
INTRODUCTION
Cancer includes a large group of malignancies that are believed to occur as a combined consequence of genetic and environmental factors, since many genetic traits have been demonstrated to suppress or promote carcinogenesis as cancer suppressors or oncogenes, and environmental factors may help prevent or promote cancers [1–3]. But individuals born to a family that has high genetic risks to cancers do not necessarily develop cancers in their lifetime. Conversely, many cancer patients do not have a high-risk familial genetic background nor contacts to known harmful environmental factors like tobacco, industrial pollutants, etc.
A recent study shows that the cancer risk is strongly correlated with the number of cell divisions [4]. Based on this finding, the authors explain that the majority of cancerous variation is due to random mutations arising during DNA replication in stem cells, which the authors call “bad luck”. Indeed, it is not hard to understand that the more cycles of DNA replication a cell undergoes, the more random errors it will accumulate and hence the more chances a malignant mutation appears, leading to cancer as a result. Therefore, it is a quite reasonable explanation to a fact recognized for over a century that some tissue types give rise to cancers much more frequently than other types due to their greater turnover frequencies. However, it does not explain why the majority of people in the general populations, especially those under various genetic or environmental risks to cancer, do not develop cancer for their eighty years or even longer lifetime in any tissue. This question has led us to believe that there must be so far an unknown kind of factors, non-genetic and non-environmental in the traditional meaning, in addition to immunity, that constantly work in the healthy human body to clear malignant cells timely to prevent them from forming cancers.
Previously, we investigated the intestinal microbiota for its potential associations with human health and isolated bacteria that produce the anti-cancer agent enterodiol or secoisolariciresinol [5–9] by converting precursors contained in plants such as defatted flaxseeds through biotransformation [10–13]. Results of these studies prompted us to postulate that the intestinal microbiota might also contain microbes that attack malignant cells more directly, e.g., using de novo chemical substances, and lack of such microbial malignancy-killers may make people more vulnerable to cancer-causing factors. In this study, we screened the intestinal microbiota of two representative populations and isolated bacterial strains that produced potent anti-cancer substances. The isolated bacterial malignancy-killers were diverse by phylogeny and active against broad ranges of cancer types in vitro and in vivo. This finding demonstrates the existence of another potent anti-cancer system in the human body in addition to the cancer suppression and immune systems and reiterates the importance to keep our intestinal microbiota undisturbed by avoiding unhealthy life styles such as unbalanced dietary structure or smoking and inappropriate medical procedures such as misuse of antimicrobials.
RESULTS
Detection of anti-malignancy activities in bulk fecal specimens
We first screened the fecal specimens collected from preschool children and young adults (See Materials and Methods) for their inhibiting effects on malignant cells. For this, we mixed the fresh fecal specimens with equal volume of sterile water before centrifugation and then added a 5 μl aliquot of the filtered fecal supernatant to 100 μl of cancer cell cultures. We inspected the effects under a microscope at 6 hours intervals. From many of the fecal specimens, we detected damaging effects on cancer cells, which led to various morphological changes such as cells exhibiting pyknosis, forming vacuoles and turning round (Figure 1).
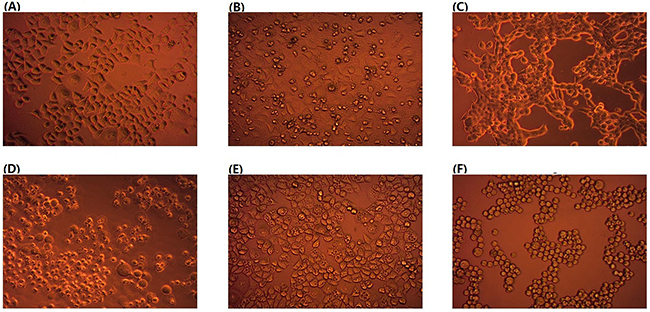
Figure 1: Effects of human fecal supernatant on HeLa cells. (A), control without fecal supernatant; (B) through (F), fecal supernatant from five different individuals (Magnification, 200 X).
Of remarkable significance, there was a strong tendency that preschool children had a much higher percentage of individuals exhibiting the anti-cancer effects than young adults (Table 1). Although the difference was statistically significant, we believed that this finding would need to be validated by larger scales of screening or more sensitive methods for the detection. Alternatively, detection of the producers of the anti-cancer substances would yield more direct evidence for the existence of the malignancy-killers and can also provide the materials for further studies or applications.
Table 1: Percentages of individuals whose fecal specimens exhibit positive anti-cancer activities in different age populations
Age group |
No. samples |
No. positive samples |
Percentage of positive samples |
|
|---|---|---|---|---|
3-7 years |
100 |
59 |
59.00% |
|
18-39 years |
113 |
12 |
10.62% |
**P<0.001 |
Total |
213 |
71 |
33.33% |
Note: The P value indicates comparison between different age groups with the chi-square test by SPSS.
Isolation of malignancy-killing bacterial strains
To isolate and identify the microbes that are responsible for the detected anti-malignancy activities, we spread diluted fecal specimens onto agar plates (see Materials and Methods), picked single colonies and grew the bacteria in liquid medium for up to 168 hours before inspecting their effects on cancer cells. From the collected 213 fecal specimens, we obtained 76 strains that showed various anti-malignancy effects. Supplementary Table 1 shows the detailed information about the participants and the anti-malignancy bacterial strains isolated from them. Notably, we isolated cancer-killing bacteria not only from fecal specimens tested positive but also from those tested negative in the supernatant for anti-malignancy activities, demonstrating that the final expression of the cancer-killing effects of these bacteria might be the net result of modulations of multiple biological events and of interactions among different microbes in the gut.
Different strains exhibited various anticancer activities against solid malignant tumors, including cervical cancer cell line HeLa, colorectal cancer cell lines SW480, HCT116 and HT29, and hematological malignancies, including human acute promyelocytic leukemia M3 cell line NB4, a primary culture of freshly isolated acute monocytic leukemia M5 strain ZYZ and chronic lymphocytic leukemia strain ZBC. Obviously, a majority of the 76 bacterial strains could kill solid tumors, especially the colorectal cancer, with much higher efficacy than they kill leukemia cells; however, we did find strains that killed leukemia cells highly effectively (Figure 2).
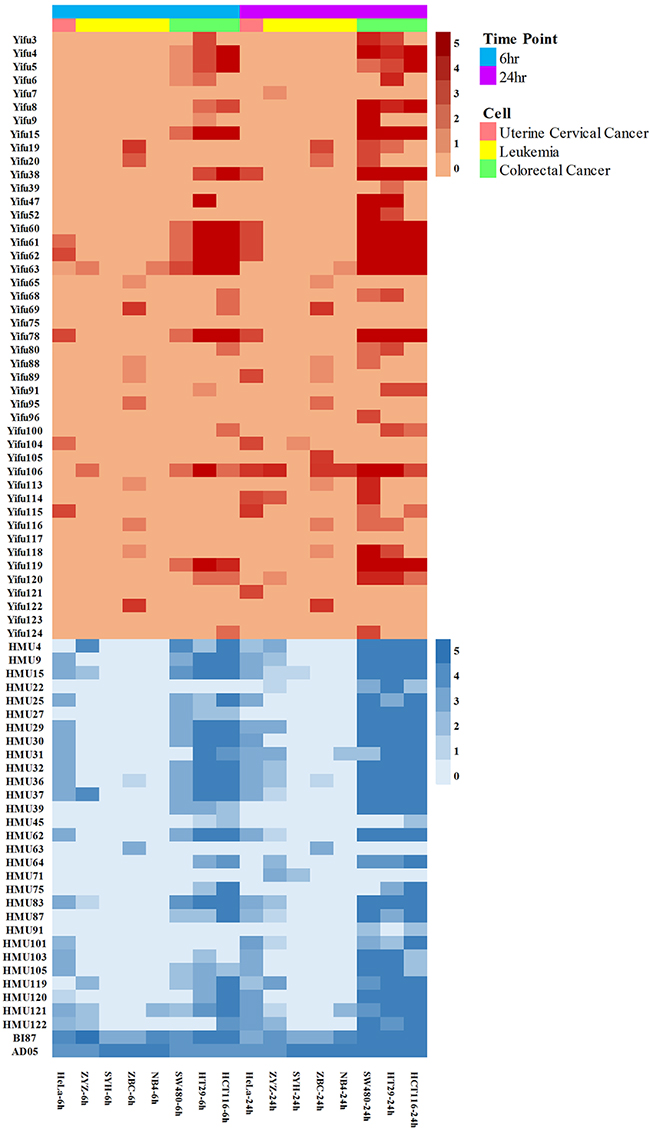
Figure 2: Anti-cancer effects of the bacterial strains against cervical cancer, leukemia and colorectal cancer.
