
INTRODUCTION
Osteoarthritis (OA), a degenerative joint disease, is characterized by joint pain, stiffness and swelling, degeneration of articular cartilage, intra-articular inflammation with synovitis and changes in peri-articular and sub-chondral bone [1]. OA is reported to be the fourth leading cause of disability, which affects 27% of the population in industrialized countries with age above 45 years old in 2012 [2, 3]. Incidence of OA has been suggested to increase with age, rising sharply over the age of 50 years old and leveling off after 80 years old [4]. Subchondral bone sclerosis and progressive cartilage degaradation are thought as hallmarks of OA [5]. Degeneration of articular cartilage is proven to be the obvious feature of OA that eventually causes joint destruction, because of the imbalance of anabolic activities [6, 7]. Treatment for degenerative OA usually includes pharmacological and non-pharmacological modalities [8, 9]. Although degenerative OA has poor prognosis, it is significant to explore the mechanisms for the pathogenesis and treatment of degenerative OA.
Toll-like receptors (TLRs) are evolutionarily conserved molecules that promote immune responses through recognition of microbial-associated molecular patterns (MAMPs); TLR signaling is indispensable for proper activation of immune responses during infections [10]. The TLR family is best characterized as group of innate immune receptors in terms of known ligands, functional relevance and downstream signaling pathways [11]. TLR signaling has been implicated in the pathogenesis of sepsis, asthma, atherosclerosis, autoimmune disorders [12–15]. While pathogen recognition begins at the receptor level, specific transcriptional response and immunological outcome are ultimately determined by the signaling components downstream and the way they interact with each other [16]. Nuclear factor-kappa B (NF-κB) has been recognized to be the master orchestrator of TLR-induced responses; all TLR signals converge on NF-κB and activation of NF-κB is critical for TLR function [17]. NF-κB is a transcription factor regulating gene expression, which can control multiple cellular functions, such as inflammatory and stress-induced responses and survival [18]. NF-κB is a critical regulator of innate immunity/inflammation; aberrant NF-κB regulation has involved in many cancers[19–22]. Besides, TLR2 expression increases in OA chondrocytes, and synoviocytes could produce RANKL, receptor activator of NF-κB ligand, which is involved in cartilage degradation and joint destruction [23, 24]. Although TLR-2/NF-κB signaling pathway might participate in the occurrence of degenerative OA, the inner influence mechanism is still unclear. Consistently, as a pro-inflammatory cytokine, IL-1β is a primary instigator in cartilage degradation in OA [25]. Also, previous study has proven that up-regulated matrix metalloprotease-13 (MMP-13) expression is essential in the OA pathogenesis [26]. Therefore, this study aims to explore the effects of TLR-2/NF-κB signaling pathway on the occurrence of degenerative OA.
RESULTS
Baseline characteristics of subjects in the OA and normal groups
Baseline characteristics of subjects in the OA and normal groups were shown in Table 1, in which age, gender, smoking, drinking and tea drinking habits had no significant difference and BMI, family history and exercise had statistically significance.
Table 1: Comparisons of baseline characteristics of subjects in the OA and normal groups
Index |
Case group (n = 231) |
Normal group (n=198) |
P value |
|---|---|---|---|
Age |
0.336 |
||
≥ 60 |
174 |
141 |
|
< 60 |
57 |
57 |
|
Gender |
0.368 |
||
Male |
79 |
76 |
|
Female |
152 |
122 |
|
BMI (kg/m2) |
< 0.001 |
||
< 23 |
111 |
99 |
|
23 ~ 25 |
64 |
81 |
|
> 25 |
56 |
18 |
|
Smoking |
0.132 |
||
Yes |
116 |
85 |
|
No |
115 |
113 |
|
Drinking |
0.196 |
||
Yes |
80 |
57 |
|
No |
151 |
141 |
|
Family history |
< 0.001 |
||
Yes |
138 |
83 |
|
No |
93 |
115 |
|
Exercise |
0.008 |
||
< 1 time/month |
24 |
42 |
|
< 1 time/week |
66 |
50 |
|
> 2 times/week |
151 |
106 |
|
Tea drinking habit |
0.146 |
||
Yes |
201 |
181 |
|
No |
30 |
17 |
Note: OA, osteoarthritis.
Structure changes of the normal articular cartilage tissues and OA articular cartilage tissues
Structure changes of the normal articular cartilage tissues and OA articular cartilage tissues observed under the microscope were showed in Figure 1. According to the HE staining, the normal chondrocytes arranged in neat rows; number and morphology were normal; cartilage structure was clear and the tidal line was continuous and complete. While the OA chondrocytes showed hyperplasia, hypertrophy and disorder; they clustered or disappeared; the surface layer showed different degrees of damage, accompanied with angiogenesis. According to the results of safranin O/fast green FCF staining, with the aggravation of the lesion, the red area of the normal cartilage tissues turned light in color; the area of dyeing loss expanded until complete dyeing loss occurred; the tidal line was also damaged; new blood vessels were seen to connect marrow cavity through the tidal line. The cartilage pathological scoring results showed that there were 198 cases in the normal group, 104 cases and 127 cases in the OA mild lesion group and the OA moderate lesion group, respectively.
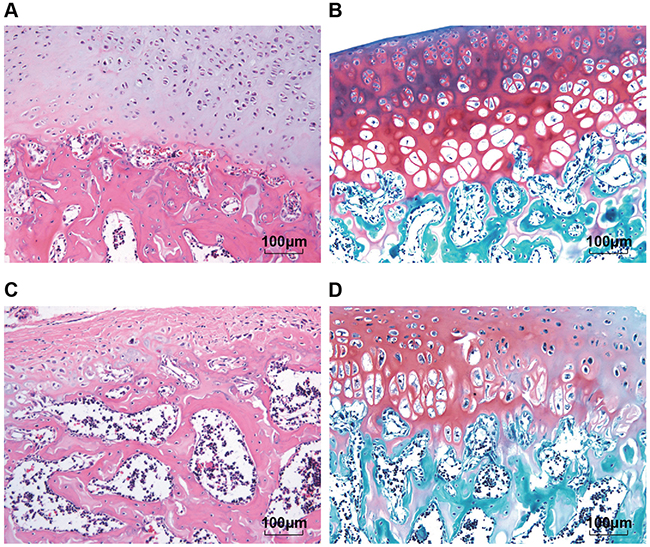
Figure 1: Comparisons of structure changes of the normal articular cartilage tissues and OA articular cartilage tissues according to the HE staining and safranin O/fast green FCF staining (200 ×). Notes: (A) and (B) were the result of normal cartilage tissues, respectively by HE staining and safranin O and fast green FCF staining; (C) and (D) were the result of OA cartilage tissues, respectively by HE staining and safranin O and fast green FCF staining; HE, Hematoxylin-Eosin; OA, osteoarthritis.
The expressions of TLR-2, NF-κB and MMP-13 among three groups
According to the results of the immunochemistry (Figure 2A), the positive expressions of TLR-2 and NF-κB were presented with brown yellow particles deposition in the nucleus of articular cartilage and a small amount in the cytoplasm of articular cartilage; while MMP-13 positive expression was presented with brown yellow particles deposition in the cytoplasm. Negative expression was indicated when only cell profile was observed, and the nucleus and cytoplasm were not colored. In the normal group, TLR-2, NF-κB and MMP-13 positively expressed in 17, 16 and 18 cases, independently, and the positive rates were 8.59%, 8.08% and 9.09%, respectively. In the OA mild lesion group, the cases of TLR-2, NF-κB and MMP-13 positively expressed were 52, 53 and 59, respectively, and the corresponding positive rates were 50.00%, 50.96% and 56.73%, respectively. In the OA moderate lesion group, the cases of TLR-2, NF-κB and MMP-13 positively expressed were 112, 119 and 117, respectively, and the corresponding positive rates were 88.19%, 93.70% and 92.13%, respectively. In pairwise comparisons, the difference of the positive expressions of TLR-2, NF-κB and MMP-13 was statistically significant (all P < 0.05); the positive expressions of TLR-2, NF-κB and MMP-13 were positively correlated with the degree of lesion (Figure 2B).
