
INTRODUCTION
With increasing numbers of patients deferring definitive therapy for prostate cancer in favor of active surveillance, there is an unmet need for molecular biomarkers that help to distinguish indolent and lethal prostate tumors. PTEN gene deletion remains one of the few common genomic alterations in prostate cancer that is reproducibly associated with poor outcomes [1–13]. Because PTEN loss is commonly focal and subclonal in primary prostate tumors [4, 14–16], in situ methodologies for PTEN loss detection may be preferable to methods that assess copy number variation based on nucleic acid extraction, such as sequencing. Immunohistochemistry (IHC) and fluorescence in situ hybridization (FISH) have both been used to assess for PTEN loss in formalin fixed paraffin embedded (FFPE) tissues. Of the two, IHC-based detection of PTEN loss in prostate cancer is less expensive and less time-consuming for the routine screening of prostate tumor specimens, and may be easier to adapt to the current pathology work flow for risk assessment in prostate cancer. In cases of focal loss, detection of PTEN gene deletion by FISH can be especially challenging and more easily accomplished by IHC. Finally, in addition to genomic deletion, PTEN protein levels may be altered by small insertions/deletions or point mutations in the gene or even by microRNA- or epigenetic-regulated mechanisms which would not be detectable by FISH [4, 17–19]. However, only a few studies have directly compared PTEN IHC and PTEN FISH in large cohorts of prostate tumors with clinical outcome information [4, 17, 18, 20–24]
.We previously optimized and genetically validated a PTEN IHC assay for the detection of PTEN loss in prostate cancer specimens [4, 23, 24]. Using this assay, we showed that PTEN loss is associated with PTEN gene deletion [24], and independently associated with an increased risk of biochemical recurrence [7, 25] and lethal prostate cancer [13] in several large, multi-institutional cohorts of patients largely treated by radical prostatectomy. Similarly, PTEN loss by FISH has been reported to be associated with an increased risk of biochemical recurrence in a large cohort of patients treated by radical prostatectomy at the University Medical Center Hamburg-Eppendorf, Hamburg, Germany [5, 8]. Here, in the largest cohort ever studied by both techniques, we evaluated the performance of PTEN IHC in the Hamburg cohort and compared it to previously reported PTEN FISH results. We demonstrate that PTEN IHC and FISH results are largely concordant and associated with a similar increase in risk of biochemical recurrence in multivariable models. Finally, taking advantage of the relatively large cohort to examine the clinical outcomes of cases with discordant results by IHC and FISH, we find that these methods may provide complementary information in a subset of cases.
RESULTS
PTEN IHC was initially assessed in a total of 9033 tumors for the current study, of which 22% (n=2005) showed any PTEN loss (including 20% or n=1794 with homogeneous PTEN loss and 2% or n=211 with heterogeneous loss), 67% (n=6075) showed intact PTEN protein and 11% (953) showed ambiguous PTEN IHC (Figure 1). Of these cases, 52% (4732/9033) had evaluable PTEN FISH data available from previous studies, and FISH images from the tumors from the current study are available in these published manuscripts [5, 8]. Of these, 23% (1087/4732) showed any PTEN loss by IHC, including 20% (966/4732) with homogeneous PTEN loss and 3% (121/4732) with heterogeneous loss. An additional 70% (3330/4732) showed intact PTEN protein by IHC and 7% (315/4732) showed ambiguous PTEN IHC results. The remainder of the manuscript will focus on the subset of cases with evaluable IHC and FISH results (n=4417). The rates of PTEN gene and PTEN protein loss were quite similar in the subset with both FISH and IHC results compared to the entire evaluable cohort with interpretable results for each assay reported separately (Table 1). Among this subset of cases, PTEN IHC loss was found in 25% (1087/4417) of cases, including 22% with homogeneous PTEN loss and 3% with heterogeneous PTEN loss. The remaining 75% (3330/4417) had intact PTEN protein. Among these cases with IHC and FISH interpretable results, PTEN gene deletions were found in 21% (952/4417) of cases, including 12% (533/4417) with homozygous gene deletion and 9% (419/4417) with heterozygous gene deletion. The remaining 79% (3465/4417) had normal PTEN by FISH, similar to what was reported previously [5, 8].
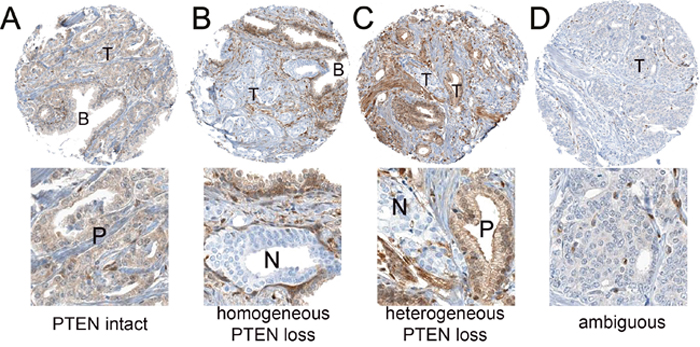
Figure 1: Representative PTEN immunohistochemistry results. (A) PTEN intact in tumor cells (T), with equivalent staining in nearby benign glands (B). Higher magnification inset below shows positive (P) staining tumor glands. (B) PTEN homogeneous loss in tumor glands (T), with intact staining in nearby benign glands (B) and stroma. Higher magnification inset below shows negative (N) staining tumor glands. (C) PTEN heterogeneous loss, with staining loss in some but not all sampled tumor cells (T). Higher magnification inset below shows positive (P) staining and negative (N) staining tumor glands. (D) PTEN ambiguous staining. PTEN is decreased but not lost in tumor glands and absence of background benign glands for comparison makes this case difficult to interpret. Higher magnification inset below shows glands with ambiguous PTEN immunostaining.
