
INTRODUCTION
Gastric cancer (GC) is the one of the most common carcinoma and the second leading cause of cancer-related death worldwide [1, 2]. The only potentially curative treatment for GC is R0 surgical resection [3]. However, most GC is diagnosed at an advanced stage [4], which is the major cause of unsatisfied overall survival (OS). In addition, more than 50% of patients will experience disease recurrence after surgery [5]. Therefore, despite advances in surgical intervention and chemotherapy, the overall prognosis of patients with advanced GC remains poor [6]. Thus, it is still an urgent need to identify novel biomarkers which correlate with GC tumorigenesis and progression.
Serine/arginine-rich domain proteins (SR proteins) mainly consist of the splicing factors for small nuclear ribonucleoprotein particles (snRNPs) and non-snRNP that concentrate in 'speckles' in the nucleus of interphase cells. SR protein-specific kinase-1 (SRPK1) was firstly identified in 1994 and can phosphorylate one of the SR proteins, namely SC35 [7]. The association of phosphorylation of SR proteins with SRPK1 activity, together with the fact that SRPK1 was the only major kinase for SR proteins obtained during purification, strongly suggests that SRPK1 is responsible for phosphorylation of SR proteins during the cell cycle in vivo [8–10]. Also, SRPK1 is highly expressed in testis, revealing its role in regulating cell division [11].
SRPK1 as well as its downstream targets have been shown to be involved in numerous biological and pathological processes. Dysregulation of SRPK1 has been reported in several cancer types, such as colon, breast, prostate, pancreas and lung cancer [12–15]. High expression of SRPK1 can induce hyper-phosphorylation of SRSF1 protein [16], subsequently increase the transcription and translation of certain proteins, including VEGFs, rising up its role as an oncogenic protein. However, a recent study reported that ablation of SRPK1 in mouse embryonic fibroblasts induced cell transformation and promote tumorigenesis [17]. Therefore, the role of SRPK1 seems distinct in different cell types.
Here in this study, we firstly explored the expression level of SRPK1 in clinical GC tissues. Statistical analyzes revealed that SRPK1 high expression was correlated with advanced tumor stage and poor prognosis. The clinical results promoted us to further investigate the underlying mechanisms of SRPK1 in regulating GC progression. Cellular experiments showed that SRPK1 can enhance cell proliferation and invasion via AKT and ERK signaling pathways, whereas PP2A (protein phosphatase 2A) and DUSP6 (dual specificity phosphatase 6) both attenuated its oncogenic effects.
RESULTS
Patients characteristics
For the 158 cases enrolled in this study, 105 cases (66.5%) were male. Most of the tumor localization was in the gastric body (67/158, 42.4%) or pylorus (61/158, 38.6%). At the time of tumor resection, the largest tumor diameter was less than 5.0 cm in 91 patients (57.6%), and the tumor already invaded into the subserosa or serosa layer (tumor invasion depth as T3-T4) in 104 cases (65.8%). Nighty-five patients (60.1%) were classified as TNM stage III-IV at the time of diagnosis. The serious invasion depth and advanced TNM stage mainly resulted from the unobvious disease phenomenon at early stage. The detailed patients’ information was provided in Table 1.
Table 1: Overview for the characteristics of the cohort
Clinicopathologic features |
Case number |
Percentage |
|---|---|---|
Age |
|
|
≤55 ys |
61 |
38.6% |
>55 ys |
97 |
61.4% |
Gender |
|
|
Female |
53 |
33.5% |
Male |
105 |
66.5% |
Localization |
|
|
Upper |
30 |
19.0% |
Middle |
67 |
42.4% |
Lower |
61 |
38.6% |
Tumor size |
|
|
≤5.0 cm |
91 |
57.6% |
>5.0 cm |
67 |
42.4% |
Invasion depth |
|
|
T1-T2 |
54 |
34.2% |
T3-T4 |
104 |
65.8% |
Differentiation |
|
|
Well |
15 |
9.5% |
Modern |
74 |
46.8% |
Poor |
69 |
43.7% |
TNM stage |
|
|
I-II |
63 |
39.9% |
III-IV |
95 |
60.1% |
SRPK1 expression |
|
|
Low |
75 |
47.5% |
High |
83 |
52.5% |
High SRPK1 expression was correlated with advanced tumor stage
SRPK1 expression was localized to the cytoplasm (Figure 1A) in GC tissues, and 52.5% (83/158) of the cases were categorized as high expression, while 47.5% (75/158) were grouped as low expression. Importantly, the protein expression level of SRPK1 was positively associated with the tumor stage (P<0.001, Figure 1B, Table 2). Therefore, we further collected 28 pairs of fresh-frozen tissues, among them, 4 cases were diagnosed with TNM stage I, 7 cases with stage II, 14 cases with stage III, and the other 3 cases with stage IV. By conducting RT-qPCR assay, we found that the mRNA level of SRPK1 was also correlated with TNM stages (P=0.047, Figure 1C), which was consistent with the protein levels.