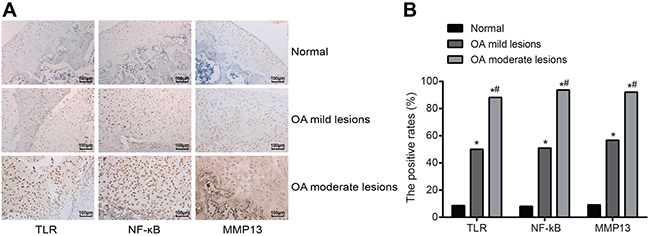
Figure 2: Comparisons of positive expressions of TLR-2, NF-κB and MMP-13 by immunochemistry among three groups. Notes: (A) Detection of protein expressions of TLR-2, NF-κB and MMP-13 by immunochemistry among three groups (positive expression was presented as the brown yellow particles deposition, in the nucleus and part of cytoplasm of articular chondrocytes for TLR-2 and NF-κB and nucleus for MMP-13); (B) Positive protein expressions of TLR-2, NF-κB and MMP-13 in the chondrocytes by immunochemistry among three groups; *, compared with the normal group, P < 0.05; #, compared with the OA moderate lesion group, P < 0.05; TLR-2, toll-like receptor-2; NF-κB, nuclear transcription factor-kappa B; MMP-13, matrix metalloprotease-13; OA, osteoarthritis.
Correlation between the expressions of TLR, NF-kB, MMP13 and clinicopathological characteristics of OA patients
Correlation between the expressions of TLR, NF-kB, MMP13 and clinicopathological characteristics of OA patients were shown in Table 2. The positive expressions of TLR, NF-kB, MMP13 were associated with BMI, family history, exercise, and WOMAC scores of OA patients (all P < 0.05), but age, gender, smoking, drinking and tea drinking habits showed no significant difference.
Table 2: Correlation between the expressions of TLR, NF-kB, MMP13 and clinicopathological characteristics of OA patients
Characteristic |
TLR |
NF-kB |
MMP13 |
||||||
|---|---|---|---|---|---|---|---|---|---|
Positive |
Negative |
P |
Positive |
Negative |
P |
Positive |
Negative |
P |
|
Case |
Case |
Case |
Case |
Case |
Case |
||||
Age (years) |
0.875 |
0.586 |
0.357 |
||||||
≥ 60 |
124 |
50 |
1248 |
46 |
130 |
44 |
|||
< 60 |
40 |
17 |
44 |
13 |
46 |
11 |
|||
Gender |
0.071 |
0.489 |
0.117 |
||||||
Male |
62 |
17 |
61 |
18 |
65 |
14 |
|||
Female |
102 |
50 |
111 |
41 |
111 |
41 |
|||
BMI (kg/m2) |
0.014 |
0.012 |
0.002 |
||||||
< 23 |
71 |
40 |
76 |
35 |
76 |
35 |
|||
23 ~ 25 |
45 |
19 |
46 |
18 |
48 |
16 |
|||
> 25 |
48 |
8 |
50 |
6 |
52 |
4 |
|||
Smoking |
0.495 |
0.163 |
0.142 |
||||||
Yes |
80 |
36 |
91 |
25 |
94 |
25 |
|||
No |
84 |
31 |
81 |
34 |
82 |
34 |
|||
Drinking |
0.584 |
0.891 |
0.199 |
||||||
Yes |
55 |
25 |
60 |
20 |
57 |
23 |
|||
No |
109 |
42 |
112 |
39 |
119 |
32 |
|||
Family history |
0.038 |
< 0.001 |
0.002 |
||||||
Yes |
105 |
33 |
117 |
21 |
115 |
23 |
|||
No |
59 |
34 |
55 |
38 |
61 |
32 |
|||
Exercise |
0.008 |
0.022 |
0.044 |
||||||
Few |
11 |
13 |
18 |
6 |
19 |
5 |
|||
A few |
45 |
21 |
41 |
25 |
43 |
23 |
|||
Often |
108 |
33 |
113 |
28 |
114 |
27 |
|||
Tea drinking habit |
0.575 |
0.88 |
0.948 |
||||||
Yes |
20 |
10 |
22 |
8 |
23 |
7 |
|||
No |
144 |
57 |
150 |
51 |
153 |
48 |
|||
WOMAC scores |
0.01 |
0.027 |
0.023 |
||||||
Mild |
72 |
46 |
79 |
39 |
81 |
37 |
|||
Moderate |
73 |
20 |
76 |
17 |
78 |
15 |
|||
Severe |
19 |
1 |
17 |
3 |
17 |
3 |
|||
Note: TLR-2, toll-like receptor-2; NF-κB, nuclear transcription factor kappa B; MMP-13, matrix metalloprotease-13; OA, osteoarthritis.
Logistic regression analysis of the risk factors of OA
Whether having OA or not was served as the dependent variable and the age, sex, BMI, smoking, drinking, family history, exercise, and drinking habits were included in logistic regression analysis. And the results showed that TLR, NF-kB, MMP13, BMI, family history and exercise were related to OA (all P < 0.05), in which LR, NF-kB, MMP13, BMI and family history were risk factors (all EXP(B) > 1, P < 0.05) and exercise was protection factor (EXP(B) < 1, P < 0.05). Also, the age, sex, smoking, drinking, drinking habits and OA had no obvious correlation (all P > 0.05) (Table 3).
Table 3: Logistic regression analysis of the factors related to OA
Factor |
B |
S.E. |
Wald |
Sig, |
Exp (B) |
95% C.I |
|---|---|---|---|---|---|---|
Age (years) |
0.24 |
0.27 |
0.78 |
0.376 |
1.27 |
0.75 - 2.15 |
Gender |
-0.41 |
0.25 |
2.76 |
0.097 |
0.66 |
0.41 - 1.08 |
BMI (kg/m2) |
||||||
< 23 |
10 |
0.007 |
||||
23 ~ 25 |
0.6 |
0.31 |
3.8 |
0.051 |
1.83 |
1.00 - 3.35 |
> 25 |
1.17 |
0.37 |
10 |
0.002 |
3.23 |
1.56 - 6.70 |
Smoking |
0.19 |
0.24 |
0.63 |
0.428 |
1.21 |
0.76 - 1.92 |
Drinking |
0.42 |
0.26 |
2.62 |
0.106 |
1.52 |
0.92 - 2.53 |
Family history |
0.65 |
0.27 |
5.92 |
0.015 |
1.92 |
1.14 - 3.26 |
Exercise |
||||||
< 1 time/month |
12.17 |
0.002 |
||||
< 1 time/week |
-1.07 |
0.37 |
8.34 |
0.004 |
0.34 |
0.16 - 0.71 |
> 2 times/week |
0.28 |
0.28 |
0.99 |
0.321 |
1.32 |
0.76 - 2.27 |
Tea drinking habit |
0.37 |
0.4 |
0.87 |
0.352 |
1.45 |
0.67 - 3.14 |
TLR |
1.23 |
0.25 |
24.48 |
< 0.001 |
3.42 |
2.10 - 5.56 |
NFKB |
0.92 |
0.26 |
12.51 |
< 0.001 |
2.52 |
1.51 - 4.20 |
MMP13 |
1.37 |
0.25 |
29.35 |
< 0.001 |
3.93 |
2.39 - 6.44 |
Note: TLR-2, toll-like receptor-2; NF-κB, nuclear transcription factor kappa B; MMP-13, matrix metalloprotease-13; OA, osteoarthritis; B: constant value; S.E.: standard error; Sig: significant; CI: confidence interval.