Table 1: Comparison of PTEN IHC and PTEN FISH results across all cases with available data
n (%) |
ambiguous IHC |
PTEN IHC intact |
PTEN IHC loss heterogeneous |
PTEN IHC loss homogeneous |
|---|---|---|---|---|
PTEN FISH normal |
280 (8%) |
3098 (83%) |
74 (2%) |
293 (8%) |
PTEN FISH heterozygous deletion |
18 (4%) |
146 (33%) |
23 (5%) |
250 (57%) |
PTEN FISH homozygous deletion |
17 (3%) |
86 (16%) |
24 (4%) |
423 (77%) |
Overall, there was a high concordance between PTEN IHC and FISH (p<0.0001). 93% (3098/3330) of tumors with intact PTEN IHC showed absence of PTEN gene deletion and 66% (720/1087) of cases with PTEN protein loss by IHC showed PTEN gene deletion by FISH. Similarly, 89% (3098/3465) of tumors with normal PTEN by FISH showed intact PTEN IHC and 76% (720/952) of cases with PTEN gene deletion by FISH showed PTEN protein loss by IHC. Overall, 84% (447/533) of cases with PTEN homozygous gene deletion had PTEN protein loss by IHC. 65% (273/419) of tumors with PTEN heterozygous gene deletion showed PTEN protein loss by IHC. Of the discordant cases with PTEN loss by IHC and normal PTEN FISH results, 20% showed heterogeneous PTEN loss. Notably, 20% (74/367) of the discordant cases (loss of PTEN protein expression by IHC and normal PTEN by FISH analysis) showed heterogeneous PTEN protein loss in some, but not all, sampled tumor glands, compared to only 11% (121/1087) of cases with PTEN IHC loss overall which showed heterogeneous PTEN loss. This suggests the possibility that tumor heterogeneity could explain at least some of the discordant results.
The negative predictive value for intact PTEN IHC was 93% (3098/3330) for lack of any gene deletion and 97% (3244/3330) for lack of homozygous PTEN deletion. The positive predictive value of PTEN IHC loss for presence of any PTEN gene deletion (homozygous or heterozygous) was 66% (720/1087) overall, or 70% (673/966) for homogeneous PTEN protein loss and 39% (47/121) for heterogeneous PTEN protein loss.
Associations between PTEN status and clinical-pathologic variables are shown in Table 2. PTEN loss by IHC was associated with a number of clinical-pathologic parameters (Table 2) and similar results are available for PTEN FISH in the same cohort in a previously published manuscript [5]. Increasing frequency of PTEN loss was seen in association with increasing pathologic stage (p<0.0001), increasing Gleason score (p<0.0001), presence of lymph node metastases (p<0.0001), higher pre-operative PSA levels (p<0.0001) and with a higher frequency of positive surgical margins (p<0.0001). Accordingly, patients with PTEN IHC loss had decreased PSA recurrence-free survival compared to patients with intact or ambiguous PTEN IHC status in univariable analyses (p<0.0001, Figure 2A). On multivariable analysis including pre-operative PSA level, pathologic tumor stage, Gleason score, lymph node status and margin status, PTEN loss by IHC remained significantly associated with decreased PSA recurrence-free survival, with a hazard ratio of 1.3 (95% CI:1.16-1.47), a hazard ratio roughly equivalent to that seen for positive lymph node status (Table 3).
Table 2: Associations between PTEN IHC status and clinical-pathologic variables
n evaluable |
% PTEN IHC intact |
% PTEN IHC loss |
p value |
|
|---|---|---|---|---|
All cancers |
7,813 |
75.7 |
24.2 |
|
Tumor stage |
||||
pT2 |
4,675 |
84.8 |
15.2 |
|
pT3a |
1,917 |
68.2 |
31.9 |
<0.0001 |
pT3b-pT4 |
1,181 |
52.5 |
47.4 |
|
Gleason grade |
||||
≤3+3 |
1,325 |
85.9 |
14 |
|
3+4 |
4,529 |
78.7 |
21.2 |
|
4+3 |
1,445 |
62.5 |
37.5 |
<0.0001 |
≥4+4 |
467 |
59.3 |
40.7 |
|
Gleason grade quantification |
||||
≤3+3 |
1,244 |
85.1 |
14.9 |
|
3+4 (≤5% pattern 4) |
952 |
86.5 |
13.4 |
|
3+4 (6-10% pattern 4) |
976 |
81.9 |
18 |
|
3+4 (11-20% pattern 4) |
814 |
74.7 |
25.3 |
|
3+4 (21-30% pattern 4) |
486 |
72.7 |
27.4 |
|
3+4 (31-49% pattern 4) |
379 |
69.4 |
30.6 |
|
3+4 (Tertiary pattern 5) |
259 |
75.3 |
24.7 |
<0.0001 |
4+3 (50-60% pattern 4) |
325 |
63.7 |
36.3 |
|
4+3 (61-80% pattern 4) |
305 |
62.9 |
37 |
|
4+3 (>80% pattern 4) |
71 |
67.6 |
32.4 |
|
4+3 (Tertiary pattern 5) |
425 |
61 |
39.1 |
|
≥4+4 |
314 |
57 |
43 |
|
Lymph node metastasis |
||||
N0 |
4,794 |
75.3 |
24.7 |
|
N+ |
592 |
51.3 |
48.6 |
<0.0001 |
Preop. PSA level (ng/ml) |
||||
<4 |
903 |
70.9 |
29.1 |
|
4-10 |
4,582 |
77.4 |
22.6 |
|
10-20 |
1,634 |
75.7 |
24.4 |
<0.0001 |
>20 |
603 |
72.5 |
27.5 |
|
Surgical margin |
||||
negative |
6,019 |
78 |
22 |
|
positive |
1,619 |
68 |
32 |
<0.0001 |
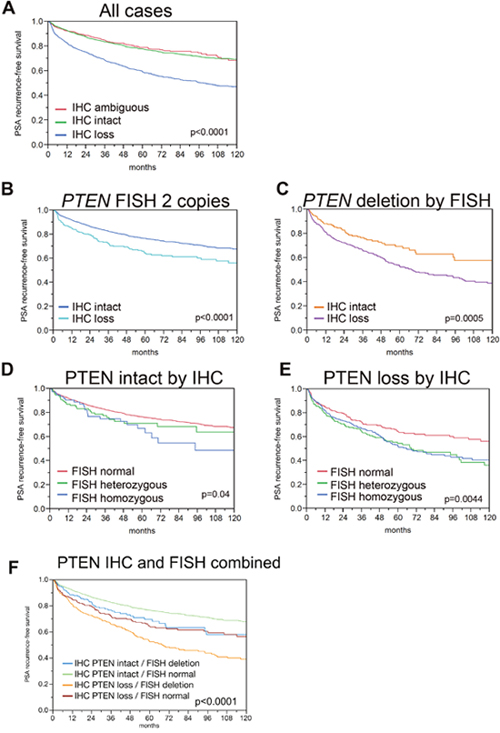
