

INTRODUCTION
Prostate cancer is a common solid malignancy and has high mortality [1]. Exosomes are small extracellular vesicles (EV) ranging from 50 to 150 nm in diameter. Exosomes have a double membrane structure with various cargo contents, such as miRNAs, mRNAs, proteins, lipids and viral particles [2]. Exosomes are released by the exocytosis of multivesicular bodies (MVBs) (Figure 1) [3]. The materials in vesicles can be transferred and alter signaling pathways in the recipient cells [4]. Exosomes are present in human body fluids such as the blood, urine and saliva, and can be isolated from cell culture medium [5]. The lipid bilayer membrane of exosomes protects their cargo from RNases and proteases, which allows them to act as good delivery vector in therapy [6].
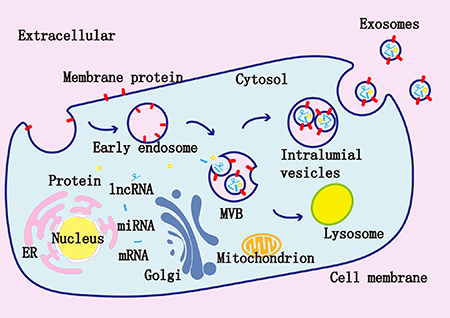
Figure 1: Exosomes are composed of a lipid bilayer and a variety of molecules derived from their original cells such as miRNAs, mRNAs, and proteins. Inside the cells, early endosomes are formed via endocytosis, early endosomes then develop to late endosomes, which form multivesicular bodies (MVBs) via the invagination of the membranes. The intraluminal vesicles (ILVs) are present in MVBs. Finally, MVBs fuse with the cell membrane and the ILVs will be released as exosomes.
Exosomes in prostate cancer progression
Tumor masses may arise from cancer stem cells which possess stem-like self-renewing ability [7]. Cancer stem cells were first found in leukemia, and later in other solid tumors including prostate cancer [8, 9]. Exosomes from cancer stem cells support prostate cancer tumorigenesis through promoting angiogenesis [10]. Recent studies suggest that exosomes from tumor microenvironment are important regulators to enhance prostate cells survival, proliferation, angiogenesis and the evasion of immune surveillance, which contribute to prostate cancer progression [10–12]. In particular, Soekmadji et al. discussed the potential of exosomes to provide candidate biomarkers for prostate cancer [13].
Tumor microenvironments are comprised of different types of cells, extracellular matrix, soluble factors, signaling molecules, and exosomes [14]. The cells include fibroblasts, inflammatory cells, lymphocytes, endothelial cells, epithelial cells, and mesenchymal stem cells. Soluble factors include growth factors, cytokines, and chemokines [15]. Carcinoma-associated fibroblasts (CAFs) known as myofibroblasts are induced and maintained by transforming growth factor-β (TGF-β) [15–19]. Prostate cancer cells derived exosomes can present TGF-β to transform fibroblasts to myofibroblasts via the activation of TGF-β/SMAD3 signaling [20, 21]. MiR-155 secreted from cancer derived exosomes can repress the expression of its target tumor protein 53-induced nuclear protein 1 (TP53INP1) to dictate CAF-like phenotypes in fibroblasts [22]. CAFs derived exosomes can transfer the miRNAs into neighboring epithelia causing the explosive growth of prostate cancer cells [23, 24]. CD81, miR-21 and miR-409 in CAFs derived exosomes affect invasion, proliferation, chemoresistance, and metabolism of cancer cells [25]. miR-21 could repress the expression of its targets apoptotic peptidase activating factor 1 (APAF1) and programmed cell death 4 (PDCD4) to inhibit the apoptosis and confer chemoresistance of cancer cells [26].
Cancer cells derived exosomes are also involved in the regulation of signaling pathways. C-Src, insulin-like growth factor I receptor and focal adhesion kinase are enriched in exosomes [27]. Androgen receptor (AR) can mediate the transcription of genes involved in prostate cancer cell proliferation and survival [28]. CD9 is an upstream regulator of AR, and exosomes can deliver CD9 to modulate paracrine signaling to mediate the growth of androgen deprived prostate cancer [29].
Angiogenesis plays a key role in the development of prostate cancer [30]. Cancer derived exosomes can induce angiogenesis. For instance, the exosomes of prostate and ovarian cancer cells transfer sphingomyelin and CD147 into endothelial cells to support the vascularization [25]. Exosomes also regulate immunity. Lundholm et al. found that exosomes of prostate cancer impaired cytotoxic function of lymphocytes, and decreased NKG2D receptor expression on natural killer cells and CD8+ T cells to promote tumor evasion from immune surveillance [31]. Other immunoregulatory molecules in cancer-derived exosomes such as FasL, TGF-β, galectin-9 and HSP72 support the immune escape of cancer cells [25]. In addition, exosomes from cancer cells activate Fas/FasL pathway to induce the apoptosis of CD8+ T cells [32]. Therefore, exosomes from both cancer cells and tumor microenvironment cooperate to promote prostate cancer progression.
