
INTRODUCTION
Breast cancer is the most frequently occurring cancer and the leading cause of cancer-related death among women worldwide, with an estimated 1.7 million incident cases and 521,900 deaths in 2012 [1]. Epidemiological studies have shown that obesity, advanced maternal age at first birth, estrogen and progestin use, physical inactivity and alcohol consumption are associated with an increased risk of breast cancer in women [2–5]. Some of these factors also influence the prognosis of patients after treatment. However, people sharing the same conditions and familial aggregation have different lifetime risks, indicating that genetic factors are essential to breast cancer etiology [6, 7].
Besides genetic mutations, epigenetic mechanisms are also important in the tumorigenesis of breast cancer [8–10]. Epigenetic modifications include DNA methylation, histone modification and noncoding RNA (ncRNA). A significant fraction of the human genome is transcribed as ncRNAs [11]. The functional relevance of ncRNAs is particularly evident for microRNAs (miRNAs) and long noncoding RNAs (lncRNAs) [12]. Over the last few years, reports have indicated that miRNAs are involved in all aspects of the carcinogenic process and can be used as biomarkers for early risk stratification and long-term survival prediction [13, 14]. Specific miRNA expression profiles can enable accurate tumor classification, targeted therapy and individualized intervention [15, 16].
Circular RNA (circRNA) is another novel class of endogenous ncRNA molecules [17]. CircRNAs were found in viroids, viruses and tetrahymena decades ago, but they were initially considered to be by-products of aberrant RNA splicing or splicing errors due to their low expression [18, 19]. Thanks to technological breakthroughs in high-throughput sequencing and computational approaches, circRNAs have drawn increasing interest, especially with the discovery of their vast abundance, diverse functions, and frequent tissue-specific expression [20, 21]. The expression of circRNAs is outstandingly high compared with the expression of linear RNA isoforms of the same genes [22, 23]. Recent evidence has indicated that circRNAs can regulate gene expression by sequestering specific miRNAs or buffering their repression of mRNA targets [17, 24]. For example, the circRNA Cdr1as, also known as CiRS-7, has been shown to act as a powerful miR-7 sponge/inhibitor in the developing midbrain of zebrafish [25].
Aberrant expression of circRNAs has been shown to occur in colorectal, basal cell and bladder carcinoma [26–28]. Recently, the involvement of circRNAs in breast cancer has also been explored, but only preliminary findings have been reported and there has been a lack of experimental and clinical evidence [29, 30]. Nair et al. developed a Circ-Seq workflow to identify expressed circRNAs, and found that circRNAs may be markers of cell proliferation in breast cancer and associate with cancer subtypes [30]. Galasso et al. used a bioinformatics detection tool to explore the predictive value of circRNAs in breast cancer, but the size of the cohort was small [29].
Here, we performed a molecular epidemiological study in a Chinese population to establish the circRNA expression profile and identify deregulated circRNAs in the carcinogenesis of human breast cancer.
RESULTS
Characteristics of the study population
In the first stage, we used the Arraystar Human circRNA Array to sequence four paired breast cancer samples from patients with invasive ductal breast cancer (Supplementary Table 1). In the validation stage, we recruited 51 breast cancer patients. The mean (range) age was 46.5 (18-79) years. The characteristics of the patients are presented in Table 1.
Table 1: Characteristics of study subjects in the validation study (n=51)
Characteristics |
Number |
% |
|---|---|---|
Age (years) |
|
|
≤60 |
31 |
60.78 |
>60 |
20 |
39.22 |
ER |
|
|
positive |
26 |
50.98 |
negative |
25 |
49.02 |
PR |
|
|
positive |
19 |
37.25 |
negative |
32 |
62.75 |
HER2 |
|
|
positive |
30 |
58.82 |
negative |
21 |
41.18 |
Tumor stage |
|
|
T1 |
16 |
31.37 |
T2 |
30 |
58.82 |
T3 |
5 |
9.80 |
Lymphatic metastasis |
|
|
N0 |
25 |
49.02 |
N1 |
17 |
33.33 |
N2 |
9 |
17.65 |
ER: estrogen receptor; PR: progestin receptor; HER2: human epidermal growth factor receptor-2.
Findings from the circRNA microarray
Differentially expressed circRNAs were detected in the four matched tissue samples. When we set the filter criteria as a fold-change ≥2 and a P-value <0.05, we found that a total of 715 circRNAs were significantly upregulated and 440 circRNAs were significantly downregulated in the breast cancer lesions compared with adjacent normal-appearing tissues. Considering that false positives can be caused by multi-comparisons, we used the false discovery rate (FDR) method to adjust the P values. After FDR correction, a total of 16 circRNAs were found to be significantly upregulated, while 5 circRNAs were significantly downregulated. Figure 1 is a hierarchical cluster displaying the levels of circRNAs in breast cancer and adjacent normal-appearing tissues. Figure 2 is a scatterplot describing the variation in circRNA expression between the two groups. Figure 3 is a volcano plot visualizing the statistical significance of differentially expressed circRNAs between cases and controls.
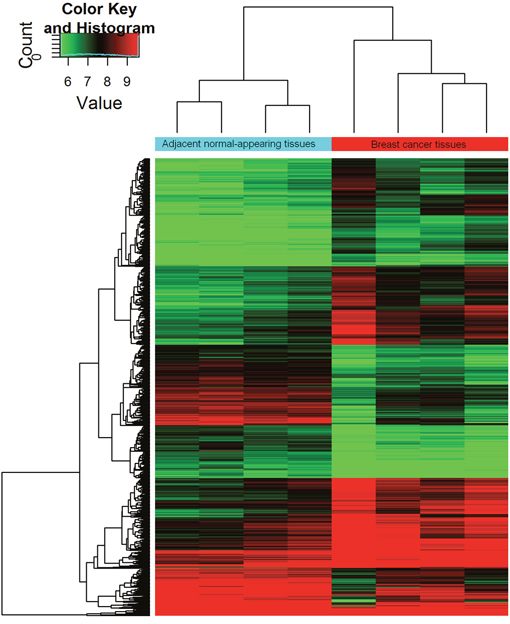
Figure 1: Hierarchical cluster of circRNA expression profiles from the microarray. The color scale runs from green (low intensity), through black (medium intensity), to red (strong intensity).