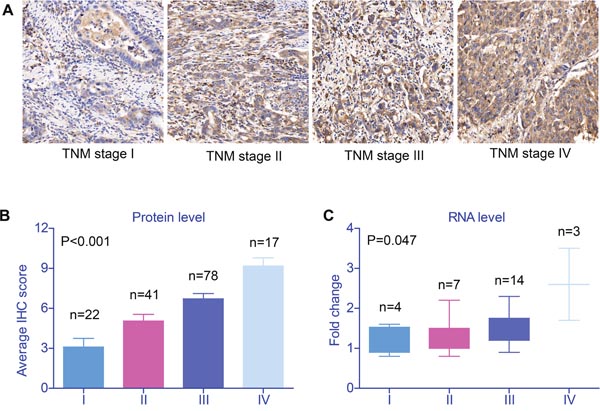
Figure 1: Validation of SRPK1 expression in gastric cancer tissues. (A) SRPK1 protein expression as determined in an IHC assay of tissues from patients with gastric adenocarcinoma. Among 158 patients, tissues from 83 showed high SRPK1 staining, mainly in the cytoplasm and partly in the nucleus of cancer cells (IHC score≥8). Moreover, both the protein (B) and RNA (C) levels of SRPK1 in gastric adenocarcinoma tissues were positively correlated with tumor stage (P<0.001 and P=0.047, respectively). Data was from three independent experiments and statistical analyzes were conducted with One-way ANOVA test, P value was showed correspondingly.
Table 2: Correlations between SRPK1 expression with clinicopathological factors
Clinicopathologic features |
SRPK1 expression level |
Chi-square |
|
|---|---|---|---|
Low (n=75) |
High (n=83) |
||
Age |
|
|
|
≤55 ys |
30 |
31 |
0.733 |
>55 ys |
45 |
52 |
|
Gender |
|
|
|
Female |
30 |
23 |
0.102 |
Male |
45 |
60 |
|
Localization |
|
|
|
Upper |
11 |
19 |
0.414 |
Middle |
33 |
34 |
|
Lower |
31 |
30 |
|
Tumor size |
|
|
|
≤5.0 cm |
48 |
43 |
0.121 |
>5.0 cm |
27 |
40 |
|
Invasion depth |
|
|
|
T1-T2 |
35 |
19 |
0.002* |
T3-T4 |
40 |
64 |
|
Differentiation |
|
|
|
Well |
8 |
7 |
0.658 |
Modern |
37 |
37 |
|
Poor |
30 |
39 |
|
TNM stage |
|
|
|
I-II |
44 |
19 |
<0.001* |
III-IV |
31 |
64 |
|
SRPK1 was an unfavorable biomarker for the prognosis of GC patients
The overall 5-year survival (5-year OS) of the cohort was 59.6%, and the median survival time was 53.0 months. Kaplan–Meier plots (Figure 2) showed that patients with cardia tumor location showed poorer prognosis (P=0.016). Conventional prognostic factors also included tumor invasion depth (P=0.044) and TNM stage (P<0.001). Interestingly, high expression of SRPK1 also indicated unfavorable clinical outcomes (P=0.001, Table 3). In contrast, patients’ age, gender, tumor size or tumor differentiation showed no statistical significance.
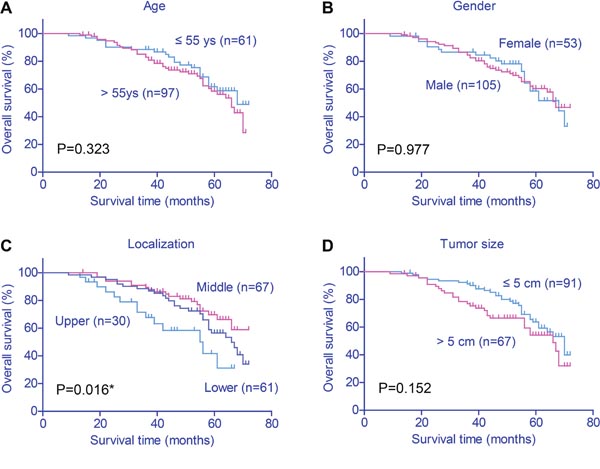
Figure 2: Kaplan–Meier survival plots of gastric cancer patients. The effects of various clinicopathological variables on prognosis were assessed by Kaplan–Meier survival analysis and log-rank test. Tumor location (P=0.016), invasion depth (P=0.044), TNM stage (P<0.001) and SRPK1 expression (P=0.001) were all associated with the clinical outcomes of gastric cancer patients.
Table 3: Kaplan-Meier univariate survival analysis for GC
Clinicopathologic features |
5-year OS |
OS months |
Univariate |
|---|---|---|---|
Age |
|
|
|
≤55 ys |
61.6% |
59.7 ± 2.3 |
0.323 |
>55 ys |
58.4% |
57.1 ± 1.8 |
|
Gender |
|
|
|
Female |
58.5% |
58.1 ± 2.4 |
0.977 |
Male |
60.3% |
58.5 ± 1.8 |
|
Localization |
|
|
|
Upper |
41.7% |
48.4 ± 3.6 |
0.016* |
Middle |
69.7% |
61.9 ± 2.0 |
|
Lower |
56.5% |
58.5 ± 2.2 |
|
Tumor size |
|
|
|
≤5.0 cm |
63.6% |
60.6 ± 1.7 |
0.152 |
>5.0 cm |
54.2% |
55.1 ± 2.5 |
|
Invasion depth |
|
|
|
T1-T2 |
68.9% |
62.5 ± 2.0 |
0.044* |
T3-T4 |
54.5% |
55.9 ± 1.9 |
|
Differentiation |
|
|
|
Well |
86.2% |
66.0 ± 3.0 |
0.092 |
Modern |
62.3% |
59.8 ± 2.0 |
|
Poor |
51.1% |
54.3 ± 2.4 |
|
TNM stage |
|
|
|
I-II |
77.3% |
64.8 ± 1.6 |
<0.001* |
III-IV |
46.6% |
52.2 ± 1.9 |
|
SRPK1 expression |
|
|
|
Low |
72.0% |
63.9 ± 1.4 |
0.001* |
High |
46.0% |
52.2 ± 2.3 |
|
Abbreviations: GC, gastric cancer; OS, overall survival; SRPK1, serine-arginine protein kinase 1.