Figure 2: Kaplan-Meier analysis for PSA recurrence-free survival by PTEN IHC and FISH status. (A) All cases with PTEN IHC and FISH results, stratified by PTEN IHC intact (n=2,970, ncensored=2,372) and PTEN IHC loss (n=992, ncensored=615). (B) All cases with normal PTEN FISH results, stratified by PTEN IHC intact (n=2,756 ncensored=2214) and PTEN IHC loss (n=332, ncensored=229). (C) All cases with PTEN deletion by FISH stratified by PTEN IHC intact (n=214, ncensored=158) and PTEN IHC loss (n=660, ncensored=386). (D) All cases with PTEN intact by IHC, stratified by PTEN FISH normal (n=2,756, ncensored=2,214), PTEN FISH heterozygous deletion (n=137, ncensored=104) and PTEN FISH homozygous deletion (n=77, ncensored=54). For comparison between PTEN normal and PTEN heterozygous deletion, p=0.1016. For comparison between PTEN normal and PTEN homozygous deletion, p=0.0393. For comparison between PTEN homozygous deletion and PTEN heterozygous deletion, p=0.5776. (E) All cases with PTEN loss by IHC, stratified by PTEN FISH heterozygous deletion (n=248 ncensored=144) and PTEN FISH homozygous deletion (n=412, ncensored=242). For comparison between PTEN normal and PTEN heterozygous deletion, p=0.0017. For comparison between PTEN normal and PTEN homozygous deletion, p=0.0044. For comparison between PTEN homozygous deletion and PTEN heterozygous deletion, p=0.4777.
Table 3: Multivariable analysis of association of PTEN IHC status with clinical-pathologic variables
HR |
95% CI |
p-value |
||
|---|---|---|---|---|
4-10 vs <4 |
1.1 |
0.91-1.43 |
||
PSA level |
10-20 vs 4-10 |
1.3 |
1.12-1.45 |
<0.0001 |
>20 vs 10-20 |
1.3 |
1.12-1.51 |
||
pT3a vs pT2 |
1.9 |
1.66-2.25 |
||
pT stage |
pT3b vs pT3a |
1.5 |
1.28-1.69 |
<0.0001 |
pT4 vs pT3b |
1.3 |
0.91-1.76 |
||
3+4 vs ≤3+3 |
2.2 |
1.67-2.81 |
||
pGleason |
4+3 vs 3+4 |
2 |
1.8-2.32 |
<0.0001 |
≥4+4 vs 4+3 |
1.2 |
1-1.4 |
||
pN Stage |
N+ vs N0 |
1.4 |
1.21-1.65 |
<0.0001 |
margin status |
positive vs negative |
1.2 |
1.03-1.32 |
0.0164 |
PTEN |
loss vs intact |
1.3 |
1.16-1.47 |
<0.0001 |
We performed ROC analyses using PSA recurrence (biochemical recurrence, BCR) as the categorical response to estimate whether addition of PTEN IHC and/or FISH can improve the predictive power beyond established prognostic parameters including pT stage, Gleason grade, nodal stage and pre-surgical PSA value as the basic model. Addition of PTEN IHC increased the area under the ROC curve (AUC) from 0.701 to 0.741, and addition of PTEN FISH increased the AUC from 0.701 to 0.745. If both PTEN IHC and FISH were added to the basic model, there was only a slight further increase to 0.749.
Next, we examined the association of PTEN loss by IHC in combination with the PTEN FISH results with outcomes in univariable analysis. PTEN loss by IHC was associated with decreased PSA recurrence-free survival among tumors with PTEN gene deletion and among tumors with normal PTEN by FISH (p=0.0005 and p<0.0001 respectively) (Figure 2B, 2C). Conversely, among cases with intact PTEN by IHC, homozygous (p=0.04) but not heterozygous (p=0.10) PTEN gene deletion was weakly associated with decreased PSA recurrence-free survival compared to cases with PTEN FISH normal (Figure 2D). However among cases with PTEN loss by IHC, both homozygous (p=0.0044) and heterozygous (p=0.0017) PTEN gene deletion were associated with decreased PSA recurrence-free survival compared to cases with PTEN FISH normal (Figure 2E). The combined impact of the IHC and FISH findings on patient prognosis are shown in Figure 2F. The best prognosis was found for cancers with intact PTEN protein and normal PTEN copy numbers, while cancer with PTEN protein loss (IHC) and deletion (FISH) had the worst outcome (p<0.0001). An intermediate prognosis was found for tumors harboring only one alteration (i.e., PTEN protein loss or deletion). There was no difference between cancers with intact PTEN protein but deletion by FISH or loss of PTEN protein but normal copy numbers (p=0.4174), but both intermediate groups were significantly different from cancers with concordant lack or presence of PTEN alterations (p≤0.0125 each).
DISCUSSION
There is an increasing need for tissue-based prognostic biomarkers in prostate cancer with growing numbers of patients being enrolled on active surveillance protocols rather than undergoing definitive therapy. Loss of the PTEN tumor suppressor gene is arguably one of the most reproducible and best validated genetic biomarkers in prostate cancer. Indeed, its loss has been associated with increased risk of prostate cancer recurrence [1, 5, 7, 8, 12, 25] and death [9, 13, 26] in numerous independent studies. Because PTEN is most commonly inactivated in prostate cancer by large scale genomic deletion, PTEN FISH has historically been used to query PTEN status in tissue samples. However, interphase FISH is relatively expensive and time-consuming to perform, requiring the detailed enumeration of fluorescent signals in 20-100 nuclei under oil-immersion microscopy.
To address these challenges, several groups have developed immunohistochemistry (IHC) assays to query PTEN status in archived formalin fixed tissue samples [4, 27, 28]. While a number of such assays have been published, for the most part, these assays have largely been compared to PTEN FISH in only small scale studies with less than 200 samples [4, 17, 18, 20–24]. In the few larger studies that compared IHC and FISH, including one previous study on the cohort described in the current study, there was no significant [5] or only weak [9] correlation between the assays, likely due to failure of the IHC assay to validate.