The mRNA expressions of TLR-2, NF-κB and MMP-13, IL-1, IL-6 and TNF-α among three groups
As shown in Figure 3A, compared with the normal group, the OA mild and moderate lesion groups showed significantly increased mRNA expressions of TLR-2, NF-κB and MMP-13 (all P < 0.05); the OA moderate lesion group showed significantly higher mRNA expressions of TLR-2, NF-κB and MMP-13 than those in the OA mild lesion group (all P < 0.05). The mRNA expressions of the related inflammatory factors are shown in Figure 3B, specifically, compared with the normal group, the OA mild and moderate lesion groups showed significantly increased mRNA expressions of IL-1, IL-6 and TNF-α (all P < 0.05); the OA moderate lesion group showed significantly higher mRNA expressions of IL-1, IL-6 and TNF-α than those in the OA mild lesion group (all P < 0.05). The mRNA expressions of TLR-2, NF-κB, MMP-13, IL-1, IL-6 and TNF-α were positively correlated with the degree of lesion.
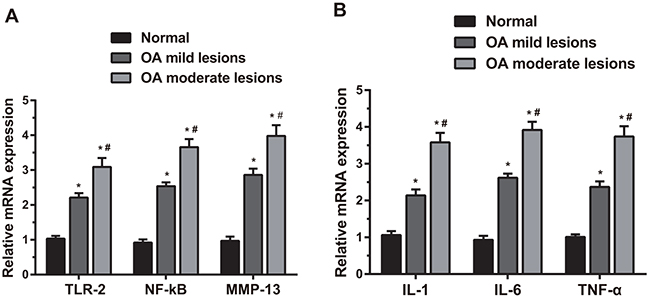
Figure 3: Comparisons of mRNA expressions of TLR-2, NF-κB, MMP-13, IL-1, IL-6 and TNF-α in the cartilage tissues by qRT-PCR among three groups. Notes: (A). Comparisons of mRNA expressions of TLR-2, NF-κB and MMP-13 among three groups; (B). comparisons of mRNA expressions of IL-1, IL-6 and TNF-α among three groups; *, compared with the normal group, P < 0.05; #, compared with the OA moderate lesion group, P < 0.05; OA, osteoarthritis.
Isolation, culture and identification of chondrocytes
Primary human chondrocytes were presented in a triangle with the cell cluster as the center, or in irregular shape; they crawled around. The chondrocytes of the first generation had less cluster compared with the primary cells, which with uniform morphology; the cells were slightly sparse in density, and with pseudopodia connected cells (Figure 4A, 4B). Type 2 collagen usually was presented in brown yellow after immunochemistry; we observed quantity of brown yellow particles in the cytoplasm and surrounding of chondrocytes (Figure 4C). According to the toluidine blue staining, the chondrocytes were mainly in light blue, which indicated the existence of proteoglycan and other extracellular matrix secreted by the chondrocytes (Figure 4D).
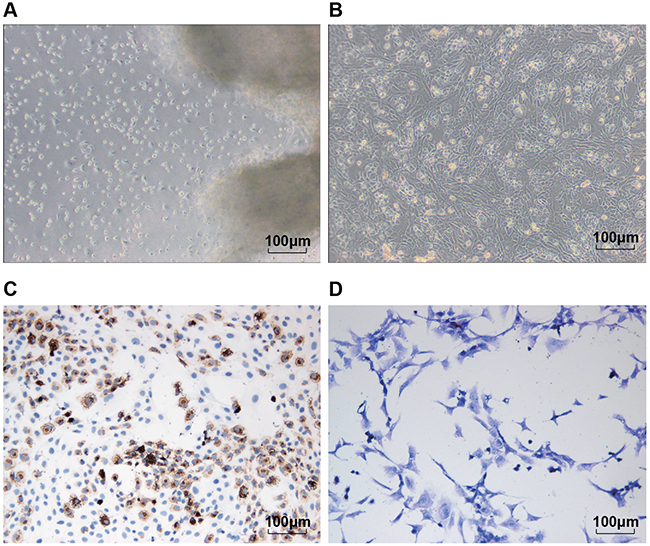
Figure 4: Isolation, culture and identification of chondrocytes. Notes: (A) and (B) were respectively the primary and first generation of the separated chondrocytes (×100); (C) type II collagen staining results of the chondrocytes (×200); (D) toluidine blue staining results of the chondrocytes (×200).
Comparisons of the proliferation of chondrocytes among four groups
According to the MTT assay (Figure 5), the OD value which indicated proliferation of cells in each group was not statistically significant at 24 h (all P > 0.05). At 48 h and 72 h, the chondrocytes in the IL-1β group, the IL-1β + anti-TLR-2 group and the IL-1β + PDTC group were significantly inhibited in growth compared with the control group (all P < 0.05). At 48 h and 72 h, the chondrocytes in the IL-1β + anti-TLR-2 group and the IL-1β + PDTC group showed faster cell growth compared with those in the IL-1β group (all P < 0.05). The chondrocytes in the IL-1β + anti-TLR-2 group and the IL-1β + PDTC group showed no significant difference in proliferation at the three time points (all P > 0.05).
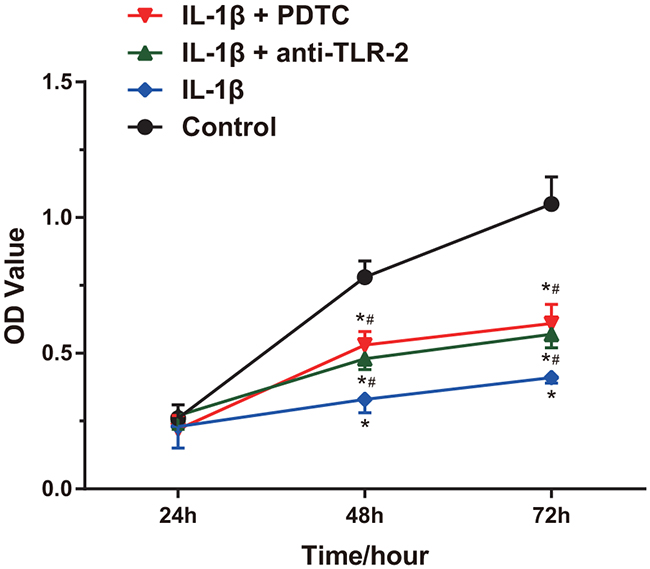
Figure 5: Comparisons of proliferation of chondrocytes among four groups. Notes: IL-1β significantly inhibited cell growth; *, compared with the control group, P < 0.05; #, compared with the IL-1β group, P < 0.05; OD, optic density.
Comparisons of cell apoptosis of chondrocytes among four groups
According to the Annexin V/PI method (Figure 6), the apoptosis rates of the cells in the control group, the IL-1β group, the IL-1β + anti-TLR-2 group and the IL-1β + PDTC group after treated 48 h were (2.97 ± 0.33)%, (22.15 ± 2.17)%, (13.72 ± 1.13)% and (12.54 ± 1.02)%, respectively. Whether interfered by anti-TLR-2 and PDTC or not, the apoptosis rate of the chondrocytes in the IL-1β group increased significantly compared with the control group (all P < 0.05). The apoptosis rate of the chondrocytes in the IL-1β + anti-TLR-2 and IL-1β + PDTC groups decreased significantly compared with the IL-1β group (all P < 0.05). The IL-1β + anti-TLR-2 group and the IL-1β + PDTC group showed no significant difference in the apoptosis rate of the chondrocytes.