The Kaplan–Meier survival results promoted us to further investigate whether SRPK1 can serve as an independent prognostic biomarker. Cox regression hazard analysis demonstrated that both advanced TNM stage (P=0.014) and high SRPK1 expression (P=0.035, Table 4) were significant risk factors for the OS of GC patients.
Table 4: Multivariate analysis for hazard factors of GC survival
Variables |
HR (95% CI) |
Multivariate P value |
|---|---|---|
Localization |
|
|
Middle or lower |
0.800 (0.545-1.176) |
0.256 |
Upper |
Reference |
|
Invasion depth |
|
|
T1-T2 |
0.848 (0.413-1.743) |
0.654 |
T3-T4 |
Reference |
|
TNM stage |
|
|
III-IV |
2.609 (1.218-5.590) |
0.014* |
I-II |
Reference |
|
SRPK1 expression |
|
|
High |
1.818 (1.044-3.165) |
0.035* |
Low |
Reference |
|
Abbreviations: GC, gastric cancer; HR, hazard ratio; SRPK1, serine-arginine protein kinase 1.
SRPK1 promoted proliferation and invasion of GC cells
The clinical results indicated the potential oncogenic role of SRPK1 in GC, and next we performed cellular studies to verify its detailed functions and mechanisms. Western blot results showed that SRPK1 expression was higher in gastric adenocarcinoma cells (SUN-1 and AGS cells) than normal gastric epithelial cells (GES-1 cells, Figure 3A). Then we conducted overexpression and knockdown of SRPK1 in both SUN-1 and AGS cells (Figure 3B). The proliferation and invasion capacities of tumor cells were tested by CCK-8 and Matrigel-Transwell assays, respectively. As expected, both the cell viability and invasion were up-regulated upon SRPK1 overexpression, whereas SRPK1-siRNA impaired the oncogenic characteristics (Figures 3C-3F).
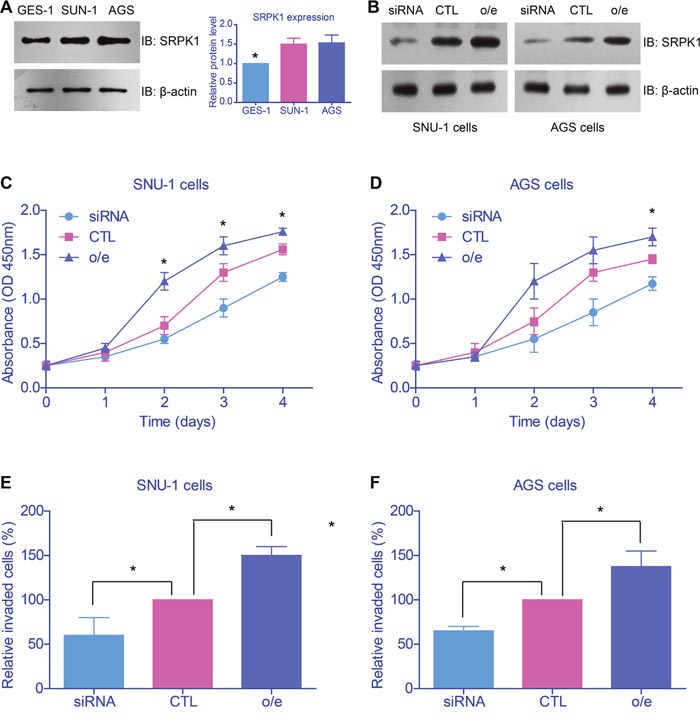
Figure 3: SRPK1 promoted proliferation and invasion of gastric cancer cell lines. (A) Western blot results showed that SRPK1 expression was higher in gastric adenocarcinoma cells (SUN-1 and AGS cells) than normal gastric epithelial cells (GES-1 cells). (B) Transfection efficiency of SRPK1 overexpression (o/e) and knockdown in both SUN-1 and AGS cells were confirmed. The proliferation (C, D) and invasion (E, F) capacities of tumor cells were tested by CCK-8 and Matrigel-Transwell assays, respectively. Both the cell viability and invasion were up-regulated upon SRPK1 overexpression, whereas SRPK1-siRNA impaired the oncogenic characteristics.
AKT and ERK were downstream effectors of SRPK1 in tumorigenesis
Taking into consideration that SRPK1 was an important kinase, we then wanted to test whether its kinetic activity was indispensable in tumor promoting. We generated the kinase-dead mutant (KDM) of SRPK1 by mutating the critical catalytic lysine of amino acid 109 into alanine [18]. Immunoblotting results revealed that SRPK1 overexpression increased the mRNA and protein levels of epithelial-mesenchymal transition (EMT) biomarkers (Figures 4A, 4B). Moreover, IHC results implicated significant correlation between SRPK1 and twist1 (Supplementary Figure 1). On the other hand, the SRPK1-KDM showed no significant oncogenic effects on Slug and Twist1, two of the most important EMT markers. Consistently, transfection of SRPK1-KDM neither exhibited changes on proliferation nor invasion profiles, compared to control cells (Figures 4C, 4D).
