
INTRODUCTION
Foxp3+CD4+ regulatory T cells (Tregs) play a crucial role in the maintenance of immune homeostasis and prevention of excessive immune responses [1–3]. Tregs comprise two subsets with two distinct origins and functions: thymus-derived Tregs (tTregs) and peripherally-induced Tregs (pTregs) that are generated from naïve CD4+ T cells by numbers of triggers in the peripheral organs such as gut, skin and lung [4]. Additionally, in the presence of IL-2 and TGF-β1, the conversion from CD25- Foxp3- CD4+ T cells into induced Foxp3+ Tregs (iTregs) can be achieved in in vitro T cell cultures [5]. However, these Tregs are less stable and can lose their Foxp3 expression and suppressive activity after the transfer into mice. Previously, it was shown that DNA methylation status at the Foxp3 locus is crucial for maintaining a stable Treg lineage. The instable Foxp3 expression by in vitro generated iTregs is associated with decreased demethylation of the Treg-specific demethylated region (TSDR) at the Foxp3 locus, which is highly demethylated within tTreg and pTreg subsets [6–8]. Interestingly, mice with a deletion in TGF-β1 or animals lacking specifically Smad2 and Smad3 in T cells have been show to exhibit normal development of tTregs but have significantly decreased numbers of pTregs [9, 10], suggesting that TGF-β signalling pathway is essential for the maintenance of Foxp3 expression and function of Tregs in peripheral organs. Although the presence of TGF-β1 in the gut is essential for optimal pTreg development, recent findings suggest that some exogenous factors such as gut microbiota-derived short chain fatty acids (SCFAs) facilitate expansion of colonic pTregs by enhancing the acetylation of histone H3 at the Foxp3 locus [11, 12]. On the other hand, the activation of classical NF-κB signalling pathway has been shown to be crucial for the development of Tregs in the thymus [13–15]. Especially, the Treg cell-intrinsic expression of c-Rel is essential for the thymic induction of Foxp3 [16–19]. Similarly, the atypical inhibitor of NF-κB IκBNS is also involved in the development of Tregs by regulating the transition of thymic immature Treg precursors into mature Tregs [20]. Furthermore, mice lacking proteins involved in NF-κB activation such as PKCΘ, Bcl10, CARMA1 and MALT1 display impaired Treg cell development [14, 21]. These data revealed that NF-κB, although originally described as a pro-inflammatory factor, is one of key regulators of development of tTregs with anti-inflammatory properties. However, c-Rel deficient mice still exhibit moderate Foxp3+ Treg cell frequencies in the peripheral organs prompting us to investigate the origin of these cells.
The selective in vivo expansion of Tregs has the potential to treat autoimmune diseases. Recently, it has been shown that the treatment of mice with IL-2/anti-IL-2 complex, generated by using JES6-1 antibody, increases the number of Tregs and protects from asthma, experimental autoimmune encephalomyelitis (EAE) and type 1 diabetes [22–24]. These results highlight the central role for IL-2 and Tregs in efficient treatment of autoimmune diseases. The members of Ikaros family of transcription factors, Helios and Eos appear to be specifically expressed in Tregs [25, 26]. A recent study has described Eos as an important factor that directly interacts with Foxp3 and induces chromatin modifications resulting in specific gene silencing in Tregs [26]. Helios has been found to be preferentially expressed in Tregs of thymic origin, thus potentially acting as a crucial marker for the discrimination between tTregs and pTregs. Its expression in T cells does not require the activity of Foxp3, as Helios has also been detected in the subsets of DN2-DN4 thymocytes [25].
Here, we show that c-Rel deficient mice have significantly less Helios+Foxp3+ but not Helios-Foxp3+ Tregs. Interestingly, only Helios+Foxp3+ Tregs were specifically enriched after treatment of WT and even rel-/-mice with IL-2/anti-IL-2 mAb. Surprisingly, we found a strong accumulation of pTregs in aged rel-/- mice. Furthermore, the expansion of mucosal Tregs during acute colitis as well as normal generation of iTregs in the absence of c-Rel indicate that this transcription factor is essential for the development of tTregs but not for that of pTregs and iTregs.
RESULTS
In vivo administration of the IL-2/JES6-1 complex induces expansion of Tregs in c-Rel deficient mice
IL-2 plays an essential role for the thymic development of Tregs and the maintenance of Treg homeostasis in the peripheral tissues [27]. Defective IL-2 production in c-Rel deficient mice was reported previously [28]. Here, we confirm that CD4+ T cells isolated from rel-/- mice and stimulated for 24 h with anti-CD3/anti-CD28 antibodies secrete significantly less IL-2 than their wild-type (WT) counterparts (Figure 1A). Recent studies have shown that Tregs selectively expand after treatment of mice with IL-2 and anti-IL-2 mAb JES6-1 [22, 23]. To analyse whether exogenous in vivo supply of IL-2 can reverse Treg-intrinsic requirements for c-Rel, we administered IL-2/JES6-1 complex in WT and rel-/- animals. Consistent with earlier reports, we found that biological activity of IL-2 is greatly enhanced by association with JES6-1 mAb. When compared to the PBS-treated WT mice, there was a significant increase of Tregs in spleen and mesenteric lymph nodes (mLN) after treatment with IL-2/JES6-1. The proportion of Foxp3+ Tregs within CD4+ T cell population rose to 30-50 % in these organs as compared to basic level of 10-15 % in PBS-treated animals. Remarkably, the injection of IL-2/JES6-1 also resulted in a strong expansion of Tregs in rel-/- mice (Figure 1B and 1C). The substantial increase in frequencies of Tregs was also reflected in increased total Treg cell numbers in examined organs (Figure 1D). Kinetic studies demonstrated that three injections of the IL-2/JES6-1 complex resulted in a brief expansion of Tregs which peaked on day 5 and gradually declined on days 8 and 15. This finding refers to both WT and rel-/- mice, although the Treg expansion in c-Rel deficient mice did not reach the percentage and total cell numbers of WT Tregs (Figure 1E). In conclusion, we found that the short-course treatment of mice with IL-2/JES6-1 led to the substantial expansion of Tregs not only in WT but also in rel-/- mice, which normally exhibit reduced Treg frequencies and numbers. Taken together, our results demonstrate that signals derived by exogenous IL-2 can partially compensate for defects in c-Rel deficient Tregs.