To improve on existing assays, we used a commercially available rabbit monoclonal antibody against PTEN coupled with an automated staining system to develop and validate a clinical-grade PTEN IHC test. Our simple scoring system for this assay has shown high inter-observer reproducibility in a number of cohorts, with κ values exceeding 0.9 [13, 25]. Using this assay, our group previously demonstrated that PTEN protein loss is associated with an increased risk of recurrence and progression in surgically treated cohorts of prostate cancer patients [6, 7, 25]. More importantly, PTEN loss using our assay is independently associated with increased risk of lethal prostate cancer in an independent, large population-based cohort [13]. More recently, we have examined the correlation between a 4-color FISH assay and PTEN IHC results in the Canary Retrospective Tissue Microarray cohort and found excellent concordance [24]. However this previous study included only a small number of cases with discordant status by FISH and IHC which precluded any meaningful analysis of outcomes in this group.
In the current study, we compared PTEN IHC and FISH results across the largest available radical prostatectomy cohort with clinical follow-up. Successful analysis of more than 9000 tumors with PTEN IHC and more than 6700 tumors with PTEN FISH resulted in more than 4400 tumors with concurrent PTEN FISH and IHC results for comparison. Overall, there was an excellent concordance between the two techniques. Reasons for discordance between PTEN IHC and FISH likely include tumor heterogeneity [14, 16, 29]. Because IHC and FISH analysis were performed several years apart, the TMA sections used for each were not adjacent to one another and may, thus, represent different subclones of the same tumor in a fraction of cases, a weakness of the current study design that was unavoidable for logistical reasons. Intratumoral heterogeneity in PTEN loss can be seen in upwards of 50% of primary prostate tumors [13, 14, 16] and might be one important explanation for the 15% of cases with homozygous PTEN deletion by FISH that had intact PTEN by IHC. Among cases with normal or hemizygous PTEN by FISH yet PTEN loss by IHC, another explanation could be the occurrence of alterations that are undetectable by our FISH assay yet lead to protein loss, such as truncating mutations in PTEN (nonsense, frameshift and splice site mutations), structural rearrangements, epigenetic alterations or modifications influencing protein stability and half-life [16, 30–33]. Finally, it cannot be excluded that a fraction of samples were misclassified due to technical reasons. FISH scoring cut-offs are determined by studying a relatively small number of cases with known PTEN genomic status, however because of the statistical nature of these cut-off determinations, a small fraction of cases may be misclassified by FISH [34]. Likewise, IHC can only detect protein concentrations beyond the detection limit of IHC, and the amount of detectable protein may be influenced by many non-biological factors such as tissue quality, protein preservation, antibody concentration and antigen retrieval efficacy [35, 36].
Though we have conducted smaller comparisons of our IHC assay with FISH previously [24], the large number of cases in the current study enabled us to examine the clinical significance of discordance between PTEN IHC and FISH. Given that many factors other than the gene copy number can influence protein levels, it seems unlikely that discrepant IHC/FISH findings are merely due to failure of one of the two tests. The fact that the presence of one alteration (either on the DNA or on the protein level) is associated with a significant worsening of the patient’s prognosis, and that the prognostic impact of isolated protein or DNA alterations is virtually identical but less severe compared to when both types of alterations co-occur, may fit with the proposed dose-dependency of PTEN’s tumor suppressor function [37]. Our data thus suggest that PTEN protein loss and gene deletion represent complementary mechanisms of PTEN inactivation and each provides complementary prognostic information. In fact, multivariate ROC-AUC modelling including PTEN IHC and PTEN FISH in addition to established prognostic parameters such as pT stage, Gleason score, nodal stage and pre-surgical PSA indicates that both IHC and FISH similarly improve predictive accuracy. That PTEN loss by IHC identified cancers with markedly reduced PSA recurrence-free survival independently of the FISH status is consistent with the idea that a “direct” analysis of the protein – i.e., the “active” component of a gene - may be superior to “indirect” analysis of its mere copy number state. However, that FISH analysis also predicted prognosis in the subset of cancers with PTEN loss by IHC, demonstrates that copy number analysis may hold relevant prognostic potential beyond the protein level. A possible explanation is that genomic deletion can also be a surrogate marker for genetic instability, which is generally linked to poor outcome [38]. Profiling studies using next generation sequencing have shown that prostate cancers often harbor multiple additional structural genomic alterations [31, 32, 38, 39], and we have demonstrated earlier in the same cohort as used in the present study that many of these deletions are linked to poor outcome [5, 40–44]. Finally, we also cannot exclude the possibility that since the FISH and IHC assays were not conducted on adjacent sections, performing both assays was a mechanism to evaluate the tumor for PTEN loss in two separate areas, identifying more cases with heterogeneous PTEN loss than either single test alone.
Irrespective of the reasons leading to discrepant IHC and FISH findings, the strong and independent prognostic impact of both methods suggests that re-testing with a second method could be justified in a subset of cases, particularly in cases with PTEN deletion or lack of deletion by FISH or PTEN protein loss by IHC. Although a negative result with one test had a high negative predictive value for the other method (IHC: 93%, FISH: 89%), these figures do also demonstrate that about 10% of all cancers (accounting for 45% of PTEN-deficient cases) harbor PTEN alterations that remain undetected if only one method is employed. Our data do not argue for a particular sequence of PTEN testing. Arguments in favor of first-line FISH are that FISH eliminates the need to compare staining intensities between cancer and normal cells and that it gives a clear-cut “yes/no” answer with regards to the genomic status of the tumor. Arguments for using IHC as a first test include the lower costs and shorter analysis time, the higher rate of technical failure with FISH, as well as the fact that the IHC test is more easily integrated into standard pathology laboratory work flow. IHC analysis could, therefore, be the method of choice in places where no FISH analysis is possible.