Exosomes in prostate cancer metastasis
Most deaths of advanced prostate cancer patients are due to the metastasis of prostate cancer. Exosomes derived from tumors can be taken by the cells of specific organs and assist the formation of the pre-metastatic niche. Prostate cancer has metastatic organotropism of the bone [33]. Normal human cells can express prostate-specific genes after culturing with exosomes derived from prostate cancer tissues [34]. Exosomes from metastatic prostate cancer patients showed high contents of miR-21 and miR-141, which regulated osteoclastogenesis and osteoblastogenesis [35, 36]. Prostate cancer derived exosomes contained TGF-β which induced the conversion from bone marrow mesenchymal stem cells to fibroblasts [37]. Exosomes can prepare pre-metastatic niche. For example, exosomal miR-21, miR-375 and miR-141 help cancer cells overcome the low-androgen conditions in distant metastatic organs [10].
In addition, prostate cancer derived exosomes carried integrin α3 and integrin β1 which promoted the migration and invasion of epithelial cells [38]. The integrin αvβ6 was transferred by exosomes and its expression was high in prostate cancer. The recipient cells will internalize integrin αvβ6 and express them on the surface [39]. Integrin αvβ3 is highly expressed in many types of tumor and promotes the metastatic phenotype. In prostate cancer cells, integrin αvβ3 was co-expressed with synaptophysin which was considered a biomarker for aggressive neuroendocrine prostate cancer [40]. These exosomal integrins will activate Src phosphorylation and increase the expression of pro-inflammatory S100 in recipient cells, and have the potential to predict organ-specific metastasis [41].
The epithelial-mesenchymal transition (EMT) plays a pivotal role in the conversion from benign to malignant cancers. Cancer derived exosomes can promote EMT via miRNAs and prepare the pre-metastatic niche [42]. Several signaling pathways such as TGF-β1, Wnt, EGF and HGF participate in the induction of EMT [43–46]. The exosomes from human breast milk could promote EMT via TGFβ2 [20]. miR-409 in exosomes from prostate cancer promoted EMT through the repression of tumor suppressor genes such as Ras suppressor 1 and stromal antigen 2 [23].
Metastasis is a highly inefficient process. Only 0.01% circulating tumor cells (CTCs) shed from the primary tumors into the bloodstream and lymphatics can form metastatic lesions in distant organs [47]. EMT markers such as twist and vimentin were expressed at higher levels in CTCs of patients with metastatic breast cancer than in those of patients in the early stage [48]. Metastases-initiating cells (MICs) are special CTCs with sternness and enhance the growth, survival and colonization of prostate cancer cells in distant metastatic organs [11]. MICs have the ability to alter tumor microenvironment to promote reprogramming of non-tumorigenic prostate cancerous and non-cancerous epithelial and stromal cells, leading to their transformation and de-differentiation [49, 50]. Exosomes derived from MICs can promote EMT of prostate cancer cells through the activation of RANKL, FOXM1, and c-Myc [11].
Exosomes in prostate cancer drug resistance
Exosomes contribute to chemoresistance of cancer cells by complicated mechanisms. In cancer cells, chemotherapeutic drugs could be exported via exosomes [51]. Exosomes can shield cancer cells from therapeutic antibody attack, leading to the failure of antibody therapy [52]. Exosomal contents play an important role in the drug resistance of prostate cancer cells. For example, miR-34 in prostate cancer cells and cell-derived exosomes targeted Bcl-2 to regulate the response to docetaxel [53]. Exosomes could confer docetaxel-resistant cancer cells to docetaxel-sensitive cancer cells [54]. A recent study identified 29 deregulated miRNAs in exosomes from paclitaxel resistant prostate cancer cells, and these exosome-derived miRNAs may contribute to prostate cancer chemoresistance [55].
AR is a key transcription regulator that is highly expressed in prostate cancer. AR isoform encoded by splice variant 7 lacks the ligand-binding domain and is associated with the resistance to hormonal prostate cancer therapies, especially enzalutamide and abiraterone [56]. Androgen-receptor splice variant 7 messenger RNA (AR-V7) can be isolated from exosomal RNA in the blood and is a valuable resistance marker [56].
Exosomes for the diagnosis of prostate cancer
Present diagnostic markers such as prostate specific antigen (PSA) and carbohydrate antigens have substantial drawbacks such as false-negatives, false-positives and lack of tumor-type specificity [57]. Tumor biopsy is the only definitive method of diagnosis, but it is invasive. Novel prostate cancer biomarkers are required for clinical application. Exosomes can be isolated from human body fluids such as the blood, urine and saliva [58].