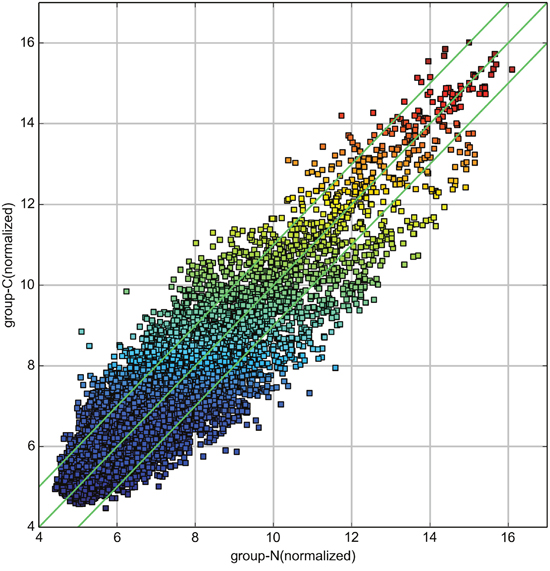
Figure 2: Scatterplot of circRNA signal values between breast cancer and adjacent normal-appearing tissues. The values spotted in the X and Y axes represent the normalized signals of samples in the two groups (log2-scaled). The green lines represent fold-changes. The circRNAs above the upper green line and below the lower green line are those with expression fold-changes >2.0 between breast cancer and normal-appearing tissues.
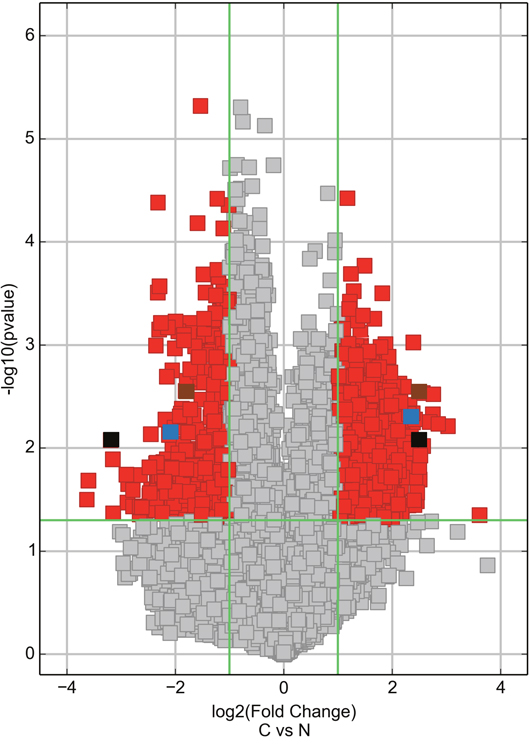
Figure 3: Volcano plot visualizing the differential expression of circRNAs between breast cancer and adjacent normal-appearing tissues. The vertical green lines correspond to up- and downregulation >2.0-fold. The horizontal green line represents a P-value of 0.05. The red squares indicate differentially expressed circRNAs with statistical significance. On the right side, the brown square represents hsa_circ_103110, the blue square represents hsa_circ_104689, and the black square represents hsa_circ_104821. On the left side, the brown square represents hsa_circ_100219, the blue square represents hsa_circ_006054, and the black square represents hsa_circ_406697. C: breast cancer tissue. N: adjacent normal-appearing tissue.
Validation of deregulated circRNAs
To test whether the differentially expressed circRNAs discovered through the microarray were bona fide, we selected six potentially significant circRNAs for validation by the quantitative real-time reverse transcription PCR (qRT-PCR). We used the following criteria: (1) length around 200 to 3000 bp; (2) fold-change >2; (3) p-value <0.01; (4) raw intensity >200; (5) exonic-related circRNAs; and (6) conservative. These six circRNAs are highlighted in the volcano plot (Figure 3). The scatterplot revealed that hsa_circ_103110, hsa_circ_104689 and hsa_circ_104821 levels were elevated in cancer tissues, whereas hsa_circ_006054, hsa_circ_100219 and hsa_circ_406697 were downregulated in breast cancer lesions (Figure 4). Subgroup analysis demonstrated that progestin receptor negativity (PR-) was associated with upregulation of hsa_circ_104689 and hsa_circ_104821, and with downregulation of hsa_circ_406697 (Supplementary Table 2). We further evaluated the diagnostic value of these six circRNAs for breast cancer by plotting a receiver operating characteristic curve. We found that hsa_circ_100219 had the highest diagnostic accuracy, with an area under the curve (AUC) of 0.78 (95% CI: 0.69-0.88) (Table 2). When hsa_circ_006054, hsa_circ_100219 and hsa_circ_406697 were combined, the AUC increased to 0.82 (95% CI: 0.73-0.90) (Figure 5).
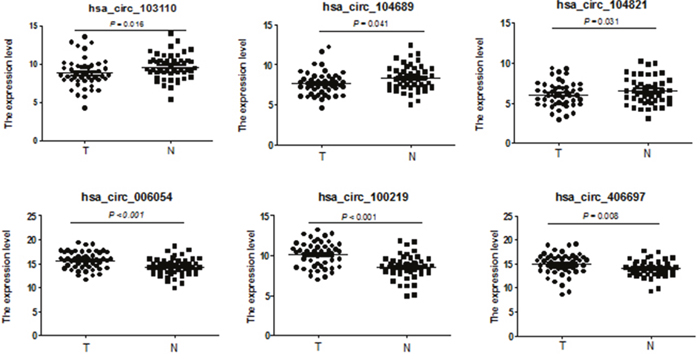
Figure 4: Validation of novel circRNAs by qRT-PCR in breast cancer and adjacent normal-appearing tissues. Scatterplots display the relative expression of specific circRNAs between breast cancer (T) and adjacent normal-appearing tissues (N). Marks above the reference line indicate downregulated circRNAs.