There are a number of limitations of the current study. Technical difficulties associated with optimizing FISH hybridization on TMA slides meant that more than half of the cases with PTEN IHC results did not have accompanying FISH results. Though we showed that the samples with data by both methodologies were essentially comparable to the larger set with IHC results, we cannot exclude some bias in the cases that were excluded for unavailable FISH results and thus not studied by both methods. In addition, the design of the TMA set in the current study sampled only one core of tumor tissue for each case. Tumor heterogeneity is a major factor in all TMA and prostate biopsy studies, and the one-core-per-cancer sampling strategy in our TMA is not suitable to address inter- or even intratumoral heterogeneity. The very low likelihood to have relevant heterogeneity among the about 500 tumor cells that are typically present in a 0.6 mm TMA spot is perfectly reflected by the markedly lower rate of heterogeneous PTEN IHC loss (2%) in the current study as compared to other TMA cohorts with 3-4 tumor cores sampled from each case [13, 25]. However, it is noteworthy, that the amount of tissue studied in a minute TMA spot closely resembles that of a typical core needle biopsy, making our one-spot-per-cancer approach a suitable surrogate for molecular analyses on diagnostic biopsies. Finally, the “true” PTEN status is unknown in cases with discordant FISH and IHC results. Though FISH can detect deletions resulting from genomic rearrangement that are the most common mechanism of loss in prostate cancer, it will miss very small structural variations, indels and missense mutations that have been found by next generation sequencing in up to 5% of cancers. In addition, IHC does not provide information about protein activity, and there is no established threshold to distinguish between “sufficient” and “insufficient” levels of the protein with respect to downstream oncogenic signaling.
In conclusion, in the largest radical prostatectomy cohort studied to date with clinical outcome information, PTEN loss by our clinical-grade IHC assay is highly concordant with PTEN gene status by FISH, and associated with poor outcomes in the disease. That a relevant fraction of about 10% cancers yield discrepant results between FISH and IHC analysis, which are similarly linked to tumor aggressiveness, suggests that clinically relevant PTEN alterations can be missed if only one method is applied. Our findings thus argue for a combination of both methods in order to obtain the most accurate information on PTEN status with current state-of–the-art diagnostic in situ methods.
MATERIALS AND METHODS
Subject selection and tissue microarray design
The features of this cohort have been previously described in detail elsewhere [5]. Briefly, the cohort consists of radical prostatectomy specimens from 13,665 consecutive patients undergoing radical prostatectomy between 1992 and 2008 at the Department of Urology, University Medical Center Hamburg-Eppendorf, Hamburg, Germany. PTEN immunohistochemistry (IHC) was assessed in a total of 9033 tumors for the current study and 4732 of these patients had PTEN fluorescence in situ hybridization (FISH) data available for comparison. Clinical follow-up data were available for 4203 cases. The median follow-up was 46.7 months (range, 1 to 219 months). None of the patients received neoadjuvant endocrine therapy. In all patients, PSA values were measured quarterly in the first year, followed by biannual measurements in the second year and annual measurements after the third year following surgery. Recurrence was defined as a postoperative PSA of 0.2 ng/mL, increasing thereafter. The first PSA value of 0.2 ng/mL or greater was used to define the time of recurrence. Salvage therapy was initiated in cases of biochemical relapse. Patients without evidence of tumor recurrence were censored at the last follow-up. All cancers were arrayed on 30 tissue microarrays blocks, where each tumor was sampled once, utilizing 0.6 mm cores. The area selected for sampling was guided not by Gleason grade, but to maximize tumor content available for analysis in the TMA core.
Immunohistochemistry assays
PTEN immunohistochemistry (IHC) was performed as recently reported [13, 25]. Briefly, the protocol uses the Ventana automated staining platform (Ventana Discovery Ultra, Ventana Medical Systems, Tucson, AZ) and a rabbit anti-human PTEN antibody (Clone D4.3 XP; Cell Signaling Technologies, Danvers, MA).
Immunohistochemistry scoring
After staining for PTEN, all TMAs were scanned at 20x magnification (Aperio, Leica Microsystems, Buffalo Grove, IL) and segmented into TMAJ for scoring (http://tmaj.pathology.jhmi.edu/). PTEN protein status was blindly and independently scored by a trained pathologist (TLL) using a previously validated scoring system (Figure 1). A tissue core was considered to have PTEN protein loss if the intensity of cytoplasmic and nuclear staining was markedly decreased or entirely negative across >10% of tumor cells compared to surrounding benign glands and/or stroma, which provide internal positive controls for PTEN protein expression [4, 13]. If the tumor core showed PTEN protein expressed in >90% of sampled tumor glands, the tumor was scored as PTEN intact. If PTEN was lost in <100% of the tumor cells sampled in a given core, the core was annotated as showing heterogeneous PTEN loss in some, but not all, cancer glands (focal loss). Alternatively, if the core showed PTEN loss in 100% of sampled tumor glands, the core was annotated as showing homogeneous PTEN loss. Finally, a small percentage of cores were scored as having ambiguous PTEN IHC results. This occurred when the intensity of the tumor cell staining was light or absent in the absence of evaluable internal benign glands or stromal staining.
Initial blinded analysis of PTEN FISH
PTEN FISH was performed as previously described [5, 8]. Briefly, a dual-color FISH probe set was used consisting of two SpectrumGreen-labeled bacterial artificial chromosome clones (RP11-380G5 and RP11-813O3; Source Bioscience, Nottingham, UK) and a SpectrumOrange-labeled commercial centromere 10 probe (06J36-090; Abbott, Wiesbaden, Germany) as a reference. The predominant red and green signal numbers were recorded for each FISH probe. A total of 659 tissue spots were excluded from FISH analysis because basal cell marker 34βE12 analysis indicated lack of tumor cells. Thresholds for PTEN FISH analysis were established from 0.6-mm tissue spots from seven tumors with a known PTEN deletion (four with a heterozygous and three with a homozygous deletion), based on single-nucleotide polymorphism (SNP) array copy number analysis. In five of these tumors, PTEN signal losses by FISH were found in all analyzed tissue blocks. The two remaining cancers had tissue blocks with and without PTEN deletion, indicating the presence of intratumoral heterogeneity. In all seven cases, tumor blocks with PTEN deletion had FISH signal losses in most (at least 60%) tumor cells. According to these findings, homozygous deletion of PTEN was defined as complete absence of PTEN FISH probe signals in ≥60% of tumor nuclei of the tissue spot, with the presence of one or two PTEN FISH signals in adjacent normal cells. Tissue spots with a lack of PTEN signals in all (tumor and normal cells) or lack of any normal cells as an internal control for successful hybridization of the PTEN probe were excluded from analysis. Heterozygous deletion of PTEN was defined as the presence of fewer PTEN signals than centromere 10 probe signals in ≥60% tumor nuclei.