Phylogenetic distribution of the anti-malignancy bacteria
To identify and characterize the bacteria that produced anti-malignancy effects, we shortlisted fifty-three strains for identification by 16S rDNA sequencing and compared them with known bacteria in the NCBI database (https://www.ncbi.nlm.nih.gov/genome/). Phylogenetic analyses showed that they mostly belonged to Actinobacteria but also contained lineages belonging to other phyla, including Proteobacteria and Firmicutes (Table 2), which are distantly related from one another in evolution (Supplementary Figure 1). Within the Actinobacteria phylum, the phylogenetic distribution of the bacteria was also broad (Supplementary Figure 2), with bacteria in the Streptomyces genus being most abundant in both the kindergarten (71.4%) and young adult (81.0%) groups (Figure 3). Another common genus to both groups was Rhodococcus. Overall, phylogenetic diversity of the anti-malignancy bacteria was higher in the kindergarten group than the young adult group on the genus level (Figure 4A). Some rare species, such as those of Amycolatopsis jiangsuensis and Gordonia sputi, were isolated from the kindergarten but not the young adult group. Within the genus of Streptomyces, the species were so diverse between the two groups that only one species, Streptomyces lividans, was found to be common to both of them (Figure 4B). These results suggest that the composition of the human intestinal microbiota may vary with age as previously reported [17], but more refined correlations, especially regarding the abundance changes of cancer-killing bacteria with age, would require larger sample sizes in the investigation to establish.
Table 2: Phylogenetic positions of the bacterial strains with strong anti-cancer activities
Strain |
Sample ID |
Known bacteria with closest relatedness (Accession numbers) |
Ident |
Phylum |
|---|---|---|---|---|
Yifu7 |
KG-JZH |
Amycolatopsis jiangsuensis KLBMP 1262 (NR_109638.1) |
99% |
Actinobacteria |
Yifu8 |
KG-JZH |
Streptomyces pactum CW (KX372557.1) |
100% |
Actinobacteria |
Yifu20 |
KG-HYX |
Streptomyces lividans T38 (KU317912.1) |
100% |
Actinobacteria |
Yifu38 |
KG-SR |
Streptomyces lavendofoliae NBRC 12882 (NR_112318.1) |
100% |
Actinobacteria |
Yifu52 |
KG-HBY |
Gordonia sputi IFM 10047 (FJ536305.1) |
100% |
Actinobacteria |
Yifu60 |
KG-ZYC |
Streptomyces cinnamocastaneus NBRC 14278 (AB184588.1) |
99% |
Actinobacteria |
Yifu61 |
KG-ZYC |
Streptomyces cinnamocastaneus NBRC 14278 (AB184588.1) |
99% |
Actinobacteria |
Yifu62 |
KG-ZYC |
Brevibacterium halotolerans IHB B 18060 (KU605238.1) |
99% |
Firmicutes |
Yifu63 |
KG-ZYC |
Bacillus amyloliquefaciens GC49 (KF158228.1) |
100% |
Firmicutes |
Yifu65 |
KG-WZH |
Streptomyces cinnamocastaneus NBRC 14278 (AB184588.1) |
95% |
Actinobacteria |
Yifu68 |
KG-JYX |
Streptomyces aurantiacus BCCO 10_398 (KP718492.1) |
99% |
Actinobacteria |
Yifu69 |
KG-JYX |
Streptomyces microflavus KMJ-8 (KJ020690.1) |
99% |
Actinobacteria |
Yifu75 |
KG-ZJH |
Bacillus aryabhattai L42 (KU179345.1) |
100% |
Firmicutes |
Yifu78 |
KG-SJX |
Bacillus amyloliquefaciens GC49 (KF158228.1) |
100% |
Firmicutes |
Yifu80 |
KG-PJY |
Acinetobacter lwoffii F4-2-26 (KX350033.1) |
99% |
Proteobacteria |
Yifu88 |
KG-CBC |
Acinetobacter lwoffii HaTc14 (KX150815.1) |
99% |
Proteobacteria |
Yifu89 |
KG-CBC |
Streptomyces fragilis 214109 (JN180226.1) |
99% |
Actinobacteria |
Yifu91 |
KG-WX |
Streptomyces clavifer F29 (KU324446.1) |
100% |
Actinobacteria |
Yifu95 |
KG-WJZ |
Rhodococcus corynebacterioides SCG11 (KU995335.1) |
99% |
Actinobacteria |
Yifu100 |
KG-YQT |
Streptomyces olivaceus SCSIOZ-SH16 (KC747484.1) |
100% |
Actinobacteria |
Yifu104 |
KG-TZ |
Brevibacterium halotolerans IHB B 18060 (KU605238.1) |
100% |
Firmicutes |
Yifu105 |
KG-XZH |
Acinetobacter lwoffii 272XG8 (KF818627.1) |
99% |
Proteobacteria |
Yifu113 |
KG-YPY |
Bacillus megaterium F4-2-27 (KX350034.1) |
100% |
Firmicutes |
Yifu115 |
KG-XWR |
Streptomyces lividans T38 (KU317912.1) |
99% |
Actinobacteria |
Yifu116 |
KG-XWR |
Mycobacterium arabiense W13104 (KJ676965.1) |
99% |
Actinobacteria |
Yifu117 |
KG-LEQ |
Streptomyces pactum CW (KX372557.1) |
100% |
Actinobacteria |
Yifu120 |
KG-ZJH |
Gordonia sputi Z1-2 (KJ571101.1) |
99% |
Actinobacteria |
Yifu121 |
KG-WX |
Streptomyces olivaceus FoRh46 (KM370070.1) |
100% |
Actinobacteria |
Yifu122 |
KG-WX |
Williamsia muralis 3541 (JN180186.1) |
99% |
Actinobacteria |
Yifu126 |
KG-LTW |
Brevibacterium halotolerans IHB B 18060 (KU605238.1) |
100% |
Firmicutes |
Yifu128 |
KG-WYH |
Streptomyces pactum CW (KX372557.1) |
99% |
Actinobacteria |
HMU4 |
YA-AB |