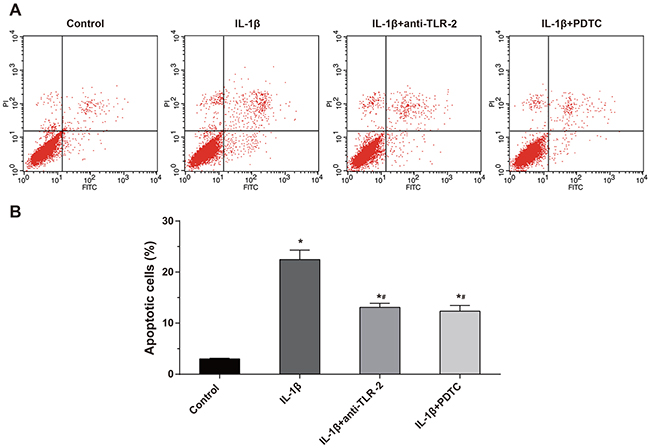
Figure 6: Comparisons of apoptosis of chondrocytes by Annexin V/PI method among four groups. Notes: (A) Apoptosis of chondrocytes among four groups by flow cytometry; (B) histogram of the apoptosis rate in each group; *, compared with the control group, P < 0.05; #, compared with the IL-1β group, P < 0.05.
Comparisons of the protein expressions of TLR, NF-kB, p-NF-kB and MMP13 among four groups
As shown in Figure 7, compared with the control group, the IL-1β group, the IL-1β + anti-TLR-2 group and the IL-1β + PDTC group showed significantly increased protein expressions of TLR-2, NF-κB, p-NF-κB and MMP-13 (all P < 0.05). Compared with the chondrocytes in the IL-1β group, the cells in the IL-1β + anti-TLR-2 and IL-1β + PDTC groups showed decreased protein expressions of NF-κB, p-NF-κB and MMP-13 (all P < 0.05); the cells in the IL-1β + anti-TLR-2 group showed decreased protein expression of TLR-2 (P < 0.05); the cells in the IL-1β + PDTC group showed no significant difference in the protein expression of TLR-2. The IL-1β + anti-TLR-2 group and the IL-1β + PDTC group showed no significant difference in the protein expressions of TLR-2, NF-κB, p-NF-κB and MMP-13.
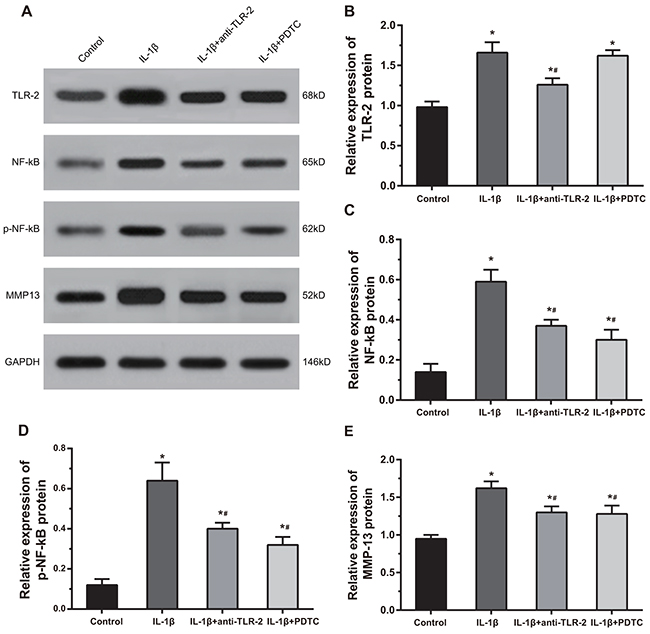
Figure 7: Comparisons of protein expressions of TLR-2, NF-κB, p-NF-κB and MMP-13 by Western blotting among four groups. Notes: (A) Protein bands of TLR-2, NF-κB, p-NF-κB and MMP-13 in chondrocytes among four groups; (B-E) histograms of the protein expressions TLR-2, NF-κB, p-NF-κB and MMP-13 in chondrocytes; p-NF-κB, phosphorylated- nuclear transcription factor-kappa B; *, compared with the control group, P < 0.05; #, compared with the IL-1β group, P < 0.05.
Comparisons of the expressions of IL-1, IL-6 and TNF-α among four groups
As shown in Figure 8, compared with the control, IL-1β, IL-1β + anti-TLR-2 and IL-1β + PDTC groups showed significantly increased expressions of IL-1, IL-6 and TNF-α (all P < 0.05). Compared with the IL-1β group, the cell supernatants in the IL-1β + anti-TLR-2 and IL-1β + PDTC groups showed obvious decreased expressions of IL-1, IL-6 and TNF-α (all P < 0.05). The cell supernatant in the IL-1β + anti-TLR-2 and IL-1β + PDTC groups showed no significant differences in the expressions of IL-1, IL-6 and TNF-α.
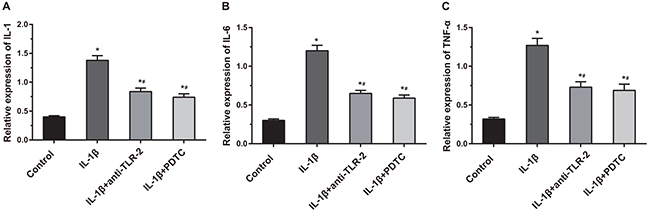
Figure 8: Comparisons of related inflammatory factors (IL-1, IL-6 and TNF-α) among four groups. Notes: (A) Histogram of IL-1; (B) histogram of IL-6; (C) histogram of TNF-α; *, compared with the control group, P < 0.05; #, compared with the IL-1β group, P < 0.05.
DISCUSSION
Progressive loss of articular cartilage in OA patients was recognized to be attributed to an interactional imbalance of anabolic, anti-catabolic, anti- and pro-inflammatory and anti- and pro-apoptotic activities [27–29]. By comparing the normal and OA cartilage tissues and cells, we explored the role of TLR-2/NF-κB signaling pathway in OA and the expressions of related inflammatory factors, with the main conclusion that expressions of TLR-2, NF-κB and MMP-13 and related inflammatory factors were up-regulated with the increase of degree of OA lesions, indicating that TLR-2/NF-κB signaling pathway can contribute to occurrence of OA.
Our study has illustrated that the positive expressions of TLR, NF-kB, MMP13 were associated with BMI, family history, exercise, and WOMAC scores of OA patients. As a degenerative joint disease, OA is characterized by progressive articular cartilage destruction, which leads to pain, joint swelling and reduced mobility [7]. Chondrocytes, stimulated by cytokines and growth factors, are the sole cells residing in articular cartilage, and maintain ECM through a homeostatic balance between anabolic and catabolic activities [30, 31]. Aberrances in the patterns of gene expression of chondrocytes, including expression increase of catabolic cytokines and degradative enzymes, which can cause destruction of the ECM [32]. Inflammation has been implicated in the pathogenesis of OA by shifting the balance from the anabolic toward catabolic state with progressive cartilage loss [33]. IL-1β, a pro-inflammatory cytokine, is recognized as a primary instigator in cartilage degradation in OA [34, 35]. Previous studies have illustrated that TLR2 expression increases in OA chondrocytes [24, 36]. IL-1β functions through autocrine and paracrine signaling pathways to promote synthesis of degradative enzymes, including MMPs and aggrecanases, which break down ECM actively [37]. MMP-13 can degrade aggrecan and the proteoglycan molecule, playing a dual role in matrix destruction [38]. Increased IL-1β expression is primary course for OA [39, 40] which may be a marker of inflammation in OA; therefore, it is not difficult to understand the increase of MMP-13 in OA, as well as related inflammatory factors, including IL-1, IL-6 and TNF-α. As involved in immune responses, TLR-2 and NF-κB have no reason not to be found up-regulated in OA, an inflammatory environment. And it is self-explanatory that with the increase of the degree of the lesions, the expressions of TLR-2, NF-κB and the related inflammatory factors also increased. TLR-2 was reported to be expressed by articular chondrocytes and the expressions were increased in OA lesions [41].