Statistics
Statistical calculations were performed with JMP® 10.0.2 software (SAS Institute Inc., NC, USA). Contingency tables and the chi2-test were performed to search for associations between PTEN alterations and tumor phenotype. Survival curves were calculated according to Kaplan-Meier. The Log-Rank test was applied to detect significant differences between groups. Cox proportional hazards regression analysis was performed to test the statistical independence and significance between pathological, molecular and clinical variables. Logistic regression was used to quantify the area under the receiver-operator curve (ROC-AUC).
CONFLICTS OF INTEREST
TLL has received research funding from Ventana Medical Systems.
FUNDING
Funding for this research was provided in part by the the CDMRP Transformative Impact Award (W81XWH-12-PCRP-TIA, TLL), a CDMRP Idea Award (W81XWH-13-1-0271, TLL).
REFERENCES
1. Yoshimoto M, Cunha IW, Coudry RA, Fonseca FP, Torres CH, Soares FA, Squire JA. FISH analysis of 107 prostate cancers shows that PTEN genomic deletion is associated with poor clinical outcome. Br J Cancer. 2007; 97:678–85. https://doi.org/10.1038/sj.bjc.6603924.
2. McCall P, Witton CJ, Grimsley S, Nielsen KV, Edwards J. Is PTEN loss associated with clinical outcome measures in human prostate cancer? Br J Cancer. 2008; 99:1296–301. https://doi.org/10.1038/sj.bjc.6604680.
3. Yoshimoto M, Joshua AM, Cunha IW, Coudry RA, Fonseca FP, Ludkovski O, Zielenska M, Soares FA, Squire JA. Absence of TMPRSS2:ERG fusions and PTEN losses in prostate cancer is associated with a favorable outcome. Mod Pathol. 2008; 21:1451–60. https://doi.org/10.1038/modpathol.2008.96.
4. Lotan TL, Gurel B, Sutcliffe S, Esopi D, Liu W, Xu J, Hicks JL, Park BH, Humphreys E, Partin AW, Han M, Netto GJ, Isaacs WB, De Marzo AM. PTEN protein loss by immunostaining: analytic validation and prognostic indicator for a high risk surgical cohort of prostate cancer patients. Clin Cancer Res. 2011; 17:6563–73. https://doi.org/10.1158/1078-0432.CCR-11-1244.
5. Krohn A, Diedler T, Burkhardt L, Mayer PS, De Silva C, Meyer-Kornblum M, Kötschau D, Tennstedt P, Huang J, Gerhäuser C, Mader M, Kurtz S, Sirma H, et al. Genomic deletion of PTEN is associated with tumor progression and early PSA recurrence in ERG fusion-positive and fusion-negative prostate cancer. Am J Pathol. 2012; 181:401–12. https://doi.org/10.1016/j.ajpath.2012.04.026.
6. Antonarakis ES, Keizman D, Zhang Z, Gurel B, Lotan TL, Hicks JL, Fedor HL, Carducci MA, De Marzo AM, Eisenberger MA. An immunohistochemical signature comprising PTEN, MYC, and Ki67 predicts progression in prostate cancer patients receiving adjuvant docetaxel after prostatectomy. Cancer. 2012; 118:6063–71. https://doi.org/10.1002/cncr.27689.
7. Chaux A, Peskoe SB, Gonzalez-Roibon N, Schultz L, Albadine R, Hicks J, De Marzo AM, Platz EA, Netto GJ. Loss of PTEN expression is associated with increased risk of recurrence after prostatectomy for clinically localized prostate cancer. Mod Pathol. 2012; 25:1543–49. https://doi.org/10.1038/modpathol.2012.104.
8. Steurer S, Mayer PS, Adam M, Krohn A, Koop C, Ospina-Klinck D, Tehrani AA, Simon R, Tennstedt P, Graefen M, Wittmer C, Brors B, Plass C, et al. TMPRSS2-ERG fusions are strongly linked to young patient age in low-grade prostate cancer. Eur Urol. 2014; 66:978–81. https://doi.org/10.1016/j.eururo.2014.06.027.
9. Cuzick J, Yang ZH, Fisher G, Tikishvili E, Stone S, Lanchbury JS, Camacho N, Merson S, Brewer D, Cooper CS, Clark J, Berney DM, Møller H, et al, and Transatlantic Prostate Group. Prognostic value of PTEN loss in men with conservatively managed localised prostate cancer. Br J Cancer. 2013; 108:2582–89. https://doi.org/10.1038/bjc.2013.248.
10. Liu W, Xie CC, Thomas CY, Kim ST, Lindberg J, Egevad L, Wang Z, Zhang Z, Sun J, Sun J, Koty PP, Kader AK, Cramer SD, et al. Genetic markers associated with early cancer-specific mortality following prostatectomy. Cancer. 2013; 119:2405–12. https://doi.org/10.1002/cncr.27954.
11. Mithal P, Allott E, Gerber L, Reid J, Welbourn W, Tikishvili E, Park J, Younus A, Sangale Z, Lanchbury JS, Stone S, Freedland SJ. PTEN loss in biopsy tissue predicts poor clinical outcomes in prostate cancer. Int J Urol. 2014; 21:1209–14. https://doi.org/10.1111/iju.12571.
12. Troyer DA, Jamaspishvili T, Wei W, Feng Z, Good J, Hawley S, Fazli L, McKenney JK, Simko J, Hurtado-Coll A, Carroll PR, Gleave M, Lance R, et al. A multicenter study shows PTEN deletion is strongly associated with seminal vesicle involvement and extracapsular extension in localized prostate cancer. Prostate. 2015; 75:1206–15. https://doi.org/10.1002/pros.23003.
13. Ahearn TU, Pettersson A, Ebot EM, Gerke T, Graff RE, Morais CL, Hicks JL, Wilson KM, Rider JR, Sesso HD, Fiorentino M, Flavin R, Finn S, et al. A Prospective Investigation of PTEN Loss and ERG Expression in Lethal Prostate Cancer. J Natl Cancer Inst. 2015; 108:djv346. https://doi.org/10.1093/jnci/djv346.