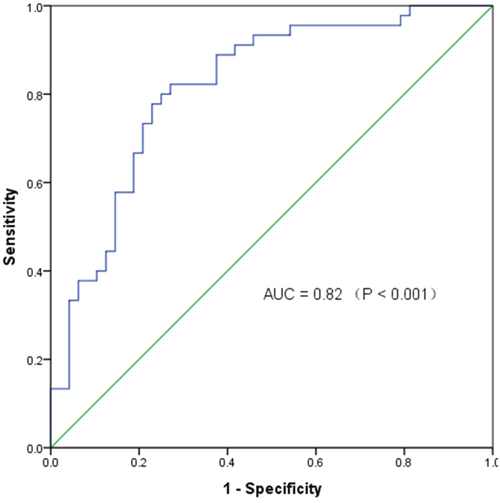
Figure 5: The receiver operating characteristic curve of specific circRNAs in distinguishing breast cancer. AUC: area under the curve; Combination of circRNAs: hsa_circ_006054, hsa_circ_100219 and hsa_circ_406697.
Table 2: Expression and diagnostic value of specific circRNAs in breast cancer
CircRNAs |
Expressiona |
t |
P |
AUC (95% CI) |
Sensitivity (95% CI) |
Specificity (95% CI) |
Cut-off valueb |
|
|---|---|---|---|---|---|---|---|---|
Breast cancer |
Adjacent tissues |
|||||||
hsa_circ_103110 |
8.79±1.86 |
9.62±1.63 |
2.49 |
0.016 |
0.63 (0.52-0.74) |
0.63 (0.48-0.76) |
0.63 (0.48-0.76) |
8.97 |
hsa_circ_104689 |
7.66±1.48 |
8.31±1.57 |
2.09 |
0.041 |
0.61 (0.50-0.73) |
0.57 (0.42-0.71) |
0.55 (0.40-0.69) |
7.67 |
hsa_circ_104821 |
6.07±1.55 |
6.52±1.68 |
2.22 |
0.031 |
0.60 (0.49-0.71) |
0.57 (0.42-0.71) |
0.57 (0.42-0.71) |
6.04 |
hsa_circ_006054 |
15.66±1.82 |
14.29±1.75 |
4.30 |
<0.001c |
0.71 (0.61-0.81) |
0.65 (0.50-0.78) |
0.69 (0.54-0.81) |
14.84 |
hsa_circ_100219 |
10.13±1.58 |
8.53±1.43 |
5.70 |
<0.001c |
0.78 (0.69-0.88) |
0.69 (0.54-0.81) |
0.71 (0.56-0.83) |
8.95 |
hsa_circ_406697 |
14.85±2.27 |
13.96±1.68 |
2.76 |
0.008c |
0.64 (0.52-0.75) |
0.63 (0.48-0.76) |
0.63 (0.48-0.76) |
14.24 |
a: The expression of each circRNA was calculated from the ΔCt and expressed as mean±standard deviation; b: We used the median as the cut-off value; c: Significant after Bonferroni correction (P<0.008); AUC: area under the curve; CI: confidence interval.
GO enrichment and KEGG analysis
To explore how circRNAs regulate parental gene transcription, we performed Gene Ontology (GO) enrichment analysis for the genes targeted by the circRNAs that were found to be differentially expressed in our qRT-PCR results. Target genes of upregulated circRNAs in breast cancer were involved in the developmental process, positive regulation of gene expression and positive regulation of biological processes (Supplementary Figure 1A), whereas target genes of downregulated circRNAs in breast cancer were involved in the transmembrane receptor protein tyrosine kinase signaling pathway and the developmental process (Supplementary Figure 1B). Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis revealed that pathways such as the Hippo signaling pathway and the WNT signaling pathway were related to the upregulated circRNAs, while pathways such as the RAP1 signaling pathway and the RAS signaling pathway were related to the downregulated circRNAs (Supplementary Figure 2A & 2B).
MiRNA response element (MRE) sequence analysis
As shown in Table 3, dominant MREs were targeted by differentially expressed circRNAs. Some of these miRNAs have been reported to be associated with breast cancer [31–38]. By referring to published literature, we identified the following miRNA-circRNA matches: hsa_miR_339_5p and hsa_circ_103110; hsa_miR_143_5p and hsa_circ_104689; hsa_miR_409_3p/hsa_miR_153_3p/hsa_miR_145_5p and hsa_circ_104821; hsa_miR_298/hsa_miR_485_3p and hsa_circ_100219; and hsa_miR_100_3p and hsa_circ_406697. MRE sequence analysis for the differentially expressed circRNAs revealed a variety of features boosting MRE efficacy (Supplementary Figure 3).
