
INTRODUCTION
Globally, approximately 15% of heterosexual couples suffer from infertility [1–6], in which non-obstructive azoospermia (NOA) and severe oligozoospermia represent two of the predominant phenotypes relating to severely defective spermatogenesis. Although several factors can lead to male infertility, including malformations of the reproductive tract (e.g., varicocele or cryptorchidism, hypogonadotrophic hypogonadism, karyotype anomalies and Y chromosome microdeletions), infection, and chemical exposure, the effect of genetic predisposition upon male infertility remains to be fully clarified [2]. Therefore, an understanding of the genetic basis of reproductive failure is essential in managing an infertile couple appropriately.
Protamines (PRM), which were first isolated from spermatozoa one century ago, play essential roles in sperm chromatin condensation [7]. Two types of protamine were identified in mammals: protamine 1 (PRM1), which is present in vertebrate species, and protamine 2 (PRM2), which only exists in some mammalian species, including humans and mice [8]. PRM1 and PRM2 (NC_000016.9, GI: 224589807) are closely linked in a stretch of DNA 13–15 kb long on human chromosome 16p13.3, along with the gene encoding transition protein 2 (TNP2); collectively, these genes are categorized as members of the protamine gene family [2]. Previous research has shown that the biological and physiological functions of protamine are involved in a variety of mechanisms: (i) paternal genome packing; (ii) competition and removal of transcription factors and other proteins from the spermatid; and (iii) imprinting of the paternal genome during spermatogenesis [2, 7]. Mutations or polymorphisms within PRM induce conformational changes of the encoded proteins and alter their incorporation into sperm chromatin, leading to sperm defects, although the underlying mechanisms remain largely unknown [8, 9]. Considered as one of the most perplexing disorders in the reproductive field, male factor infertility is prevalent, and its incidence is rising; worryingly, however, the etiology of this condition remains largely elusive.
Since the association between PRM polymorphisms and the risk of male infertility was first reported in 2003, there have been additional investigations of the association between rs201365, rs1646022 rs2070923 and rs737008 and the risk of male infertility among different ethnicities [10–16]. However, the results arising from these studies have been mixed or contradictory, most probably due to relatively small sample size. Interestingly, published association studies of male infertility in Chinese populations did not identify SNPs as susceptible loci, perhaps owing to the stringent P values required to avoid false-positive findings, which dramatically reduces the possibility of revealing a modest effect of some common SNPs upon male infertility, particularly for those SNPs which are potentially functional. Therefore, in the present study, we further investigated the association of some potentially functional PRM SNPs (rs2301365, rs737008, rs35576928, rs1646022 and rs2070923) and TNP1 (rs62189545) with the risk of male infertility in a large scale study of a Chinese population.
RESULTS
Clinical data of the study population
The clinical characteristics of the study population are described in Table 1. Consistently, no significant differences were observed in terms of the relative distributions of age, abstinence time, semen volume, pH of semen and serum hormone index (Testosterone: T, Estradiol: E2 and Luteinizing hormone: LH) when considering cases and controls from study sets (P > 0.05 for all), with the exception of sperm concentration, progressive mobility and Follicle stimulating estrogen (FSH) status (P < 0.05).
Table 1: Characteristics of the study population
Characteristics |
Case (mean ± SD) |
Control (mean ± SD) |
P |
|---|---|---|---|
All subjects |
636 |
442 |
|
Age (year) |
28.56 ± 4.30 |
28.37 ± 4.23 |
0.545 |
Abstinence time (day) |
4.33 ± 1.60 |
5.08 ± 4.09 |
0.521 |
Semen volume (mL) |
3.65 ± 1.76 |
3.51 ± 1.08 |
0.295 |
Sperm concentration (106/mL) |
12.32 ± 15.49 |
72.77 ± 45.21 |
0.000 |
Progressive mobility (%) |
15.29 ± 15.06 |
42.02 ± 9.04 |
0.000 |
pH value of semen |
7.38 ± 0.06 |
7.37 ± 0.07 |
0.404 |
Serum Hormone Index |
|||
T (nmol/L) |
13.85 ± 5.32 |
12.43 ± 4.83 |
0.188 |
E2 (pmol/L) |
115.47 ± 67.21 |
103.87 ± 77.35 |
0.506 |
LH (IU/L) |
6.28 ± 4.90 |
4.62 ± 7.49 |
0.163 |
FSH (FSH IU/L) |
14.69 ± 15.70 |
4.72 ± 2.51 |
0.001 |
T: testosterone; E2: estradiol; LH: lutenizing hormone; FSH: follicle-stimulating hormone. Bold font means significant difference when compared with controls (P < 0.05)
Genetic analyses
Logistic regression analysis
In the control group, all genotype frequencies of SNPs were in line with the Hardy-Weinberg equilibrium (P > 0.05). Genotype distributions of the selected SNPs in cases and controls are summarized in Table 2. For PRM1, rs35576928 showed a GG genotype in all samples. Among the five SNPs (rs737008, rs2301365, rs2070923, rs1646022 and rs62180545) for PRM1, PRM2 and TNP1, no significant differences were found between the cases and controls. Similarly, no significant differences were observed in the distributions of genotype among cases and controls for all SNPs within subgroups (i.e., NOA, severe oligozoospermia, and other types of infertility).