Streptomyces lividans T38 (KU317912.1) |
100% |
Actinobacteria |
HMU9 |
YA-HSJ |
Streptomyces sampsonii C.P.68 (KF991641.1) |
99% |
Actinobacteria |
BI87 |
YA-ZSY |
Streptomyces carpaticus BTSS-501 (HQ711933.1) |
99% |
Actinobacteria |
HMU15 |
YA-TZL |
Nocardia harenae YIM 131100 (KX502989.1) |
99% |
Actinobacteria |
HMU25 |
YA-LTY |
Streptomyces lividans T38 (KU317912.1) |
100% |
Actinobacteria |
HMU27 |
YA-CPP |
Saccharothrix texasensis 3536 (JN180184.1) |
99% |
Actinobacteria |
HMU29 |
YA-CH |
Streptomyces sampsonii GACK10 (KP970678.1) |
100% |
Actinobacteria |
HMU31 |
YA-WQY |
Streptomyces sampsonii GACK10 (KP970678.1) |
99% |
Actinobacteria |
HMU32 |
YA-WQY |
Streptomyces lividans T38 (KU317912.1) |
100% |
Actinobacteria |
HMU37 |
YA-FC |
Streptomyces violascens YJ-R42 (KF876851.1) |
100% |
Actinobacteria |
HMU39 |
YA-ZS |
Streptomyces fimicarius 14-5 (KJ571079.1) |
99% |
Actinobacteria |
HMU63 |
YA-LHM |
Rhodococcus fascians YIM 131099 (KX502988.1) |
100% |
Actinobacteria |
HMU64 |
YA-LHM |
Rhodococcus cercidiphylli YIM 131061 (KX502977.1) |
100% |
Actinobacteria |
HMU71 |
YA-DYHN |
Streptococcus pyogenes Sp15 (FJ662831.1) |
99% |
Firmicutes |
HMU83 |
YA-WSL |
Streptomyces plumbiresistens XB-2S-3 (JF439616.1) |
100% |
Actinobacteria |
HMU87 |
YA-JHN |
Streptomyces griseoplanus BCCO 10_1190 (KP718551.1) |
100% |
Actinobacteria |
HMU101 |
YA-SYH |
Streptomyces lavendulae xsd08101 (FJ481058.1) |
100% |
Actinobacteria |
HMU103 |
YA-SYH |
Streptomyces zaomyceticus HBUM174505 (EU841617.1) |
100% |
Actinobacteria |
HMU105 |
YA-SYH |
Streptomyces albidoflavus IHBA 9992 (KR085950.1) |
99% |
Actinobacteria |
HMU121 |
YA-SYH |
Streptomyces longwoodensis NBRC 14251 (NR_041161.1) |
99% |
Actinobacteria |
HMU122 |
YA-SYH |
Streptomyces lavendulae F1 (KU324431.1) |
100% |
Actinobacteria |
AD05 |
YA-AR |
Streptomyces lividans T38 (KU317912.1) |
100% |
Actinobacteria |
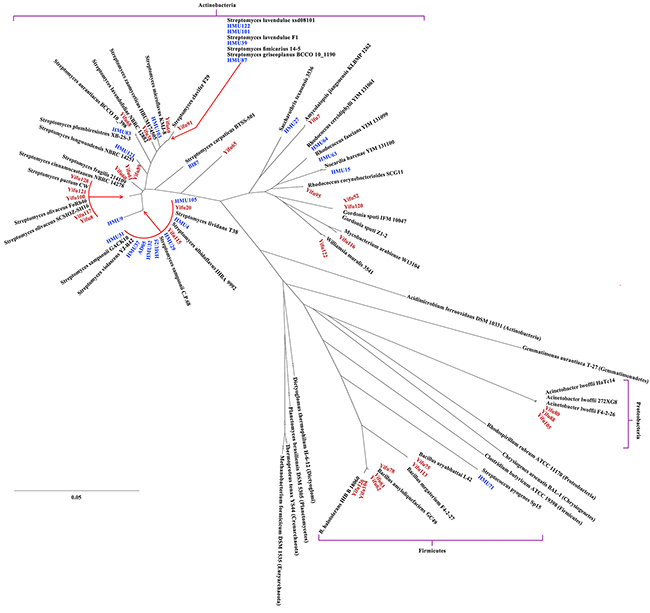
Figure 3: Phylogenetic distribution of the anti-cancer bacteria isolated in this study. Two archeal strains and strains of several representative bacterial phyla were included to illustrate the relative phylogenetic positions of the anti-cancer bacteria. Red, strains isolated from preschool children; blue, strains isolated from young adults.
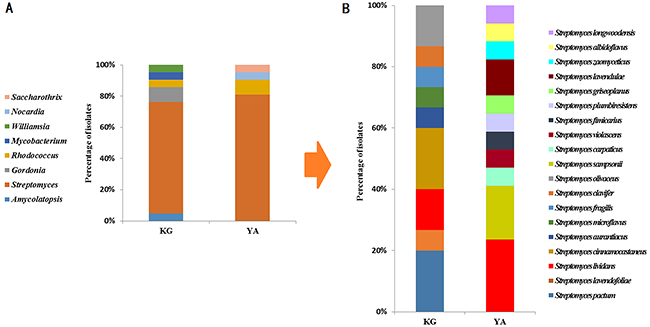
Figure 4: The relative abundance of human gut actinomycetes from kindergarten children and young adults. (A) comparison at the genus level of Actinobacteria; and (B) comparison at the species level of Streptomyces.
Broad ranges of cancer types to attack: differential efficacies
As most anti-malignancy bacterial strains exhibited broad ranges of cancer types to attack (see Figure 2), we focused on two selected strains, AD05 and BI87, to further investigate their spectra of anti-malignancy activities.
AD05 had exceptionally strong anti-malignancy activities and high stability of the effects compared to many other strains. Although most of the 76 bacterial strains exhibited strong anti-malignancy effects only on solid cancer types, AD05 exerted as strong anti-malignancy effects on leukemia as on solid cancers. We extracted the supernatant of AD05 culture with ethyl acetate and re-dissolved the dried matter in the medium. Similar to the AD05 culture supernatant, the ethyl acetate extract of AD05 also showed broad anti-cancer effects on cancer types such as ovarian cancer (A2780 and SKOV3), cervical cancer (HeLa), colorectal cancer (HCT116, SW620), human acute promyelocytic leukemia M3 (NB4), as well as a primary culture of freshly isolated chronic lymphocytic leukemia strain (ZBC) (Figure 5A). In order to examine the growth inhibitory activity of the AD05 extract on different human cancer cell lines, we used MTT assay to measure the cell viability after treatment with different concentrations. The AD05 extract exhibited significant growth inhibitory activity (P<0.05) against all the cell types tested in a dose dependent manner (Figure 5B).
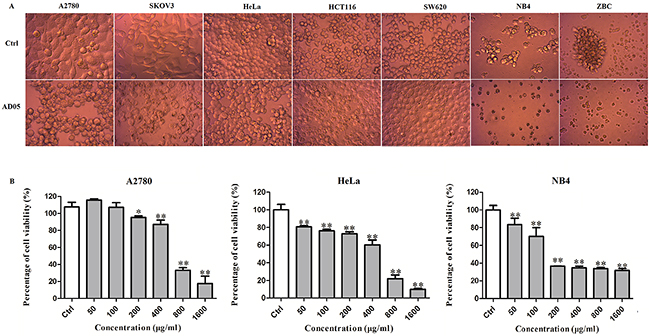
Figure 5: Anti-cancer activities of AD05 culture against human cancer cell lines. (A) Morphology of different cancer cell lines after AD05 treatment. (B) Dose dependence of anti-cancer effects of AD05 culture extract on cervical cancer cell line HeLa, ovarian cancer cell line A2780 and human acute promyelocytic leukemia M3 cell line NB4 estimated by MTT assays. Cells were treated with various concentrations of AD05 (0, 50, 100, 200, 400, 800 and 1600 μg/ml for 24h. Data were presented as mean ± standard deviation (SD) of at least three independent experiments. *P<0.05, **P<0.01 versus control.