Table 3: The differentially expressed circRNAs and microRNA response elements
CircRNAs |
Alias (circBase) |
MRE1 |
MRE2 |
MRE3 |
MRE4 |
MRE5 |
|---|---|---|---|---|---|---|
hsa_circ_103110 |
hsa_circ_0004771 |
hsa-miR-653-5p |
hsa-miR-339-5p |
hsa-miR-330-5p |
hsa-miR-595 |
hsa-miR-629-3p |
hsa_circ_104689 |
hsa_circ_0001824 |
hsa-miR-627-3p |
hsa-miR-143-5p |
hsa-miR-656-3p |
hsa-miR-376a-5p |
hsa-miR-222-5p |
hsa_circ_104821 |
hsa_circ_0001875 |
hsa-miR-409-3p |
hsa-miR-153-3p |
hsa-miR-145-5p |
hsa-miR-9-5p |
hsa-miR-152-3p |
hsa_circ_006054 |
hsa_circ_0006054 |
hsa-miR-5001-3p |
hsa-miR-718 |
hsa-miR-4793-3p |
hsa-miR-4524a-5p |
hsa-miR-513a-5p |
hsa_circ_100219 |
hsa_circ_0004619 |
hsa-miR-135b-3p |
hsa-miR-298 |
hsa-miR-485-3p |
hsa-miR-182-5p |
hsa-miR-593-5p |
hsa_circ_406697 |
- |
hsa-miR-6873-3p |
hsa-miR-6833-3p |
hsa-miR-6845-3p |
hsa-miR-6742-5p |
hsa-miR-100-3p |
MRE: microRNA response element.
DISCUSSION
CircRNAs may be generated from exonic or intronic sequences [39] and function as miRNA sponges, regulators of splicing and transcription, and modifiers of parental gene expression [39, 40]. There is evidence that circRNAs are differentially expressed in basal cell carcinoma [27], laryngeal cancer [41] and breast cancer [29, 30]. In the current two-stage epidemiological study, we constructed a profile of differentially expressed circRNAs and explored their involvement in breast cancer. Bioinformatics indicated that specific circRNAs might promote the carcinogenesis of breast cancer by sequestering miRNAs and participating in cancer-related pathways. Our study highlighted the importance of circRNAs in tumorigenesis and suggested that deregulated circRNAs may be diagnostic biomarkers of breast cancer.
In this study, we sequenced six circRNAs of interest following a microarray screening. Among them, hsa_circ_103110, hsa_circ_104689 and hsa_circ_104821 were upregulated in breast cancer tissues. Hsa_circ_103110 is encoded by the NRIP1 (nuclear receptor interacting protein 1) gene, the protein product of which stimulates the transcriptional activity of the estrogen receptor and is critical for promoting the progression and development of breast cancer [42, 43]. Hsa_circ_104689 is spliced from ASAP1 (ArfGAP with SH3 domain, ankyrin repeat and PH domain 1), which encodes an oncoprotein associated with colorectal cancer, laryngeal squamous cell cancer and epithelial ovarian cancer [44–46]. Hsa_circ_104821 is derived from FAM120A (family with sequence similarity 120A), and its encoded protein is a signaling partner that activates the FAK and PI3K pathways in colon cancer metastasis [47]. On the other hand, hsa_circ_006054, hsa_circ_100219 (also known as hsa_circ_0004619) and hsa_circ_406697 were downregulated in breast cancer tissues. Hsa_circ_006054 aligns with the gene KIAA0355, which encodes an uncharacterized protein that may be involved in colorectal carcinogenesis [48]. Hsa_circ_100219 is derived from FAF1 (Fas associated factor 1), which encodes a protein that binds to FAS antigen and initiates or enhances apoptosis. FAF1 also functions as a tumor suppressor, and ectopic FAF1 expression reduces the migration of cancer cells in vitro and invasion/metastasis in vivo [49]. Hsa_circ_100219 is matched with miR-135b [50], while hsa-miR-135b has been found to be up-regulated in cutaneous squamous cell carcinoma [51]. This circRNA had the highest diagnostic accuracy in the current study. Hsa_circ_406697 aligns with the gene RBM22 (RNA binding motif protein 22). RBM22 encodes an RNA binding protein which plays a role in cell division and may be involved in pre-mRNA splicing [52, 53].
GO terms provide proofs of concept for target genes that may regulate crucial biological processes during the development of human diseases. The Hippo signaling pathway has been reported to activate microprocessor which is necessary in mediating the genesis of miRNAs from the primary miRNA transcript, and link cell-density-dependent miRNA biogenesis to cancer [54]. The WNT signaling pathway underlies a wide range of human pathologies and is one of the most canonical cancer-related signaling pathways [55]. The RAP1 signaling pathway is important for both major processes of vessel formation and angiogenesis, while the RAS signaling pathway is a key determinant of the metastatic dissemination of luminal breast cancer [56, 57].
The function of circRNAs remains unclear. An intriguing possibility is that circRNAs act as microRNA sponges. Oncogenic miRNAs like hsa-miR-339-5p, hsa-miR-143-5p, hsa-miR-409-3p, hsa-miR-153-3p and hsa-miR-145-5p have been reported to be downregulated in breast cancer [31–35]. These miRNAs were matched with upregulated circRNAs in our study. Other miRNAs, like hsa-miR-298, hsa-miR-485-3p, and hsa-miR-100, which were matched with downregulated circRNAs in our study, have also been reported to be related to breast cancer [36–38].
Some limitations must be considered in the interpretation of our results. First, due to the relatively low levels of circRNA and the minimum detection thresholds of current methods, the possibility of obtaining false negatives when evaluating circRNA expression cannot be avoided. Second, the sample size was limited and the associations need to be further confirmed. The molecules associated with the present circRNAs, such as miRNAs or proteins, should be experimentally identified and characterized in the future. Third, circulating biomarkers are more acceptable than tissue biomarkers and have greater value in clinical applications. Further studies will be needed to evaluate the diagnostic value of circRNA levels in peripheral blood samples.
In summary, our study provided a profile of circRNAs in breast cancer and adjacent normal-appearing tissues. We discovered that hsa_circ_103110, hsa_circ_104689 and hsa_circ_104821 were upregulated, while hsa_circ_006054, hsa_circ_100219 and hsa_circ_406697 were downregulated in breast cancer tissues. Specific circRNAs are important promoters of carcinogenesis, as they participate in cancer-related pathways and sequester miRNAs, and thus may be useful biomarkers of breast cancer.
