
INTRODUCTION
Selenium, an essential non-metal trace element [1], is crucial to the function of several enzymes, including glutathioneperoxidases and thioredoxin reductases [2]. Selenium deficiency is related to tumorigenesis, and selenium recruitment reduces the risk of cancer in humans [3, 4]. Furthermore, selenium compounds can prevent tumor formation [5], mainly by inducing apoptosis [6]. For example, Yang and Wang found that Na5SeV5O18.H20, a heteropoly compound containing selenium, had anti-tumor effects both in vitro and in vivo [7]. When used as therapeutic agents in tumor patients, selenium compounds are well-tolerated [8].
The possible anti-tumor effects and related underlying mechanisms for another selenium-containing heteropoly compound, (NH4)2H15Se2VIMo10V3O52·2H2O (abbreviation: Se2Mo10V3), have not yet been studied. Here, we examined the anti-tumor effects of Se2Mo10V3 and the mechanisms underlying those effects.
RESULTS
Se2Mo10V3 inhibited proliferation in K562 cells
As shown in Figure 1, Se2Mo10V3 (12.5-100 mg/L) inhibited proliferation in K562 cells. The inhibitory rates for the 12.5, 25, 50, and 100 mg/L doses were 15.91±2.94, 16.64±0.97, 17.93±3.92, and 34.65±0.73% after 24 h (all P<0.05), 17.02±3.77, 20.78±0.15, 35.79±1.83, and 59.51±1.16% after 48 h (all P<0.05), and 30.12±3.76, 48.07±2.16, 62.93±1.63, and 92.77±1.51% after 72 h (all P<0.01), respectively, with IC50 values of >100 mg/L after 24 h, 78.72±2.82 mg/L after 48 h, and 24.94±0.88 mg/L after 72 h. The inhibitory rate in control cells was set to 0. In positive control K562 cells treated with carboplatin (12.5-100 mg/L), IC50 values were >100 mg/L after 24 h, 52.39 mg/L after 48 h, and 25.00 mg/L after 72 h. Similar inhibitory effects were observed in A549, CNE, SPCA-1, and Lewis cells after Se2Mo10V3 treatment (data not shown).
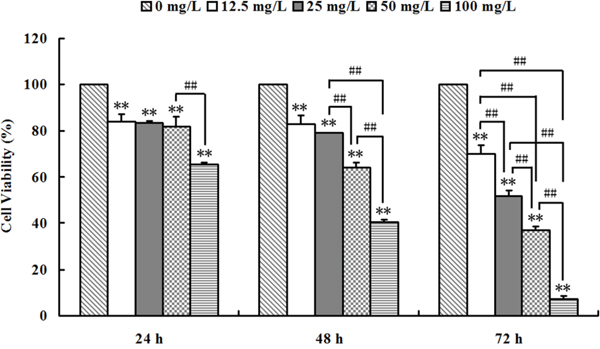
Figure 1: Se2Mo10V3 inhibited proliferation in K562 cells in an MTT assay. Data are expressed as means ± S.D. of three independent experiments. *P<0.05, **P<0.01 versus control cells.
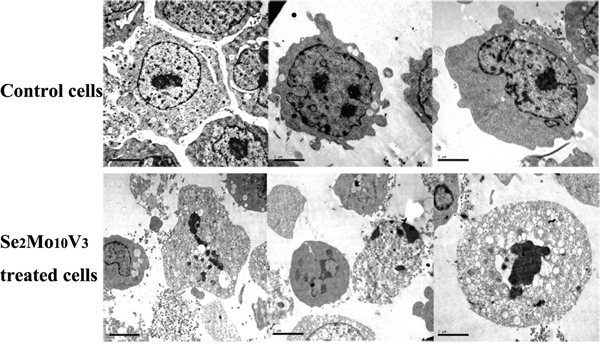
Figure 2: 50 mg/L Se2Mo10V3 induced apoptosis in K562 cells in a TEM expreiment. Scale = 2.00 μm. Data are representative of three independent experiments.