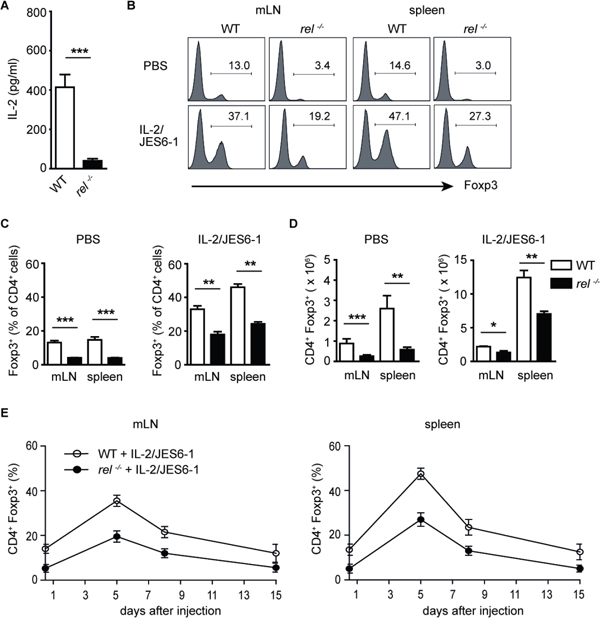
Figure 1: In vivo expansion of Foxp3+ Tregs with IL-2/JES6-1 complex in rel-/- mice. (A) CD4+ T cells purified from wild-type (WT) and rel-/- mice were stimulated with anti-CD3 plus anti-CD28 for 24h. IL-2 secretion was measured by ELISA. Data are means ± SEM; ***P<0.001. (B) WT and rel-/- mice were treated i.p. daily for three days (days 0, 1, 2) with IL-2/JES6-1. The expansion of Tregs was analyzed on day 5 by the intracellular staining of Foxp3 in mLN and spleen. The histograms are gated on CD4+ T cells. Data are representative of two independent experiments. (C) and (D) The percentage (C) and total cell number (D) of Tregs in mLN and spleen on day 5 after treatment of mice with IL-2/JES6-1 (n = 6 mice per group). Results are means ± SEM; *P<0.05, **P<0.01, ***P<0.001. (E) A kinetic analysis of Treg expansion after IL-2/JES6-1 treatment of WT and rel-/- mice. Foxp3 frequencies among CD4+ T cells on days 0, 5, 8 and 15 following three injections of IL-2/ JES6-1 are shown (n = 6 per group). Error bars represent SEM.
Preferential expansion of Helios+Foxp3+ Tregs after IL-2/JES6-1 treatment
Previously, it was shown that administration of the IL-2/JES6-1 stabilized the Foxp3 expression and expanded the numbers of transferred iTregs [29]. Although IL-2 administration has been extensively used to boost Treg cell numbers in vivo, it is still unknown whether IL-2 specifically expands tTregs or pTregs in the absence of antigen challenge. To investigate if both Treg subsets, tTregs and pTregs, can be expanded by exogenous IL-2, the percentage of Helios+Foxp3+ and Helios-Foxp3+ cells was compared among CD4+ T cells in lymphoid tissues of WT and rel-/- mice after IL-2/JES6-1 administration. Surprisingly, the treatment with IL-2/JES6-1 induced strong tTreg but not pTreg cell expansion in spleen, mLN and inguinal (i) LN at day 5 in WT mice (Figure 2A and 2B). Remarkably, this effect was even more pronounced in c-Rel deficient mice. In fact, in the spleen of IL-2/JES6-1-treated rel-/- mice, approximately a 15-fold increase in the frequency of Helios+Foxp3+ Tregs was found as compared with naïve animals (Figure 2A and 2C). Furthermore, the significant reduction of Helios+Foxp3+ but not of Helios-Foxp3+ Treg population was observed in naïve rel-/- mice, confirming that c-Rel is indeed a crucial factor required for thymic differentiation of Tregs (Figure 2A).
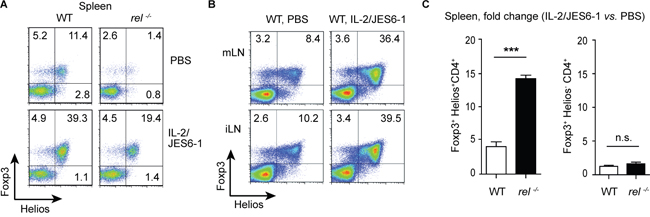
Figure 2: Rapid expansion of Helios+Foxp3+ Tregs after IL-2/JES6-1 treatment. (A) The effect of the IL-2/JES6-1 complex on expansion of splenic Helios+Foxp3+ and Helios-Foxp3+ Tregs was analyzed on day 5 after treatment. Data are representative of three independent experiments. (B) IL-2/JES6-1-mediated expansion of Helios+Foxp3+ Tregs in mLN and inguinal (i) LN on day 5 after treatment of WT mice. A representative of three similar experiments is shown. The dot plots (A and B) are gated on CD4+ T cells. (C) Data from (A) are shown as a fold change for IL-2/JES6-1-expanded splenic Tregs as compared with Tregs from PBS-treated mice. Results are means ± SEM; n.s., not significant, ***P<0.001.
The unexpected finding that only Helios+Foxp3+ Tregs robustly proliferate and respond to the supplementation with exogenous IL-2 prompted us to investigate the possible reason for their predominant expansion. IL-2 complexed to JES6-1 has been described to preferentially activate cells that express CD25 and co-injection of CD25-depleting PC61 mAb was able to block Treg cell expansion [23]. Firstly, the treatment of WT mice with CD25-depleting Ab (two i.p. injections of PC61 mAb) targeted both Treg subsets as Foxp3 expression was substantially reduced in Helios+ and Helios- Treg populations as compared to control mice (Supplementary Figure 1A). However, when we compared the expression of CD25 for each Treg population, we found that the CD25 expression levels were higher in Helios+ Treg than in Helios- Treg population, which suggests that Helios+ Treg subset might have a proliferative advantage by responding rapidly to lower IL-2 concentrations (Supplementary Figure 1B). These data are in agreement with a very recent study that shows the preferential expansion of human Helios+ Tregs after treatment of patients with low dose IL-2 [30].
c-Rel is dispensable for the expansion of mucosal Tregs in acute colitis
We next performed a histological and flow cytometry analysis of Foxp3 expression in the small intestine and colon of WT and rel-/- mice. As expected, intestinal Tregs were reduced in c-Rel deficient animals (Figure 3A-3C). Germ-free (GF) mice have also reduced numbers of colonic Tregs that selectively expand after colonization with commensal bacteria [31, 32]. Here, we show that the percentages of colonic Helios-Foxp3+CD4+ cells were almost 5 times lower in GF than in conventional mice, while the percentage of Helios+Foxp3+ subset was unchanged (Supplementary Figure 2A and 2B). Therefore, it is tempting to speculate that, in accordance with the original report [25], the most Helios-Foxp3+ Tregs are indeed generated in the peripheral tissues.