Table 2: Logistic regression analysis of associations between the genotype of PRM1/2 and TNP1 and male infertility risk
Genotype |
Control |
Case |
NOA and SO |
Other infertility |
||||||
|---|---|---|---|---|---|---|---|---|---|---|
N |
P |
OR(95% CI) |
N |
P |
OR(95% CI) |
N |
P |
OR(95% CI) |
||
PRM1 rs737008 230G>T |
||||||||||
GG |
237 |
339 |
0.332 |
ref |
298 |
0.463 |
ref |
41 |
0.216 |
ref |
GT |
159 |
250 |
0.567 |
1.10(0.80-1.53) |
205 |
0.877 |
1.03(0.73-1.45) |
45 |
0.133 |
1.63(0.86-3.08) |
TT |
46 |
47 |
0.233 |
0.71(0.40-1.25) |
41 |
0.246 |
0.71(0.39-1.27) |
6 |
0.627 |
0.73(0.21-2.59) |
GT/TT |
205 |
297 |
0.937 |
1.01(0.74-1.38) |
246 |
0.780 |
0.96(0.69-1.32) |
51 |
0.257 |
1.43(0.77-2.64) |
PRM1 rs2301365 c.-190C>A |
||||||||||
CC |
277 |
378 |
0.655 |
ref |
329 |
0.790 |
ref |
49 |
0.448 |
ref |
CA |
144 |
229 |
0.357 |
1.17(0.84-1.62) |
192 |
0.508 |
1.12(0.80-1.58) |
37 |
0.245 |
1.46(0.77-2.78) |
AA |
21 |
29 |
0.884 |
1.06(0.50-2.22) |
23 |
0.942 |
0.97(0.44-2.13) |
6 |
0.463 |
1.63(0.44-6.03) |
CA/AA |
165 |
258 |
0.520 |
1.11(0.81-1.52) |
215 |
0.556 |
1.10(0.79-1.53) |
43 |
0.209 |
1.48(0.80-2.75) |
PRM2 rs2070923 373T>G |
||||||||||
TT |
233 |
333 |
0.210 |
ref |
292 |
0.307 |
ref |
41 |
0.241 |
ref |
TG |
162 |
258 |
0.510 |
1.12(0.81-1.55) |
213 |
0.774 |
1.05(0.75-1.48) |
45 |
0.162 |
1.57(0.83-2.97) |
GG |
47 |
45 |
0.153 |
0.66(0.38-1.17) |
39 |
0.162 |
0.66(0.36-1.19) |
6 |
0.577 |
0.70(0.20-2.47) |
TG/GG |
209 |
303 |
0.937 |
1.01(0.74-1.38) |
252 |
0.810 |
0.96(0.70-1.33) |
51 |
0.310 |
1.38(0.74-2.54) |
PRM2 rs1646022 298C>G |
||||||||||
CC |
235 |
335 |
0.157 |
ref |
284 |
0.208 |
ref |
51 |
0.538 |
ref |
CG |
166 |
266 |
0.466 |
1.13(0.82-1.56) |
229 |
0.432 |
1.14(0.82-1.60) |
37 |
0.913 |
1.04(0.55-1.96) |
GG |
41 |
35 |
0.115 |
0.61(0.33-1.13) |
31 |
0.170 |
0.64(0.33-1.21) |
4 |
0.285 |
0.44(0.10-1.97) |
CG/GG |
207 |
301 |
0.875 |
1.03(0.75-1.40) |
260 |
0.793 |
1.04(0.76-1.44) |
41 |
0.789 |
0.92(0.50-1.70) |
TNP1 rs62180545 c.139+75A>G |
||||||||||
AA |
389 |
577 |
0.511 |
ref |
489 |
0.734 |
ref |
88 |
0.337 |
ref |
AG |
52 |
57 |
0.247 |
0.74(0.44-1.23) |
53 |
0.436 |
0.81(0.48-1.37) |
4 |
0.141 |
0.34(0.28-1.44) |
GG |
1 |
2 |
0.983 |
0.97(0.60-15.57) |
2 |
0.924 |
1.14(0.07-18.39) |
0 |
1.000 |
- |
AG/GG |
53 |
59 |
0.253 |
0.75(0.45-1.23) |
55 |
0.453 |
0.82(0.49-1.37) |
4 |
0.131 |
0.33(0.08-1.40) |
ref: reference; CI: confidence interval, OR: odds ratio. The results were in bold, if the 95% CI excluded 1 or P < 0.05; NOA: non-obstructive azoospermia; SO: severe oligozoospermia.