Strain BI87 also exhibited a broad killing spectrum of cancers including ovarian cancer (ES2 and A2780), cervical cancer (HeLa), colorectal cancer (HCT116), leukemia (NB4), primary cultures of a freshly isolated acute monocytic leukemia M5 strain ZYZ and of a chronic lymphocytic leukemia strain ZBC (Figure 6A), and the MTT assays demonstrated that the effects were dose dependent (Figure 6B).
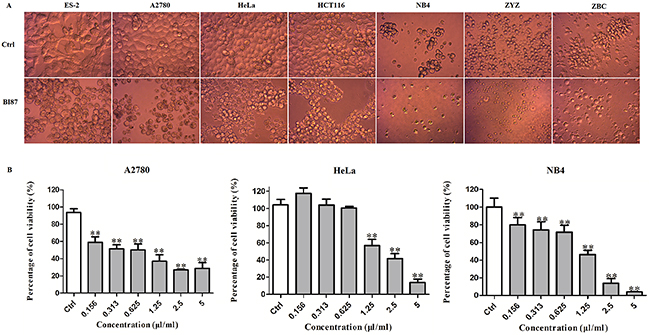
Figure 6: Anticancer activities of BI87 culture against human cancer cell lines. (A) Morphology of different cancer cell lines after BI87 treatment. (B) Dose dependence of anti-cancer effects of BI87 culture on cervical cancer cell line HeLa, ovarian cancer cell line A2780 and human acute promyelocytic leukemia M3 cell line NB4 estimated by MTT assays. Cells were treated with the various concentrations of BI87 (0, 0.156, 0.313, 0.625, 1.25, 2.5 and 5 μl/ml) for 24h. Data were presented as mean ± standard deviation (SD) of at least three independent experiments. *P<0.05, **P<0.01 versus control.
BI87 suppressed cell proliferation by inducing cell apoptosis
To determine the possible mechanism of the anti-cancer effects of BI87 and AD05, we detected the induction of apoptosis after treatment. Six hours after treatment with different concentrations, cells were double stained with Annexin V and PI and subjected to flow cytometry to quantitatively analyze the apoptotic effects. As illustrated in Figure 7, the percentages of total apoptotic cells, including the early apoptotic portion (Annexin V positive) and the late apoptotic portion (Annexin V and PI positive), were dose-dependently increased with increasing concentrations of BI87 in leukemia cell lines. Under the same experiment conditions, no significant differences were detected between the AD05 culture-treated group and the controls (data not shown). These results suggest that the BI87 but not AD05 culture could suppress cell proliferation by inducing cell apoptosis.
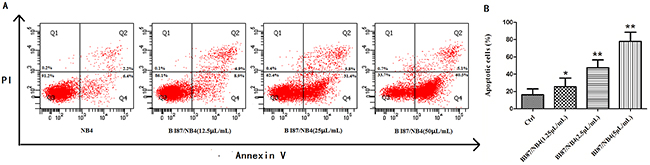
Figure 7: BI87 culture induced cancer cells apoptosis. (A) The apoptosis induced by BI87 culture detected by flow cytometry. (B) Quantitative analysis of apoptotic cells (*P<0.05, **P<0.01 versus control).
AD05 and BI87 inhibited both growth and metastasis of ovarian cancer in vivo
To test the anti-cancer effects of AD05 and BI87 in vivo, we generated a subcutaneous xenograft tumor model by transplanting ovarian cancer ES-2 cells into nude mice (Figure 8A and 8B). AD05 and BI87 treatment as single agent resulted in significant tumor volume and weight reduction compared to treatment with PBS as control (Figure 8E, 8F, 8G and 8H). The inhibition rate of tumor weight in AD05 group was 42.16%, which was a little higher than that in the BI87 group (32.02%; Figure 8F). Of great importance, no significant differences were found in spleen or body weight among the mice (Figure 8C, 8D), indicating that there was little or no harm of the anti-cancer compounds to the general health status of the mice.
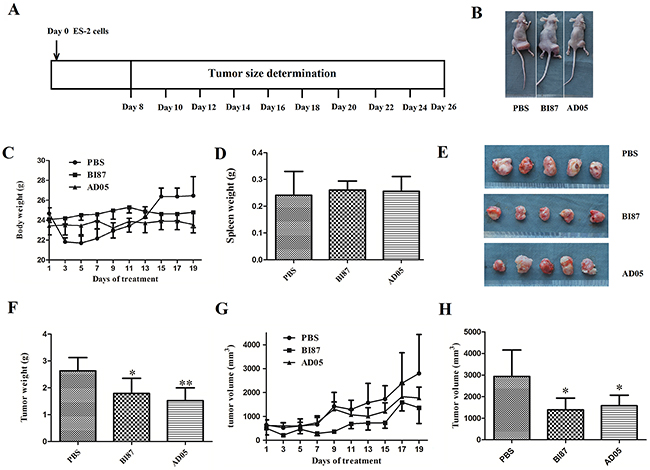
Figure 8: Tumor growth suppression in vivo by the cultures of BI87 and AD05. (A) Cancer cell inoculation and tumor size measurement schedule. (B) The subcutaneous xenograft tumor models by transplanting human ovarian cancer ES-2 cells into nude mice. (C) Body weight changes over the 19 days of tumor growth. (D) Statistical comparison of spleen weight among the animals of the three groups. The data represent the means±SD. No significant differences were seen between BI87/AD05 treated animals and controls. The P values were 0.6644/0.7540 respectively. (E) Tumor growth at day 26 in the BI87- and AD05-treated and PBS groups. (F) The removed tumors were weighted and statistically analyzed (n=5). The data represent the means ±SD. *P<0.05, **P<0.01 versus control. (G) Tumor volume changes over the 19 days of tumor growth. (H) Tumor volume comparison among the animals of the three groups at day 26. The data represent the means ±SD. *P<0.05 versus control.
Histological examinations of the tumor tissues revealed remarkable differences in the cancer cell morphology between mice of the experimental and control groups. Specifically, several necrotic foci with pyknotic and fragmented cell nuclei were observed in sections derived from BI87/AD05-injected tumors but not in sections derived from the controls, demonstrating that the supernatant of BI87/AD05 induced cell death (Figure 9A, B, C).
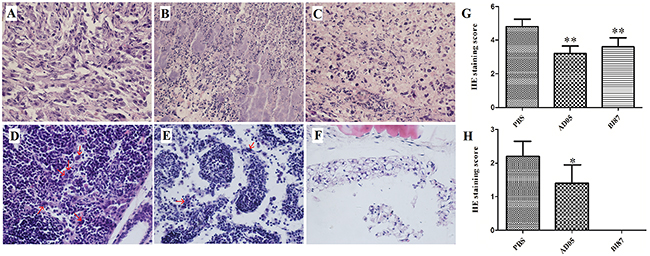
Figure 9: Histological examination of metastasis of cancer cells in nude mice with or without treatment (HE stain; magnification×400). Sections of tumor tissues from mice treated with PBS (A), AD05 (B) or BI87 (C), and sections of tissues on opposite side (without tumor growth by naked eyes) from mice treated with PBS (D), AD05 (E) or BI87 (F). (G) The statistical analyses of the HE staining score for the right thigh. (H) The statistical analyses of the HE staining score for the left thigh. The score 0 to 5 used here depended on the percentages of cancer cells stained by HE (0: 0%, 1: <5%, 2: 5%-25%, 3: 25%-50%, 4: 50%-75% and 5: >75%). It was determined by checking 15slides under the microscope (×400). The score for each slide was calculated from 5 different vision fields randomly.