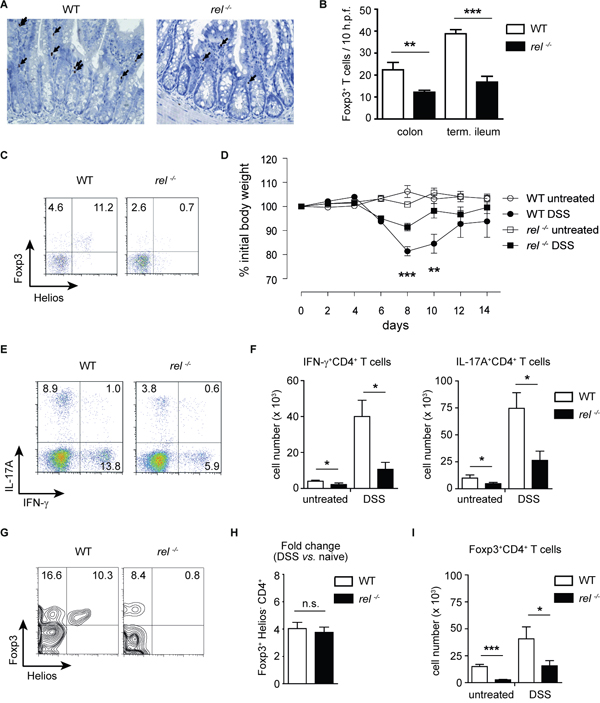
Figure 3: Expansion of mucosal Tregs in rel-/- mice after induction of colitis. (A) and (B) Colons and small intestines from WT and rel-/- mice were stained for Foxp3 (indicated by arrows) using immunohistochemistry analysis (magnification: 200 x). Data are shown as a number of Foxp3+ cells/ 10 h.p.f. (high power fields). Number of animals: 4-5 mice per group. Data are means ± SEM; **P<0.01, ***P<0.001. (C) Frequencies of Helios+Foxp3+ and Helios-Foxp3+ Treg cells in the colon of naïve WT and rel-/- mice. A representative of three similar experiments is shown. (D) Colitis was induced in WT and rel-/- mice by orally administering 3 % DSS for 5 days and the change of initial weight was monitored over a period of 14 days. Data represent mean ± SEM (n = 8 mice per group). **P<0.01, ***P<0.001. (E) and (F) Frequency (E) and total cell number (F) of colonic IFN-γ+ and IL-17A+ CD4+ T cells in untreated and DSS-treated WT and rel-/- mice, measured by FACS analysis on day 10 after colitis induction. Data in (F) are displayed as means ± SEM; *P<0.05. (G) and (H) The percentage of Helios-Foxp3+ Tregs within colonic CD4+ T cells after colitis induction by 3 % DSS. Results in (H) are displayed as fold change as compared to naïve mice. n.s., not significant. (I) Absolute cell numbers of colonic Tregs in untreated and DSS-treated WT and rel-/- mice. Results are shown as means ± SEM; *P<0.05, ***P<0.001.
To analyse the impact of c-Rel on the expansion of mucosal pTregs, we induced acute colitis in WT and rel-/- animals and tested whether the accumulation of pTregs at gut mucosal sites is dependent on this transcription factor. Following oral administration of dextran sodium sulfate (DSS), the measurement of the loss of initial body weight and colon length revealed a milder disease course in rel-/- mice as compared to WT animals (Figure 3D, and Supplementary Figure 3A). In our previous study, we found no reduction in expression of effector cytokines by in vitro generated Th17 and Th1 cells [18]. In contrast to these data, on day 10 after colitis induction, a decreased production of IL-17A and IFN-γ by colonic CD4+ T cells was detected in rel-/- mice in comparison to WT animals (Figure 3E and 3F). This discrepancy could be attributed to reduced colonic secretion of proinflammatory cytokines such as IL-12, IL-23, IL-6 and IL-1β (which are known to considerably impact on T cell differentiation) in DSS-treated c-Rel deficient mice (Supplementary Figure 3B and 3C). Notably, the FACS analysis revealed a dispensable role for c-Rel for the expansion of colonic Helios-Foxp3+ pTregs, as in both, DSS-treated WT and rel-/- mice, the frequency of these cells increased 4-fold as compared to naïve mice (Figure 3C, 3G and 3H). Similarly, the absolute number of colonic Tregs increased during the progression of colitis in both WT and rel-/- animals (Figure 3I).
Selective accumulation of Helios-Foxp3+ Tregs in aged c-Rel deficient mice
During the aging, the immune system undergoes several changes which are not beneficial for the host, including age-related Treg accumulation and suppressed immune phenotype [33]. A recent study has demonstrated that, in addition to the expansion of Helios-Foxp3+ pTregs, also tTregs with high expression of Helios and Neuropilin 1 (Nrp1) accumulate in aged mice. Furthermore, the aged Tregs were shown to be functional and had a broad Vβ repertoire [34]. In accordance with previous data, we found increased proportion of Tregs in all investigated tissues in Foxp3-RFP reporter mice (Figure 4A). Moreover, we confirmed that the frequency of both Helios-Foxp3+ and Helios+Foxp3+ Tregs was increased in aged WT mice as compared to young animals (Figure 4B). Surprisingly, by analysing the Treg subsets in young and old c-Rel deficient mice, we found that in the absence of this transcription factor, the aged Tregs were enriched only in Helios- population (Figure 4B-4D). Taken together, the surprising observation of a substantial increase of pTregs in c-Rel deficient aged mice implies that, in contrary to its crucial role in thymic Treg development, the transcription factor c-Rel is not required for the accumulation of pTregs.
