med Chem. 2011; 46:5154–5161.
30. Marrazzo A, Fiorito J, Zappala L, Prezzavento O, Ronsisvalle S, Pasquinucci L, Scoto GM, Bernardini R, Ronsisvalle G. Antiproliferative activity of phenylbutyrate ester of haloperidol metabolite II [(+/–)-MRJF4] in prostate cancer cells. Eur J Med Chem. 2011; 46:433–438.
31. Parenti C, Marrazzo A, Arico G, Parenti R, Pasquinucci L, Ronsisvalle S, Ronsisvalle G, Scoto GM. The antagonistic effect of the sigma 1 receptor ligand (+)-MR200 on persistent pain induced by inflammation. Inflamm Res. 2014; 63:231–237.
32. Korpis K, Weber F, Brune S, Wunsch B, Bednarski PJ. Involvement of apoptosis and autophagy in the death of RPMI 8226 multiple myeloma cells by two enantiomeric sigma receptor ligands. Bioorg Med Chem. 2014; 22:221–233.
33. Richelson E, Souder T. Binding of antipsychotic drugs to human brain receptors focus on newer generation compounds. Life Sci. 2000; 68:29–39.
34. Niso M, Abate C, Contino M, Ferorelli S, Azzariti A, Perrone R, Colabufo NA, Berardi F. Sigma-2 receptor agonists as possible antitumor agents in resistant tumors: hints for collateral sensitivity. ChemMedChem. 2013; 8:2026–2035.
35. Aydar E, Onganer P, Perrett R, Djamgoz MB, Palmer CP. The expression and functional characterization of sigma (sigma) 1 receptors in breast cancer cell lines. Cancer Lett. 2006; 242:245–257.
36. Schmidt HR, Zheng S, Gurpinar E, Koehl A, Manglik A, Kruse AC. Crystal structure of the human sigma1 receptor. Nature. 2016; 532:527–530.
37. Balasuriya D, D’Sa L, Talker R, Dupuis E, Maurin F, Martin P, Borgese F, Soriani O, Edwardson JM. A direct interaction between the sigma-1 receptor and the hERG voltage-gated K+ channel revealed by atomic force microscopy and homogeneous time-resolved fluorescence (HTRF(R)). The Journal of biological chemistry. 2014; 289:32353–32363.
38. Heiss K, Raffaele M, Vanella L, Murabito P, Prezzavento O, Marrazzo A, Arico G, Castracani CC, Barbagallo I, Zappala A, Avola R, Li Volti G. (+)-Pentazocine attenuates SH-SY5Y cell death, oxidative stress and microglial migration induced by conditioned medium from activated microglia. Neurosci Lett. 2017; 642:86–90.
39. Moritz C, Berardi F, Abate C, Peri F. Live imaging reveals a new role for the sigma-1 (sigma1) receptor in allowing microglia to leave brain injuries. Neurosci Lett. 2015; 591:13–18.
40. Rubinsztein DC, Gestwicki JE, Murphy LO, Klionsky DJ. Potential therapeutic applications of autophagy. Nat Rev Drug Discov. 2007; 6:304–312.
41. Levine B, Klionsky DJ. Development by self-digestion: molecular mechanisms and biological functions of autophagy. Dev Cell. 2004; 6:463–477.
42. Hippert MM, O’Toole PS, Thorburn A. Autophagy in cancer: good, bad, or both? Cancer Res. 2006; 66:9349–9351.
43. Levine B, Kroemer G. Autophagy in the pathogenesis of disease. Cell. 2008; 132:27–42.
44. Mathew R, Karp CM, Beaudoin B, Vuong N, Chen G, Chen HY, Bray K, Reddy A, Bhanot G, Gelinas C, Dipaola RS, Karantza-Wadsworth V, White E. Autophagy suppresses tumorigenesis through elimination of p62. Cell. 2009; 137:1062–1075.
45. Zeng C, Rothfuss J, Zhang J, Chu W, Vangveravong S, Tu Z, Pan F, Chang KC, Hotchkiss R, Mach RH. Sigma-2 ligands induce tumour cell death by multiple signalling pathways. Br J Cancer. 2012; 106:693–701.
46. Megalizzi V, Mathieu V, Mijatovic T, Gailly P, Debeir O, De Neve N, Van Damme M, Bontempi G, Haibe-Kains B, Decaestecker C, Kondo Y, Kiss R, Lefranc F. 4-IBP, a sigma1 receptor agonist, decreases the migration of human cancer cells, including glioblastoma cells, in vitro and sensitizes them in vitro and in vivo to cytotoxic insults of proapoptotic and proautophagic drugs. Neoplasia. 2007; 9:358–369.
47. Sacerdoti D, Colombrita C, Ghattas MH, Ismaeil EF, Scapagnini G, Bolognesi M, Li Volti G, Abraham NG. Heme oxygenase-1 transduction in endothelial cells causes downregulation of monocyte chemoattractant protein-1 and of genes involved in inflammation and growth. Cell Mol Biol (Noisy-le-grand). 2005; 51:363–370.
48. Tibullo D, Barbagallo I, Giallongo C, La Cava P, Parrinello N, Vanella L, Stagno F, Palumbo GA, Li Volti G, Di Raimondo F. Nuclear translocation of heme oxygenase-1 confers resistance to imatinib in chronic myeloid leukemia cells. Curr Pharm Des. 2013; 19:2765–2770.
49. Li Volti G, Salomone S, Sorrenti V, Mangiameli A, Urso V, Siarkos I, Galvano F, Salamone F. Effect of silibinin on endothelial dysfunction and ADMA levels in obese diabetic mice. Cardiovasc Diabetol. 2011; 10:62.
50. Torella D, Leosco D, Indolfi C, Curcio A, Coppola C, Ellison GM, Russo VG, Torella M, Li Volti G, Rengo F, Chiariello M. Aging exacerbates negative remodeling and impairs endothelial regeneration after balloon injury. Am J Physiol Heart Circ Physiol. 2004; 287:H2850–2860.
51. Salomone F, Barbagallo I, Puzzo L, Piazza C, Li Volti G. Efficacy of adipose tissue-mesenchymal stem cell transplantation in rats with acetaminophen liver injury. Stem Cell Res. 2013; 11:1037–1044.
52. Ghandi M, Sherafat F, Sadeghzadeh M, Alirezapour B. One-pot synthesis and sigma receptor binding studies of novel spirocyclic-2,6-diketopiperazine derivatives. Bioorg Med Chem Lett. 2016; 26:2676–2679.