Se2Mo10V3 induced apoptotic morphology in K562 cells
As shown in Figure 3, typical indicators of apoptosis, such as chromatin fragmentation, condensation and margination, cytoplasm vacuoles, and disappearance of cytolemma microvilli, were observed to a greater extent in K562 cells treated with 50 mg/L Se2Mo10V3 than in control cells, which were characterized by normal chromatin, uniform cytoplasm, the presence of cytolemma microvilli, and diastolic cell bodies.
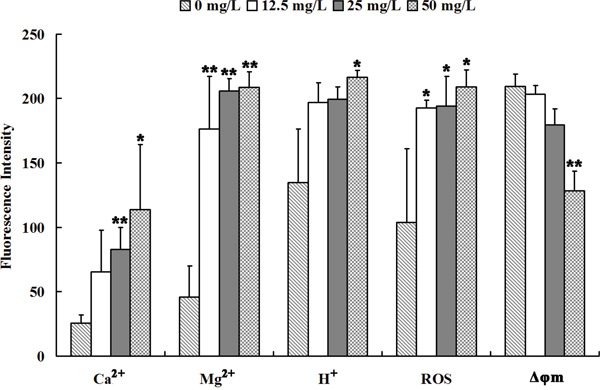
Figure 3: Se2Mo10V3 increased Ca2+, Mg2+, H+, and ROS levels and decreased Δφm in K562 cells in a CLSM experiment. Data are expressed as means ± S.D. of three independent experiments. *P<0.05, **P<0.01 versus control cells.
Se2Mo10V3 increased Ca2+, Mg2+, H+, and ROS levels and decreased Δφm in K562 cells
The results of CLSM are shown in Figure 4. Intracellular Ca2+ levels increased after treatment with 25 or 50 mg/L Se2Mo10V3 (both P<0.05). Intracellular Mg2+ levels also increased after treatment with 12.5, 25, or 50 mg/L Se2Mo10V3 (all P<0.01). Intracellular H+ levels also increased, indicating that intracellular pH decreased, after treatment with 50 mg/L Se2Mo10V3 (P<0.05). In addition, intracellular ROS levels increased after treatment with 12.5, 25, or 50 mg/L Se2Mo10V3 (all P<0.05). Finally, Δφm was markedly reduced after treatment with 50 mg/L Se2Mo10V3 (P<0.01).
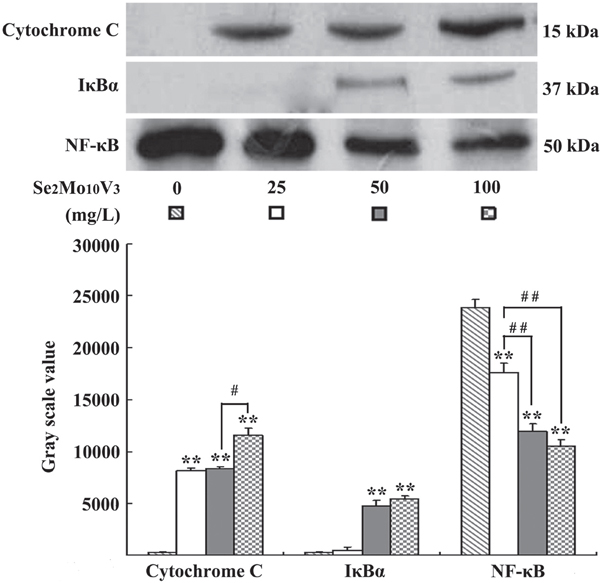
Figure 4: Se2Mo10V3 promoted cytoplasmic cytochrome C level, inhibited cytoplasmic IκBα degradation and reduced nuclear NF-κB level in K562 cells in a Western blotting experiment. Data are expressed as means ± S.D. of three independent experiments. *P<0.05, **P<0.01 versus control cells.