MATERIALS AND METHODS
Ethics statement
The Institutional Review Board of Nanjing Medical University (Nanjing, China) approved this study. Written informed consent was obtained from all participants included in the study.
Study design
We designed a two-stage study. First, we used the Arraystar Human circRNA Array V2 (8x15K, Arraystar) to construct a genome-wide circRNA profile. Then, we selected candidate circRNAs for validation using qRT-PCR with a relatively large sample size.
Patients and specimens
We recruited breast cancer patients from the Affiliated Hospital of Jiangsu University, the People's Hospital of Yixing and the First Affiliated Hospital of Suzhou University from March to May 2016. Patients were included if they were: (1) women; (2) with a pathologic diagnosis of breast cancer; (3) without previous cancer history; (4) without HIV/AIDS; (5) >18 years old; (6) having undergone mastectomy; (7) with informed consent. Breast cancer lesions and adjacent normal-appearing tissues were collected from patients who underwent surgical breast resection. The corresponding adjacent normal-appearing tissues were located >5 cm from the edge of the tumors. All patients had no history of radiotherapy or chemotherapy before specimen collection. The specimens were placed in RNA storage solutions (Shanghai Biotechnology Corporation, Shanghai, China) and stored at –80 °C in an ultra-low temperature refrigerator. Tumor stage was determined according to the Classification of Malignant Tumors Staging System (TNM) by the American Joint Committee on Cancer [58].
RNA isolation
Total RNA was isolated with an RNeasy Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. The quality and quantity of RNA were measured with a NanoDrop ND-2000 (Thermo Fisher Scientific, Waltham, Massachusetts, United States). Additionally, RNA integrity was assessed through standard denaturing agarose gel electrophoresis.
CircRNA microarray
The Arraystar Human Circular RNA Microarray V2 (Catalog No: AS-CR-H-V2.0, Arraystar Inc., MD, USA) was used to identify circRNAs with differential expression between breast cancer lesions and adjacent normal-appearing tissues. The array covers 13,617 human circRNAs with stringent experimental support, carefully and comprehensively collected from circRNA studies and landmark publications. Sample labeling and array hybridization were performed according to the manufacturer’s protocol.
qRT-PCR
During the validation stage, we selected 51 breast cancer samples, matched with adjacent normal-appearing tissues. cDNAs were synthesized with the Prime-Script RT reagent kit (TaKaRa, Japan) from 500 ng of total RNA. Subsequently, we performed a SYBR method-based qRT-PCR reaction in a total volume of 10 μL, including 0.2 μL/10 μM forward/reverse primers, 0.2 μL 50× ROX reference dye I, 1 μL cDNA, 5 μL 2× SYBR Premix Ex Taq II and 3.4 μL double-distilled water. The cycling program entailed initiation at 95°C for 30 sec, followed by 40 cycles of 95°C for 5 sec and a pre-selected annealing temperature for 30 sec. The best annealing temperatures were 63°C for hsa_circ_103110, hsa_circ_104689, hsa_circ_104821 and hsa_circ_100219, and 65°C for hsa_circ_006054 and hsa_circ_406697. Results were obtained from three independent wells. The relative expression of each circRNA was calculated from the ΔCt. Divergent primers were designed to amplify the circRNA-specific back-splice junctions (Table 4). We used GAPDH as an internal control. The primers were designed through Primer3 web (http://primer3.ut.ee/), verified through primer-BLAST (https://www.ncbi.nlm.nih.gov/tools/primer-blast/), and synthesized by Realgene (Nanjing, China). The appearance of a single peak in the melting curve of the qRT–PCR indicated the specificity of the PCR results (Supplementary Figure 4).
Table 4: Primers designed for RT-PCR
CircRNA |
Forward (5’-3’) |
Reverse (5’-3’) |
|---|---|---|
hsa_circ_103110 |
CCGGATGACATCAGAGCTACT |
ACACTTCCGTCTGTCTCCAA |
hsa_circ_104689 |
TGGCAGTGAAAAGAAGGGGT |
TGAAAGAAATGTGGCATGTGAGA |
hsa_circ_104821 |
CCACCACATTACTTAGGTTGCA |
CGTTCCGGCTCAGTTTTAGG |
hsa_circ_006054 |
TCCTGTGACTGAAGTGCTGA |
GTCTAGATGTCGCCAGTCCA |
hsa_circ_100219 |
TGCTACAGACGACTCAGAGA |
AGATGATGAAGGTGGTGGCA |
hsa_circ_406697 |
GAGACAGATTTAAGGCCTGCC |
GGTAGATGTGGCTTTCCCCA |
Bioinformatics and data analysis
We performed GO analysis to annotate genes meaningfully in terms of their biological processes, cellular components and molecular functions. The -log10 (P-value) yields an enrichment score representing the significance of GO term enrichment among differentially expressed genes. KEGG analysis was performed to determine the involvement of target genes in different biological pathways. Here, the -log10 (P-value) yields an enrichment score indicating the significance of pathway correlations. To further elucidate the correlations between circRNAs and miRNAs, we predicted circRNA/miRNA interactions using miRNA target prediction software from Arraystar, which refers to TargetScan (http://www.targetscan.org/) and miRanda (http://www.microrna.org/).
Statistical analysis
The fold-change of each circRNA was computed from the profile difference between the cancer and control groups, and the significance was analyzed with a t-test. A receiver operating characteristic curve was plotted, and the AUC, sensitivity and specificity were calculated to assess the ability of circRNAs to differentiate between breast cancer and adjacent normal-appearing tissues. Statistical analyses were performed with R software version 3.3.1 (https://www.r-project.org/) and GraphPad Prism 5 (GraphPad Software, La Jolla, CA).
CONFLICTS OF INTEREST
The authors declare that they have no competing interest.