Exosomes can protect miRNAs against RNase degradation [59]. Huang et al. found that miR-1290 and miR-375 had the potential of predicting the prognosis of castration-resistant prostate cancer [60]. Exosomal miR-34a could induce docetaxel sensitivity in docetaxel-resistant prostate cancer cells by inhibiting Bcl-2 [30]. Exosomal miR-34a can be used as a predictive biomarker for the response to docetaxel [53]. A recent study showed that miR-182 of miR-183 cluster family was detected in prostate cancer cells derived exosomes from the serum [61].
Like the miRNAs, the proteins in exosomes can be the biomarkers for prostate cancer. Hosseini-Beheshti et al. characterized exosomal proteins from prostate cancer cells and identified annexin A2, calsyntenin 1, fatty acid synthesis, filamin C, folate hydrolase-1, and growth differentiation factor 15, which may be specific for prostate cancer diagnosis [62]. Duijvesz et al. identified biomarker exportin-1 [63]. Webber et al. found that Notch3, milk fat globule epidermal growth factor-factor 8, and inter-alpha-trypsin inhibitor heavy chain H4 were enriched in prostate cancer exosomes [64]. Khan et al. reported that exosomal survivin was a potential biomarker for early detection of prostate cancer [65]. In addition, prostate cancer antigen 3 (PCA3), flotillin 2, Rab3B and late endosomal/lysosomal adaptor, MAPK and mTOR activator 1 (LAMTOR1) of exosomes could be diagnostic markers for prostate cancer [66, 67]. Exosomal lncRNAs also have the potential to be the biomarkers of prostate cancer. Exosomal lncRNAs may be involved in prostate cancer carcinogenesis and can be utilized for prostate cancer diagnosis [68].
Noninvasive and simple diagnostic assays are required for prostate cancer diagnosis. A novel noninvasive Urine Exosome Gene Expression Assay has been applied to reduce the number of unnecessary biopsies [69]. Moreover, a PCR-free efficient diagnostic method was developed for simultaneous and multiplexed detection of exosomal miRNAs [70]. These improvements of detection technology facilitate the application of exosomes for prostate cancer diagnosis.
Exosomes in prostate cancer therapy
Exosomes can be used as a delivery vector to target cancer cells and the contents can escape the attack by immune system [71]. Adipose-derived stromal cells (ASCs) derived exosomal miR-145 could reduce the activity of Bcl-xL and promote prostate cancer cell apoptosis via caspase-3/7 pathway. Therefore, ASCs derived exosomes can be used in prostate cancer therapy [72]. Engineered microvesicles can carry suicide mRNA/protein to inhibit Schwannoma growth [73]. Saari et al. used exosomes as the carriers to deliver paclitaxel to autologous prostate cancer cells and showed increased cytotoxic effect [74]. Encapsulation of anti-inflammatory agent curcumin in exosomes achieved a high concentration of curcumin in target tissues [75].
Exosomes are also utilized in tumor vaccination. Tumor derived exosomes often contain tumor specific antigens to activate dendritic cells which induce anti-tumor response of T lymphocytes [76, 77]. Dendritic cells derived exosomes activate NK cells [78]. A recent study showed an efficient exosome-based tumor antigens-adjuvant co-delivery system. CpG DNA modified exosomes derived from tumor cells could deliver tumor antigens to antigen presenting cells efficiently and show promise in cancer immunotherapy [79].
A new tool was developed for intracellular delivery of target proteins which was named exosomes for protein loading via optically reversible protein–protein interactions (EXPLORs) [71]. Nanoscale exosome-mimics (EMs) could be designed to produce sufficient quantity of vectors used for drug or gene delivery in cancer therapy [80]. A recent study showed that exosomes engineered as doxorubicin delivery platform for targeted therapy achieved high therapy efficiency [81]. However, all these studies are in the experimental stage. Further preclinical studies are needed to validate the potential of exosomes in prostate cancer therapy.
Perspective
Tumor derived exosomes orchestrate a series of processes, such as coagulation, vascular leakiness, and reprogramming of stromal recipient cells to provide pre-metastatic niche and promote subsequent metastasis [82]. In addition, exosomes released by prostate cancer cells in tumor-bone interface promote osteoclast fusion and differentiation to support the metastasis of prostate cancer to the bone [83]. In summary, accumulating evidences confirm that exosomes are implicated in the progression and metastasis of prostate cancer (Figure 2). Many biological molecules are encapsulated in the exosomes from prostate cancer such as miRNAs, lncRNAs and proteins, and their expression levels differ from those of normal prostate cells. The easy isolation of exosomes from body fluid enables them as potential biomarkers of prostate cancer [84, 85]. Furthermore, the lipid bilayer membrane of exosomes makes them promising carriers of drugs and other therapeutic molecules to target prostate cancer. In the near future, we would expect that the power of this nano-sized vesicles would be realized to promote the clinical application of exosomes in prostate cancer diagnosis and therapy.