Se2Mo10V3 promoted cytoplasmic cytochrome C level, inhibited cytoplasmic IκBα degradation and reduced nuclear NF-κB level in K562 cells
Western blotting results are shown in Figure 5. Cytoplasmic cytochrome C levels were increased in cells treated with 25, 50, or 100 mg/L Se2Mo10V3 (all P<0.01), which indicated that the release of cytochrome C from the mitochondria into the cytoplasm was increased. Cytoplasmic IκBα degradations were decreased in cells treated with 50 or 100 mg/L, but not with 25 mg/L Se2Mo10V3 (both P<0.01), and nuclear NF-κB levels were reduced in cells treated with 25, 50, or 100 mg/L Se2Mo10V3 (all P<0.01), which indicated that the translocation of NF-κB from the cytoplasm to the nucleus was inhibited.
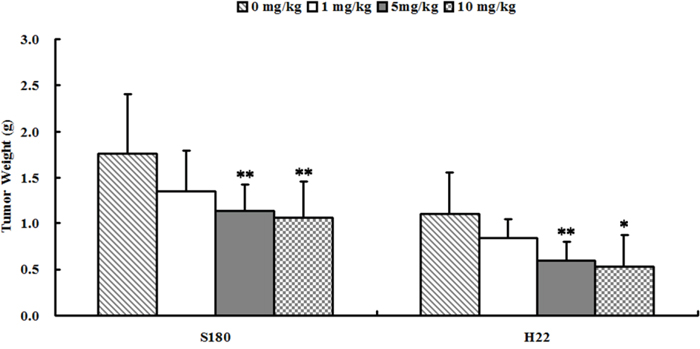
Figure 5: Se2Mo10V3 inhibited the growth of S180 and H22 xenografts in vivo. Data are expressed as means ± S.D. (n=10). *P<0.05, **P<0.01 versus control tumor-bearing mice.
Se2Mo10V3 inhibited S180 and H22 cell growth in vivo
As shown in Figure 2, 5 or 10 mg/kg Se2Mo10V3 (i.g.) inhibited the growth of S180 cells by 35.6 and 39.4%, respectively (all P<0.01). In S180 tumor-bearing mice, tumor weights were 1.762±0.642, 1.351±0.442, 1.135±0.275, and 1.067±0.379 g after treatment with 0, 1, 5, and 10 mg/kg Se2Mo10V3, respectively. Five or 10 mg/kg Se2Mo10V3 (i.g.) similarly inhibited the growth by 46.4 and 52.6%, respectively, in H22 cells (both P<0.05). In H22 tumor-bearing mice, tumor weights were 1.113±0.447, 0.851±0.203, 0.597±0.212, and 0.528±0.351 g after treatment with 0, 1, 5, and 10 mg/kg Se2Mo10V3, respectively.
DISCUSSION
In this study, we demonstrated that Se2Mo10V3, a heteropoly compound containing selenium, had anti-tumor effects both in vitro and in vivo. Furthermore, we explored the mechanisms underlying its anti-tumor effects in K562 cells.
Apoptosis, a genetically controlled mechanism essential for the maintenance of tissue homeostasis and proper development, is crucial to the anti-tumor effects of many chemotherapeutics [9]. Here, we observed typical apoptotic morphology, including chromatin fragmentation, condensation and margination, cytoplasm vacuoles, and the absence of cytolemma microvilli, in Se2Mo10V3-treated K562 cells here. Ca2+ and Mg2+ are key intracellular messengers that play important roles in the induction of apoptosis. Increases in cytosolic Ca2+ and Mg2+ levels have been linked to the activation of a Ca2+/Mg2+ dependent endonuclease which cleaves DNA to generate nucleosomal fragments (180-200 bp) during apoptosis [10, 11]. Here, we confirmed that Se2Mo10V3 increased intracellular Ca2+ and Mg2+ levels in K562 cells. Accumulating data also indicates that ROS have beneficial anti-cancer effects. For instance, increasing ROS levels can enhance apoptosis and thereby inhibit tumor growth [12]. Selenium can also induce the generation of ROS and apoptosis in tumor cells [13]. Moreover, Na5SeV5O18.H20, an analog of Se2Mo10V3, induced an increase in ROS levels in K562 cells [7]; here, Se2Mo10V3 also increased ROS levels in K562 cells. Efficient pH maintenance reduces apoptosis, suggesting that intracellular pH homeostasis and tumor development may be linked [14], and decreasing pH is a hallmark of apoptosis [7]. Consistent with these findings, we observed here that Se2Mo10V3 increased H+ levels, which corresponds to a decrease in pH, in K562 cells. During apoptosis, increasing mitochondrial permeability reduces Δφm and triggers release of cytochrome C into the cytoplasm [15]. Here, Se2Mo10V3 decreased Δφm and triggered the subsequent release of cytochrome C in K562 cells. Changes in the concentration of intracellular ions such as Ca2+ and Mg2+ may also induce mitochondrial impairment and reduce Δφm [16]. It is therefore possible that, in Se2Mo10V3-treated K562 cells, decreased Δφm and cytochrome C release were triggered in part by increased levels of Ca2+ and Mg2+. In conclusion, the results of this study together with previous findings indicate conclusively that Se2Mo10V3 induces the apoptosis of K562 cells.