Haplotype analysis
One of the goals of the present study was to establish the most shared haplotypes present in the PRM1 and PRM2 genes in the Chinese Han population, taking advantage of the proximity of the two protamine genes and the fact that we have detected three common SNPs in PRM1 and two common SNPs in PRM2. With this information, we defined five haplotypes present in 97.3% of the chromosomes in the Han population, and six additional rare haplotypes present in 2.7% of the chromosomes in our population (Table 3). A linkage disequilibrium (LD) plot for the five SNPs is shown in Figure 1, indicating that rs737008 was strongly linked with both rs2301365 and rs2070923.
Table 3: Haplotype analysis of five SNPs of PRM1/2
Haplotype |
Control |
Case |
NOA and SO |
Other infertility |
||||||
|---|---|---|---|---|---|---|---|---|---|---|
N |
P |
OR(95% CI) |
N |
P |
OR(95% CI) |
N |
P |
OR(95% CI) |
||
rs737008/rs35576928/rs2070923/rs1646022/rs2301365 |
||||||||||
GCTCC |
0.492 |
0.696 |
0.000 |
2.40 (1.91-3.02) |
0.712 |
0.000 |
2.61(2.05-3.33) |
0.581 |
0.131 |
1.41(0.90-2.19) |
GCTGC |
0.216 |
0.026 |
0.000 |
0.10 (0.06-0.16) |
0.015 |
0.000 |
0.05(0.03-0.11) |
0.111 |
0.016 |
0.45(0.23-0.87) |
TCGCA |
0.167 |
0.026 |
0.000 |
0.13(0.08-0.22) |
0.006 |
0.000 |
0.03(0.01-0.09) |
0.148 |
0.618 |
0.86(0.47-1.57) |
TCGCC |
0.057 |
0.008 |
0.000 |
0.13(0.05-0.33) |
0.006 |
0.000 |
0.11(0.04-0.32) |
0.026 |
0.211 |
0.44(0.12-1.64) |
TCGGA |
0.041 |
0.196 |
0.000 |
5.73(3.71-8.84) |
0.207 |
0.000 |
6.18(3.98-9.58) |
0.118 |
0.002 |
3.09(1.48-6.48) |
TCGGC |
0.02 |
0.04 |
0.036 |
2.03(1.04-3.96) |
0.042 |
0.026 |
2.14(1.08-4.23) |
0.016 |
- |
- |
The statistically significant results were in bold, if the 95% CI excluded 1 or P < 0.05.
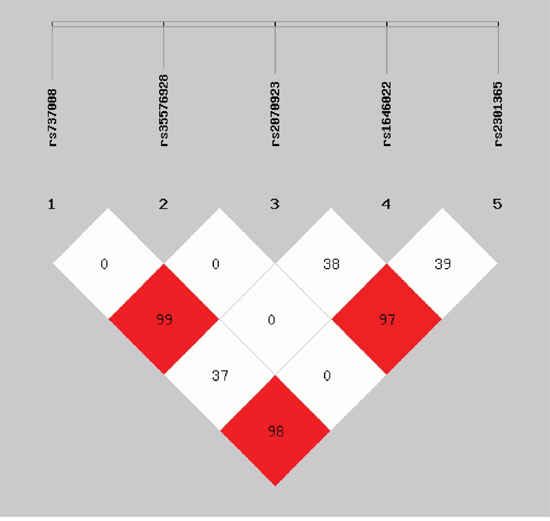
Figure 1: Linkage disequilibrium (LD) plot for five SNPs of PRM1 and PRM2. The D value is displayed as a percentage. One LD block has been identified and shows the involvement of five SNPs. Data showed that rs737008 was strongly linked with both rs2301365 and rs2070923.
The frequency of the shared haplotypes was significantly different between infertile patients and healthy individuals (P < 0.05). As shown in Table 3, six haplotypes were found to be associated with male infertility and especially for the subgroups of NOA and severe oligozoospermia. Three haplotypes exhibited significantly protective effects against male infertility when compared with controls (for the GCTGC haplotype: P = 0.000, OR = 0.10, 95%CI = 0.06–0.16; for the TCGCA haplotype: P = 0.000, OR = 0.13, 95%CI = 0.08–0.22; for the TCGCC haplotype: P = 0.000, OR = 0.10, 95%CI = 0.05–0.33). However, three haplotypes were associated with an increased risk of male infertility (for the TCGGA haplotype: P = 0.000, OR = 5.73, 95%CI = 3.71–8.84; for the GCTCC haplotype: P = 0.000, OR = 2.40, 95%CI = 1.91–3.02; for the TCGGC haplotype: P = 0.036, OR = 2.03, 95%CI = 1.04–3.96). In the subgroup analysis of NOA and severe oligozoospermia, a similar positive effect was found. Nevertheless, in the subgroup analysis of other types of infertility, we unfortunately failed to find a similar effect across several haplotypes, except for the GCTGC haplotype (P = 0.016, OR = 0.45, 95%CI = 0.23–0.87) and the TCGGA haplotype (P = 0.002, OR = 3.09, 95%CI = 1.48–6.48).