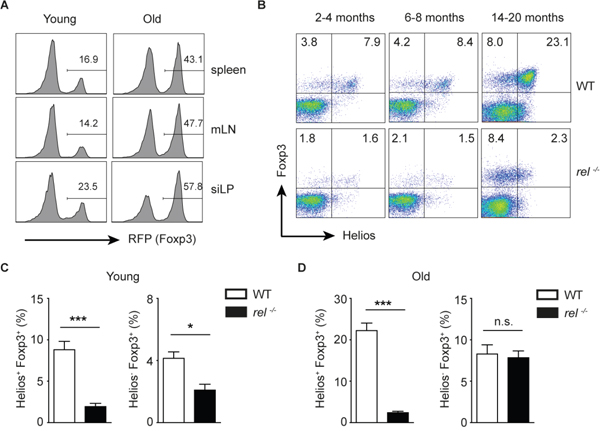
Figure 4: Preferential accumulation of Helios-Foxp3+ Tregs in aged rel-/- mice. (A) The frequency of RFP+ cells within CD4+ T cells in spleen, mLN and lamina propria of the small intestine (siLP) in naïve young (8-14 weeks) and old (14-20 months) Foxp3-RFP reporter mice. The cells are gated on CD4+ gate. A representative of three experiments is shown. (B-D) Young (2-4 months), middle-aged (6-8 moths) and old (14-20 months) naive WT and rel-/- mice were analyzed for their Helios and Foxp3 expression within splenic CD4+ T cells. A representative of two experiments is shown within CD4 gate (B). Data (C and D) are frequencies of indicated cells within CD4+ T cell population and are displayed as means ± SEM; n.s., not significant, *P<0.05, ***P<0.001.
c-Rel is not required for the differentiation of iTregs in the presence of exogenous IL-2
It remains obscure whether the observed defective cell numbers of Tregs in peripheral organs of rel-/- mice are only due to the lack of the thymic activity of c-Rel. We here show that the stimulation of CD3 and co-stimulatory molecule CD28 with anti-CD3/anti-CD28 for 24 hours is sufficient to increase c-Rel expression and translocate this transcription factor into the nucleus of activated CD4+ T cells (Figure 5A). To investigate the impact of c-Rel on the generation of iTregs, we isolated naïve CD4+ T cells from spleen and LN of WT and c-Rel deficient mice and cultured them in the presence of TGF-β1 and IL-2. Activation of STAT5 upon binding of IL-2 is necessary for subsequent proliferation and survival of Tregs (23). We thus wondered whether this signalling pathway is impaired in T cells lacking c-Rel. In the western blot analysis, a normal phosphorylation of STAT5 was observed in both, WT and c-Rel deficient CD4+ T cells treated with IL-2. Thus, although a partial defect in CD25 expression was previously described for rel-/- T cells [28], CD4+ T cells from rel-/- mice are able to sense the exogenous IL-2 and to response to it by activating STAT5 signalling pathway (Figure 5B and Supplementary Figure 4). Moreover, we were not able to find any defective expression of Foxp3 in T cell cultures lacking c-Rel as compared to WT cells under optimal iTreg-inducing conditions (Figure 5C and 5D). On the other hand, we confirm a defective Treg development in rel-/- thymocytes (Figure 5E), which was previously observed by several groups [16–18]. Together, the transcription factor c-Rel, although essential for tTreg generation, is dispensable for in vitro conversion of naïve CD4+ T cells into Foxp3+ iTregs in the presence of exogenous IL-2.
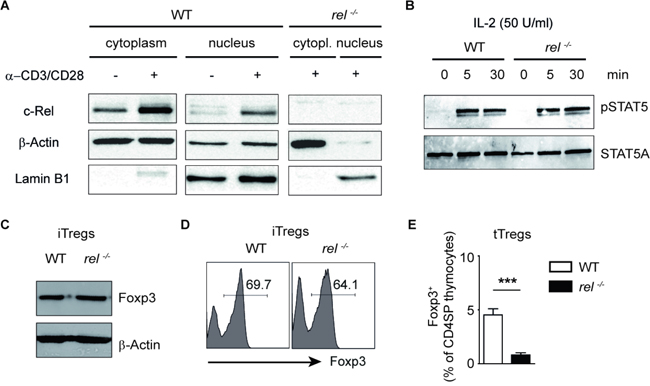
Figure 5: Normal in vitro generation of Foxp3+ Tregs in the absence of c-Rel. (A) Western blot analysis of c-Rel expression in cytoplasmic and nuclear extracts of WT and rel-/- CD4+ T cells after activation with plate-bound anti-CD3 and soluble anti-CD28 antibodies for 24 hours. Lamin B1 and β-Actin served as loading controls. (B) Western blot analysis of naïve CD4+ T cells purified from spleens of WT and rel-/- mice and stimulated with IL-2 (50 U/ml) for indicated time points. Phosphorylation of STAT5 was analyzed by using anti-phospho-STAT5 and anti-STAT5A antibodies. Three similar experiments were performed. (C) Immunoblot analysis for Foxp3 and β-Actin was performed on day three after culturing the purified naïve CD4+ T cells under optimal Treg-inducing conditions (2 ng/ml TGF-β1 and 50 U/ml IL-2). A representative of two experiments is shown. (D) FACS Analysis showing the frequency of Foxp3+ cells on day 3 of the cell culture. Naïve CD4+ T cells were treated as described in (C). (E) The percentage of Foxp3+ Tregs within CD4+CD8- thymocytes (CD4 single positive, CD4SP) in WT and rel-/- mice. Results are means ± SEM of three independent experiments; ***P<0.001.
DISCUSSION
The molecular basis of Treg development is characterized by epigenetic regulations within the Foxp3 locus [35–37]. Investigation of proximal regions at the Foxp3 locus revealed the existence of several important regulatory elements termed conserved non-coding sequences 1-3 (CNS 1-3) that play a decisive role in the control of Foxp3 gene expression [19, 38]. Novel findings suggest that c-Rel acts as “pioneer” transcription factor during the Treg development in thymus by binding at the CNS3 region of Foxp3 locus [14, 19]. Additionally, the expression of IL-2, a cytokine important for Treg homeostasis, is under direct transcriptional control of c-Rel [28, 39, 40]. Steady-state IL-2 levels are indispensable for survival of Foxp3+ Tregs [41–45]. Interestingly, we here show that defective Foxp3 expression and Tregs numbers in rel-/- mice can be partially compensated by adding exogenous IL-2. Two recent clinical studies demonstrate that the administering IL-2 at low doses provides a therapeutic window to promote self-tolerance and suppress adverse immune responses in humans. Both reports suggest that beneficial effects are mediated by boosting Treg cell numbers [46, 47]. By treating mice with IL-2/JES6-1 complexes, we were able to show that the Helios expression in Tregs is linked to the selective, IL-2-mediated enrichment of these cells. Moreover, the increase of Helios+Foxp3+ Tregs was even more pronounced in c-Rel deficient mice when compared to the naïve animals.
The expression of transcription factor Helios in Tregs has been proposed to be independent from signals required for induction of Foxp3 [48]. Although defining the origin of functional Treg subsets is difficult, Helios has been suggested to be a suitable marker for thymus-derived tTregs [4]. Recent two studies have demonstrated that also Neuropilin 1 (Nrp1) is specifically expressed on tTregs but not on pTregs [49, 50]. While these reports showed a strong correlation between Nrp1 and Helios expression exclusively on tTregs, other researchers challenged this concept demonstrating that Helios might be involved in various cellular processes in T cells such as activation, proliferation, apoptosis of autoreactive thymocytes, Th2 development as well as T cell anergy and exhaustion [51–55].