To determine whether Se2Mo10V3-induced apoptosis is mediated by the NF-κB/IκBα pathway, we also examined changes in NF-κB and IκBα. NF-κB translocation is normally inhibited by IκB proteins that sequester it in the cytoplasm after they are phosphorylated by IκB kinase. Degradation of IκBα thus allows nuclear translocation of NF-κB [17, 18]. It has been reported that NF-κB/IκBα is constitutively active in breast, colon, pancreatic, ovarian, lymphoma, and melanoma cancer cells. Furthermore, inhibition of this pathway using chemotherapeutics may be able to reverse or halt the growth and spreading of tumors [19]. Selenium compounds also inhibit NF-κB in tumor cells [17]. Here, Se2Mo10V3 inhibited the degradation of IκBα and the translocation of NF-κB in K562 cells. Shishodia and colleagues demonstrated that translocated NF-κB activates the expression of anti-apoptotic genes such as Bcl-2 and Bcl-xL, and the suppression of NF-κB inhibited the expression of these genes, thereby promoting apoptosis [20]. Immunocytochemistry also revealed that Se2Mo10V3 treatment decreased Bcl-2 expression and increased Bax expression in K562 (data not shown). An elevated Bax/Bcl-2 ratio increases spontaneous self-oligomerization of those molecules, which facilitates the reduction of Δφm, the release of cytochrome C into the cytoplasm, and, ultimately, apoptosis [21]. Based on these findings, it is likely that decreased Δφm and the release of cytochrome C in Se2Mo10V3-treated K562 cells are triggered in part by an elevated Bax/Bcl-2 ratio.
In summary, Se2Mo10V3 inhibits tumor growth both in vitro and in vivo and induces apoptosis in K562 cells at least in part by inhibiting NF-κB/IκBα. Our results indicate that Se2Mo10V3 may be a useful component of clinical therapy. Further research is needed to examine the anti-leukocythemia effects of Se2Mo10V3 in vivo and to evaluate its preclinical safety on animals.
MATERIALS AND METHODS
Drugs and chemicals
Se2Mo10V3 (yellow crystal, purity >99%) was synthesized by the State Key Laboratory of Applied Organic Chemistry, Lanzhou University, China. Briefly, solution 1 (0.29 g NH4VO3 dissolved in 10 mL hot water) and solution 2 (1.85 g (NH4)6Mo7O24·4H2O dissolved in 8 mL hot water) were blended and 0.063 g/mL Na2SeO4 were added while stirring (solution 3). Solution 3 was adjusted to a pH of 2 using H2SO4 (1:1), and its color changed from orange red to crimson (solution 4). Solution 4 was stirred with reflux at 90°C for 10 h (solution 5). Solution 5 was filtered while hot. The filtrate was placed at room temperature and the yellow crystals separated out quickly. After vacuum filtration, the crystal was washed using an aqueous solution (pH was adjusted to 2 using H2SO4 (1:1)) and dried. The weight of the crude product was 0.68g. The crude product was recrystallized 2 times using an aqueous solution (pH=2). 1.12g yellow crystal was obtained (Se2Mo10V3). The yellow crystal decomposed readily in the air and was slightly soluble in water, methanol, DMSO, and acetone [22]. RPMI-1640 nutrient solution was purchased from Invitrogen. Fetal bovine serum was purchased from Sijiqing Co. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) was purchased from Sigma.