As metastasis is a common cause of cancer exacerbation and patient death, we looked into the possible effects of AD05 and BI87 on the migration or survival of cancer cells at remote sites from the primary tumor by examining legs on the opposite side (the tumor was inoculated and grew in the right thigh and tissue was taken from the left thigh where no tumor growth was seen for metastasis examination). The animal experiments showed sharp contrast between AD05- or BI87-treated and control mice. In the control, the interstitial tissue of the left thigh was heavily infiltrated with cancer cells (Figure 9D) even though no tumor growth was seen by naked eyes; in AD05-treated mice, infiltrated cancer cells were only seldom found (Figure 9E); and in BI87-treated mice, no infiltrated cancer cells were found (Figure 9F). These results demonstrated that AD05 and BI87 cultures could not only kill cancer cells but also prevent tumor metastasis effectively, with BI87 having higher performance than AD05. The statistical analyses of the HE staining scores also confirmed these effects (Figure 9G, H).
GC-MS analysis of BI87 extract
Gas chromatography-mass spectrometry analysis identified three compounds present in the ethyl acetate extract of BI87 original sample (Table 3), including (1) 3-Phenethylbenzonitrile, (2) Tetrahydro-4,4,6-trimethyl-2H-pyran-2-one, and (3) 1,6:3,4-Dianhydro-2-deoxy-β-d-ribo-hexopyranose (Figure 10). Additionally, twelve compounds were identified in the methanol extract of BI87 baked sample (Table 3), including (4) Thiodiglycolic anhydride, (5) Dianhydromannitol, (6) N-Benzyloxy-2-isopropoxycarbonylazetidine, (7) Furfuryl alcohol, (8) 7-Methoxy-8-oxa-1-arabicycle(5,1,0)octane, (9) [1,2,3,4]]Tetrazolo[1,5-b][1,2,4]triazine,5,6,7,8-tetrahydro-, (10) 2-Hydroxy-2-cyclopenten-1-one, (11) Mefruside, (12) 3-Amino-2-oxazolidinone, (13) (+/-)-3-Hydroxy-gamma-butyrolactone, (14) 2-Furancarboxylic acid, tetrahydro-5-oxo-, and (15) Isosorbide dinitrate (Figure 10). These compounds were among the most abundant components in the bacterial culture supernatant, although their involvement in the anti-malignancy activities awaits further investigations.
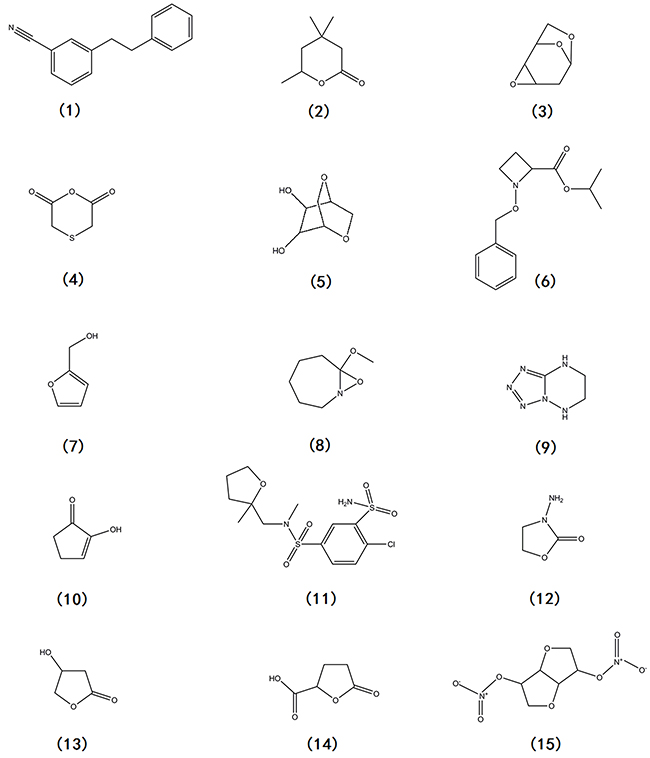
Figure 10: Chemical structures of constituents detected in BI87 extract.
Table 3: Chemical constituents identified in Streptomyces sp. BI87 extract
No. |
Constituents |
Class |
Retention time (min) |
Molecular formula |
Mol. weight |
References |
|---|---|---|---|---|---|---|
1 |
3-Phenethylbenzonitrile |
Benzonitrile |
4.191 |
C15H13N |
207 |
Molander, Gary A. et al., 2002 |
2 |
Tetrahydro-4,4,6-trimethyl-2H-pyran-2-one |
Furan |
10.191 |
C8H14O2 |
142 |
Filip Boratynski et al., 2010 |
3 |
1,6:3,4-Dianhydro-2-deoxy-β-d-ribo-hexopyranose |
Pytanose |
18.478 |
C6H8O3 |
128 |
Crandall, Jack K.et al., 1983 |
4 |
Thiodiglycolic anhydride |
Anhydride |
1.948 |
C4H4O3S |
132 |
Dar’in, Dmitry et al., 2015 |
5 |
Dianhydromannitol |
Mannitol |
2.551 |
C6H10O4 |
146 |
No paper |
6 |
N-Benzyloxy-2-isopropoxycarbonylazetidine |
Azetidine |
5.233 |
C14H19NO3 |
249 |
Kostyanovskii, R.G.et al., 1974 |
7 |
Furfuryl alcohol |
Furan |
7.382 |
C5H6O2 |
98 |
Nicolas Bosq et al.,2015 |
8 |
7-Methoxy-8-oxa-1-arabicycle(5,1,0)octane |
Azepane |
8.056 |
C7H13NO2 |
143 |
Donald H. Aue et al., 1974 |
9 |
[1,2,3,4]]Tetrazolo[1,5-b][1,2,4]triazine, 5,6,7,8-tetrahydro- |
Tetrazolo |
8.986 |
C3H6N6 |
126 |
Rodney L.Willer et al., 1987 |
10 |
2-Hydroxy-2-cyclopenten-1-one |
Cyclopentene |
9.153 |
C5H6O2 |
98 |
W. Kreiser et al., 1996 |
11 |
Mefruside |
Sulfonamides |
10.166 |
C13H19ClN2O5S2 |
382 |
Perst V. et al., 2002 |
12 |
3-Amino-2-oxazolidinone |
Oxazolidinone |
13.246 |
C3H6N2O2 |
102 |
Gregory K. Friestad et al., 2000 |
13 |
(+/-)-3-Hydroxy-gamma-butyrolactone |
Butyrolactone |
13.669 |
C4H6O3 |
102 |
Perosa A. et al., 2002 |
14 |
2-Furancarboxylic acid, tetrahydro-5-oxo- |
Furancarboxylic acid |
14.529 |
C5H6O4 |
130 |
Montagnat.Oliver D. et al., 2010 |
15 |
Isosorbide dinitrate |
Isosorbide |
15.003 |
C6H8N2O8 |
236 |
McComb MN et al., 2016 |
DISCUSSION
To our knowledge, it is the first report demonstrating the widespread existence of phylogenetically diverse bacteria that produce cancer-killing substances in the human intestine. Of particular significance, the cancer-killers have broad spectra of malignancy types to act on, including solid tumors as well as leukemia. This finding reflects a very important possibility that the anti-malignancy compounds might work not only locally (i.e., against colorectal cancers) but also systemically (i.e., against cancers in organs not in physical vicinity with the bacteria that produce the anti-malignancy compounds, such as the bone marrow, uterus, liver, ovaries, breasts, etc.), clearing stochastically appearing malignant cells at the originating site before they form lumps (solid tumors) or spreading to the whole body (leukemia and metastatic solid cancer cells) at very early stages. To date, however, investigations about bacteria and cancer have focused largely on the bacterial roles in causing (rather than clearing) cancers, especially locally, such as the well-known examples of gut bacteria causing colorectal cancer (see excellent recent reviews) [18–20]. Anti-malignancy microbes have been described mostly on viruses [21, 22], but very few on bacteria [23]. The great phylogenetic diversity of the bacteria to produce anti-malignancy substances and their broad ranges of cancer types to target demonstrate the potential clinical impacts of such bacteria and their products on malignancy therapeutics.