In this study, we observed more prominent reduction of Helios+Foxp3+ than of Helios-Foxp3+ Treg population in c-Rel deficient mice. The transcription factor c-Rel has been shown to regulate multiple steps in the thymic development of Foxp3-expressing CD4+ Tregs [56]. We and others have recently shown that mice lacking c-Rel possess only 15-25 % of normal Treg cell numbers and frequencies in the thymus and peripheral lymphoid organs [13, 15–18]. Interestingly, the remaining c-Rel deficient Tregs have normal immune suppressive properties in vivo [16]. In view of evidence of preferential accumulation of Helios-Foxp3+ pTregs in aged rel-/- mice and normal iTreg generation and expansion of mucosal Tregs in the absence of c-Rel, we suggest the following model of c-Rel-dependent and -independent regulation of Tregs: in the thymus, the development of Tregs requires the triggering of the T cell receptor (TCR), whereby the transcription factor c-Rel links the TCR-mediated signals to the induction of Foxp3 expression. c-Rel might bind to CNS3 in cooperation with other NF-κB-associated molecules such as IκBNS and p50 [20]. Additionally, the IL-2 production by non-Treg cells, which is needed for survival and proliferation of both tTregs and pTregs, is also transcriptionally regulated by c-Rel [28, 40]. IL-2 stimulation of Tregs activates STAT5 that binds together with RUNX1 to CNS2 (also known as TSDR) regulatory element of Foxp3 locus, which is an important step for stable Foxp3 expression and maintenance of Treg subsets. On the other hand, in pTregs and iTregs, the c-Rel-independent binding of SMAD3 and NFAT (which also binds together with AP1 to the IL2 promoter and controls IL2 gene expression, [57]) to CNS1 is sufficient to generate Tregs in an extra-thymic manner (Figure 6). Additionally, retinoic acid-activated heterodimers consisting of RAR and RXR are also able to bind to CNS1 to induce differentiation of pTregs [58–60].
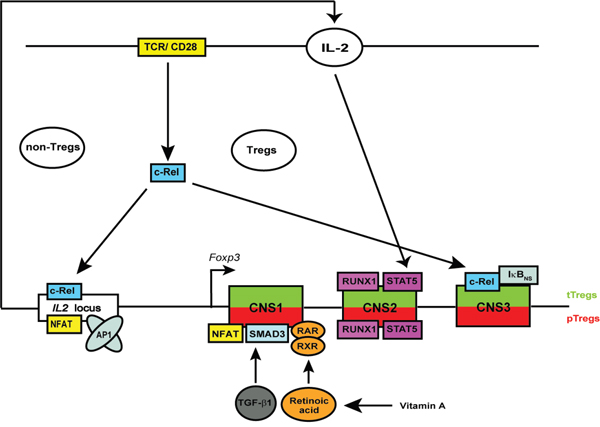
Figure 6: Schematic overview of the proposed mechanism of c-Rel-mediated regulation of Tregs. c-Rel regulates the Treg development and homeostasis by binding to the IL2 locus in non-Tregs and to the Foxp3 locus in developing thymic Tregs, respectively. c-Rel binds together with IκBNS to conserved non-coding sequence 3 (CNS3) of Foxp3 locus. Several other transcription factors such as STAT5, RUNX1 as well as NFAT, SMAD3, RAR and RXR can bind to CNS2 and CNS1, respectively. Tregs in the peripheral organs and in in vitro CD4+ T cell cultures develop independently of c-Rel. Abbreviations: TCR, T cell receptor; TGF, transforming growth factor; STAT, signal transducer and activator of transcription; NFAT, nuclear factor of activated T-cells; AP1, activator protein 1; RUNX1, runt-related transcription factor 1; RAR, retinoic acid receptor; RXR, retinoid X receptor.
MATERIALS AND METHODS
Mice
Conventional C57BL/6 mice were purchased from Charles River Laboratories (Sulzfeld, Germany) and kept under specific pathogen-free (SPF) conditions. Germ-free (GF) mice on C57BL/6 background, Foxp3-RFP reporter mice and rel-/- mice were bred at the animal facility of the Biomedical Research Center (University of Marburg, Germany). All experiments were performed according to the German animal protection law.
In vivo administration of IL-2/JES6-1 complex
Wild-type (WT) and rel-/- mice were treated with three daily injection (days 0,1, 2) of recombinant mouse IL-2 (eBioscience) with anti-IL-2 mAb (clone JES6-1, eBioscience). Recombinant IL-2 was mixed with JES6-1 and incubated for 30 min at 37°C as previously described [23]. Mice were daily i.p. injected with 6 μg IL-2/JES6-1 complex at optimal molar ratio of approximately 2:1 (1 μg/58 pmol of IL-2 and 5μg/33 pmol of JES6-1) and the phenotype of expanded cells was analysed on day 5. For kinetics studies, mLN, spleen and liver cell suspensions were analysed on day 0, 5 and 8.
Intracellular flow-cytometry
Single cell suspensions were performed from spleens, mLN and iLN by mechanical disruption and passage through pre-separation filter. Colonic lamina propria mononuclear cells (LPMCs) were isolated as previously described [61]. Subsequently, cells were stained with anti-CD4 (eBioscience), fixed and permeabilazed in 3% saponine buffer before intracellular staining. Foxp3- and Helios-expressing cells were routinely detected by using the Foxp3 staining kit (eBioscience), anti-Foxp3 (FJK-16s, eBioscience) and anti-Helios (22F6, eBioscience) Abs. Intracellular detection of IL-17A (eBio17B7) and IFN-γ (XMG1.2) was performed after restimulation of cells with PMA (50 mg/ml)/ionomycin (750 ng/ml) in the presence of brefeldin A (10 mg/ml, all substances were obtained from Sigma-Aldrich) for 4 h. Both Abs were obtained from eBioscience. The stained cells were analyzed by flow cytometry using the FlowJo software (Tree Star).
Induction of acute colitis
Colitis was induced in sex- and age-matched WT and rel-/- mice by giving them 3% (w/v) dextran sodium sulfate (DSS, molecular weight 35-50 kDa, MP Biomedicals). DSS was administered orally into the drinking water for 5 days. The body weight was monitored throughout the experiment for 14 days.