Cell culture
The human chronic myelogenous leukemia K562 cell line was supplied by Shanghai Cell Bank, Chinese Academy of Sciences and grown in RPMI-1640 nutrient solution supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 100 units/mL penicillin, and 100 μg/mL streptomycin in a humidified 5% CO2 incubator at 37°C. The in vitro experiments were performed using a concentration of 1×105 cells/mL.
MTT assay
Cytotoxicity was measured in an MTT assay with slight modifications. K562 cells were treated with 12.5, 25, 50, or 100 mg/L Se2Mo10V3 for 24, 48, or 72 h. MTT solution was then added and the cells were incubated for an additional 4 h. The formazan crystals were dissolved with SDS solution and quantified using a microplate reader (ELx800, Bio-TEK). Cytotoxicity is presented as percent inhibition relative to the control cells. The half-maximal inhibitory concentrations (IC50) for Se2Mo10V3 in K562 cells were calculated using IC50 calculation software.
Transmission electron microscopy (TEM)
K562 cell morphology was examined using TEM. Cells were treated with 50 mg/L Se2Mo10V3 for 24 h. Cell precipitates were then fixed with glutaraldehyde, post- fixed with osmium tetroxide, dehydrated with a graded alcohol series, immersed in epoxy resin and acetone, and embedded in epoxy resin. Ultra-thin sections were then prepared, dyed using uranyl acetate and lead citrate, and examined with a JEM-100 CX-II TEM (Jeol).
Confocal laser scanning microscopy (CLSM)
Levels of Ca2+, Mg2+, H+, and reactive oxygen species (ROS) and mitochondrial membrane potential (Δφm) in K562 cells were determined by CLSM. Cells were treated with 12.5, 25, or 50 mg/L Se2Mo10V3 for 24 h. The cells were then incubated with Fluo-3/AM (5 μmol/L), Mag-fluo-4 (5 mmol/L), Carboxy SNARF-1/AM (10 μmol/L), 2′,7′-dichlorofluorescin diacetete D-399 (5 μg/mL), and Mito Tracker Green FM (1.25 μmol/L) (Molecular Probes, OR, USA) for 30-45 min at 37°C to determine the intracellular Ca2+, Mg2+, H+, and ROS levels and Δφm, respectively. Cells were then washed with RPMI-1640 medium and examined under a TCS SP2 CLSM (Leica, Germany). The results were analyzed with Leica Confocal software.
Western blotting
Cytochrome C, IκBα, and NF-κB in K562 cells were detected by Western blotting. Cells were treated with 25, 50, or 100 mg/L Se2Mo10V3 for 24 h, resuspended in buffer A (10 mM Hepes-NaOH (pH 7.8), 15 mM KCl, 1 mM MgCl2, 0.1 mM EDTA, 1 mM DTT, 1 mM PMSF, 1 mg/L Leupeptin, and 1% NP-40), and incubated on ice for 30 min. The samples were then centrifuged at 12,000 ×g for 30 min at 4°C and the supernatants were used as cytoplasmic extracts. The remaining pellets were resuspended in buffer B (20 mM Hepes-NaOH (pH 7.9), 1.5 mM MgCl2, 0.42 M NaCl, 0.2 mM EDTA, 25% glycerol, 0.5 mM DTT, 0.5 mM PMSF, and 1 mg/L Leupeptin), incubated on ice for 30 min, and centrifuged at 14,000 ×g for 30 min at 4°C. The supernatants were used as nuclear extracts. All supernatants were quantified and run on 12% or 15% SDS polyacrylamide gels. Separated proteins were transferred from the gels to nitrocellulose membranes. The remaining procedures were performed according to the instructions provided with the LumiGLO Western blotting kit (KPL, MD, USA). Mouse monoclonal anti-cytochrome C (1:1000) (Invitrogen, CA, USA), rabbit monoclonal anti-IκBα (1:1000) (Abcam, MA, USA), and rabbit monoclonal anti-NF-κB (1:250) (Zymed Laboratory, CA, USA) were used as primary antibodies. Immunoblotting results were semi-quantified using Quantity One software (Bio-Rad).