To further explore the effects of TLR-2 and NF-κB on OA, we introduced IL-1β inducement. Observed from our results, the chondrocytes in the IL-1β group, IL-1β + anti-TLR-2 group and IL-1β + PDTC group were significantly inhibited in growth compared with the control group; while the chondrocytes in the IL-1β + anti-TLR-2 and IL-1β + PDTC groups showed faster cell growth compared with those in the IL-1β group. In addition, the apoptosis rate of the chondrocytes in the IL-1β + anti-TLR-2 and IL-1β + PDTC groups decreased significantly compared with the IL-1β group. Furthermore, the cell supernatant in the IL-1β + anti-TLR-2 and IL-1β + PDTC groups showed decreased expressions of IL-1, IL-6 and TNF-α, compared with that in the IL-1β group. IL-1β has been demonstrated to inhibit matrix protein synthesis and induce matrix degrading enzymes and other pro-inflammatory cytokines, including IL-6 [42, 43]. A previous study showed that mice with IL-1β or IL-6 knockout suffered from accelerated OA development, indicating their critical role in cartilage biology [33]. Ortved KF et al. used posttranscriptional silencing of IL-1β to control the catabolic cascade involved in OA and the activation of the catabolic cascade is crucial for degradation of cartilage [32]. Therefore, we used IL-1β inducement to help compare and observe the effects of TLR-2 and NF-κB on OA.
OA is characterized by degeneration of the articular cartilage [6]. The findings in our study suggested that TLR-2 and NF-κB could contribute to suppressing the growth and promoting the apoptosis of chondrocytes, indicating that inhibiting TLR-2/NF-κB signaling pathway might improve chondrocytes viability and antagonize chondrocytes damage induced by IL-1β. IL-1β has been implicated in the degeneration of articular cartilage for its induction of proteoglycan loss and matrix degradation [44]. It was reported that acute injury induced production of pro-inflammatory cytokines and catabolic enzymes, promoting chondrocyte apoptosis and degraded cartilage [45]. Pro-inflammatory cytokines, like IL-1β and TNF-α, were demonstrated to differentially regulate the apoptotic pathway in human chondrocytes [46]. TLR ligands could activate the synoviocytes, which could secrete pro-inflammatory cytokines and chemokines, such as TNF-α, IL-15, RANTES (regulated on activation normal T-cell expressed and secreted) and granulocyte and monocyte chemotactic protein 2, contributing to synovitis maintenance and inflammatory cell infiltration [47–49]. Activated synoviocytes could also produce MMPs and RANKL (receptor activator of NF-κB ligand), which are involved in cartilage degradation and joint destruction [50–52]. Therefore, TLRs may be critical in mediating synovial inflammation in experimental arthritis [53]. Zhu W et al. demonstrated that the induction of TLR-3 in fibroblast-like synoviocytes (FLSs) was due to T cell-derived inflammatory stimulation and further mediated FLS activation in arthritis [47].
Stimulation of TLRs with their ligands could activate NF-κB [54]. NF-κB activation is known to be pro-inflammatory [17]. And the suppression of NF-κB has been related with anti-inflammatory activity [55]. Accumulating studies reported that TLR-mediated responses were key inducers of pro-inflammatory cytokines, which directly or indirectly resulted in development of lymphatic pathology [56–59]. Studies reported that ligands for TLRs (including TLR-2, 3, 4, 9) induced or exacerbated arthritis in experimental models[52, 60]. NF-κB has been reported to regulate expressions of genes involved in OA [61]. TLR-2/NF-κB signaling pathway has been implicated in many studies related with inflammation or immune [62–64]. The specific mechanism behind how it affected the proliferation and apoptosis of chondrocytes was not elucidated yet and needed further exploration.
In conclusion, the expressions of TLR-2, NF-κB and MMP-13 and related inflammatory factors increased with the increase of degree of OA lesions, indicating that TLR-2/NF-κB signaling pathway can contribute to occurrence of OA; inhibiting TLR-2/NF-κB signaling pathway. However, the inner mechanisms of related inflammatory factors and degenerative OA are still unclear, more researches are needed to be conducted to provide a target for the degenerative OA treatment.
MATERIALS AND METHODS
Ethic statement
The study was approved by the Ethics Committee of Huaihe Hospital of Henan University and all the study subjects have signed an informed consent.
Sample collection
A total of 231 patients with degenerative OA receiving total knee arthroplasty (TKA) in Huaihe Hospital of Henan University from January 2013 to December 2015 were selected for our study. The diagnosis was in accordance with the diagnostic criteria for knee OA in 2001 revised by American College of Rheumatology (ACR; formerly, the American Rheumatism Association) [65]. Among these 231 patients, there were 79 males and 152 females, with age ranging from 36 to 96 years old and the mean age 65.42 ± 8.74 years old. Clinical criteria: (1) Most of time nearly 1 with knee pain; (2) With bony crepitus; (3) early morning stiffness ≤ 30 min; (4) Age ≥ 38; (5) With bony enlargement. Clinical and radiological criteria: (1) Most of time nearly 1 with knee pain; (2) X-ray forming osteophyte; (3) Joint fluid examination consistent with OA; (4) Age ≥ 40; (5) early morning stiffness ≤ 30 min; (6) With bony crepitus. Knee OA can be diagnosed when conforming to 1 + 2 or 1 + 3 + 5 + 6 or 1 + 4 + 5 + 6. The articular cartilage samples were collected from patients with traumatic amputation as OA group. Totally 198 patients received amputation because of trauma in the Emergency Department of our hospital, including 76 males and 122 females, with age ranging from 38 to 81 years old and the mean age 64.03 ± 7.70 years old. The articular cartilages were collected for sampling as the normal group. Patients with obvious osteoporosis or neoplastic lesion or non-primary arthritis, for example, rheumatoid arthritis, suppurative arthritis or trauma-induced arthritis, were excluded.