14. Gumuskaya B, Gurel B, Fedor H, Tan HL, Weier CA, Hicks JL, Haffner MC, Lotan TL, De Marzo AM. Assessing the order of critical alterations in prostate cancer development and progression by IHC: further evidence that PTEN loss occurs subsequent to ERG gene fusion. Prostate Cancer Prostatic Dis. 2013; 16:209–15.
15. Bismar TA, Yoshimoto M, Duan Q, Liu S, Sircar K, Squire JA. Interactions and relationships of PTEN, ERG, SPINK1 and AR in castration-resistant prostate cancer. Histopathology. 2012; 60:645–52. https://doi.org/10.1111/j.1365-2559.2011.04116.x.
16. Krohn A, Freudenthaler F, Harasimowicz S, Kluth M, Fuchs S, Burkhardt L, Stahl P, C Tsourlakis M, Bauer M, Tennstedt P, Graefen M, Steurer S, Sirma H, et al. Heterogeneity and chronology of PTEN deletion and ERG fusion in prostate cancer. Mod Pathol. 2014; 27:1612–20. https://doi.org/10.1038/modpathol.2014.70.
17. Han B, Mehra R, Lonigro RJ, Wang L, Suleman K, Menon A, Palanisamy N, Tomlins SA, Chinnaiyan AM, Shah RB. Fluorescence in situ hybridization study shows association of PTEN deletion with ERG rearrangement during prostate cancer progression. Mod Pathol. 2009; 22:1083–93. https://doi.org/10.1038/modpathol.2009.69.
18. Verhagen PC, van Duijn PW, Hermans KG, Looijenga LH, van Gurp RJ, Stoop H, van der Kwast TH, Trapman J. The PTEN gene in locally progressive prostate cancer is preferentially inactivated by bi-allelic gene deletion. J Pathol. 2006; 208:699–707. https://doi.org/10.1002/path.1929.
19. Poliseno L, Salmena L, Riccardi L, Fornari A, Song MS, Hobbs RM, Sportoletti P, Varmeh S, Egia A, Fedele G, Rameh L, Loda M, Pandolfi PP. Identification of the miR-106b~25 microRNA cluster as a proto-oncogenic PTEN-targeting intron that cooperates with its host gene MCM7 in transformation. Sci Signal. 2010; 3:ra29. https://doi.org/10.1126/scisignal.2000594.
20. Yoshimoto M, Cutz JC, Nuin PA, Joshua AM, Bayani J, Evans AJ, Zielenska M, Squire JA. Interphase FISH analysis of PTEN in histologic sections shows genomic deletions in 68% of primary prostate cancer and 23% of high-grade prostatic intra-epithelial neoplasias. Cancer Genet Cytogenet. 2006; 169:128–37. https://doi.org/10.1016/j.cancergencyto.2006.04.003.
21. Ferraldeschi R, Nava Rodrigues D, Riisnaes R, Miranda S, Figueiredo I, Rescigno P, Ravi P, Pezaro C, Omlin A, Lorente D, Zafeiriou Z, Mateo J, Altavilla A, et al. PTEN protein loss and clinical outcome from castration-resistant prostate cancer treated with abiraterone acetate. Eur Urol. 2015; 67:795–802. https://doi.org/10.1016/j.eururo.2014.10.027.
22. Lotan TL, Carvalho FL, Peskoe SB, Hicks JL, Good J, Fedor H, Humphreys E, Han M, Platz EA, Squire JA, De Marzo AM, Berman DM. PTEN loss is associated with upgrading of prostate cancer from biopsy to radical prostatectomy. Mod Pathol. 2015; 28:128–37. https://doi.org/10.1038/modpathol.2014.85.
23. Picanço-Albuquerque CG, Morais CL, Carvalho FL, Peskoe SB, Hicks JL, Ludkovski O, Vidotto T, Fedor H, Humphreys E, Han M, Platz EA, De Marzo AM, Berman DM, et al. In prostate cancer needle biopsies, detections of PTEN loss by fluorescence in situ hybridization (FISH) and by immunohistochemistry (IHC) are concordant and show consistent association with upgrading. Virchows Arch. 2016; 468:607–17. https://doi.org/10.1007/s00428-016-1904-2.
24. Lotan TL, Wei W, Ludkovski O, Morais CL, Guedes LB, Jamaspishvili T, Lopez K, Hawley ST, Feng Z, Fazli L, Hurtado-Coll A, McKenney JK, Simko J, et al. Analytic validation of a clinical-grade PTEN immunohistochemistry assay in prostate cancer by comparison with PTEN FISH. Mod Pathol. 2016; 29:904–14. https://doi.org/10.1038/modpathol.2016.88.
25. Lotan TL, Wei W, Morais CL, Hawley ST, Fazli L, Hurtado-Coll A, Troyer D, McKenney JK, Simko J, Carroll PR, Gleave M, Lance R, Lin DW, et al. PTEN Loss as Determined by Clinical-grade Immunohistochemistry Assay Is Associated with Worse Recurrence-free Survival in Prostate Cancer. Eur Urol Focus. 2016; 2:180–88. https://doi.org/10.1016/j.euf.2015.07.005.
26. Reid AH, Attard G, Ambroisine L, Fisher G, Kovacs G, Brewer D, Clark J, Flohr P, Edwards S, Berney DM, Foster CS, Fletcher A, Gerald WL, et al, and Transatlantic Prostate Group. Molecular characterisation of ERG, ETV1 and PTEN gene loci identifies patients at low and high risk of death from prostate cancer. Br J Cancer. 2010; 102:678–84. https://doi.org/10.1038/sj.bjc.6605554.
27. Sangale Z, Prass C, Carlson A, Tikishvili E, Degrado J, Lanchbury J, Stone S. A robust immunohistochemical assay for detecting PTEN expression in human tumors. Appl Immunohistochem Mol Morphol. 2011; 19:173–83. https://doi.org/10.1097/PAI.0b013e3181f1da13.