Animals and xenograft tumor experiments
5-week old female mice (species: Kun-Ming; strain: Swiss) weighing 20.0±2.0 g were purchased from the GLP Laboratory, Lanzhou University (Grade II, Certificate No. 14-005). They were housed at 23±1 °C under 12h light/12h dark conditions with ad libitum access to food and water. All animal experiments were performed in accordance with relevant guidelines and regulations approved by the Experimental Animal Research Committee of Lanzhou University. According to standard xenograft tumor research protocols, mice were given subcutaneous injections of 3×106 sarcoma 180 (S180) or hepatoma 22 (H22) cells in the exponential growth phase into the right axillary fossa on day 0. On day 1, mice were randomly divided into control, low, middle, and high dose groups and were treated daily with 0.9% normal saline or 1, 5, or 10 mg/kg Se2Mo10V3, i.g., respectively (n=10 per group). The mice were sacrificed on day 10 and the tumors were removed and weighed. Inhibition of tumor growth was evaluated using percent inhibition relative to tumor weights in control tumor-bearing mice.
Statistical analysis
Data are shown as means ± S.D. of three independent experiments. Statistical analysis were performed using one-way ANOVA and the LSD method in SPSS 22.0 software. Differences were considered statistically significant when P<0.05.
Abbreviations
Se2Mo10V3, (NH4)2H15Se2VIMo10V3O52·2H2O. IC50, half-maximal inhibitory concentration. TEM, transmission electron microscope. Δφm, mitochondrial membrane potential. CLSM, confocallaser scanning microscopy. S180, sarcoma 180. H22, hepatoma 22.
Author contributions
H.-N. Z. performed the experiments. W.-L. F. helped to perform the in vitro experiment. C.-N. A. wrote the manuscript, prepared the figures and supplied publication costs. W.-G. L. designed the project. All authors reviewed the manuscript.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
FUNDING
This work was supported by Science and Technology Program of Beijing Municipal Education Commission (KM201810025002) and Health Science Key Program of Hebei Province (ZD20140029).
REFERENCES
1. Alfthan G, Eurola M, Ekholm P, Venäläinen ER, Root T, Korkalainen K, Hartikainen H, Salminen P, Hietaniemi V, Aspila P, Aro A. Effects of nationwide addition of selenium to fertilizers on foods, and animal and human health in Finland: from deficiency to optimal selenium status of the population. J Trace Elem Med Bio. 2015; 31: 142-147.
2. Navarro-Alarcon M, Cabrera-Vique C. Selenium in food and the human body: a review. Sci Total Environ. 2008; 400: 115-141.
3. Klein EA. Selenium: epidemiology and basic science. J Urol. 2004; 171: S50-S53.
4. Endo M, Hasegawa H, Kaneko T, Kanno C, Monma T, Kano M, Shinohara F, Takahashi T. Antitumor activity of selenium compounds and its underlying mechanism in human oral squamous cell carcinoma cells: a preliminary study. J Oral Maxil Surg. 2017; 29: 17-23.
5. de Miranda JX, Andrade Fde O, Conti AD, Dagli ML, Moreno FS, Ong TP. Effects of selenium compounds on proliferation and epigenetic marks of breast cancer cells. J Trace Elem Med Biol. 2014; 28: 486-491.
6. Zeng HW, Combs GF Jr. Selenium as an anticancer nutrient: roles in cell proliferation and tumor cell invasion. J Nutr Biochem. 2008; 19: 1-7.