FUNDING
The present study was supported by the National Natural Science Foundation of China (81172268), the Social Development Project in Jiangsu Province (BE2015694), the Scientific Research Innovation Project for Graduate Students in Jiangsu Province (KYLX16_1125), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAAD). The funding agencies had no role in the study design, data collection, analysis, decision to publish, or preparation of the manuscript.
REFERENCES
1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015; 65:87-108. doi: 10.3322/caac.21262.
2. Guo Y, Warren Andersen S, Shu XO, Michailidou K, Bolla MK, Wang Q, Garcia-Closas M, Milne RL, Schmidt MK, Chang-Claude J, Dunning A, Bojesen SE, Ahsan H, et al. Genetically Predicted Body Mass Index and Breast Cancer Risk: Mendelian Randomization Analyses of Data from 145,000 Women of European Descent. PLoS Med. 2016; 13:e1002105. doi: 10.1371/journal.pmed.1002105.
3. Lambertini M, Santoro L, Del Mastro L, Nguyen B, Livraghi L, Ugolini D, Peccatori FA, Azim HA Jr. Reproductive behaviors and risk of developing breast cancer according to tumor subtype: A systematic review and meta-analysis of epidemiological studies. Cancer Treat Rev. 2016; 49:65-76. doi: 10.1016/j.ctrv.2016.07.006.
4. Chlebowski RT, Manson JE, Anderson GL, Cauley JA, Aragaki AK, Stefanick ML, Lane DS, Johnson KC, Wactawski-Wende J, Chen C, Qi L, Yasmeen S, Newcomb PA, Prentice RL. Estrogen plus progestin and breast cancer incidence and mortality in the Women's Health Initiative Observational Study. J Natl Cancer Inst. 2013; 105:526-35. doi: 10.1093/jnci/djt043.
5. Rice MS, Eliassen AH, Hankinson SE, Lenart EB, Willett WC, Tamimi RM. Breast Cancer Research in the Nurses' Health Studies: Exposures Across the Life Course. Am J Public Health. 2016; 106:1592-8. doi: 10.2105/AJPH.2016.303325.
6. Zeng C, Guo X, Long J, Kuchenbaecker KB, Droit A, Michailidou K, Ghoussaini M, Kar S, Freeman A, Hopper JL, Milne RL, Bolla MK, Wang Q, et al. Identification of independent association signals and putative functional variants for breast cancer risk through fine-scale mapping of the 12p11 locus. Breast Cancer Res. 2016; 18:64. doi: 10.1186/s13058-016-0718-0.
7. Zhang B, Beeghly-Fadiel A, Long J, Zheng W. Genetic variants associated with breast-cancer risk: comprehensive research synopsis, meta-analysis, and epidemiological evidence. Lancet Oncol. 2011; 12:477-88. doi: 10.1016/S1470-2045(11)70076-6.
8. Karsli-Ceppioglu S, Dagdemir A, Judes G, Ngollo M, Penault-Llorca F, Pajon A, Bignon YJ, Bernard-Gallon D. Epigenetic mechanisms of breast cancer: an update of the current knowledge. Epigenomics. 2014; 6:651-64. doi: 10.2217/epi.14.59.
9. Wu L, Shen Y, Peng X, Zhang S, Wang M, Xu G, Zheng X, Wang J, Lu C. Aberrant promoter methylation of cancer-related genes in human breast cancer. Oncol Lett. 2016; 12:5145-55. doi: 10.3892/ol.2016.5351.
10. Wu L, Wang F, Xu R, Zhang S, Peng X, Feng Y, Wang J, Lu C. Promoter methylation of BRCA1 in the prognosis of breast cancer: a meta-analysis. Breast Cancer Res Treat. 2013; 142:619-27. doi: 10.1007/s10549-013-2774-9.
11. Venkatesh T, Suresh PS, Tsutsumi R. Non-coding RNAs: Functions and applications in endocrine-related cancer. Mol Cell Endocrinol. 2015; 416:88-96. doi: 10.1016/j.mce.2015.08.026.
12. Huang T, Alvarez A, Hu B, Cheng SY. Noncoding RNAs in cancer and cancer stem cells. Chin J Cancer. 2013; 32:582-93. doi: 10.5732/cjc.013.10170.
13. Mulrane L, McGee SF, Gallagher WM, O'Connor DP. miRNA dysregulation in breast cancer. Cancer Res. 2013; 73:6554-62. doi: 10.1158/0008-5472.CAN-13-1841.
14. Sidiropoulos KG, Ding Q, Pampalakis G, White NM, Boulos P, Sotiropoulou G, Yousef GM. KLK6-regulated miRNA networks activate oncogenic pathways in breast cancer subtypes. Mol Oncol. 2016; 10:993-1007. doi: 10.1016/j.molonc.2016.03.008.
15. Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA, Downing JR, Jacks T, Horvitz HR, Golub TR. MicroRNA expression profiles classify human cancers. Nature. 2005; 435:834-8. doi: 10.1038/nature03702.
16. Hannafon BN, Trigoso YD, Calloway CL, Zhao YD, Lum DH, Welm AL, Zhao ZJ, Blick KE, Dooley WC, Ding WQ. Plasma exosome microRNAs are indicative of breast cancer. Breast Cancer Res. 2016; 18:90. doi: 10.1186/s13058-016-0753-x.
17. Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer M, Loewer A, Ziebold U, Landthaler M, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013; 495:333-8. doi: 10.1038/nature11928.
18. Sanger HL, Klotz G, Riesner D, Gross HJ, Kleinschmidt AK. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc Natl Acad Sci U S A. 1976; 73:3852-6. doi:
19. Capel B, Swain A, Nicolis S, Hacker A, Walter M, Koopman P, Goodfellow P, Lovell-Badge R. Circular transcripts of the testis-determining gene Sry in adult mouse testis. Cell. 1993; 73:1019-30. doi:
20. Jeck WR, Sharpless NE. Detecting and characterizing circular RNAs. Nat Biotechnol. 2014; 32:453-61. doi: 10.1038/nbt.2890.