Gene–gene interaction analysis
We further evaluated the associations between the combined types of the selected shared SNPs and male infertility (Table 4 and Supplementary Table 1). Individuals with a combined genotype of CGGT (rs1646022/rs737008) were associated with male infertility susceptibility, accompanied by a 1.58-fold increased risk of infertility (P = 0.047, OR = 1.58, 95%CI = 1.01–2.47). At the same time, individuals with the CGAC genotype (rs1646022/rs2301365) were associated with male infertility, accompanied by a 1.68-fold increased risk of infertility (P = 0.023, OR = 1.68, 95%CI = 1.07–2.64).
Table 4: Gene–gene interactions of PRM1 and PRM2 and male infertility risk
Genotype |
Control |
Case |
NOA and SO |
Other infertility |
||||||
|---|---|---|---|---|---|---|---|---|---|---|
N |
OR(95% CI) |
P |
N |
OR(95% CI) |
P |
N |
OR(95% CI) |
P |
||
rs1646022/rs737008 |
||||||||||
GGCC |
115 |
308 |
ref |
0.000 |
280 |
ref |
0.000 |
28 |
ref |
0.559 |
CCGT |
94 |
23 |
0.09(0.05-0.18) |
0.000 |
4 |
0.02(0.00-0.07) |
0.000 |
19 |
0.87(0.36-2.08) |
0.753 |
CCTT |
27 |
4 |
0.05(0.01-0.23) |
0.000 |
0 |
- |
0.998 |
4 |
0.60(0.13-2.86) |
0.521 |
CGGG |
96 |
29 |
0.11(0.06-0.21) |
0.000 |
16 |
0.07(0.03-0.14) |
0.000 |
13 |
0.59(0.23-1.55) |
0.284 |
CGGT |
53 |
225 |
1.58(1.01-2.47) |
0.047 |
201 |
1.55(0.98-2.45) |
0.059 |
24 |
1.85(0.78-4.36) |
0.162 |
CGTT |
16 |
12 |
0.27(0.10-0.74) |
0.011 |
12 |
0.30(0.11-0.81) |
0.180 |
0 |
- |
0.999 |
GGGG |
26 |
2 |
0.03(0.00-0.21) |
0.001 |
2 |
0.03(0.00-0.24) |
0.001 |
0 |
- |
0.998 |
GGGT |
12 |
2 |
0.06(0.01-0.48) |
0.008 |
0 |
- |
0.989 |
2 |
0.67(0.08-5.68) |
0.711 |
GGTT |
3 |
31 |
4.28(0.96-19.06) |
0.056 |
29 |
4.41(0.98-19.74) |
0.053 |
2 |
3.00(0.26-35.33) |
0.383 |
rs1646022/rs2301365 |
||||||||||
CCCC |
139 |
309 |
ref |
0.000 |
278 |
ref |
0 |
31 |
ref |
0.756 |
CCAA |
15 |
4 |
0.13(0.03-0.60) |
0.009 |
0 |
- |
0.999 |
4 |
1.28(0.26-6.36) |
0.767 |
CCAC |
83 |
22 |
0.12(0.06-0.23) |
0.000 |
6 |
0.04(0.01-0.12) |
0.000 |
16 |
0.84(0.34-2.07) |
0.699 |
CGAA |
5 |
6 |
0.48(0.11-2.21) |
0.349 |
6 |
0.54(0.12-2.46) |
0.425 |
0 |
- |
0.999 |
CGAC |
52 |
193 |
1.68(1.07-2.64) |
0.023 |
174 |
1.68(1.07-2.66) |
0.026 |
19 |
1.68(0.70-4.02) |
0.246 |
CGCC |
108 |
67 |
0.27(0.17-0.44) |
0.000 |
49 |
0.22(0.13-038) |
0.000 |
18 |
0.72(0.30-1.710 |
0.453 |
GGAA |
1 |
19 |
6.46(0.81-51.19) |
0.078 |
17 |
6.47(0.81-51.83) |
0.079 |
2 |
6.38(0.38-107.11) |
0.198 |
GGAC |
9 |
14 |
0.65(0.22-1.90) |
0.426 |
12 |
0.62(0.20-1.89) |
0.396 |
2 |
0.91(0.11-7.90) |
0.932 |
GGCC |
30 |
2 |
0.03(0.00-0.22) |
0.001 |
2 |
0.03(0.00-0.25) |
0.001 |
0 |
- |
0.998 |
The statistically significant results were in bold, if the 95% CI excluded 1 or P < 0.05.