Among all known bacteria, actinobacteria are the best producers of bioactive secondary metabolites, producing more than two-thirds of all naturally derived antibiotics in current clinical use, as well as many anticancer, antihelmintic, and antifungal compounds [24]. Actinomycetes are closely related to the production and life of human beings. They are widely distributed in nature, and isolates from soils and oceans have so far been very extensively investigated. According to recent reports, more than 95% of the bacteria in the intestinal microbiota can be assigned to one of four major phyla: Firmicutes, Bacteroidetes, Actinobacteria and Protecteobacteria [25]. In the present study, we found that most of the cancer-killing bacteria that we had identified are actinomycetes, which might be a novel source to explore new active compounds for drug discovery.
A particularly important finding in this study is that the preschool children population seemed to have a higher ratio of individuals to possess the malignancy-killing activities than the older population (see Table 1), suggesting a fading trend of the anti-malignancy activities with age. The fact that we isolated cancer-killing bacteria also from fecal specimens tested negative for the anti-malignancy activities in the fresh supernatant indicates that the malignancy-killers might be even more widespread in the human populations than shown here but the eventual expression of their malignancy-killing effects might require optimal balancing and coordination of multiple factors, which may involve human genetic predisposition as well as environmental disturbance such as dysbiosis due to certain kinds of diets or abusive use of antimicrobials, which calls for further investigation.
An exceptionally interesting fact is that one individual may have phylogenetically very diverse malignancy-killers, as seen in participant KG-HYX, who had the malignancy-killers belonging to two phyla, Firmicutes and Actinobacteria; within a phylum, the diversity was also enormous (see Table 2 and Supplementary Figure 2). It can be anticipated that larger scales of screening of the intestinal microbes, not only bacteria but possibly also other kinds of microorganisms like fungi or even protozoa, may reveal even greater diversity of malignancy-killers. As such, mechanisms used by the malignancy-killers for the anti-cancer activities may involve extensive molecular pathways.
Our findings in this study bring up four updates to the biomedical research fields, especially regarding carcinogenesis investigations and clinical therapeutics of cancers. First, the widespread existence of malignancy-killers in the human populations at large provides a novel explanation to the fact that many people at high genetic or environmental risks to cancer may not have cancer in their long lifetime, revealing a long ignored arsenal against cancers in addition to the immune system. Second, the importance of maintaining a normal microbiota, such as by avoiding antimicrobial abuse, is reiterated to reduce cancer risks. Third, as the isolated bacterial strains that produced the anti-malignancy activities are phylogenetically very diverse and the damages caused by the cancer-killing effects of the bacteria to the cancer cells are morphologically remarkably distinct, the anti-malignancy substances produced by the bacteria, and hence the mechanisms in cancer killing, may be highly different, encouraging the combined use of the different substances for even greater effects in the treatment of cancers. Finally, the natural anti-cancer substances may become novel lead compounds for cancer drug development.
MATERIALS AND METHODS
Collection of fecal specimens and isolation of bacteria
Fecal specimens were collected from 100 preschool children of Yifu Kindergarten, 3-7 years old, and 113 young adults, 18-39 years old, including faculty, postdoctoral fellows, and graduate and undergraduate students, Harbin Medical University, Harbin, China. We obtained the informed consents and detailed information of the participants including age, sex, and family history. The fecal specimens were mixed with equal volume of sterile distilled water and centrifuged at 10000 rpm/min for 30 min. The supernatant was saved for detection of anti-cancer effects and the pellets were treated as follows for bacterial isolation. One gram of the fecal pellet was suspended in 10 ml PBS (KH2PO4 0.27g, NaHPO4 1.42g, NaCl 8g, KCl 0.2g and 1L distilled water) and shaken for 30 minutes at 60 rpm/min. After the shaking, we placed the tubes on the bench and waited for 10 min for the large debris to become settled to the bottom of the tubes. We then inoculated the upper part of the samples by to 10:1, 100:1 and 1000:1 dilutions into the Gauze No. 1 medium (solution starch 20 g, KNO3 1 g, K2HPO4 0.5g, MgSO4·7H2O 0.5g, NaCl 0.5g, FeSO4·7H2O 0.01g, K2Cr2O7 50mg and distilled water added to 1L) and incubated the mixture at 28°C. Periodically, we spread the liquid culture onto Gauze No. 1 agar plates and picked up colonies with different morphological characteristics, with a focus on actinobacteria-like bacteria. The chosen colonies were transferred into fresh Gauze No.1 liquid medium for further culture and saturated pure cultures were obtained for tests as well as for preservation in 25% (v/v) glycerol at -80°C.
Cell culture
Human solid cancer cells of several cell lines, including cervical cancer cell line HeLa, ovarian cancer cell lines SKOV3 and A2780, and colorectal cancer cell lines HCT116 and SW620, were cultured in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine serum. Ovarian cancer cell line ES-2 was cultured in McCoy’s 5A medium with 10% fetal bovine serum. Human acute promyelocytic leukemia NB4 cells were cultured in RPMI 1640 medium, supplemented with 10% fetal bovine serum, 100 units/mL penicillin and 100 μg/mL streptomycin. All the cultures were maintained in an incubator at 37°C with 5% CO2 in a humidified atmosphere.
Isolation and cultivation of leukemia cells
Heparin-anticoagulated peripheral blood from a leukemia patient was centrifuged at 1500 rpm/min for 10 min. The plasma was removed and the blood cells were mixed with equal volume of cell culture medium without serum. The mixture was then added slowly to the top of two volumes of leukocyte separation medium (Tianjin Haoyang biological technology co., LTD) before centrifugation at 2000 rpm/min for 20 min. The leukemia cells in the leukocyte layer were transferred to a fresh 15 ml sterile centrifuge tube, washed with PBS and cultured in RPMI 1640 medium supplemented with 10% fetal calf serum in an incubator at 37°C with 5% CO2 in a humidified atmosphere.
Morphological assessment
Morphological changes of cells treated with fecal supernatant or extractions of the metabolites from the bacterial cultures were inspected by phase contrast inverted microscopy (Zeiss Axiocam Erc 5s, Germany). The performance of the experiments and the determination of experimental results were completed blindly and separately by at least two different persons. The cells treated with the same volume of Gauze No.1 medium and the cells without treatment served as control groups.