21. Cheng J, Metge F, Dieterich C. Specific identification and quantification of circular RNAs from sequencing data. Bioinformatics. 2016; 32:1094-6. doi: 10.1093/bioinformatics/btv656.
22. Memczak S, Papavasileiou P, Peters O, Rajewsky N. Identification and Characterization of Circular RNAs As a New Class of Putative Biomarkers in Human Blood. PLoS One. 2015; 10:e0141214. doi: 10.1371/journal.pone.0141214.
23. Guo JU, Agarwal V, Guo H, Bartel DP. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014; 15:409. doi: 10.1186/s13059-014-0409-z.
24. Hansen TB, Jensen TI, Clausen BH, Bramsen JB, Finsen B, Damgaard CK, Kjems J. Natural RNA circles function as efficient microRNA sponges. Nature. 2013; 495:384-8. doi: 10.1038/nature11993.
25. Xu H, Guo S, Li W, Yu P. The circular RNA Cdr1as, via miR-7 and its targets, regulates insulin transcription and secretion in islet cells. Sci Rep. 2015; 5:12453. doi: 10.1038/srep12453.
26. Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J, Chen D, Gu J, He X, Huang S. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell Res. 2015; 25:981-4. doi: 10.1038/cr.2015.82.
27. Sand M, Bechara FG, Sand D, Gambichler T, Hahn SA, Bromba M, Stockfleth E, Hessam S. Circular RNA expression in basal cell carcinoma. Epigenomics. 2016; 8:619-32. doi: 10.2217/epi-2015-0019.
28. Huang M, Zhong Z, Lv M, Shu J, Tian Q, Chen J. Comprehensive analysis of differentially expressed profiles of lncRNAs and circRNAs with associated co-expression and ceRNA networks in bladder carcinoma. Oncotarget. 2016; 7:47186-200. doi: 10.18632/oncotarget.9706.
29. Galasso M, Costantino G, Pasquali L, Minotti L, Baldassari F, Corra F, Agnoletto C, Volinia S. Profiling of the Predicted Circular RNAs in Ductal In Situ and Invasive Breast Cancer: A Pilot Study. Int J Genomics. 2016; 2016:4503840. doi: 10.1155/2016/4503840.
30. Nair AA, Niu N, Tang X, Thompson KJ, Wang L, Kocher JP, Subramanian S, Kalari KR. Circular RNAs and their associations with breast cancer subtypes. Oncotarget. 2016; 7:80967-79. doi: 10.18632/oncotarget.13134.
31. Wu ZS, Wu Q, Wang CQ, Wang XN, Wang Y, Zhao JJ, Mao SS, Zhang GH, Zhang N, Xu XC. MiR-339-5p inhibits breast cancer cell migration and invasion in vitro and may be a potential biomarker for breast cancer prognosis. BMC Cancer. 2010; 10:542. doi: 10.1186/1471-2407-10-542.
32. Ng EK, Li R, Shin VY, Siu JM, Ma ES, Kwong A. MicroRNA-143 is downregulated in breast cancer and regulates DNA methyltransferases 3A in breast cancer cells. Tumour Biol. 2014; 35:2591-8. doi: 10.1007/s13277-013-1341-7.
33. Zhang G, Liu Z, Xu H, Yang Q. miR-409-3p suppresses breast cancer cell growth and invasion by targeting Akt1. Biochem Biophys Res Commun. 2016; 469:189-95. doi: 10.1016/j.bbrc.2015.11.099.
34. Li W, Zhai L, Zhao C, Lv S. MiR-153 inhibits epithelial-mesenchymal transition by targeting metadherin in human breast cancer. Breast Cancer Res Treat. 2015; 150:501-9. doi: 10.1007/s10549-015-3346-y.
35. Chang YY, Kuo WH, Hung JH, Lee CY, Lee YH, Chang YC, Lin WC, Shen CY, Huang CS, Hsieh FJ, Lai LC, Tsai MH, Chang KJ, Chuang EY. Deregulated microRNAs in triple-negative breast cancer revealed by deep sequencing. Mol Cancer. 2015; 14:36. doi: 10.1186/s12943-015-0301-9.
36. Bao L, Hazari S, Mehra S, Kaushal D, Moroz K, Dash S. Increased expression of P-glycoprotein and doxorubicin chemoresistance of metastatic breast cancer is regulated by miR-298. Am J Pathol. 2012; 180:2490-503. doi: 10.1016/j.ajpath.2012.02.024.
37. Anaya-Ruiz M, Bandala C, Perez-Santos JL. miR-485 Acts as a Tumor Suppressor by Inhibiting Cell Growth and Migration in Breast Carcinoma T47D Cells. Asian Pacific Journal of Cancer Prevention. 2013; 14:3757-60. doi: 10.7314/Apjcp.2013.14.6.3757.
38. Deng L, Shang L, Bai S, Chen J, He X, Martin-Trevino R, Chen S, Li XY, Meng X, Yu B, Wang X, Liu Y, McDermott SP, et al. MicroRNA100 inhibits self-renewal of breast cancer stem-like cells and breast tumor development. Cancer Res. 2014; 74:6648-60. doi: 10.1158/0008-5472.CAN-13-3710.
39. Qu S, Yang X, Li X, Wang J, Gao Y, Shang R, Sun W, Dou K, Li H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015; 365:141-8. doi: 10.1016/j.canlet.2015.06.003.
40. Qian Y, Lu Y, Rui C, Qian Y, Cai M, Jia R. Potential Significance of Circular RNA in Human Placental Tissue for Patients with Preeclampsia. Cell Physiol Biochem. 2016; 39:1380-90. doi: 10.1159/000447842.