For rs737008 in PRM1 and rs1646022 in PRM2, we found a protective effect for male infertility in several combined genotypes (for GTCC: P = 0.000, OR = 0.09, 95%CI = 0.05–0.18; for TTCC: P = 0.000, OR = 0.05, 95%CI = 0.01–0.23; for GGCG: P = 0.000, OR = 0.11, 95%CI = 0.06–0.21; for GTCG: P = 0.011, OR = 0.27, 95%CI = 0.10–0.74; for GGGG: P = 0.001, OR = 0.03, 95%CI = 0.00–0.21; for GTCG: P = 0.008, OR = 0.06, 95%CI = 0.01–0.48). Similarly, for rs2301365 in PRM1 and rs1646022 in PRM2, there was a significant protective effect for male infertility in several combined genotypes (for AACC: P = 0.009, OR = 0.13, 95%CI = 0.03–0.60; for ACCC: P = 0.000, OR = 0.12, 95%CI = 0.06–0.23; for CCCG: P = 0.000, OR = 0.27, 95%CI = 0.17–0.44; for CCGG: P = 0.001, OR = 0.03, 95%CI = 0.00–0.22). However, we failed to find a significant difference for other combined genotypes (Table 4 and Supplementary Table 1). The basic information of six SNPs was described in the Supplementary Table 2.
DISCUSSION
In mammals, male germ cells differentiate from haploid round spermatids into flagella-possessing motile sperm in a process called spermiogenesis [17, 18]. This process is different from somatic cell differentiation in that the majority of the core histones are replaced sequentially, first by transition proteins and then by protamines, thus facilitating chromatin hyper-compaction [7, 18, 19]. This histone-to-protamine transition process represents an excellent model for the investigation of how epigenetic regulators interact with each other to remodel chromatin architecture [20–22].
Thus far, only a few studies have analyzed the correlation between PRM1/2 and TNP1 polymorphisms and particular phenotypes of male infertility [10, 13, 15, 16, 23, 24]. However, recent studies in the field have highlighted the critical role of rs2301365 (c.-190 C>A), located in the 5′-UTR of the PRM1 gene, in controlling haploid-specific developmental programming [2, 9, 11, 15]. Similarly, Jiang et al.,[2] reported that rs737008 (c.230 C>A) and rs1646022 (c.298 G>C) had strong protective effects over male infertility in several subgroups. However, no significant difference was found for other SNPs when comparing infertile cases and controls. Other studies investigated the role of several SNPs in male infertility, but failed to detect any differences in allele frequency between infertile patients and healthy men [2, 13, 14, 23, 25, 26].
These previous results may be attributable to a multitude of factors. Firstly, studies with relatively small sample sizes may lack the necessary power to allow accurate conclusions to be drawn. Secondly, the SNPs investigated may be located on non-sensitive sites. We cannot exclude the possibility that these SNPs may simply represent genetic markers of male infertility in linkage disequilibrium with other mutations or variations, which do play a role in male infertility. Thirdly, the settings and experimental methods differ across different studies. Finally, races living in different latitudes with extreme weather are under the influence of the environment, climatic conditions (air temperature, solar radiation, ultraviolet intensity) and varied dietary habits during the lengthy evolution process, which may affect the mode of action and potency of the SNPs, thus leading to differences in results among populations from certain regions [22, 27–29].
Haplotype analysis is a useful tool in revealing the potential associations of genes that are hidden by the evaluation of PRM1 and PRM2 SNPs individually. The present study identified high linkage disequilibrium among the selected SNPs. We identified five haplotypes that were present in 97.3% of the chromosomes in the Han population, and six additional rare haplotypes that were present in 2.7% of the chromosomes in our population. Several shared SNPs were strongly associated with rs737008 rs2301365 and rs2070923. Three haplotypes (GCTGC, TCGCA and TCGCC) were shown to be significant protective genetic factors for spermatogenesis, compared with the controls. In addition, three haplotypes (TCGGA, GCTCC and TCGGC) were associated with an increased risk of spermatogenesis and male infertility. In the subgroup analysis of NOA and severe oligozoospermia, similar positive effects were also found. Nevertheless, in the subgroup analysis of other types of infertility, we unfortunately failed to find a similar effect for several haplotypes, except for GCTGC and TCGGA. The potential mechanisms underlying these observations need to be investigated further.
Using gene–gene interaction analysis, we successfully examined 12 combined genotypes that were significantly associated with male infertility. The combined genotypes of CGGT (rs1646022/rs737008) and CGAC (rs1646022/rs2301365) were two risk factors for male infertility, accompanied by a 1.58–1.68-fold increased risk of infertility. We also found an intensely beneficial effect upon male infertility for several combined genotypes of rs737008 and rs1646022, such as GTCC, TTCC, GGCG, GTCG and GGGG. Similarly, a significant protective role for male infertility was identified for several combined genotypes of rs737008 and rs1646022 was found, such as AACC, ACCC, CCCG, and CCGG. Previous studies have shown that rs737008 and rs2301365 in PRM1, and rs1646022 in PRM2, were significantly associated with male infertility. This is because interference in the expression of PRM1/2 is essential for normal spermatogenesis. These results suggested that several SNPs in PRM1/2 might be an independent risk factor for male infertility.