28. Ugalde-Olano A, Egia A, Fernández-Ruiz S, Loizaga-Iriarte A, Zuñiga-García P, Garcia S, Royo F, Lacasa-Viscasillas I, Castro E, Cortazar AR, Zabala-Letona A, Martín-Martín N, Arruabarrena-Aristorena A, et al. Methodological aspects of the molecular and histological study of prostate cancer: focus on PTEN. Methods. 2015; 77-78:25–30. https://doi.org/10.1016/j.ymeth.2015.02.005.
29. Bismar TA, Yoshimoto M, Vollmer RT, Duan Q, Firszt M, Corcos J, Squire JA. PTEN genomic deletion is an early event associated with ERG gene rearrangements in prostate cancer. BJU Int. 2011; 107:477–85. https://doi.org/10.1111/j.1464-410X.2010.09470.x.
30. Taylor BS, Schultz N, Hieronymus H, Gopalan A, Xiao Y, Carver BS, Arora VK, Kaushik P, Cerami E, Reva B, Antipin Y, Mitsiades N, Landers T, et al. Integrative genomic profiling of human prostate cancer. Cancer Cell. 2010; 18:11–22. https://doi.org/10.1016/j.ccr.2010.05.026.
31. Berger MF, Lawrence MS, Demichelis F, Drier Y, Cibulskis K, Sivachenko AY, Sboner A, Esgueva R, Pflueger D, Sougnez C, Onofrio R, Carter SL, Park K, et al. The genomic complexity of primary human prostate cancer. Nature. 2011; 470:214–20. https://doi.org/10.1038/nature09744.
32. Grasso CS, Wu YM, Robinson DR, Cao X, Dhanasekaran SM, Khan AP, Quist MJ, Jing X, Lonigro RJ, Brenner JC, Asangani IA, Ateeq B, Chun SY, et al. The mutational landscape of lethal castration-resistant prostate cancer. Nature. 2012; 487:239–43. https://doi.org/10.1038/nature11125.
33. Robinson D, Van Allen EM, Wu YM, Schultz N, Lonigro RJ, Mosquera JM, Montgomery B, Taplin ME, Pritchard CC, Attard G, Beltran H, Abida W, Bradley RK, et al. Integrative clinical genomics of advanced prostate cancer. Cell. 2015; 161:1215–28. https://doi.org/10.1016/j.cell.2015.05.001.
34. Yoshimoto M, Ludkovski O, Good J, Gooding RJ, McGowan-Jordan J, Boag A, Evans A, Tsao MS, Nuin P, Squire JA. Abstract 63: incorporation of flanking probes reduces truncation losses for fluorescence in situ hybridization analysis of recurrent genomic deletions in tumor sections. Cancer Res. 2013; 73:63. https://doi.org/10.1158/1538-7445.AM2013-63.
35. Hicks DG, Boyce BF. The challenge and importance of standardizing pre-analytical variables in surgical pathology specimens for clinical care and translational research. Biotech Histochem. 2012; 87:14–17. https://doi.org/10.3109/10520295.2011.591832.
36. Tennstedt P, Sauter G. Quality aspects of TMA analysis. Methods Mol Biol. 2010; 664:17–26. https://doi.org/10.1007/978-1-60761-806-5_2.
37. Carracedo A, Alimonti A, Pandolfi PP. PTEN level in tumor suppression: how much is too little? Cancer Res. 2011; 71:629–33. https://doi.org/10.1158/0008-5472.CAN-10-2488.
38. Hieronymus H, Schultz N, Gopalan A, Carver BS, Chang MT, Xiao Y, Heguy A, Huberman K, Bernstein M, Assel M, Murali R, Vickers A, Scardino PT, et al. Copy number alteration burden predicts prostate cancer relapse. Proc Natl Acad Sci USA. 2014; 111:11139–44. https://doi.org/10.1073/pnas.1411446111.
39. Barbieri CE, Baca SC, Lawrence MS, Demichelis F, Blattner M, Theurillat JP, White TA, Stojanov P, Van Allen E, Stransky N, Nickerson E, Chae SS, Boysen G, et al. Exome sequencing identifies recurrent SPOP, FOXA1 and MED12 mutations in prostate cancer. Nat Genet. 2012; 44:685–89. https://doi.org/10.1038/ng.2279.
40. Kluth M, Runte F, Barow P, Omari J, Abdelaziz ZM, Paustian L, Steurer S, Christina Tsourlakis M, Fisch M, Graefen M, Tennstedt P, Huland H, Michl U, et al. Concurrent deletion of 16q23 and PTEN is an independent prognostic feature in prostate cancer. Int J Cancer. 2015; 137:2354–63. https://doi.org/10.1002/ijc.29613.
41. Kluth M, Hesse J, Heinl A, Krohn A, Steurer S, Sirma H, Simon R, Mayer PS, Schumacher U, Grupp K, Izbicki JR, Pantel K, Dikomey E, et al. Genomic deletion of MAP3K7 at 6q12-22 is associated with early PSA recurrence in prostate cancer and absence of TMPRSS2:ERG fusions. Mod Pathol. 2013; 26:975–83. https://doi.org/10.1038/modpathol.2012.236.
42. Kluth M, Graunke M, Möller-Koop C, Hube-Magg C, Minner S, Michl U, Graefen M, Huland H, Pompe R, Jacobsen F, Hinsch A, Wittmer C, Lebok P, et al. Deletion of 18q is a strong and independent prognostic feature in prostate cancer. Oncotarget. 2016; 7:86339–49. https://doi.org/10.18632/oncotarget.13404.
43. Kluth M, Amschler NN, Galal R, Möller-Koop C, Barrow P, Tsourlakis MC, Jacobsen F, Hinsch A, Wittmer C, Steurer S, Krech T, Büscheck F, Clauditz TS, et al. Deletion of 8p is an independent prognostic parameter in prostate cancer. Oncotarget. 2017; 8:379–92. https://doi.org/10.18632/oncotarget.13425.
44. Kluth M, Ahrary R, Hube-Magg C, Ahmed M, Volta H, Schwemin C, Steurer S, Wittmer C, Wilczak W, Burandt E, Krech T, Adam M, Michl U, et al. Genomic deletion of chromosome 12p is an independent prognostic marker in prostate cancer. Oncotarget. 2015; 6:27966–79. https://doi.org/10.18632/oncotarget.4626.