41. Xuan L, Qu L, Zhou H, Wang P, Yu H, Wu T, Wang X, Li Q, Tian L, Liu M, Sun Y. Circular RNA: a novel biomarker for progressive laryngeal cancer. Am J Transl Res. 2016; 8:932-9. doi:
42. Aziz MH, Chen X, Zhang Q, DeFrain C, Osland J, Luo Y, Shi X, Yuan R. Suppressing NRIP1 inhibits growth of breast cancer cells in vitro and in vivo. Oncotarget. 2015; 6:39714-24. doi: 10.18632/oncotarget.5356.
43. Augereau P, Badia E, Fuentes M, Rabenoelina F, Corniou M, Derocq D, Balaguer P, Cavailles V. Transcriptional regulation of the human NRIP1/RIP140 gene by estrogen is modulated by dioxin signalling. Mol Pharmacol. 2006; 69:1338-46. doi: 10.1124/mol.105.017376.
44. Muller T, Stein U, Poletti A, Garzia L, Rothley M, Plaumann D, Thiele W, Bauer M, Galasso A, Schlag P, Pankratz M, Zollo M, Sleeman JP. ASAP1 promotes tumor cell motility and invasiveness, stimulates metastasis formation in vivo, and correlates with poor survival in colorectal cancer patients. Oncogene. 2010; 29:2393-403. doi: 10.1038/onc.2010.6.
45. Li M, Tian L, Yao H, Lu J, Ge J, Guo Y, Liu M, Xiao H. ASAP1 mediates the invasive phenotype of human laryngeal squamous cell carcinoma to affect survival prognosis. Oncol Rep. 2014; 31:2676-82. doi: 10.3892/or.2014.3150.
46. Hou T, Yang C, Tong C, Zhang H, Xiao J, Li J. Overexpression of ASAP1 is associated with poor prognosis in epithelial ovarian cancer. Int J Clin Exp Pathol. 2014; 7:280-7. doi:
47. Bartolome RA, Garcia-Palmero I, Torres S, Lopez-Lucendo M, Balyasnikova IV, Casal JI. IL13 Receptor alpha2 Signaling Requires a Scaffold Protein, FAM120A, to Activate the FAK and PI3K Pathways in Colon Cancer Metastasis. Cancer Res. 2015; 75:2434-44. doi: 10.1158/0008-5472.CAN-14-3650.
48. Chadwick RB, Jiang GL, Bennington GA, Yuan B, Johnson CK, Stevens MW, Niemann TH, Peltomaki P, Huang S, de la Chapelle A. Candidate tumor suppressor RIZ is frequently involved in colorectal carcinogenesis. Proc Natl Acad Sci U S A. 2000; 97:2662-7. doi: 10.1073/pnas.040579497.
49. Zhang L, Zhou F, Li Y, Drabsch Y, Zhang J, van Dam H, ten Dijke P. Fas-associated factor 1 is a scaffold protein that promotes beta-transducin repeat-containing protein (beta-TrCP)-mediated beta-catenin ubiquitination and degradation. J Biol Chem. 2012; 287:30701-10. doi: 10.1074/jbc.M112.353524.
50. Sand M, Bechara FG, Gambichler T, Sand D, Bromba M, Hahn SA, Stockfleth E, Hessam S. Circular RNA expression in cutaneous squamous cell carcinoma. J Dermatol Sci. 2016; 83:210-8. doi: 10.1016/j.jdermsci.2016.05.012.
51. Sand M, Skrygan M, Georgas D, Sand D, Hahn SA, Gambichler T, Altmeyer P, Bechara FG. Microarray analysis of microRNA expression in cutaneous squamous cell carcinoma. J Dermatol Sci. 2012; 68:119-26. doi: 10.1016/j.jdermsci.2012.09.004.
52. Rasche N, Dybkov O, Schmitzova J, Akyildiz B, Fabrizio P, Luhrmann R. Cwc2 and its human homologue RBM22 promote an active conformation of the spliceosome catalytic centre. EMBO J. 2012; 31:1591-604. doi: 10.1038/emboj.2011.502.
53. He F, Wang CT, Gou LT. RNA-binding motif protein RBM22 is required for normal development of zebrafish embryos. Genet Mol Res. 2009; 8:1466-73. doi: 10.4238/vol8-4gmr683.
54. Mori M, Triboulet R, Mohseni M, Schlegelmilch K, Shrestha K, Camargo FD, Gregory RI. Hippo signaling regulates microprocessor and links cell-density-dependent miRNA biogenesis to cancer. Cell. 2014; 156:893-906. doi: 10.1016/j.cell.2013.12.043.
55. Clevers H, Nusse R. Wnt/beta-catenin signaling and disease. Cell. 2012; 149:1192-205. doi: 10.1016/j.cell.2012.05.012.
56. Chrzanowska-Wodnicka M. Distinct functions for Rap1 signaling in vascular morphogenesis and dysfunction. Exp Cell Res. 2013; 319:2350-9. doi: 10.1016/j.yexcr.2013.07.022.
57. Wright KL, Adams JR, Liu JC, Loch AJ, Wong RG, Jo CE, Beck LA, Santhanam DR, Weiss L, Mei X, Lane TF, Koralov SB, Done SJ, et al. Ras Signaling Is a Key Determinant for Metastatic Dissemination and Poor Survival of Luminal Breast Cancer Patients. Cancer Res. 2015; 75:4960-72. doi: 10.1158/0008-5472.CAN-14-2992.
58. Edge SB, Compton CC. The American Joint Committee on Cancer: the 7th edition of the AJCC cancer staging manual and the future of TNM. Ann Surg Oncol. 2010; 17:1471-4. doi: 10.1245/s10434-010-0985-4.