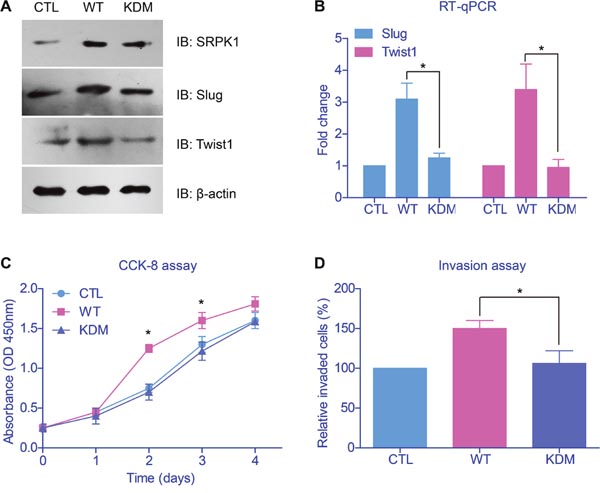
Figure 4: Catalytic activity of SRPK1 was critical in promoting tumor progression. (A) Immunoblotting results revealed that wild type SRPK1 (WT) overexpression increased the protein levels of epithelial-mesenchymal transition (EMT) biomarkers. However, transfection of the kinase-dead mutant of SRPK1 (KDM) showed no significant oncogenic alteration on Slug and Twist1. (B) The mRNA levels of EMT markers were tested by RT-qPCR and showed comparable results with protein levels. Overexpression of SRPK1-KDM neither showed effect on cell proliferation (C) nor invasion (D), compared with control groups.
After we confirmed that the oncogenic role of SRPK1 was functioned by its catalytic activity, we then explored its down-stream effectors. Cellular studies showed that both the phosphorylation of AKT and ERK were enhanced upon SRPK1-overexpression, whereas no significant change with SRPK1-KDM transfection (Figure 5A). Therefore, SRPK1 can phosphorylate and activate AKT and ERK, both are critical in controlling cell proliferation and invasion [19].
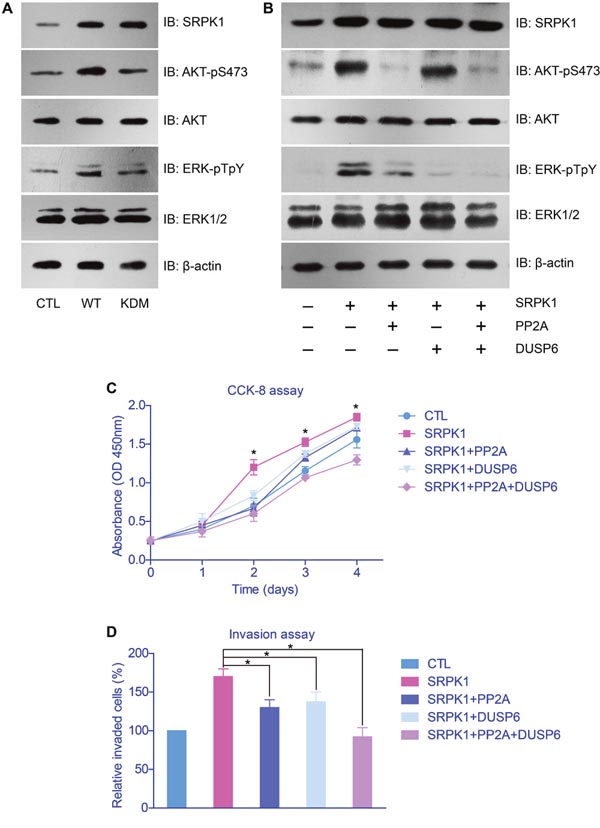
Figure 5: AKT and ERK were down-stream effectors of SRPK1 in gastric cancer cells. (A) SRPK1-overexpression significantly elevated the phosphorylation levels of AKT and ERK proteins, whereas SRPK1-KMD showed little effect. (B) The activation function of AKT and ERK by SRPK1 can be impaired upon overexpression of PP2A and DUSP6, respectively. The facilitated effects of SRPK1 on cell proliferation (C) and invasion (D) were also down-regulated by PP2A and DUSP6, indicating the critical role of AKT and ERK in SRPK1-mediated tumor progression.
PP2A and DUSP6 attenuated the oncogenic effects of SRPK1 with indirect manners
To study the functional mechanisms of SRPK1 in promoting GC progression, we further tested the signaling network on AKT and ERK phosphorylation. It has been reported that PP2A was a phosphatase toward phosphor-AKT [20] and DUSP6 was a phosphatase for phosphor-ERK [21]. By transfected cells with PP2A and/or DUSP6, we found that the activation of AKT and ERK were inhibited, correspondingly (Figure 5B). In addition, the proliferation and invasion capacities of tumor cells were significantly down-regulated after overexpressed PP2A and/or DUSP6, compared with control groups (Figures 5C, 5D).