Histological staining
Cartilage block with subchondral bone was cut into 1.0 cm × 1.0 cm × 0.5 cm sized blocks, which were then fixed for 3 days with neutral formalin solution, followed by decalcification for 14 days in 30% formic acid solution, dehydration with ethanol in conventionally gradient, paraffin embedding, cutting into 5 μm slices and drying for further use. The Hematoxylin-Eosin (HE) staining was performed as follows. After dewaxing and hydration, the cartilage samples were stained for 5 min in Harris alum hematoxylin (Fuzhou Maixin Biotechnology, Co., Ltd., Fuzhou, China), followed by washing and color separation for 10 s in 0.5% hydrochloric acid alcohol. After washing, the samples were stained for 40 s in eosin (Fuzhou Maixin Biotechnology, Co., Ltd., Fuzhou, China), followed by dehydration, transparency, mounting with neutral balsam. Observed under a microscope, the nucleus of the chondrocytes appeared blue and the other tissues appeared pink. The procedures of safranin O/fast green FCF staining: After dewaxed and hydrated, the cartilage samples were stained for 5 min in Harris alum hematoxylin, followed by washing and color separation for 10 s in 0.5% hydrochloric acid alcohol. After washing again, the samples were placed in 0.2% fast green solution (Shanghai Sangon Biological Engineering Technology & Services Co., Ltd., Shanghai, China) for 1 min, 1% ethylic acid solution for 30 s and 0.1% safranin O solution (Shanghai Sangon Biological Engineering Technology & Services Co., Ltd.) for 15 min, followed by dehydration, transparency and mounting with neutral balsam. The normal cartilage appeared red and the background appeared green. According to the improved Mankin cartilage pathological score standard [66], the articular cartilage samples were graded from four aspects, namely cartilage structure, cartilage cell morphology, the results of safranin O/fast green FCF staining and tidal line, as the normal group (0 ~ 2 points), the OA mild lesion group (3 ~ 7 points) and the OA moderate lesion group (8 ~ 11 points).
Immunohistochemistry
After fixation and decalcification, the samples were cut into conventional slices, followed by dewaxing with xylene and gradient alcohol hydration. The slices were added with drops of 3% H2O2 and then rested for 15 min to block endogenous peroxidase, followed by washing with phosphate buffered saline (PBS) for three times. The cartilage slices were added with heated sodium citrate buffer (10 mM) and microwaved for 6 min for antigen repair. The procedure was repeated twice, followed by cooling and washing with PBS. The cartilage slices were then treated with 5% goat serum for 10 min, in order to obstruct the binding of nonspecific antibody. After added with drops of the first antibodies of TLR-2, NF-κB and matrix metalloprotease-13 (MMP-13) (Cell Signaling Technologies, Beverly, MA, USA), the slices were incubated at 37°C for 30 min and washed with PBS. After being added with horse radish peroxidase labelled second antibody (Cell Signaling Technologies, Beverly, MA, USA), the slices were incubated at 37°C for 30 min and washed with PBS, followed by staining for 10 min with diaminobenzidine (DAB) (Sigma-Aldrich Chemical Company, St Louis MO, USA), washing with running water, re-staining with hematoxylin, mounting with neutral balsam and lastly observing under a microscope and picture-taking.
Quantitative real-time polymerase chain reaction (qRT-PCR)
The tissue samples were cut into thin slices and then grinded using a liquid nitrogen method. According to the reagent specification of Trizol (Invitrogen Inc., Carlsbad, CA, USA), the total RNA of the tissue samples were extracted in Trizol one-step method. Reverse transcription of the RNA was performed in a two-step method following the reagent specification (Fermentas Inc., Hanover, MD, USA). The reaction conditions were as follows: 70°C for 10 min, ice-bath for 2 min, 42°C for 60 min and 70°C for 10 min. The cDNA obtained was reserved temporarily in a refrigerator at -80°C. The qRT-PCR was carried out using TaqMan probing method. The reaction system was operated according to the reagent specification (Fermentas Inc., Hanover, MD, USA). The primer sequences are shown in Table 4. The reaction conditions: pre-denaturation at 95°C for 30 s, denaturation at 95°C for 10 s, annealing at 60°C for 20 s and extension at 70°C for 10 s, with totally 40 cycles. qRT-PCR instrument (Bio-Rad, Inc., Hercules, CA, USA; model: Bio-Rad iQ5) was applied for detection, with β-actin as an internal reference for target gene, relative quantitative method for calculating and 2-ΔΔCt representing relative expression multiple of each target gene. Each experiment was repeated 5 times.
Table 4: Primer sequences of qRT-PCR
Target |
Primer sequence |
|---|---|
TLR-2 |
F: 5’-CCTGTGCAATTTGACCATTG-3’ |
R: 5’-AAGCATTCCCACCTTTGTTG-3’ |
|
NF-κB |
F: 5’-CCTGGATGACTCTTGGGAAA-3’ |
R: 5’-TCAGCCAGCTGTTTCATGTC-3’ |
|
MMP-13 |
F: 5’-TGAGAGTCATGCCAACAAATTC-3’ |
R: 5’-CAGCCACGCATAGTCATGTAGA-3’ |
|
IL-1 |
F: 5’-GGACAAGCTGAGGAAGATGC-3’ |
R: 5’-TCCATATCCTGTCCCTGGAG-3’ |
|
IL-6 |
F: 5’-GAGCTTCAGGCAGGCAGTATC-3’ |
R: 5’-GTATAGATTCTTTCCTTTGAGGC-3’ |
|
TNF-α |
F: 5’-TCAGAGGGCCTGTACCTCAT-3’ |
R: 5’-GGAAGACCCCTCCCAGATAG-3’ |
|
β-actin |
F: 5’-AGCGAGCATCCCCCAAAGYY-3’ |
R: 5’-GGGCACGAAGGCTCATCATT-3’ |
Note: qRT-PCR, quantitative real-time polymerase chain reaction; TLR-2, toll-like receptor-2; NF-κB, nuclear transcription factor kappa B; MMP-13, matrix metalloprotease-13; IL-1, interleukin-1; IL-6, interleukin-6; TNF-α, tumor necrosis factor alpha; F, forward; R, reverse.
Isolation, culture and detection of chondrocytes
The cartilage collected from the OA patients in the surgery was cut into sections sized 1 ~ 2 mm3, followed by PBS washing and 0.25% type II collagen enzyme (Sigma-Aldrich Chemical Company) digestion overnight. The digestive juice was filtered by a 200 mesh cell filter (Corning Glass Works, Corning, N.Y., USA), and then the chondrocytes were centrifuged at 1000g for 5 min. After discarding the supernatant, the cells were added with Dulbecco minimum essential medium (DMEM) containing 10% fetal bovine serum (FBS) (Gibco Company, Grand Island, NY, USA), followed by suspension and culture at 37°C with 5% CO2. After growing full of the incubator, the chondrocytes were digested with 0.25% pancreatic enzyme (Gibco Company) and cultured for passage. Well-grown chondrocytes were collected to make climbing films, which were stained with toluidine blue (Beijing Solarbio Science & Technology Co., Ltd., Beijing, China) according to the reagent specification of immunochemistry staining, and labelled with type II collagen (USCN Life Science & Technology Company, Double Lake, Missouri City, TX, USA), followed by dehydration with absolute alcohol, mounting with neutral balsam and observing under a microscope and picture-taking.
Cell grouping
The isolated and cultured chondrocytes were inoculated into corresponding plates or culture dishes at an appropriate density at 37°C with 5% CO2. After adherent, the cells were treated for 48 h with antibody of interleukin-1β (IL-1β; at 10 ng/ml; Sigma-Aldrich Chemical Company) and TLR-2 (at 50 ng/ml; Sigma-Aldrich Chemical Company), and NF-κB inhibitor pyrrolidinedithiocarbamic acid (PDTC; 100 uM; Sigma-Aldrich Chemical Company). The cells were divided into 4 groups, namely the control group, the IL-1β group, the IL-1β + anti-TLR-2 group and the IL-1β + PDTC group.
MTT assay
The chondrocytes were inoculated into a 96-well plate (104 cells in each well). The three experiment groups were treated respectively with anti-TLR-2, NF-κB inhibitor PDTC and IL-1β; and three wells were set for each group. After being incubated for a period of time at 37°C with 5% CO2, each well was added with 20 μL MTT solution (5 mg/ml; Sigma-Aldrich Chemical Company) and cultured for another 4 h. After the supernatant was discarded, each well was added with 150 μL of dimethylsulfoxide (DMSO), followed by low-speed shaking for 10 min. Enzyme-linked immunosorbent assay (ELISA) was performed to detect the optical density (OD) value at the wavelength of 490 nm.