Our study has several strengths and limitations. Based on a case-controlled study, we have obtained a rewarding result in that we identified an association between several SNPs and male infertility. However, there are also some limitations. Firstly, selection bias is unavoidable on account of the hospital-based case-controlled nature of this study. Secondly, with restriction to a China Han population, it is uncertain whether our findings could be generalized to other populations. We should therefore aim to analyze samples from several regions and ethnicities in future [29, 30]. Lastly, due to technological limitations, we could not verify the function of the selected SNPs, which may have helped us to understand the precise molecular mechanisms underlying the function of these selected SNPs and their influencing upon male infertility [31, 32].
In conclusion, we found that the PRM1 and PRM2 haplotypes GCTGC, TCGCA and TCGCC exhibited significant protective effects against male infertility as compared to fertile men, except for TCGGA, GCTCC and TCGGC which could represent a significant increased risk of spermatogenesis. Our study showed that rs737008 and rs2301365 in PRM1, and rs1646022 in PRM2, were significantly associated with male infertility and that gene–gene interactions played a role in male infertility. These findings will help us to further understand the aetiology of male infertility. Further studies should include a larger number of patients, genes, and SNPs, particularly growing critical genes; this will ultimately help us to unravel the effects of individual genetic factors upon the development of male infertility.
MATERIALS AND METHODS
Ethics statement
All patients provided written informed consent for the collection of samples and their subsequent analysis. This study was conducted in accordance with the tenets of the Declaration of Helsinki and its amendments, and was approved by the ethics committee of Jinling Hospital, Nanjing University.
Study population
This case-controlled study recruited 693 infertile men for a genetic questionnaire. All patients attended the Reproductive Medicine Centre of Jinling Hospital of Nanjing University between July 2013 and January 2015. At least two semen samples were analyzed for each patient to confirm the diagnosis of NOA or severe oligozoospermia. NOA was defined as a zero sperm count after semen centrifugation at 3000g for 10 min while severe oligozoospermia was defined as the detection limit of sperm concentration of < 2 × 106/ml and when the proportion of progressive sperm was < 32%.
All patient diagnoses were based upon a comprehensive andrological examination including medical history, physical examination, semen analysis, hormone analysis, karyotype analysis and Y chromosome microdeletion screening. Patients with a history of varicocele, epididymitis, orchitis, hypogonadotrophic hypogonadism, mono- or bilateral cryptorchidism, obstruction/absence of the vas deferens, chromosomal abnormalities, and Y chromosome microdeletions, were carefully excluded. Among the 693 infertile men, ten cases of chromosomal abnormalities, and 47 cases of Y chromosome microdeletions, were excluded from the final genotype analysis. Thus, 636 study cases were finally investigated (544 cases with NOA and severe oligozoospermia and 92 cases with other types of infertility).
The control group consisted of 442 hospital-based normozoospermia men aged from 24 to 45 years, who were recruited from the same reproductive medicine center, where they were in search of assisted reproduction technology because of female infertility. All controls had fathered at least one child previously. From each patient, we collected a range of clinical characteristics, including age, semen quantity, sperm counts, hormone levels, and PR value.
Genetic analyses
A panel of six SNPs of PRM1, PRM2 and TNP1 were selected for study, based on previous investigations. Genotyping analysis of the SNPs selected for fast-track validation analysis was performed using Sequenom MassARRAY technology (San Diego, CA, USA). Genomic DNA (15 ng) was used to genotype each sample. Locus-specific PCR and detection primers were designed using MassARRAY Assay Design 3.0 software at the Department of Reproduction and Genetics at the affiliated Jinling Hospital of Nanjing University. DNA samples were amplified by multiplex PCRs and the amplification products were then used for locus-specific single-base extension reactions. The resulting products were desalted and transferred to a 384-elementSpectroCHIP array (Sequenom). Allele detection was performed using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (Sequenom). Mass spectrograms were analysed by MassARRAY Typer software. SNPs detected with a call rate lower than 90% in the cases and controls, or beyond the Hardy–Weinberg equilibrium in the controls (P < 0.05), were excluded.