However, we didn’t find any interaction between SRPK1 with PP2A or DUSP6 (Figure 6A), indicating that PP2A and DUSP6 may regulate down-stream effectors of SRPK1 rather than SRPK1 itself (Figure 6B).
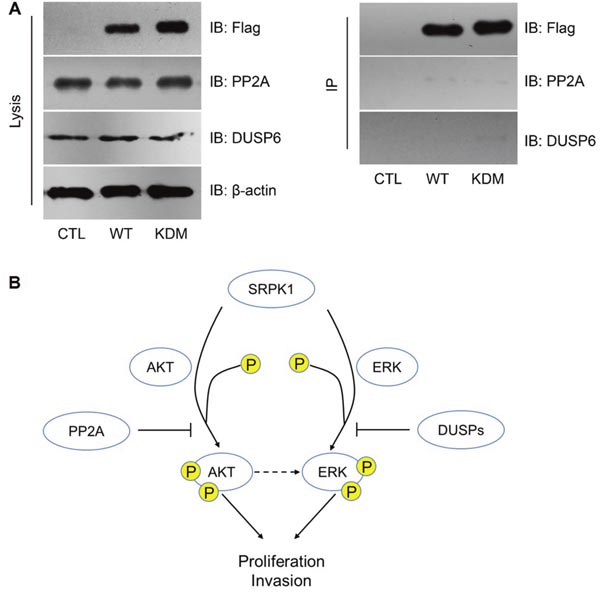
Figure 6: PP2A and DUSP6 inhibited SRPK1 signaling by indirect manners. (A) Neither PP2A nor DUSP6 was interacted with SRPK1, as reflected by immunoprecipitation (IP) assay. (B) Schematically model of the functional network involving SRPK1, AKT, ERK, PP2A and DUSP6.
DISCUSSION
SRPK1 is a protein kinase that specifically phosphorylates proteins containing serine–arginine-rich (SR) domains [22]. SR proteins are involved in regulating several RNA-processing pathways, including RNA stability, alternative splicing, and translation [23]. Both SRPK1 and its downstream targets have been shown to be involved in a number of biological and pathological processes [24]. Although there have been several studies reporting the dysregulation of SRPK1 in malignancies [25], there was no evidence about its role in GC.
Our study detected the expression of SRPK1 in GC for the first time, and demonstrated that SRPK1 was up-regulated in GC cells and tissues. Immunostaining analysis showed that the expression level of SRPK1 protein in histological sections was significantly correlated with clinical characteristics and reduced survival time of gastric cancer patients. Multivariate analysis revealed that SRPK1 expression might be an independent prognostic indicator of survival in GC patients. Taken together, our study suggests that SRPK1 is a novel marker for the prognosis of GC. Furthermore, ectopic overexpression of SRPK1 promoted, while silencing expression of SRPK1 inhibited, the proliferation and invasion of GC cells. In addition, we found that AKT and ERK are involved in the SRPK1 signaling pathway, which was interdicted by PP2A and DUSP6 phosphatases.
Interestingly, although our study revealed that SRPK1 can phosphorylate AKT, AKT was able to phosphorylate SRPK1 reciprocally [26]. However, we didn’t test the phosphorylation levels of SRPK1 due to lacking specific antibodies. Analogically, ERK may also regulate the phosphorylation and functions of SRPK1, but this will need more evidence to verify. Besides ERK and AKT, SRPK1 can activate several other tumor-related proteins in distinct tumor types, such as JNK [27] and VEGFs [28–30]. Certain up-stream regulators for SRPK1 were also identified, including RhoA [31], Wnt [15], TGF-β [27], long non-coding RNAs [25] and human papillomavirus type 1 E1^E4 protein [32].
The complicated signaling network of SRPK1 indicates its invaluable potential in drug development. Indeed, a recent study reported highly potent, selective, and cell active SRPK1 inhibitors, which also have antiangiogenic properties in vivo [33], shedding light on its novel anti-tumor applications.
CONCLUSION
SRPK1 is an oncogenic protein in GC, its high expression is correlated with advanced TNM stage and poor prognosis. SRPK1 functions via AKT and ERK signaling pathway, whereas PP2A and DUSP6 show antagonistic effects.
MATERIALS AND METHODS
Patients and samples
A total of 158 patients was retrospectively included in this study. All patients were diagnosed as gastric adenocarcinoma by pathological examinations, and all the patients underwent R0 surgical resections in Renmin Hospital Affiliated to Wuhan University from March 2000 to March 2002. During the study, we also collected 28 pairs of tumor tissues with adjacent normal tissues, and were frozen in liquid nitrogen until analysis.
Immunohistochemistry staining (IHC) and IHC evaluation
One hundred fifty-eight formalin-fixed, paraffin-embedded GC tissues were used for SRPK1 IHC experiments. After deparaffined and blocking, antigen-antibody reaction was performed at 4°C overnight. The 3,3′-diaminobenzidine (DAB) reagents were applied to determine the signal from the immune-reaction. All slides were counterstained with hematoxylin. The primary SRPK1 (Abcam, Cambridge, MA, USA; ab58002; dilution 1:100) antibody was used for the immunostaining. Primary antibody was replaced with the phosphate-buffered saline (PBS) as a negative control.