CD4+ purification and in vitro stimulation
Naïve CD4+ T cells were purified from spleens and LN by using CD4+ T cell purification kit (Miltenyi Biotec). The cells were activated by plate-bound anti-CD3 (5 mg/ml, clone 145-2C11) and soluble anti-CD28 (1.5 mg/ml, clone 37.51) and differentiated towards Foxp3-expressing Tregs by adding 2 ng/ml TGF-β1 (Peprotech), anti-IL-4 (10% culture supernatant of clone 11B11), anti-IFN-γ (5 μg/ml, clone XMG1.2) and 50 U/ml recombinant hIL-2 (Novartis). After 3 days, the expression of Foxp3 was analyzed by flow cytometry using anti-Foxp3.
Immunohistochemistry
Immunohistochemistry analysis of Foxp3 expression in the gut of WT and rel-/- mice was performed as previously described [62]. In brief, 1-2 μm sections of formalin-fixed, paraffin-embedded tissue were cut, deparaffinized, and subjected to a heat induced epitope retrieval step. Slides were rinsed in cool running water, washed in Tris-buffered saline (pH 7.4) before incubation with anti-mouse Foxp3 Ab (clone FJK-16s, eBioscience, dilution 1:50) for 30 minutes. For detection, rabbit anti-rat Ab (Dianova) was used followed by the EnvisionPO kit (DAKO). Peroxidase was developed with a highly sensitive diaminobenzidine chromogenic substrate for 5 minutes. Negative controls were performed by omitting the primary antibodies. Images were acquired using an AxioImager Z1 microscope and processed with AxioVision software (Carl Zeiss MicroImaging).
Western blot analysis
To study Stat5 phosphorylation, whole-cell lysates were prepared from purified CD4+ T cells as described earlier (19). In brief, the CompleteTM protease inhibitor cocktail (Roche Applied Science) and phosphatase inhibitors (0.5 mM Na3VO4 and 50 mM NaF) were added into RIPA lysis buffer. To determine cellular protein content, the Micro BCA Protein Assay Kit (Pierce) was used. For detection of phospho-STAT5, WT and rel-/- CD4+ T cells were isolated from spleen and LN of WT and rel-/- mice, rested for 3 h, and stimulated in vitro for 0–30 min with recombinant hIL-2 (50 U/ml, Novartis). In some experiments, CD4+ T cells were cultured under Treg-inducing conditions for 6 h and18 h before the western blot analysis, respectively. The induction of p-STAT5 was examined by using anti-p-STAT5 Ab (Tyr 694, Cell Signaling Techology). Detection of β-Actin (Sigma) and STAT5A (Santa Cruz Biotechnology) served as loading control.
Statistics
Results are generally expressed as the mean ± SEM. For statistical analysis, data were analyzed using the unpaired two-tailed Student’s t test. Results were considered statistically significant at P ≤ 0.05. F test was used to determine factorial analysis of variances between DSS-treated and untreated groups. All analyses were performed using GraphPad Prism 5.0 (GraphPad Software).
Author contribution
ML, EJ and NV performed all in vitro and in vivo experiments and analysed the data. AV supervised the investigation and wrote the manuscript. ML and NV critically revised the manuscript.
ACKNOWLEDGMENTS
We thank Anne Hellhund for technical assistance. We kindly acknowledge Dr. Anja A. Kühl for performing immunohistochemistry, Dr. Thorsten Joeris, Dr. Nicole Schmidt and Dr. Katharina Reinhard for discussing the project as well as Dr. Stephen T. Reece for providing us with CD25-depleting Ab. We also thank the staff of the animal facility, University of Marburg, for assistance with animals.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
FUNDING
This study was supported by Studienstiftung des deutschen Volkes (Maik Luu) and by the Fritz Thyssen Stiftung grant (Alexander Visekruna).
REFERENCES
1. Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. Journal of immunology. 1995; 155:1151-1164.
2. Fontenot JD, Gavin MA, Rudensky AY. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nature immunology. 2003; 4:330-336.
3. Hori S, Nomura T, Sakaguchi S. Control of regulatory T cell development by the transcription factor Foxp3. Science. 2003; 299:1057-1061.
4. Shevach EM, Thornton AM. tTregs, pTregs, and iTregs: similarities and differences. Immunological reviews. 2014; 259:88-102.
5. Feuerer M, Hill JA, Mathis D, Benoist C. Foxp3+ regulatory T cells: differentiation, specification, subphenotypes. Nature immunology. 2009; 10:689-695.
6. Polansky JK, Kretschmer K, Freyer J, Floess S, Garbe A, Baron U, Olek S, Hamann A, von Boehmer H, Huehn J. DNA methylation controls Foxp3 gene expression. European journal of immunology. 2008; 38:1654-1663.
7. Ohkura N, Hamaguchi M, Morikawa H, Sugimura K, Tanaka A, Ito Y, Osaki M, Tanaka Y, Yamashita R, Nakano N, Huehn J, Fehling HJ, Sparwasser T, et al. T cell receptor stimulation-induced epigenetic changes and Foxp3 expression are independent and complementary events required for Treg cell development. Immunity. 2012; 37:785-799.
8. Lal G, Zhang N, van der Touw W, Ding Y, Ju W, Bottinger EP, Reid SP, Levy DE, Bromberg JS. Epigenetic regulation of Foxp3 expression in regulatory T cells by DNA methylation. Journal of immunology. 2009; 182:259-273.
9. Marie JC, Letterio JJ, Gavin M, Rudensky AY. TGF-beta1 maintains suppressor function and Foxp3 expression in CD4+CD25+ regulatory T cells. The Journal of experimental medicine. 2005; 201:1061-1067.
10. Takimoto T, Wakabayashi Y, Sekiya T, Inoue N, Morita R, Ichiyama K, Takahashi R, Asakawa M, Muto G, Mori T, Hasegawa E, Saika S, Hara T, et al. Smad2 and Smad3 are redundantly essential for the TGF-beta-mediated regulation of regulatory T plasticity and Th1 development. Journal of immunology. 2010; 185:842-855.
11. Arpaia N, Rudensky AY. Microbial metabolites control gut inflammatory responses. Proceedings of the National Academy of Sciences of the United States of America. 2014; 111:2058-2059.
12. Furusawa Y, Obata Y, Fukuda S, Endo TA, Nakato G, Takahashi D, Nakanishi Y, Uetake C, Kato K, Kato T, Takahashi M, Fukuda NN, Murakami S, et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature. 2013; 504:446-450.
13. Long M, Park SG, Strickland I, Hayden MS, Ghosh S. Nuclear factor-kappaB modulates regulatory T cell development by directly regulating expression of Foxp3 transcription factor. Immunity. 2009; 31:921-931.
14. Hori S. c-Rel: a pioneer in directing regulatory T-cell lineage commitment? European journal of immunology. 2010; 40:664-667.
15. Deenick EK, Elford AR, Pellegrini M, Hall H, Mak TW, Ohashi PS. c-Rel but not NF-kappaB1 is important for T regulatory cell development. European journal of immunology. 2010; 40:677-681.