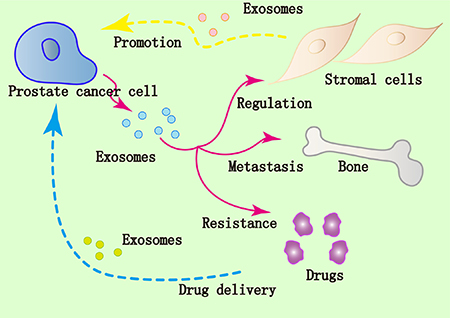
Figure 2: Implication of exosomes in prostate cancer. Prostate cancer cells modulate surrounding stromal cells via the exosomes. Affected stromal cells employ the exosomes to modulate microenvironment which can promote tumor growth and metastasis. Exosomes derived from prostate cancer cells could contribute to drug resistance of cancer. The lipid bilayer membrane of exosomes makes them promising carriers of drugs and other therapeutic molecules targeting prostate cancer.
CONFLICTS OF INTEREST
None.
FUNDING
This study was supported by grants from Jiangsu Province Science Foundation of China (No BK2007032), the Project of Invigorating Health Care through Science, Technology and Education and Jiangsu Provincial Medical Youth Talent.
REFERENCES
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016; 66:7–30.
2. Mathivanan S, Fahner CJ, Reid GE, Simpson RJ. ExoCarta 2012: database of exosomal proteins, RNA and lipids. Nucleic Acids Res. 2012; 40:D1241–44.
3. Fujita Y, Yoshioka Y, Ochiya T. Extracellular vesicle transfer of cancer pathogenic components. Cancer Sci. 2016; 107:385–90.
4. Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007; 9:654–59.
5. Lopez-Verrilli MA, Court FA. Exosomes: mediators of communication in eukaryotes. Biol Res. 2013; 46:5–11.
6. Shtam TA, Kovalev RA, Varfolomeeva EY, Makarov EM, Kil YV, Filatov MV. Exosomes are natural carriers of exogenous siRNA to human cells in vitro. Cell Commun Signal. 2013; 11:88.
7. Al-Hajj M. Cancer stem cells and oncology therapeutics. Curr Opin Oncol. 2007; 19:61–64.
8. Bonnet D, Dick JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med. 1997; 3:730–37.
9. Collins AT, Berry PA, Hyde C, Stower MJ, Maitland NJ. Prospective identification of tumorigenic prostate cancer stem cells. Cancer Res. 2005; 65:10946–51.
10. Liu CM, Hsieh CL, Shen CN, Lin CC, Shigemura K, Sung SY. Exosomes from the tumor microenvironment as reciprocal regulators that enhance prostate cancer progression. Int J Urol. 2016; 23:734–44.
11. Shiao SL, Chu GC, Chung LW. Regulation of prostate cancer progression by the tumor microenvironment. Cancer Lett. 2016; 380:340–48.
12. De Marzo AM, Platz EA, Sutcliffe S, Xu J, Grönberg H, Drake CG, Nakai Y, Isaacs WB, Nelson WG. Inflammation in prostate carcinogenesis. Nat Rev Cancer. 2007; 7:256–69.
13. Soekmadji C, Russell PJ, Nelson CC. Exosomes in prostate cancer: putting together the pieces of a puzzle. Cancers (Basel). 2013; 5:1522–44.
14. Egeblad M, Nakasone ES, Werb Z. Tumors as organs: complex tissues that interface with the entire organism. Dev Cell. 2010; 18:884–901.
15. Wiseman BS, Werb Z. Stromal effects on mammary gland development and breast cancer. Science. 2002; 296:1046–49.
16. Calvo F, Ege N, Grande-Garcia A, Hooper S, Jenkins RP, Chaudhry SI, Harrington K, Williamson P, Moeendarbary E, Charras G, Sahai E. Mechanotransduction and YAP-dependent matrix remodelling is required for the generation and maintenance of cancer-associated fibroblasts. Nat Cell Biol. 2013; 15:637–46.
17. Shimoda M, Principe S, Jackson HW, Luga V, Fang H, Molyneux SD, Shao YW, Aiken A, Waterhouse PD, Karamboulas C, Hess FM, Ohtsuka T, Okada Y, et al. Loss of the Timp gene family is sufficient for the acquisition of the CAF-like cell state. Nat Cell Biol. 2014; 16:889–901.