The immunostaining results were assigned by two independent pathologists based on both the staining intensity and proportion of positive cells. The percentage score was calculated based on the proportion of positive cells (0, none; 1, ≤25 %; 2, 26–50 %; 3, 51–75 %; 4, >75 %). The intensity score was evaluated according to the positive intensity (0, none; 1, weak; 2, moderate; 3, strong). The total IHC score for SRPK1 expression, ranging from 0 to 12, was the product of the percentage and intensity scores. The protein expression of SRPK1 was categorized as low (IHC score 1–7) or high (IHC score 8–12) for subsequent statistical analysis.
RNA isolation and reverse transcription quantitively polymerase chain reaction (RT-qPCR)
Total RNA from cultured cells and GC tissues was isolated using the RecoverAll Total Nucleic Acid Isolation Kit (Ambion) according to the manufacturer’s instruction. The cDNA was then synthesized from total RNA using the SuperScript II Reverse Trancriptase (Invitrogen). RT-qPCR was performed with the Applied Biosystems 7500 Sequence Detection system, using iQ™ SYBR Green Supermix (BioRad Laboratories, Hercules, CA, USA). The data was normalized to GAPDH housekeeping gene and calculated as 2−ΔΔCT method [34]. Sequences of the primers for RT-qPCR are shown below:
SRPK1: forward primer, 5′-TAATGATTAT TGTAAAGGAG-3′; and reverse primer, 5′-GAACAACC ATTTCTCTATTT-3′.
Slug: forward primer, 5'-CTTCCTGGTCAAGA AGCA-3'; and reverse primer, 5'-GGGAAATAATCACT GTATGTGTG-3'.
Twist1: forward primer, 5'-GGAGTCCGCAG TCTTACGAG-3'; and reverse primer, 5'-CCAGCTTGAG GGTCTGAATC-3'.
GAPDH: forward prime, 5′-ACATCCCCTCAC CAATAACAAC-3′; and reverse primer: 5′-TAGCCAAAT CATACTGCTCGTC-3′.
Cell culture and transfection
Normal human gastric epithelial cells (GES-1 cells) were purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). Human gastric adenocarcinoma cell lines, SNU-1 cell line and AGS cell line, were also obtained from ATCC. All cells were cultured in DMEM (Life Technologies, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS, Life Technologies, Grand Island, NY, USA) and 1% penicillin/streptomycin (Life Technologies) at 37°C with 5% CO2.
Homo sapiens SRPK1 cDNA in pDONR223 vector was purchased from Addgene (Addgene plasmid 23582) [35], a Flag-tag was further added into the construct by PCR and confirmed by DNA sequencing. The siRNA sequences for SRPK1 was 5'-GATCATCAAATCCAATTA-3’ [36] and synthesized by GenePharma (Shanghai, China).
Both the overexpression and siRNA transfection were conducted using Lipofectamine 2000 (Invitrogen, CA, USA) according to the manufacturer’s protocol.
Western blot
The tissues or cultured cells were lysed by radioimmunoprecipitation assay (RIPA; Santa Cruz Biotechnology, Santa Cruz, California, USA) supplemented with a protease inhibitor and phosphatase inhibitor cocktail (Roche, Shanghai, China). Then, a bicinchoninic acid protein assay kit (Biosharp, Shanghai, China) was used to detect the protein concentration. For electrophoresis, 20 μg of the protein sample was loaded on a 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gel and then transferred into polyvinylidene fluoride membrane (PVDF; Millipore, CA, USA) after electrophoresis. Then, membranes were blocked in Tris-buffered saline Tween 20 (TBST) containing 5% skimmed milk for 1 h. These membranes were incubated with primary antibody overnight at 4°C (SRPK1, ab58002, Abcam; Phospho-ERK (Thr202/Tyr204), #4370, Cell Signaling Technology). After washed with TBST for 3 times, PVDF membranes were further incubated with goat secondary antibody IgG-horseradish peroxidase at room temperature for 1 h. Finally, X-ray film (Kodak, NY, USA) was used to analyze the optical density value of target bands. β-actin was served as an internal standard for normalization. Results of densitometric analysis were measured by ImageJ software.
CCK-8 assay
Cell proliferation was measured using a Cell Counting Kit-8 (Dojindo, Kumamoto, Japan). After transfection and/or drug pre-treatment, SNU-1 and AGS cells were seeded in 96-well plates at 2 × 103 cells/well in triplicate and cultured for designated time (1, 2, 3, 4 days). At each time points, 10 μl of CCK-8 solution was added to each well and incubated for 4 h. The absorbance was measured at 450 nm using a microplate reader. All experiments were repeated three times.
Invasion assay
Cell invasion assay was conducted using Transwell chamber (Costar, Corning, NY, USA) with Matrigel (BD, NJ, USA) and polycarbonic membrane (6.5mmin diameter, 8 μm pore size). Briefly, the Transwell membrane was pre-coated with Matrigel solution and incubated at 37°C for 4 h. The transfected cells were resuspended at a density of 5 × 105 cells/mL in serum-free medium and 100 μL was added into the upper chamber. 500 μL of DMEM medium supplemented with 10% FBS was then added to the lower chamber. After incubation for 48 h, cells that had invaded to the lower side of the membrane were fixed and stained. Stained cells were counted under a microscope in five randomly chosen fields and the average number was calculated [37]. All experiments were repeated three times.