Bacterial genomic DNA extraction
Genomic DNA from bacterial strains that showed strong anti-cancer activities was isolated for 16S rDNA sequencing. For each strain, we picked a single colony and inoculated it to 5 mL Gauze No.1 medium. The incubation was conducted in a shaker at 28°C for 7 days. After that, we transferred the culture to 50 mL Gauze No.1 fresh medium to continue the incubation for 7 days. After centrifugation at 4000 rpm/min for 15 min, the supernatant was discarded. The pellets were crushed down and suspended in 5 ml 10 mM EDTA and then the suspension was centrifuged at 4000 rpm/min for 15min. Next, the pellet was suspended in 5 ml SET buffer (10 ml 1M Tris-HCl pH7.5, 25ml 0.5M EDTA pH8.0, 7.5 ml 5M NaCl, and H2O adjusted to 1L). Following that, we added 50 μl 100 mg/ml lysozyme to the suspension and kept it at 37°C for 1 hour, and then added 600 μl 10% SDS and 150 μl 20mg/ml proteinase K to the suspension and kept it at 55°C for 2 hours. After mixing the suspension with 2 ml 5M NaCl and 5 ml chloroform and shaking the mixture for 30 minute, we centrifuged it at 4000 rpm/min for 30 min and transferred the aqueous phase to an Eppendorf tube. To this, we added an equal volume of isopropanol and centrifuged the tube at 8000 rpm/min for 20 min. We washed the DNA pellet by adding 5 ml 70% ethanol and centrifuging the tube at 8000 rpm/min for 20 min. After the DNA pellet was air dried, we added 200 μl TE buffer to it and stored it at −20°C.
16S rRNA gene amplification and phylogenetic analyses
Primers for amplifying the 16S rRNA gene were designed according to the highly conserved regions of 16S rRNA gene sequences [14], and we used F (5’-AGAGTTTGATCCTGGCTCAG-3’) and R (5’-AAGGAGGTGATCCAGCCGCA-3’) here. The 25 μl PCR mixture contained 1 μl bacterial genomic DNA, 1 μl primer F and 1 μl primer R, 0.5 μl dNTP, 0.2 μl DNA polyase, 2 μl PCR buffer and 19.3 μl deionized water. The PCR conditions were as follows: the initial denaturation was set at 95°C for 5 min, followed by 10 cycles of 94°C for 30 s, 61°C for 30 s, and 72°C for 45 s, then 25 cycles of 94°C for 30 s, 53°C for 30 s, and 72°C for 45 s, with final extension for 10 min at 72°C. All amplification products were sent to Genewiz company for sequencing. The 16S rDNA sequence data reported in the present study were deposited in the Genbank nucleotide database under the accession numbers KX900586 (Yifu7), KU058402 (Yifu8), KU058420 (Yifu20), KU058403 (Yifu38), KX900587 (Yifu52), KX900588 (Yifu60), KX900589 (Yifu61), KU058404 (Yifu62), KU058414 (Yifu63), KX900590 (Yifu65), KU058415 (Yifu68), KX900591 (Yifu69), KU058405 (Yifu75), KU058416 (Yifu78), KU058417 (Yifu80), KX900592 (Yifu88), KX900593 (Yifu89), KU058418 (Yifu91), KX900594 (Yifu95), KU058406 (Yifu100), KU058419 (Yifu104), KX900595 (Yifu105), KX900596 (Yifu113), KX900597 (Yifu115), KX900598 (Yifu116), KX900599 (Yifu117), KX900600 (Yifu120), KX900601 (Yifu121), KX900602 (Yifu122), KX900603 (Yifu126), KX900604 (Yifu128), KU058421 (HMU4), KX900573 (HMU9), KU058407 (BI87), KX900574 (HMU15), KX900575 (HMU25), KU058408 (HMU27), KX900576 (HMU29), KX900577 (HMU31), KX900578 (HMU32), KX900579 (HMU37), KU058409 (HMU39), KX900580 (HMU63), KX900581 (HMU64), KU058410 (HMU71), KX900582 (HMU83), KX900583 (HMU87), KU058411 (HMU101), KU058412 (HMU103), KU058413 (HMU105), KX900584 (HMU121), KX900585 (HMU122), KU058430 (AD05). The sequences were compared for similarity with the reference species of bacteria using the NCBI BLAST. They were aligned using the Clustal X program [15] and the phylogenetic tree was constructed by MEGA 6 using the neighbor-joining algorithm [16]. Evolutionary distances for the neighbor-joining algorithm were computed using p-distance model. The stability of the tree topologies were evaluated by using the bootstrap based on 100 replications.
Extraction of the metabolites from bacterial cultures
The culture supernatant was filtered by vacuum and then extracted by equal volume of ethyl acetate for three times. The extraction was dried by rotary evaporator at 40°C and stored at 4°C.
MTT assay
Cultured cells were distributed into 96-well plates in 100 μL of medium, incubated overnight and then treated with the compounds over a range of concentrations for the indicated time. After the addition of 20 μL of 5 mg/mL MTT solution per well, the plates were incubated for another 4 h. The media were removed, and the formazan crystals were solubilized in 150 μL of DMSO and shaken for 10 min. The absorbance was measured at 490 nm using a microplate reader (Emax@ Plus, England). The percentage of cell viability was calculated as follows:
% cell viability = Absorbance of treated cells/Absorbance of untreated cells × 100%
Cell apoptosis analysis
To detect apoptosis, cells were incubated with culture of BI87 or AD05 for 6h. The cells were harvested, washed twice with cold 1xPBS, and re-suspended in 100μL 1×Binding buffer at density of 1×105 cells/mL. The cells were then stained with 5μL Annexin V and 5μL PI (BD Biosciences) for 15min in dark condition at room temperature. After staining, we added 400 µl of 1×Binding Buffer to each tube. The samples were subjected to analysis by flow cytometry (BD FACSCantoTM II). The early apoptosis was evaluated based on the percentage of Annexin V positive and PI negative cells, while the late apoptosis was evaluated based on the percentage of Annexin V positive and PI positive cells.
In vivo tumor growth assay
Female nude mice at 6-8 weeks of age were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd., and housed in a specific pathogen free facility. Mice were subcutaneously inoculated with ES-2 cells (0.33 × 106 suspended in 0.08 mL PBS for each mouse). When the tumor reached an average volume of 500 mm3, the animals were randomized into groups (n=5) and treated with sterile supernatant of the bacterial cultures or with PBS, 1ml/kg, by intra-tumor injection every other day. The measurements of tumor growth with a digital caliper were done once every other day. Tumor volumes were calculated by the two dimensional sizes of each tumor with the following formula: V = L×W2×1/2, where V is the volume, L is the length, and W is the width. At the end of experiment, the mice were weighed and sacrificed, and the tumors were dissected, weighed and stored in 4% paraformaldehyde solution for hematoxylin-eosin staining.
Hematoxylin-eosin staining
Following euthanasia of the mice, the tumor tissue was removed aseptically and immediately fixed in a 4% paraformaldehyde solution (Wuhan Boster Biological Technology Co.,Ltd). The fixed tumors were processed through graded concentrations of ethanol and xylene and were then embedded in paraffin. The tissue sections were mounted on glass slides and stained with HE. In total, 15 slides were inspected and the scoring was performed.
Gas chromatography-mass spectrometry (GC-MS) analysis
We employed the Agilent Technologies 7890B (GC) machinery equipped with 5977A Mass Selective Detector (MS), HP-5MS (5% phenyl methyl siloxane) capillary column of dimensions 30m×250μm×0.25μm and used helium as the carrier gas at 1 mL/min. For the original sample, the column temperature was programmed initially at 40°C for 3min, followed by an increase of 10°C /min to 220°C and then an increase of 25°C /min to 280°C. For the baked sample, the column temperature was programmed initially at 40°C for 5min, followed by an increase of 10°C /min to 180°C and then an increase of 20°C /min to 280°C. The MS was operated at 70eV. The constituents were identified by comparison of their mass spectral data with those from NIST 11 Spectral Library.