Flow cytometry
After treated for 48 h, the cells were collected into flow tubes and then centrifuged for 5 min at 1000r/min, and then the supernatant was discarded. The cells were washed with cooled PBS for three times and centrifuged, and then the supernatant was discarded. According to the specification of Annexin-V-FITC cell apoptosis detection Kit (Sigma-Aldrich Chemical Company), each tube was added with 150 μL of binding buffer and 5 μL of Annexin-V-FITC, followed by shaking and mixing, and incubation for 15 min in dark at room temperature. The addition of 100 μL of binding buffer and 5 μL of propidium iodide (PI; Sigma-Aldrich Chemical Company) and evenly mixing were conducted. A flow cytometry was used to determine cell apoptosis.
Western blotting
The chondrocytes treated for 48 h were collected and washed for three times with pre-cooling PBS, then lysed with protein extraction lysis solution (Beyotime Biotechnology Co., Shanghai, China) and lastly rested on ice for 30 min, followed by centrifugation at 12000g for 10 min at 4°C, separate packing and reservation at -20°C. Bovine Serum Albumin (BSA) protein with gradient density (Beijing Solarbio Science & Technology Co., Ltd., Beijing, China) was prepared; the protein concentration of each sample was determined according to Pierce BCA Protein Assay Kit (Thermo Fisher Scientific Inc., Waltham, MA). Electrophoresis was performed firstly at 60 V and then 120 V after the protein samples entered into separation gel, for 1 ~ 2 h, at 4°C in a cold room. After electrophoresis, the protein samples were transferred to polyvinylidene fluoride (PVDF) membrane using wet transferring method, for 2 h, at 4°C in a cold room. After removing the PVDF membrane, the protein samples were sealed with 5% evaporated milk and Tris-buffered saline-Tween 20 (TBST), followed by incubation for 1 ~ 2 h at room temperature. After added with the first antibodies of TLR-2, NF-κB, phosphorylated-NF-κB (p-NF-κB) and MMP-13 (Cell Signaling Technologies), the protein samples were rested overnight at 4°C, followed by washing with TBST (3 × 10 min), addition of mouse second antibody (Abcam Inc., Cambridge, MA, USA), resting for 1 h at room temperature and washing with TBST (3 × 10 min). Chemiluminescence, X-ray film tableting, developing and fixing were carried out. All the bands were analyzed for relative OD value, with the ratio of the integrated OD value of the target band and the integrated OD value of the corresponding internal reference GAPDH as the relative expression of the target protein.
ELISA
ELISA was performed to determine the cell inflammation related factors, including IL-1, IL-6 and TNF-α. The chondrocytes at the logarithmic growth phase were inoculated into a 96-well plate. After the experiment groups were treated for 48 h, the cell supernatant was collected for detection. After added with diluted 100 μL of standard or sample to be tested and completely mixed, the cell supernatant was cultured for 120 min at 37°C. The reaction plate was fully washed and then dried. Each well was then added with 100 μL of enzyme conjugate (IL-1, IL-6 and TNF-α) (Beijing QiWei YiCheng Tech Co., Ltd.), mixed and then cultured in dark for 10 min at 37°C. Each well was added with 100 μL of stop buffer. A microplate reader was applied to detect the OD value at the wavelength of 450 nm; the concentrations of IL-1, IL-6 and TNF-α were determined according to the standard curve.
Statistical analysis
All the data were analyzed using SPSS 21.0 statistic software (IBM-SPSS Inc., Chicago, IL, USA). Measurement data were presented as mean ± standard deviation; comparison among groups was analyzed using one-step variance; comparison between two groups was testified using LSD-t. Enumeration data were presented as frequency. Comparison of categorical data was validated by chi-square and hierarchical data by rank-sum test. Logistic regression analyzed factors related to OA. P < 0.05 was considered significantly different.
ACKNOWLEDGMENTS
We would like to give our sincere appreciation to the reviewers for their helpful comments on this article.
COMPETING INTERESTS
The authors have declared that no competing interests exist.
REFERENCES
1. Mobasheri A, Csaki C, Clutterbuck AL, Rahmanzadeh M, Shakibaei M. Mesenchymal stem cells in connective tissue engineering and regenerative medicine: Applications in cartilage repair and osteoarthritis therapy. Histol Histopathol. 2009; 24: 347-366.
2. Fransen M, Bridgett L, March L, Hoy D, Penserga E, Brooks P. The epidemiology of osteoarthritis in asia. Int J Rheum Dis. 2011; 14: 113-121.
3. Pabinger C, Lothaller H, Geissler A. Utilization rates of knee-arthroplasty in oecd countries. Osteoarthritis Cartilage. 2015; 23: 1664-1673.
4. Bruyere O, Cooper C, Arden N, Branco J, Brandi ML, Herrero-Beaumont G, Berenbaum F, Dennison E, Devogelaer JP, Hochberg M, Kanis J, Laslop A, McAlindon T, et al. Can we identify patients with high risk of osteoarthritis progression who will respond to treatment? A focus on epidemiology and phenotype of osteoarthritis. Drugs Aging. 2015; 32: 179-187.
5. Li G, Yin J, Gao J, Cheng TS, Pavlos NJ, Zhang C, Zheng MH. Subchondral bone in osteoarthritis: Insight into risk factors and microstructural changes. Arthritis Res Ther. 2013; 15: 223.
6. Gierman LM, van El B, van der Ham F, Koudijs A, Stoop R, Verheijen JH, Kloppenburg M, van Osch GJ, Stojanovic-Susulic V, Huizinga TW, Zuurmond AM. Profiling the secretion of soluble mediators by end stage osteoarthritis synovial tissue explants reveals a reduced responsiveness to an inflammatory trigger. PLoS One. 2013; 8: e62634.
7. Zhu Y, Yuan M, Meng HY, Wang AY, Guo QY, Wang Y, Peng J. Basic science and clinical application of platelet-rich plasma for cartilage defects and osteoarthritis: A review. Osteoarthritis Cartilage. 2013; 21: 1627-1637.
8. Hochberg MC, Altman RD, April KT, Benkhalti M, Guyatt G, McGowan J, Towheed T, Welch V, Wells G, Tugwell P, American College of R. American college of rheumatology 2012 recommendations for the use of nonpharmacologic and pharmacologic therapies in osteoarthritis of the hand, hip, and knee. Arthritis Care Res (Hoboken). 2012; 64: 465-474.
9. Conaghan PG, Dickson J, Grant RL, Guideline Development G. Care and management of osteoarthritis in adults: Summary of nice guidance. BMJ. 2008; 336: 502-503.
10. Round JL, Lee SM, Li J, Tran G, Jabri B, Chatila TA, Mazmanian SK. The toll-like receptor 2 pathway establishes colonization by a commensal of the human microbiota. Science. 2011; 332: 974-977.
11. Barton GM, Kagan JC. A cell biological view of toll-like receptor function: Regulation through compartmentalization. Nat Rev Immunol. 2009; 9: 535-542.
12. Salomao R, Brunialti MK, Gomes NE, Mendes ME, Diaz RS, Komninakis S, Machado FR, da Silva ID, Rigato O. Toll-like receptor pathway signaling is differently regulated in neutrophils and peripheral mononuclear cells of patients with sepsis, severe sepsis, and septic shock. Crit Care Med. 2009; 37: 132-139.