Statistical
All data were analyzed by SPSS 18.0 software and presented as mean ± SD. χ2 test was used to explore the association between SRPK1 expression and clinicopathological variables. Kaplan–Meier curves were constructed and the log-rank test was performed for the analysis of survival data. Multivariate analysis was performed using Cox proportional hazards model, and the hazard ratio (HR) as well as its 95% confidence interval (95% CI) were presented. For cellular experiments, all data were expressed as mean ± SD of three independent experiments performed in triplicate. Statistical significance was conducted with the Student's t-test or One-way ANOVA. P < 0.05 was considered statistically significant.
Ethical
This study was approved by the Ethic Committee of Renmin Hospital Affiliated to Wuhan University. Written consent was obtained from each patient for research purposes. This study complied with the Helsinki Declaration.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015; 65:87-108.
2. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015; 65:87-108.
3. Kamangar F, Dores GM, Anderson WF. Patterns of cancer incidence, mortality, and prevalence across five continents: defining priorities to reduce cancer disparities in different geographic regions of the world. Journal of clinical oncology. 2006; 24:2137-2150.
4. Msika S, Benhamiche AM, Jouve JL, Rat P, Faivre J. Prognostic factors after curative resection for gastric cancer. A population-based study. European journal of cancer. 2000; 36:390-396.
5. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. 2015; 65:5-29.
6. Ueda T, Volinia S, Okumura H, Shimizu M, Taccioli C, Rossi S, Alder H, Liu CG, Oue N, Yasui W, Yoshida K, Sasaki H, Nomura S, et al. Relation between microRNA expression and progression and prognosis of gastric cancer: a microRNA expression analysis. The lancet oncology. 2010; 11:136-146.
7. Gui JF, Lane WS, Fu XD. A serine kinase regulates intracellular localization of splicing factors in the cell cycle. Nature. 1994; 369:678-682.
8. Colwill K, Feng LL, Yeakley JM, Gish GD, Caceres JF, Pawson T, Fu XD. SRPK1 and Clk/Sty protein kinases show distinct substrate specificities for serine/arginine-rich splicing factors. The Journal of biological chemistry. 1996; 271:24569-24575.
9. Ma CT, Hagopian JC, Ghosh G, Fu XD, Adams JA. Regiospecific phosphorylation control of the SR protein ASF/SF2 by SRPK1. Journal of molecular biology. 2009; 390:618-634.
10. Sellis D, Drosou V, Vlachakis D, Voukkalis N, Giannakouros T, Vlassi M. Phosphorylation of the arginine/serine repeats of lamin B receptor by SRPK1-insights from molecular dynamics simulations. Biochimica et biophysica acta. 2012; 1820:44-55.
11. Papoutsopoulou S, Nikolakaki E, Chalepakis G, Kruft V, Chevaillier P, Giannakouros T. SR protein-specific kinase 1 is highly expressed in testis and phosphorylates protamine 1. Nucleic acids research. 1999; 27:2972-2980.
12. Hayes GM, Carrigan PE, Miller LJ. Serine-arginine protein kinase 1 overexpression is associated with tumorigenic imbalance in mitogen-activated protein kinase pathways in breast, colonic, and pancreatic carcinomas. Cancer research. 2007; 67:2072-2080.
13. Mavrou A, Brakspear K, Hamdollah-Zadeh M, Damodaran G, Babaei-Jadidi R, Oxley J, Gillatt DA, Ladomery MR, Harper SJ, Bates DO, Oltean S. Serine-arginine protein kinase 1 (SRPK1) inhibition as a potential novel targeted therapeutic strategy in prostate cancer. Oncogene. 2015; 34:4311-4319.
14. Odunsi K, Mhawech-Fauceglia P, Andrews C, Beck A, Amuwo O, Lele S, Black JD, Huang RY. Elevated expression of the serine-arginine protein kinase 1 gene in ovarian cancer and its role in Cisplatin cytotoxicity in vitro. PloS one. 2012; 7:e51030.
15. Gong L, Song J, Lin X, Wei F, Zhang C, Wang Z, Zhu J, Wu S, Chen Y, Liang J, Fu X, Lu J, Zhou C, Song L. Serine-arginine protein kinase 1 promotes a cancer stem cell-like phenotype through activation of Wnt/beta-catenin signalling in NSCLC. The Journal of pathology. 2016; 240:184-196.
16. da Silva MR, Moreira GA, Goncalves da Silva RA, de Almeida Alves Barbosa E, Pais Siqueira R, Teixera RR, Almeida MR, Silva Junior A, Fietto JL, Bressan GC. Splicing Regulators and Their Roles in Cancer Biology and Therapy. BioMed research international. 2015; 2015:150514.
17. Wang P, Zhou Z, Hu A, Ponte de Albuquerque C, Zhou Y, Hong L, Sierecki E, Ajiro M, Kruhlak M, Harris C, Guan KL, Zheng ZM, Newton AC, et al. Both decreased and increased SRPK1 levels promote cancer by interfering with PHLPP-mediated dephosphorylation of Akt. Molecular cell. 2014; 54:378-391.
18. Zhou B, Li Y, Deng Q, Wang H, Wang Y, Cai B, Han ZG. SRPK1 contributes to malignancy of hepatocellular carcinoma through a possible mechanism involving PI3K/Akt. Molecular and cellular biochemistry. 2013; 379:191-199.