Statistical analysis
Data were presented as mean ± standard deviation (SD) of at least three independent experiments. The MTT assay results were obtained by GraphPad Prism statistical software. The statistical significance was accepted at P < 0.05.
Ethics statement
We obtained a written informed consent from each participant or their guardian, consistent with the 1975 Declaration of Helsinki. All experimental protocols were reviewed and approved by The Ethics Committee, Harbin Medical University, and all experiments were performed in accordance with relevant guidelines and regulations.
Author Contributions
SLL conceived the research, YJZ and SLL designed the project and drafted the manuscript, XQM, XL, ZL, participated in sample collection, YJZ, DDZ, HTC, JJL, ZYZ, JHW, YXC, YZH, HL, PGW, YYJ, PQL, XP, SYC, ZLY, XYY, TL and BQY conducted the experiments, LT, YGL, GRL and SLL contributed reagents/ materials/ analysis tools. SLL finalized the manuscript. All authors read and approved the final manuscript.
ACKNOWLEDGEMENTS
We thank all participants who provided fecal specimens for their cooperation. Special thanks go to the following individuals who provided specimens and also technical assistance: Xiao-Yun Chang, Xin Fang, Gang Jin, Bailiang Li, Qing-Hai Li, Bin-Bin Liu, Yang-Peng Liu, Lauren Mak, Emilio Mastriani, Alexey Rakov, Qiang Sun, Zi-Qiao Yan, Lu Yu, Chao Yue, Zheng Zeng, Songling Zhu.
CONFLICTS OF INTEREST
The authors declare no competing financial interests.
FUNDING
This work was supported by a Heilongjiang Innovation Endowment Award for graduate studies to YJZ (YJSCX2012-214HLJ) and grants of the National Natural Science Foundation of China (NSFC81030029, 81271786, 81671980) to SLL. HDL was supported by a scholarship from China Scholarship Council, CSC No. 201508230143 and College students Innovation & Entrepreneurship project in Heilongjiang Province (201410226047, 201510226020). ZL was supported by a grant from the Natural Science Foundation of Heilongjiang Province (no. QC2013C086) and a grant from the Science Foundation of Health Department of Heilongjiang Province (no. 2012-452). ZZY was supported by Natural Science Foundation of Heilongjiang Province, China (QC2014C095) and Health and Family Planning Commission of Heilongjiang Province, China (2014-434).
REFERENCES
1. Fearon ER. Human cancer syndromes: clues to the origin and nature of cancer. Science. 1997; 278:1043-50.
2. Lichtenstein P, Holm NV, Verkasalo PK, Iliadou A, Kaprio J, Koskenvuo M, Pukkala E, Skytthe A, Hemminki K. Environmental and heritable factors in the causation of cancer-analyses of cohorts of twins from Sweden, Denmark, and Finland. N Engl J Med. 2000; 343:78-85.
3. Danaei G, Vander Hoorn S, Lopez AD, Murray CJ, Ezzati M, Comparative Risk Assessment collaborating group (Cancers). Causes of cancer in the world: comparative risk assessment of nine behavioural and environmental risk factors. Lancet. 2005; 366:1784-93.
4. Tomasetti C, Vogelstein B. Cancer etiology. Variation in cancer risk among tissues can be explained by the number of stem cell divisions. Science. 2015; 347:78-81.
5. Thompson LU, Robb P, Serraino M, Cheung F. Mammalian lignan production from various foods. Nutr Cancer. 1991; 16:43-52.
6. Adlercreutz H, Mousavi Y, Clark J, Höckerstedt K, Hämäläinen E, Wähälä K, Mäkelä T, Hase T. Dietary phytoestrogens and cancer: in vitro and in vivo studies. J Steroid Biochem Mol Biol. 1992; 41:331-37.
7. Sung MK, Lautens M, Thompson LU. Mammalian lignans inhibit the growth of estrogen-independent human colon tumor cells. Anticancer Res. 1998; 18:1405-08.
8. Thompson LU. Experimental studies on lignans and cancer. Baillieres Clin Endocrinol Metab. 1998; 12:691-705.
9. Adlercreutz H. Lignans and human health. Crit Rev Clin Lab Sci. 2007; 44:483-525.
10. Li MX, Zhu HY, Yang DH, Ma XQ, Wang CZ, Cai SQ, Liu GR, Ku BS, Liu SL. Production of secoisolariciresinol from defatted flaxseed by bacterial biotransformation. J Appl Microbiol. 2012; 113:1352-61.
11. Tao YL, Yang DH, Zhang YT, Zhang Y, Wang ZQ, Wang YS, Cai SQ, Liu SL. Cloning, expression, and characterization of the β-glucosidase hydrolyzing secoisolariciresinol diglucoside to secoisolariciresinol from Bacteroides uniformis ZL1. Appl Microbiol Biotechnol. 2014; 98:2519-31.
12. Wang CZ, Ma XQ, Yang DH, Guo ZR, Liu GR, Zhao GX, Tang J, Zhang YN, Ma M, Cai SQ, Ku BS, Liu SL. Production of enterodiol from defatted flaxseeds through biotransformation by human intestinal bacteria. BMC Microbiol. 2010; 10:115.
13. Zhu HY, Li MX, Yang DH, Tao YL, Zhang Y, Liu SL. Biotransformation of the SDG in defatted flaxseed into END co-cultured by three single bacterial colonies. Process Biochem. 2014; 49:19-24.
14. Olsen GJ, Larsen N, Woese CR. The ribosomal RNA database project. Nucleic Acids Res. 1991; 19:2017-21.
15. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997; 25:4876-82.
16. Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987; 4:406-25.
17. Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez-Bello MG, Contreras M, Magris M, Hidalgo G, Baldassano RN, Anokhin AP, Heath AC, Warner B, Reeder J, et al. Human gut microbiome viewed across age and geography. Nature. 2012; 486:222-27.
18. Schwabe RF, Jobin C. The microbiome and cancer. Nat Rev Cancer. 2013; 13:800-12.
19. Sears CL, Garrett WS. Microbes, microbiota, and colon cancer. Cell Host Microbe. 2014; 15:317-28.
20. Marchesi JR, Adams DH, Fava F, Hermes GD, Hirschfield GM, Hold G, Quraishi MN, Kinross J, Smidt H, Tuohy KM, Thomas LV, Zoetendal EG, Hart A. The gut microbiota and host health: a new clinical frontier. Gut. 2016; 65:330-39.
21. Ahmad T, Venkataraman S, AbouHaidar M, Hefferon KL. Recent Patents in Oncolytic Virotherapy. Recent Pat Biotechnol. 2016; 9:79-85.
22. Clements DR, Kim Y, Gujar SA, Lee PW. All that glitters is not gold: the need to consider desirable and undesirable immune aspects of oncolytic virus therapy. OncoImmunology. 2015; 5:e1057674.
23. Hoffman RM. Tumor-Targeting Salmonella typhimurium A1-R: an Overview. Methods Mol Biol. 2016; 1409:1-8.
24. Barka EA, Vatsa P, Sanchez L, Gaveau-Vaillant N, Jacquard C, Klenk HP, Clément C, Ouhdouch Y, van Wezel GP. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol Mol Biol Rev. 2015; 80:1-43.
25. Landman C, Quévrain E. [Gut microbiota: Description, role and pathophysiologic implications]. [Article in French]. Rev Med Interne. 2016; 37:418-23.