Statistical analysis
Differences in the distributions of demographic characteristics, selected variables, and frequencies of genotypes between cases and controls were tested by the Student’s t test (for continuous variables) or chi-square-test (for categorical variables). Hardy–Weinberg equilibrium was determined based upon the control genotyping results. A logistic regression analysis was used to analyze genotype distributions in the cases and controls. Linkage disequilibria and haplotypes were analysed with SHEsis software (http://analysis.bio-x.cn/myAnalysis.php; Shi and He, 2005). Frequencies of < 0.03 were ignored in the linkage disequilibrium analysis. The association between polymorphisms and the risk of infertility was analyzed by calculating the odds ratios (OR) and 95% confidence intervals (CI) and by comparing allele frequency or genotype of the case group with that of the control group. Strict Bonferroni correction was used to correct the P-values of the allele frequency and genotype for each SNP. Multiplicative interactions were assessed by logistic regression or Cox regression. The corrected significance level was interpreted as 0.0125 after Bonferroni correction for multiple comparisons. All tests were two-sided and carried out with SPSS software (version 22; SPSS Institute). P < 0.05 was considered statistically significant.
Abbreviations
PRM: Protamine; TNP1: Transition protein; SNP: Single nucleotide polymorphisms; NOA: Non-obstructive azoospermia; LD: Linkage disequilibrium plot; OR: odds ratios; 95%CI: 95% confidence intervals; T: Testosterone; E2: Estradiol; LH: Lutenizing hormone; FSH: Follicle-stimulating hormone; SO: Severe oligozoospermia.
Author contributions
W.J. J., Y.X. C., Y. L. and X.Y. X. conceived and designed the experiments. W.J. J., J. C. and P.R. Z. performed the experiments. W.J. J., M.X. N. and S.M. L. analyzed the data. W.J. J., W.W. L., M.M. Y., and Q.Y. W. contributed materials/analysis tools. W.J. J. wrote and revised the paper. The manuscript was reviewed and approved by all authors.
ACKNOWLEDGMENTS
We additionally appreciate the valuable comments from other members of our laboratory.
CONFLICTS OF INTEREST
None of the authors report any competing financial interest.
FUNDING
This work was supported by the Medical Health and Scientific Research Foundation of Jinling Hospital of Nanjing University (No. 2014MS098), the Key Foundation of Jiangsu Science and Technology Bureau (No. BM2015020), the Nanjing Science and Technology Development Project (No. 201503010) and the Foundation of Jinling Hospital of Nanjing University (No. 2014044 and No. 2015046).
REFERENCES
1. Krausz C, Escamilla AR, Chianese C. Genetics of male infertility: from research to clinic. Reproduction. 2015; 150:R159-R174.
2. Jiang W, Sun H, Zhang J, Zhou Q, Wu Q, Li T, Zhang C, Li W, Zhang M, Xia X. Polymorphisms in Protamine 1 and Protamine 2 predict the risk of male infertility: a meta-analysis. Sci Rep. 2015; 5:15300.
3. Agarwal A, Mulgund A, Hamada A, Chyatte MR. A unique view on male infertility around the globe. Reprod Biol Endocrinol. 2015; 13:37-46.
4. Winters BR, Walsh TJ. The epidemiology of male infertility. Urol Clin North Am. 2014; 41:195-204.
5. Mascarenhas MN, Flaxman SR, Boerma T, Vanderpoel S, Stevens GA. National, regional, and global trends in infertility prevalence since 1990: a systematic analysis of 277 health surveys. PLoS Med. 2012; 9:e1001356.
6. Castillo J, Estanyol JM, Ballesca JL, Oliva R. Human sperm chromatin epigenetic potential: genomics, proteomics, and male infertility. Asian J Androl. 2015; 17:601-609.
7. Oliva R. Protamines and male infertility. Hum Reprod Update. 2006; 12:417-435.
8. He XJ, Ruan J, Du WD, Chen G, Zhou Y, Xu S, Zuo XB, Cao YX, Zhang XJ. PRM1 variant rs35576928 (Arg> Ser) is associated with defective spermatogenesis in the Chinese Han population. Reprod Biomed Online. 2012; 25:627-634.
9. Jodar M, Oriola J, Mestre G, Castillo J, Giwercman A, Vidal-Taboada J, Ballescà J, Oliva R. Polymorphisms, haplotypes and mutations in the protamine 1 and 2 genes. Int J Androl. 2011; 34:470-485.
10. Tanaka H, Miyagawa Y, Tsujimura A, Matsumiya K, Okuyama A, Nishimune Y. Single nucleotide polymorphisms in the protamine-1 and-2 genes of fertile and infertile human male populations. Mol Hum Reprod. 2003; 9:69-73.
11. Yu Q, Yang X, Li F, Ye L, Wu Y, Mao X. [Association of PRM1-190C-> A polymorphism with teratozoospermia]. [Article in Chinese]. Zhonghua Nan Ke Xue. 2012; 18:314-317.
12. Tüttelmann F, Křenková P, Römer S, Nestorovic A, Ljujic M, Štambergová A, Macek M Jr, Macek M Sr, Nieschlag E, Gromoll J, Simoni M. A common haplotype of protamine 1 and 2 genes is associated with higher sperm counts. Int J Androl. 2010; 33:e240-e248.