19. Wang L, Ouyang F, Liu X, Wu S, Wu HM, Xu Y, Wang B, Zhu J, Xu X, Zhang L. Overexpressed CISD2 has prognostic value in human gastric cancer and promotes gastric cancer cell proliferation and tumorigenesis via AKT signaling pathway. Oncotarget. 2016; 7:3791-3805. https://doi.org/10.18632/oncotarget.6302.
20. Kuo YC, Huang KY, Yang CH, Yang YS, Lee WY, Chiang CW. Regulation of phosphorylation of Thr-308 of Akt, cell proliferation, and survival by the B55α regulatory subunit targeting of the protein phosphatase 2A holoenzyme to Akt. Journal of Biological Chemistry. 2008; 283:1882-1892.
21. Bermudez O, Marchetti S, Pages G, Gimond C. Post-translational regulation of the ERK phosphatase DUSP6/MKP3 by the mTOR pathway. Oncogene. 2008; 27:3685-3691.
22. Colwill K, Feng LL, Yeakley JM, Gish GD, Cáceres JF, Pawson T, Fu XD. SRPK1 and Clk/Sty protein kinases show distinct substrate specificities for serine/arginine-rich splicing factors. Journal of Biological Chemistry. 1996; 271:24569-24575.
23. Long JC, Caceres JF. The SR protein family of splicing factors: master regulators of gene expression. The Biochemical journal. 2009; 417:15-27.
24. Bullock N, Oltean S. The many faces of SRPK1. The Journal of pathology. 2017; 241:437-440.
25. Hu ZY, Wang XY, Guo WB, Xie LY, Huang YQ, Liu YP, Xiao LW, Li SN, Zhu HF, Li ZG, Kan H. Long non-coding RNA MALAT1 increases AKAP-9 expression by promoting SRPK1-catalyzed SRSF1 phosphorylation in colorectal cancer cells. Oncotarget. 2016; 7:11733-11743. https://doi.org/10.18632/oncotarget.7367.
26. Zhou Z, Qiu J, Liu W, Zhou Y, Plocinik RM, Li H, Hu Q, Ghosh G, Adams JA, Rosenfeld MG. The Akt-SRPK-SR axis constitutes a major pathway in transducing EGF signaling to regulate alternative splicing in the nucleus. Molecular cell. 2012; 47:422-433.
27. Ren G, Sheng L, Liu H, Sun Y, An Y, Li Y. The crucial role of SRPK1 in TGF-beta-induced proliferation and apoptosis in the esophageal squamous cell carcinomas. Medical oncology. 2015; 32:209.
28. Mavrou A, Oltean S. SRPK1 inhibition in prostate cancer: A novel anti-angiogenic treatment through modulation of VEGF alternative splicing. Pharmacological research. 2016; 107:276-281.
29. Bullock N, Potts J, Simpkin AJ, Koupparis A, Harper SJ, Oxley J, Oltean S. Serine-arginine protein kinase 1 (SRPK1), a determinant of angiogenesis, is upregulated in prostate cancer and correlates with disease stage and invasion. Journal of clinical pathology. 2016; 69:171-175.
30. Morooka S, Hoshina M, Kii I, Okabe T, Kojima H, Inoue N, Okuno Y, Denawa M, Yoshida S, Fukuhara J, Ninomiya K, Ikura T, Furuya T, et al. Identification of a Dual Inhibitor of SRPK1 and CK2 That Attenuates Pathological Angiogenesis of Macular Degeneration in Mice. Molecular pharmacology. 2015; 88:316-325.
31. Chang Y, Li L, Zhang L, Guo X, Feng Z, Zhou J, Zhou S, Feng G, Han F, Huang W, Yang J, Huang F. Plexin-B1 indirectly affects glioma invasiveness and angiogenesis by regulating the RhoA/alphavbeta3 signaling pathway and SRPK1. Tumour biology. 2016; 37:11225-11236.
32. Prescott EL, Brimacombe CL, Hartley M, Bell I, Graham S, Roberts S. Human papillomavirus type 1 E1^E4 protein is a potent inhibitor of the serine-arginine (SR) protein kinase SRPK1 and inhibits phosphorylation of host SR proteins and of the viral transcription and replication regulator E2. Journal of virology. 2014; 88:12599-12611.
33. Batson J, Toop HD. Development of Potent, Selective SRPK1 Inhibitors as Potential Topical Therapeutics for Neovascular Eye Disease. ACS Chem Biol. 2017; 12:825-832.
34. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001; 25:402-408.
35. Johannessen CM, Boehm JS, Kim SY, Thomas SR, Wardwell L, Johnson LA, Emery CM, Stransky N, Cogdill AP, Barretina J, Caponigro G, Hieronymus H, Murray RR, et al. COT drives resistance to RAF inhibition through MAP kinase pathway reactivation. Nature. 2010; 468:968-972.
36. Sanidas I, Kotoula V, Ritou E, Daans J, Lenz C, Mairhofer M, Daniilidou M, Kolbus A, Kruft V, Ponsaerts P, Nikolakaki E. The ratio of SRPK1/SRPK1a regulates erythroid differentiation in K562 leukaemic cells. Biochimica et biophysica acta. 2010; 1803:1319-1331.
37. Shang C, Sun L, Zhang J, Zhao B, Chen X, Xu H, Huang B. Silence of cancer susceptibility candidate 9 inhibits gastric cancer and reverses chemoresistance. Oncotarget. 2017; 8:15393-15398. https://doi.org/10.18632/oncotarget.14871.