18. Procopio MG, Laszlo C, Labban DA, Dong EK, Bordignon P, Jo SH, Goruppi S, Menietti E, Ostano P, Ala U, Provero P, Hoetzenecker W, Neel V, et al. Combined CSL and p53 downregulation promotes cancer-associated fibroblast activation. Nat Cell Biol. 2015; 17:1193–204.
19. Albrengues J, Bertero T, Grasset E, Bonan S, Maiel M, Bourget I, Philippe C, Herraiz Serrano C, Benamar S, Croce O, Sanz-Moreno V, Meneguzzi G, Feral CC, et al. Epigenetic switch drives the conversion of fibroblasts into proinvasive cancer-associated fibroblasts. Nat Commun. 2015; 6:10204.
20. Webber J, Steadman R, Mason MD, Tabi Z, Clayton A. Cancer exosomes trigger fibroblast to myofibroblast differentiation. Cancer Res. 2010; 70:9621–30.
21. Webber JP, Spary LK, Sanders AJ, Chowdhury R, Jiang WG, Steadman R, Wymant J, Jones AT, Kynaston H, Mason MD, Tabi Z, Clayton A. Differentiation of tumour-promoting stromal myofibroblasts by cancer exosomes. Oncogene. 2015; 34:290–302.
22. Pang W, Su J, Wang Y, Feng H, Dai X, Yuan Y, Chen X, Yao W. Pancreatic cancer-secreted miR-155 implicates in the conversion from normal fibroblasts to cancer-associated fibroblasts. Cancer Sci. 2015; 106:1362–69.
23. Josson S, Gururajan M, Sung SY, Hu P, Shao C, Zhau HE, Liu C, Lichterman J, Duan P, Li Q, Rogatko A, Posadas EM, Haga CL, Chung LW. Stromal fibroblast-derived miR-409 promotes epithelial-to-mesenchymal transition and prostate tumorigenesis. Oncogene. 2015; 34:2690–99.
24. Josson S, Gururajan M, Hu P, Shao C, Chu GY, Zhau HE, Liu C, Lao K, Lu CL, Lu YT, Lichterman J, Nandana S, Li Q, et al. miR-409-3p/-5p promotes tumorigenesis, epithelial-to-mesenchymal transition, and bone metastasis of human prostate cancer. Clin Cancer Res. 2014; 20:4636–46.
25. Naito Y, Yoshioka Y, Yamamoto Y, Ochiya T. How cancer cells dictate their microenvironment: present roles of extracellular vesicles. Cell Mol Life Sci. 2017; 74:697–713.
26. Au Yeung CL, Co NN, Tsuruga T, Yeung TL, Kwan SY, Leung CS, Li Y, Lu ES, Kwan K, Wong KK, Schmandt R, Lu KH, Mok SC. Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat Commun. 2016; 7:11150.
27. DeRita RM, Zerlanko B, Singh A, Lu H, Iozzo RV, Benovic JL, Languino LR. c-Src, Insulin-Like Growth Factor I Receptor, G-Protein-Coupled Receptor Kinases and Focal Adhesion Kinase are Enriched Into Prostate Cancer Cell Exosomes. J Cell Biochem. 2017; 118:66–73.
28. Bentel JM, Tilley WD. Androgen receptors in prostate cancer. J Endocrinol. 1996; 151:1–11.
29. Soekmadji C, Riches JD, Russell PJ, Ruelcke JE, McPherson S, Wang C, Hovens CM, Corcoran NM, Hill MM, Nelson CC, and Australian Prostate Cancer Collaboration BioResource. Modulation of paracrine signaling by CD9 positive small extracellular vesicles mediates cellular growth of androgen deprived prostate cancer. Oncotarget. 2016; 8:52237–55. https://doi.org/10.18632/oncotarget.11111.
30. Carmeliet P, Jain RK. Angiogenesis in cancer and other diseases. Nature. 2000; 407:249–57.
31. Lundholm M, Schröder M, Nagaeva O, Baranov V, Widmark A, Mincheva-Nilsson L, Wikström P. Prostate tumor-derived exosomes down-regulate NKG2D expression on natural killer cells and CD8+ T cells: mechanism of immune evasion. PLoS One. 2014; 9:e108925.
32. Steinbichler TB, Dudas J, Riechelmann H, Skvortsova II. The Role of Exosomes in Cancer Metastasis. Semin Cancer Biol. 2017. pii: S1044-579X(17)30024-X. https://doi.org/10.1016/j.semcancer.2017.02.006.
33. Roudier MP, True LD, Higano CS, Vesselle H, Ellis W, Lange P, Vessella RL. Phenotypic heterogeneity of end-stage prostate carcinoma metastatic to bone. Hum Pathol. 2003; 34:646–53.