13. Ravel C, Chantot-Bastaraud S, El Houate B, Berthaut I, Verstraete L, De Larouziere V, Lourenco D, Dumaine A, Antoine JM, Mandelbaum J, Siffroi JP, McElreavey K. Mutations in the protamine 1 gene associated with male infertility. Mol Hum Reprod. 2007; 13:461-464.
14. Imken L, Rouba H, El Houate B, Louanjli N, Barakat A, Chafik A, McElreavey K. Mutations in the protamine locus: association with spermatogenic failure? Mol Hum Reprod. 2009; 15:733-738.
15. Gázquez C, Oriola J, Mateo S, Vidal-Taboada JM, Ballescà JL, Oliva R. A common protamine 1 promoter polymorphism (-190 C-> A) correlates with abnormal sperm morphology and increased protamine P1/P2 ratio in infertile patients. J Androl. 2008; 29:540-548.
16. Aoki VW, Christensen GL, Atkins JF, Carrell DT. Identification of novel polymorphisms in the nuclear protein genes and their relationship with human sperm protamine deficiency and severe male infertility. Fertil Steril. 2006; 86:1416-1422.
17. Leitch HG, Tang WW, Surani MA. Primordial germ-cell development and epigenetic reprogramming in mammals. Curr Top Dev Biol. 2013; 104:149-187.
18. Francis S, Yelumalai S, Jones C, Coward K. Aberrant protamine content in sperm and consequential implications for infertility treatment. Hum Fertil (Camb). 2014; 17:80-89.
19. Bao JQ, Bedford MT. Epigenetic regulation of the histone-to-protamine transition during spermiogenesis. Reproduction. 2016; 151:R55-R70.
20. Miller D, Brinkworth M, Iles D. Paternal DNA packaging in spermatozoa: more than the sum of its parts? DNA, histones, protamines and epigenetics. Reproduction. 2010; 139:287-301.
21. Brunner AM, Nanni P, Mansuy IM. Epigenetic marking of sperm by post-translational modification of histones and protamines. Epigenetics Chromatin. 2014; 7:2.
22. Dada R, Kumar M, Jesudasan R, Fernandez JL, Gosalvez J, Agarwal A. Epigenetics and its role in male infertility. J Assist Reprod Genet. 2012; 29:213-223.
23. Grassetti D, Paoli D, Gallo M, D’Ambrosio A, Lombardo F, Lenzi A, Gandini L. Protamine-1 and-2 polymorphisms and gene expression in male infertility: an Italian study. J Endocrinol Invest. 2012; 35:882-888.
24. Venkatesh S, Kumar R, Deka D, Deecaraman M, Dada R. Analysis of sperm nuclear protein gene polymorphisms and DNA integrity in infertile men. Syst Biol Reprod Med. 2011; 57:124-132.
25. Kichine E, Msaidie S, Bokilo AD, Ducourneau A, Navarro A, Levy N, Terriou P, Collignon P, Boetsch G, Chiaroni J. Low-frequency protamine 1 gene transversions c.102G->T and c.-107G->C do not correlate with male infertility. J Med Genet. 2008; 45:255-256.
26. Yang XM, Liu F, Li J, Song YM, Tan YZ. Association study of protamine 2 (PRM2) gene polymorphism with male infertility in Chinese Han population. Genes Genomics. 2016; 38:311-317.
27. Godmann M, Lambrot R, Kimmins S. The dynamic epigenetic program in male germ cells: its role in spermatogenesis, testis cancer, and its response to the environment. Microsc Res Tech. 2009; 72:603-619.
28. O'Flynn O'Brien KL, Varghese AC, Agarwal A. The genetic causes of male factor infertility: a review. Fertil Steril. 2010; 93:1-12.
29. Qiu FM, Yang L, Lu XX, Chen JS, Wu D, Wei YF, Nong QQ, Zhang LS, Fang WX, Chen XL, Ling XX, Yang BY, Zhang X, et al. The MKK7 p.Glu116Lys rare variant serves as a predictor for lung cancer risk and prognosis in Chinese. PLoS Genet. 2016; 12: e1005955.
30. Zhang Z, Xu X, Ni H. Small studies may overestimate the effect sizes in critical care meta-analyses: a meta-epidemiological study. Crit Care. 2013; 17:R2.
31. He J, Shi TY, Zhu ML, Wang MY, Li QX, Wei QY. Associations of Lys939Gln and Ala499Val polymorphisms of the XPC gene with cancer susceptibility: a meta-analysis. Int J Cancer. 2013; 133:1765-1775.
32. Qin Y, Du G, Chen M, Hu W, Lu C, Wu W, Hang B, Zhou Z, Wang X, Xia Y. Combined effects of urinary phytoestrogens metabolites and polymorphisms in metabolic enzyme gene on idiopathic male infertility. Arch Toxicol. 2014; 88:1527-1536.