16. Isomura I, Palmer S, Grumont RJ, Bunting K, Hoyne G, Wilkinson N, Banerjee A, Proietto A, Gugasyan R, Wu L, McNally A, Steptoe RJ, Thomas R, et al. c-Rel is required for the development of thymic Foxp3+ CD4 regulatory T cells. The Journal of experimental medicine. 2009; 206:3001-3014.
17. Ruan Q, Kameswaran V, Tone Y, Li L, Liou HC, Greene MI, Tone M, Chen YH. Development of Foxp3(+) regulatory t cells is driven by the c-Rel enhanceosome. Immunity. 2009; 31:932-940.
18. Visekruna A, Huber M, Hellhund A, Bothur E, Reinhard K, Bollig N, Schmidt N, Joeris T, Lohoff M, Steinhoff U. c-Rel is crucial for the induction of Foxp3(+) regulatory CD4(+) T cells but not T(H)17 cells. European journal of immunology. 2010; 40:671-676.
19. Zheng Y, Josefowicz S, Chaudhry A, Peng XP, Forbush K, Rudensky AY. Role of conserved non-coding DNA elements in the Foxp3 gene in regulatory T-cell fate. Nature. 2010; 463:808-812.
20. Schuster M, Glauben R, Plaza-Sirvent C, Schreiber L, Annemann M, Floess S, Kuhl AA, Clayton LK, Sparwasser T, Schulze-Osthoff K, Pfeffer K, Huehn J, Siegmund B, Schmitz I. IkappaB(NS) protein mediates regulatory T cell development via induction of the Foxp3 transcription factor. Immunity. 2012; 37:998-1008.
21. Brustle A, Brenner D, Knobbe-Thomsen CB, Cox M, Lang PA, Lang KS, Mak TW. MALT1 is an intrinsic regulator of regulatory T cells. Cell death and differentiation. 2015.
22. Wilson MS, Pesce JT, Ramalingam TR, Thompson RW, Cheever A, Wynn TA. Suppression of murine allergic airway disease by IL-2:anti-IL-2 monoclonal antibody-induced regulatory T cells. Journal of immunology. 2008; 181:6942-6954.
23. Webster KE, Walters S, Kohler RE, Mrkvan T, Boyman O, Surh CD, Grey ST, Sprent J. In vivo expansion of T reg cells with IL-2-mAb complexes: induction of resistance to EAE and long-term acceptance of islet allografts without immunosuppression. The Journal of experimental medicine. 2009; 206:751-760.
24. Grinberg-Bleyer Y, Baeyens A, You S, Elhage R, Fourcade G, Gregoire S, Cagnard N, Carpentier W, Tang Q, Bluestone J, Chatenoud L, Klatzmann D, Salomon BL, Piaggio E. IL-2 reverses established type 1 diabetes in NOD mice by a local effect on pancreatic regulatory T cells. The Journal of experimental medicine. 2010; 207:1871-1878.
25. Thornton AM, Korty PE, Tran DQ, Wohlfert EA, Murray PE, Belkaid Y, Shevach EM. Expression of Helios, an Ikaros transcription factor family member, differentiates thymic-derived from peripherally induced Foxp3+ T regulatory cells. Journal of immunology. 2010; 184:3433-3441.
26. Pan F, Yu H, Dang EV, Barbi J, Pan X, Grosso JF, Jinasena D, Sharma SM, McCadden EM, Getnet D, Drake CG, Liu JO, Ostrowski MC, Pardoll DM. Eos mediates Foxp3-dependent gene silencing in CD4+ regulatory T cells. Science. 2009; 325:1142-1146.
27. Shevach EM. Application of IL-2 therapy to target T regulatory cell function. Trends in immunology. 2012; 33:626-632.
28. Liou HC, Jin Z, Tumang J, Andjelic S, Smith KA, Liou ML. c-Rel is crucial for lymphocyte proliferation but dispensable for T cell effector function. International immunology. 1999; 11:361-371.
29. Chen Q, Kim YC, Laurence A, Punkosdy GA, Shevach EM. IL-2 controls the stability of Foxp3 expression in TGF-beta-induced Foxp3+ T cells in vivo. Journal of immunology. 2011; 186:6329-6337.
30. Hirakawa M, Matos T, Liu H, Koreth J, Kim HT, Paul NE, Murase K, Whangbo J, Alho AC, Nikiforow S, Cutler C, Ho VT, Armand P, et al. Low-dose IL-2 selectively activates subsets of CD4+ Tregs and NK cells. JCI insight. 2016; 1:e89278.
31. Atarashi K, Tanoue T, Shima T, Imaoka A, Kuwahara T, Momose Y, Cheng G, Yamasaki S, Saito T, Ohba Y, Taniguchi T, Takeda K, Hori S, et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science. 2011; 331:337-341.
32. Atarashi K, Tanoue T, Oshima K, Suda W, Nagano Y, Nishikawa H, Fukuda S, Saito T, Narushima S, Hase K, Kim S, Fritz JV, Wilmes P, et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature. 2013; 500:232-236.
33. Lages CS, Suffia I, Velilla PA, Huang B, Warshaw G, Hildeman DA, Belkaid Y, Chougnet C. Functional regulatory T cells accumulate in aged hosts and promote chronic infectious disease reactivation. Journal of immunology. 2008; 181:1835-1848.
34. Raynor J, Karns R, Almanan M, Li KP, Divanovic S, Chougnet CA, Hildeman DA. IL-6 and ICOS Antagonize Bim and Promote Regulatory T Cell Accrual with Age. Journal of immunology. 2015; 195:944-952.
35. Huehn J, Polansky JK, Hamann A. Epigenetic control of FOXP3 expression: the key to a stable regulatory T-cell lineage? Nature reviews Immunology. 2009; 9:83-89.
36. Rudensky AY. Regulatory T cells and Foxp3. Immunological reviews. 2011; 241:260-268.
37. Ohkura N, Kitagawa Y, Sakaguchi S. Development and maintenance of regulatory T cells. Immunity. 2013; 38:414-423.
38. Josefowicz SZ, Rudensky A. Control of regulatory T cell lineage commitment and maintenance. Immunity. 2009; 30:616-625.
39. Kontgen F, Grumont RJ, Strasser A, Metcalf D, Li R, Tarlinton D, Gerondakis S. Mice lacking the c-rel proto-oncogene exhibit defects in lymphocyte proliferation, humoral immunity, and interleukin-2 expression. Genes & development. 1995; 9:1965-1977.