34. Renzulli JF 2nd, Del Tatto M, Dooner G, Aliotta J, Goldstein L, Dooner M, Colvin G, Chatterjee D, Quesenberry P. Microvesicle induction of prostate specific gene expression in normal human bone marrow cells. J Urol. 2010; 184:2165–71.
35. Sugatani T, Vacher J, Hruska KA. A microRNA expression signature of osteoclastogenesis. Blood. 2011; 117:3648–57.
36. Zhang HL, Qin XJ, Cao DL, Zhu Y, Yao XD, Zhang SL, Dai B, Ye DW. An elevated serum miR-141 level in patients with bone-metastatic prostate cancer is correlated with more bone lesions. Asian J Androl. 2013; 15:231–35.
37. Chowdhury R, Webber JP, Gurney M, Mason MD, Tabi Z, Clayton A. Cancer exosomes trigger mesenchymal stem cell differentiation into pro-angiogenic and pro-invasive myofibroblasts. Oncotarget. 2015; 6:715–31. https://doi.org/10.18632/oncotarget.2711.
38. Bijnsdorp IV, Geldof AA, Lavaei M, Piersma SR, van Moorselaar RJ, Jimenez CR. Exosomal ITGA3 interferes with non-cancerous prostate cell functions and is increased in urine exosomes of metastatic prostate cancer patients. J Extracell Vesicles. 2013; 2:1–10.
39. Fedele C, Singh A, Zerlanko BJ, Iozzo RV, Languino LR. The αvβ6 integrin is transferred intercellularly via exosomes. J Biol Chem. 2015; 290:4545–51.
40. Singh A, Fedele C, Lu H, Nevalainen MT, Keen JH, Languino LR. Exosome-mediated Transfer of αvβ3 Integrin from Tumorigenic to Nontumorigenic Cells Promotes a Migratory Phenotype. Mol Cancer Res. 2016; 14:1136–46.
41. Hoshino A, Costa-Silva B, Shen TL, Rodrigues G, Hashimoto A, Tesic Mark M, Molina H, Kohsaka S, Di Giannatale A, Ceder S, Singh S, Williams C, Soplop N, et al. Tumour exosome integrins determine organotropic metastasis. Nature. 2015; 527:329–35.
42. Banyard J, Bielenberg DR. The role of EMT and MET in cancer dissemination. Connect Tissue Res. 2015; 56:403–13.
43. Gou WF, Zhao Y, Lu H, Yang XF, Xiu YL, Zhao S, Liu JM, Zhu ZT, Sun HZ, Liu YP, Xu F, Takano Y, Zheng HC. The role of RhoC in epithelial-to-mesenchymal transition of ovarian carcinoma cells. BMC Cancer. 2014; 14:477.
44. Geng SQ, Alexandrou AT, Li JJ. Breast cancer stem cells: multiple capacities in tumor metastasis. Cancer Lett. 2014; 349:1–7.
45. Shimoda M, Khokha R. Proteolytic factors in exosomes. Proteomics. 2013; 13:1624–36.
46. Tauro BJ, Mathias RA, Greening DW, Gopal SK, Ji H, Kapp EA, Coleman BM, Hill AF, Kusebauch U, Hallows JL, Shteynberg D, Moritz RL, Zhu HJ, Simpson RJ. Oncogenic H-ras reprograms Madin-Darby canine kidney (MDCK) cell-derived exosomal proteins following epithelial-mesenchymal transition. Mol Cell Proteomics. 2013; 12:2148–59.
47. Valastyan S, Weinberg RA. Tumor metastasis: molecular insights and evolving paradigms. Cell. 2011; 147:275–92.
48. Kallergi G, Papadaki MA, Politaki E, Mavroudis D, Georgoulias V, Agelaki S. Epithelial to mesenchymal transition markers expressed in circulating tumour cells of early and metastatic breast cancer patients. Breast Cancer Res. 2011; 13:R59.
49. Chu GC, Zhau HE, Wang R, Rogatko A, Feng X, Zayzafoon M, Liu Y, Farach-Carson MC, You S, Kim J, Freeman MR, Chung LW. RANK- and c-Met-mediated signal network promotes prostate cancer metastatic colonization. Endocr Relat Cancer. 2014; 21:311–26.
50. Li Q, Li Q, Nuccio J, Liu C, Duan P, Wang R, Jones LW, Chung LW, Zhau HE. Metastasis initiating cells in primary prostate cancer tissues from transurethral resection of the prostate (TURP) predicts castration-resistant progression and survival of prostate cancer patients. Prostate. 2015; 75:1312–21.
51. Shedden K, Xie XT, Chandaroy P, Chang YT, Rosania GR. Expulsion of small molecules in vesicles shed by cancer cells: association with gene expression and chemosensitivity profiles. Cancer Res. 2003; 63:4331–37.