7. Yang JY, Wang ZR. The antitumor effects of selenium compound Na5SeV5O18·3H2O in K562 cell. Arch Pharm Res. 2006; 29: 859-865.
8. Fernandes AP, Gandin V. Selenium compounds as therapeutic agents in cancer. Biochim Biophys Acta 2015; 1850: 1642-1660.
9. Vázquez R, Riveiro ME, Vermeulen M, Mondillo C, Coombes PH, Crouch NR, Ismail F, Mulholland DA, Baldi A, Shayo C, Davio C. Toddaculin, a natural coumarin from Toddalia asiatica, induces differentiation and apoptosis in U-937 leukemic cells. Phytomedicine. 2012; 19: 737-746.
10. McConkey DJ, Orrenius S. The role of calcium in the regulation of apoptosis. J Leukocyte Biol. 1996; 59: 775-783.
11. Cain K, Inayat-Hussain SH, Kokileva L, Cohen GM. DNA cleavage in rat liver nuclei activated by Mg2+ or Ca2+ + Mg2+ is inhibited by a variety of structurally unrelated inhibitors. Biochem Cell Biol. 1994; 72: 631-638.
12. Salganik RI. The benefits and hazards of antioxidants: controlling apoptosis and other protective mechanisms in cancer patients and the human population. J Am Coll Nutr. 2001; 20: 464S-472S.
13. Jung U, Zheng X, Yoon SO, Chung AS. Se-methylselenocysteine induces apoptosis mediated by reactive oxygen species in HL-60 cells. Free Radic Bio Med. 2001; 31: 479-489.
14. Wenzel U, Daniel H. Early and late apoptosis events in human transformed and non-transformed colonocytes are independent on intracellular acidification. Cell Physiol Biochem. 2004; 14: 65-76.
15. Iannolo G, Conticello C, Memeo L, De Maria R. Apoptosis in normal and cancer stem cells. Crit Rev Oncol Hematol. 2008; 66: 42-51.
16. Bolduc JS, Denizeau F, Jumarie C. Cadmium-induced mitochondrial membrane-potential dissipation does not necessarily require cytosolic oxidative stress: studies using rhodamine-123 fluorescence unquenching. Toxicol Sci. 2004; 77: 299-306.
17. Gasparian AV, Yao YJ, Lü J, Yemelyanov AY, Lyakh LA, Slaga TJ, Budunova IV. Selenium compounds inhibit IκB kinase (IKK) and nuclear factor-κB (NF-κB) in prostate cancer cells. Mol Cancer Ther. 2002; 1: 1079-1087.
18. Wu H, Jiang K, Yin N, Ma X, Zhao G, Qiu C, Deng G. Thymol mitigates lipopolysaccharide-induced endometritis by regulating the TLR4- and ROS-mediated NF-κB signaling pathways. Oncotarget. 2017; 8: 20042-20055. doi: 10.18632/oncotarget.15373.
19. Miller SC, Huang R, Sakamuru S, Shukla SJ, Attene-Ramos MS, Shinn P, Van Leer D, Leister W, Austin CP, Xia M. Identification of known drugs that act as inhibitors of NF-kB signaling and their mechanism of action. Biochem Pharmacol. 2010; 79: 1272-1280.
20. Shishodia S, Amin HM, Lai R, Aggarwal BB. Curcumin (diferuloylmethane) inhibits constitutive NF-κB activation, induces G1/S arrest, suppresses proliferation, and induces apoptosis in mantle cell lymphoma. Biochem Pharmcol. 2005; 70: 700-713.
21. Rudner J, Elsaesser SJ, Müller AC, Belka C, Jendrossek V. Differential effects of anti-apoptotic Bcl-2 family members Mcl-1, Bcl-2, and Bcl-xL on Celecoxib-induced apoptosis. Biochem Pharmcol. 2010; 79: 10-20.
22. Ju ZH, Wu JG. Studies on some heteropoly compounds containing selenium 68-75 (Graduate Dissertation Library of Lanzhou University, 2004).