40. Rao S, Gerondakis S, Woltring D, Shannon MF. c-Rel is required for chromatin remodeling across the IL-2 gene promoter. Journal of immunology. 2003; 170:3724-3731.
41. Almeida AR, Legrand N, Papiernik M, Freitas AA. Homeostasis of peripheral CD4+ T cells: IL-2R alpha and IL-2 shape a population of regulatory cells that controls CD4+ T cell numbers. Journal of immunology. 2002; 169:4850-4860.
42. Malek TR, Yu A, Vincek V, Scibelli P, Kong L. CD4 regulatory T cells prevent lethal autoimmunity in IL-2Rbeta-deficient mice. Implications for the nonredundant function of IL-2. Immunity. 2002; 17:167-178.
43. Fontenot JD, Rasmussen JP, Gavin MA, Rudensky AY. A function for interleukin 2 in Foxp3-expressing regulatory T cells. Nature immunology. 2005; 6:1142-1151.
44. Setoguchi R, Hori S, Takahashi T, Sakaguchi S. Homeostatic maintenance of natural Foxp3(+) CD25(+) CD4(+) regulatory T cells by interleukin (IL)-2 and induction of autoimmune disease by IL-2 neutralization. The Journal of experimental medicine. 2005; 201:723-735.
45. Chinen T, Kannan AK, Levine AG, Fan X, Klein U, Zheng Y, Gasteiger G, Feng Y, Fontenot JD, Rudensky AY. An essential role for the IL-2 receptor in Treg cell function. Nature immunology. 2016; 17:1322-1333.
46. Saadoun D, Rosenzwajg M, Joly F, Six A, Carrat F, Thibault V, Sene D, Cacoub P, Klatzmann D. Regulatory T-cell responses to low-dose interleukin-2 in HCV-induced vasculitis. The New England journal of medicine. 2011; 365:2067-2077.
47. Koreth J, Matsuoka K, Kim HT, McDonough SM, Bindra B, Alyea EP 3rd, Armand P, Cutler C, Ho VT, Treister NS, Bienfang DC, Prasad S, Tzachanis D, et al. Interleukin-2 and regulatory T cells in graft-versus-host disease. The New England journal of medicine. 2011; 365:2055-2066.
48. Sugimoto N, Oida T, Hirota K, Nakamura K, Nomura T, Uchiyama T, Sakaguchi S. Foxp3-dependent and -independent molecules specific for CD25+CD4+ natural regulatory T cells revealed by DNA microarray analysis. International immunology. 2006; 18:1197-1209.
49. Yadav M, Louvet C, Davini D, Gardner JM, Martinez-Llordella M, Bailey-Bucktrout S, Anthony BA, Sverdrup FM, Head R, Kuster DJ, Ruminski P, Weiss D, Von Schack D, Bluestone JA. Neuropilin-1 distinguishes natural and inducible regulatory T cells among regulatory T cell subsets in vivo. The Journal of experimental medicine. 2012; 209:1713-1722, S1711-1719.
50. Weiss JM, Bilate AM, Gobert M, Ding Y, Curotto de Lafaille MA, Parkhurst CN, Xiong H, Dolpady J, Frey AB, Ruocco MG, Yang Y, Floess S, Huehn J, et al. Neuropilin 1 is expressed on thymus-derived natural regulatory T cells, but not mucosa-generated induced Foxp3+ T reg cells. The Journal of experimental medicine. 2012; 209:1723-1742, S1721.
51. Serre K, Benezech C, Desanti G, Bobat S, Toellner KM, Bird R, Chan S, Kastner P, Cunningham AF, Maclennan IC, Mohr E. Helios is associated with CD4 T cells differentiating to T helper 2 and follicular helper T cells in vivo independently of Foxp3 expression. PloS one. 2011; 6:e20731.
52. Akimova T, Beier UH, Wang L, Levine MH, Hancock WW. Helios expression is a marker of T cell activation and proliferation. PloS one. 2011; 6:e24226.
53. Daley SR, Hu DY, Goodnow CC. Helios marks strongly autoreactive CD4+ T cells in two major waves of thymic deletion distinguished by induction of PD-1 or NF-kappaB. The Journal of experimental medicine. 2013; 210:269-285.
54. Ross EM, Bourges D, Hogan TV, Gleeson PA, van Driel IR. Helios defines T cells being driven to tolerance in the periphery and thymus. European journal of immunology. 2014; 44:2048-2058.
55. Crawford A, Angelosanto JM, Kao C, Doering TA, Odorizzi PM, Barnett BE, Wherry EJ. Molecular and transcriptional basis of CD4(+) T cell dysfunction during chronic infection. Immunity. 2014; 40:289-302.
56. Grigoriadis G, Vasanthakumar A, Banerjee A, Grumont R, Overall S, Gleeson P, Shannon F, Gerondakis S. c-Rel controls multiple discrete steps in the thymic development of Foxp3+ CD4 regulatory T cells. PloS one. 2011; 6:e26851.
57. Muller MR, Rao A. NFAT, immunity and cancer: a transcription factor comes of age. Nature reviews Immunology. 2010; 10:645-656.
58. Tone Y, Furuuchi K, Kojima Y, Tykocinski ML, Greene MI, Tone M. Smad3 and NFAT cooperate to induce Foxp3 expression through its enhancer. Nature immunology. 2008; 9:194-202.
59. Xu L, Kitani A, Stuelten C, McGrady G, Fuss I, Strober W. Positive and negative transcriptional regulation of the Foxp3 gene is mediated by access and binding of the Smad3 protein to enhancer I. Immunity. 2010; 33:313-325.
60. Vaeth M, Schliesser U, Muller G, Reissig S, Satoh K, Tuettenberg A, Jonuleit H, Waisman A, Muller MR, Serfling E, Sawitzki BS, Berberich-Siebelt F. Dependence on nuclear factor of activated T-cells (NFAT) levels discriminates conventional T cells from Foxp3+ regulatory T cells. Proceedings of the National Academy of Sciences of the United States of America. 2012; 109:16258-16263.
61. Schmidt N, Gonzalez E, Visekruna A, Kuhl AA, Loddenkemper C, Mollenkopf H, Kaufmann SH, Steinhoff U, Joeris T. Targeting the proteasome: partial inhibition of the proteasome by bortezomib or deletion of the immunosubunit LMP7 attenuates experimental colitis. Gut. 2010; 59:896-906.
62. Rausch S, Huehn J, Loddenkemper C, Hepworth MR, Klotz C, Sparwasser T, Hamann A, Lucius R, Hartmann S. Establishment of nematode infection despite increased Th2 responses and immunopathology after selective depletion of Foxp3+ cells. European journal of immunology. 2009; 39:3066-3077.