52. Yu S, Cao H, Shen B, Feng J. Tumor-derived exosomes in cancer progression and treatment failure. Oncotarget. 2015; 6:37151–68. https://doi.org/10.18632/oncotarget.6022.
53. Corcoran C, Rani S, O’Driscoll L. miR-34a is an intracellular and exosomal predictive biomarker for response to docetaxel with clinical relevance to prostate cancer progression. Prostate. 2014; 74:1320–34.
54. Corcoran C, Rani S, O’Brien K, O’Neill A, Prencipe M, Sheikh R, Webb G, McDermott R, Watson W, Crown J, O’Driscoll L. Docetaxel-resistance in prostate cancer: evaluating associated phenotypic changes and potential for resistance transfer via exosomes. PLoS One. 2012; 7:e50999.
55. Li J, Yang X, Guan H, Mizokami A, Keller ET, Xu X, Liu X, Tan J, Hu L, Lu Y, Zhang J. Exosome-derived microRNAs contribute to prostate cancer chemoresistance. Int J Oncol. 2016; 49:838–46.
56. Taneja SS. Re: AR-V7 and resistance to enzalutamide and abiraterone in prostate cancer. J Urol. 2015; 193:538.
57. Perkins GL, Slater ED, Sanders GK, Prichard JG. Serum tumor markers. Am Fam Physician. 2003; 68:1075–82.
58. Keller S, Ridinger J, Rupp AK, Janssen JW, Altevogt P. Body fluid derived exosomes as a novel template for clinical diagnostics. J Transl Med. 2011; 9:86.
59. Koga Y, Yasunaga M, Moriya Y, Akasu T, Fujita S, Yamamoto S, Matsumura Y. Exosome can prevent RNase from degrading microRNA in feces. J Gastrointest Oncol. 2011; 2:215–22.
60. Huang X, Yuan T, Liang M, Du M, Xia S, Dittmar R, Wang D, See W, Costello BA, Quevedo F, Tan W, Nandy D, Bevan GH, et al. Exosomal miR-1290 and miR-375 as prognostic markers in castration-resistant prostate cancer. Eur Urol. 2015; 67:33–41.
61. Mihelich BL, Dambal S, Lin S, Nonn L. miR-182, of the miR-183 cluster family, is packaged in exosomes and is detected in human exosomes from serum, breast cells and prostate cells. Oncol Lett. 2016; 12:1197–203.
62. Hosseini-Beheshti E, Pham S, Adomat H, Li N, Tomlinson Guns ES. Exosomes as biomarker enriched microvesicles: characterization of exosomal proteins derived from a panel of prostate cell lines with distinct AR phenotypes. Mol Cell Proteomics. 2012; 11:863–85.
63. Duijvesz D, Burnum-Johnson KE, Gritsenko MA, Hoogland AM, Vredenbregt-van den Berg MS, Willemsen R, Luider T, Paša-Tolić L, Jenster G. Proteomic profiling of exosomes leads to the identification of novel biomarkers for prostate cancer. PLoS One. 2013; 8:e82589–82589.
64. Webber J, Stone TC, Katilius E, Smith BC, Gordon B, Mason MD, Tabi Z, Brewis IA, Clayton A. Proteomics analysis of cancer exosomes using a novel modified aptamer-based array (SOMAscan™) platform. Mol Cell Proteomics. 2014; 13:1050–64.
65. Khan S, Jutzy JM, Valenzuela MM, Turay D, Aspe JR, Ashok A, Mirshahidi S, Mercola D, Lilly MB, Wall NR. Plasma-derived exosomal survivin, a plausible biomarker for early detection of prostate cancer. PLoS One. 2012; 7:e46737–46737.
66. Nilsson J, Skog J, Nordstrand A, Baranov V, Mincheva-Nilsson L, Breakefield XO, Widmark A. Prostate cancer-derived urine exosomes: a novel approach to biomarkers for prostate cancer. Br J Cancer. 2009; 100:1603–07.
67. Wang L, Skotland T, Berge V, Sandvig K, Llorente A. Exosomal proteins as prostate cancer biomarkers in urine: from mass spectrometry discovery to immunoassay-based validation. Eur J Pharm Sci. 2017; 98:80–85.
68. Ahadi A, Khoury S, Losseva M, Tran N. A comparative analysis of lncRNAs in prostate cancer exosomes and their parental cell lines. Genom Data. 2016; 9:7–9.
69. McKiernan J, Donovan MJ, O’Neill V, Bentink S, Noerholm M, Belzer S, Skog J, Kattan MW, Partin A, Andriole G, Brown G, Wei JT, Thompson IM Jr, Carroll P. A Novel Urine Exosome Gene Expression Assay to Predict High-grade Prostate Cancer at Initial Biopsy. JAMA Oncol. 2016; 2:882–89.
70. Lee JH, Kim JA, Jeong S, Rhee WJ. Simultaneous and multiplexed detection of exosome microRNAs using molecular beacons. Biosens Bioelectron. 2016; 86:202–10.
71. Yim N, Ryu SW, Choi K, Lee KR, Lee S, Choi H, Kim J, Shaker MR, Sun W, Park JH, Kim D, Heo WD, Choi C. Exosome engineering for efficient intracellular delivery of soluble proteins using optically reversible protein-protein interaction module. Nat Commun. 2016; 7:12277.
72. Takahara K, Ii M, Inamoto T, Nakagawa T, Ibuki N, Yoshikawa Y, Tsujino T, Uchimoto T, Saito K, Takai T, Tanda N, Minami K, Uehara H, et al. microRNA-145 Mediates the Inhibitory Effect of Adipose Tissue-Derived Stromal Cells on Prostate Cancer. Stem Cells Dev. 2016; 25:1290–98.
73. Mizrak A, Bolukbasi MF, Ozdener GB, Brenner GJ, Madlener S, Erkan EP, Ströbel T, Breakefield XO, Saydam O. Genetically engineered microvesicles carrying suicide mRNA/protein inhibit schwannoma tumor growth. Mol Ther. 2013; 21:101–08.
74. Saari H, Lázaro-Ibáñez E, Viitala T, Vuorimaa-Laukkanen E, Siljander P, Yliperttula M. Microvesicle- and exosome-mediated drug delivery enhances the cytotoxicity of Paclitaxel in autologous prostate cancer cells. J Control Release. 2015; 220:727–37.
75. Sun D, Zhuang X, Xiang X, Liu Y, Zhang S, Liu C, Barnes S, Grizzle W, Miller D, Zhang HG. A novel nanoparticle drug delivery system: the anti-inflammatory activity of curcumin is enhanced when encapsulated in exosomes. Mol Ther. 2010; 18:1606–14.
76. Wolfers J, Lozier A, Raposo G, Regnault A, Théry C, Masurier C, Flament C, Pouzieux S, Faure F, Tursz T, Angevin E, Amigorena S, Zitvogel L. Tumor-derived exosomes are a source of shared tumor rejection antigens for CTL cross-priming. Nat Med. 2001; 7:297–303.
77. Andre F, Schartz NE, Movassagh M, Flament C, Pautier P, Morice P, Pomel C, Lhomme C, Escudier B, Le Chevalier T, Tursz T, Amigorena S, Raposo G, et al. Malignant effusions and immunogenic tumour-derived exosomes. Lancet. 2002; 360:295–305.
78. Viaud S, Théry C, Ploix S, Tursz T, Lapierre V, Lantz O, Zitvogel L, Chaput N. Dendritic cell-derived exosomes for cancer immunotherapy: what’s next? Cancer Res. 2010; 70:1281–85.
79. Morishita M, Takahashi Y, Matsumoto A, Nishikawa M, Takakura Y. Exosome-based tumor antigens-adjuvant co-delivery utilizing genetically engineered tumor cell-derived exosomes with immunostimulatory CpG DNA. Biomaterials. 2016; 111:55–65.
80. Yang Z, Xie J, Zhu J, Kang C, Chiang C, Wang X, Wang X, Kuang T, Chen F, Chen Z, Zhang A, Yu B, Lee RJ, et al. Functional exosome-mimic for delivery of siRNA to cancer: in vitro and in vivo evaluation. J Control Release. 2016; 243:160–71.
81. Tian Y, Li S, Song J, Ji T, Zhu M, Anderson GJ, Wei J, Nie G. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials. 2014; 35:2383–90.
82. Becker A, Thakur BK, Weiss JM, Kim HS, Peinado H, Lyden D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell. 2016; 30:836–48.
83. Karlsson T, Lundholm M, Widmark A, Persson E. Tumor Cell-Derived Exosomes from the Prostate Cancer Cell Line TRAMP-C1 Impair Osteoclast Formation and Differentiation. PLoS One. 2016; 11:e0166284.
84. Valentino A, Reclusa P, Sirera R, Giallombardo M, Camps C, Pauwels P, Crispi S, Rolfo C. Exosomal microRNAs in liquid biopsies: future biomarkers for prostate cancer. Clin Transl Oncol. 2017; 19:651–57. [Epub ahead of print].
85. Foj L, Ferrer F, Serra M, Arévalo A, Gavagnach M, Giménez N, Filella X. Exosomal and Non-Exosomal Urinary miRNAs in Prostate Cancer Detection and Prognosis. Prostate. 2017; 77:573–83.