13. Reijmerink NE, Bottema RW, Kerkhof M, Gerritsen J, Stelma FF, Thijs C, van Schayck CP, Smit HA, Brunekreef B, Koppelman GH, Postma DS. Tlr-related pathway analysis: Novel gene-gene interactions in the development of asthma and atopy. Allergy. 2010; 65: 199-207.
14. Monaco C, Gregan SM, Navin TJ, Foxwell BM, Davies AH, Feldmann M. Toll-like receptor-2 mediates inflammation and matrix degradation in human atherosclerosis. Circulation. 2009; 120: 2462-2469.
15. Reynolds JM, Pappu BP, Peng J, Martinez GJ, Zhang Y, Chung Y, Ma L, Yang XO, Nurieva RI, Tian Q, Dong C. Toll-like receptor 2 signaling in cd4(+) t lymphocytes promotes t helper 17 responses and regulates the pathogenesis of autoimmune disease. Immunity. 2010; 32: 692-702.
16. Huang G, Shi LZ, Chi H. Regulation of jnk and p38 mapk in the immune system: Signal integration, propagation and termination. Cytokine. 2009; 48: 161-169.
17. Carmody RJ, Chen YH. Nuclear factor-kappab: Activation and regulation during toll-like receptor signaling. Cell Mol Immunol. 2007; 4: 31-41.
18. Chen AC, Arany PR, Huang YY, Tomkinson EM, Sharma SK, Kharkwal GB, Saleem T, Mooney D, Yull FE, Blackwell TS, Hamblin MR. Low-level laser therapy activates nf-kb via generation of reactive oxygen species in mouse embryonic fibroblasts. PLoS One. 2011; 6: e22453.
19. Karin M. Nuclear factor-kappab in cancer development and progression. Nature. 2006; 441: 431-436.
20. De Simone V, Franze E, Ronchetti G, Colantoni A, Fantini MC, Di Fusco D, Sica GS, Sileri P, MacDonald TT, Pallone F, Monteleone G, Stolfi C. Th17-type cytokines, il-6 and tnf-alpha synergistically activate stat3 and nf-kb to promote colorectal cancer cell growth. Oncogene. 2015; 34: 3493-3503.
21. Santini D, Schiavon G, Vincenzi B, Gaeta L, Pantano F, Russo A, Ortega C, Porta C, Galluzzo S, Armento G, La Verde N, Caroti C, Treilleux I, et al. Receptor activator of nf-kb (rank) expression in primary tumors associates with bone metastasis occurrence in breast cancer patients. PLoS One. 2011; 6: e19234.
22. Galardi S, Mercatelli N, Farace MG, Ciafre SA. Nf-kb and c-jun induce the expression of the oncogenic mir-221 and mir-222 in prostate carcinoma and glioblastoma cells. Nucleic Acids Res. 2011; 39: 3892-3902.
23. Kim KW, Cho ML, Lee SH, Oh HJ, Kang CM, Ju JH, Min SY, Cho YG, Park SH, Kim HY. Human rheumatoid synovial fibroblasts promote osteoclastogenic activity by activating rankl via tlr-2 and tlr-4 activation. Immunol Lett. 2007; 110: 54-64.
24. Sillat T, Barreto G, Clarijs P, Soininen A, Ainola M, Pajarinen J, Korhonen M, Konttinen YT, Sakalyte R, Hukkanen M, Ylinen P, Nordstrom DC. Toll-like receptors in human chondrocytes and osteoarthritic cartilage. Acta Orthop. 2013; 84: 585-592.
25. Montaseri A, Busch F, Mobasheri A, Buhrmann C, Aldinger C, Rad JS, Shakibaei M. Igf-1 and pdgf-bb suppress il-1beta-induced cartilage degradation through down-regulation of nf-kappab signaling: Involvement of src/pi-3k/akt pathway. PLoS One. 2011; 6: e28663.
26. Neuhold LA, Killar L, Zhao W, Sung ML, Warner L, Kulik J, Turner J, Wu W, Billinghurst C, Meijers T, Poole AR, Babij P, DeGennaro LJ. Postnatal expression in hyaline cartilage of constitutively active human collagenase-3 (mmp-13) induces osteoarthritis in mice. J Clin Invest. 2001; 107: 35-44.
27. Gierman LM, van El B, van der Ham F, Koudijs A, Stoop R, Verheijen JH, Kloppenburg M, van Osch GJ, Stojanovic-Susulic V, Huizinga TW, Zuurmond AM. Profiling the secretion of soluble mediators by end stage osteoarthritis synovial tissue explants reveals a reduced responsiveness to an inflammatory trigger. PLoS One. 2013; 8: e62634.
28. Hunter DJ, Felson DT. Osteoarthritis. BMJ. 2006; 332: 639-642.
29. Musumeci G, Loreto C, Carnazza ML, Martinez G. Characterization of apoptosis in articular cartilage derived from the knee joints of patients with osteoarthritis. Knee Surg Sports Traumatol Arthrosc. 2011; 19: 307-313.
30. Miyaki S, Nakasa T, Otsuki S, Grogan SP, Higashiyama R, Inoue A, Kato Y, Sato T, Lotz MK, Asahara H. Microrna-140 is expressed in differentiated human articular chondrocytes and modulates interleukin-1 responses. Arthritis Rheum. 2009; 60: 2723-2730.
31. Alexopoulos LG, Youn I, Bonaldo P, Guilak F. Developmental and osteoarthritic changes in col6a1-knockout mice: Biomechanics of type vi collagen in the cartilage pericellular matrix. Arthritis Rheum. 2009; 60: 771-779.
32. Ortved KF, Austin BS, Scimeca MS, Nixon AJ. Rna interference mediated interleukin-1beta silencing in inflamed chondrocytes decreases target and downstream catabolic responses. Arthritis. 2016; 2016: 3484961. doi: 10.1155/2016/3484961.
33. Choi YS, Park JK, Kang EH, Lee YK, Kim TK, Chung JH, Zimmerer JM, Carson WE, Song YW, Lee YJ. Cytokine signaling-1 suppressor is inducible by il-1beta and inhibits the catabolic effects of il-1beta in chondrocytes: Its implication in the paradoxical joint-protective role of il-1beta. Arthritis Res Ther. 2013; 15: R191.
34. Montaseri A, Busch F, Mobasheri A, Buhrmann C, Aldinger C, Rad JS, Shakibaei M. Igf-1 and pdgf-bb suppress il-1beta-induced cartilage degradation through down-regulation of nf-kappab signaling: Involvement of src/pi-3k/akt pathway. PLoS One. 2011; 6: e28663.
35. Santangelo KS, Bertone AL. Effective reduction of the interleukin-1beta transcript in osteoarthritis-prone guinea pig chondrocytes via short hairpin rna mediated rna interference influences gene expression of mediators implicated in disease pathogenesis. Osteoarthritis Cartilage. 2011; 19: 1449-1457.
36. Kim HA, Cho ML, Choi HY, Yoon CS, Jhun JY, Oh HJ, Kim HY. The catabolic pathway mediated by toll-like receptors in human osteoarthritic chondrocytes. Arthritis Rheum. 2006; 54: 2152-2163.
37. Goldring MB, Berenbaum F. The regulation of chondrocyte function by proinflammatory mediators: Prostaglandins and nitric oxide. Clin Orthop Relat Res. 2004; S37-46.
38. Burrage PS, Mix KS, Brinckerhoff CE. Matrix metalloproteinases: Role in arthritis. Front Biosci. 2006; 11: 529-43.
39. Collodel G, Fioravanti A, Pascarelli NA, Lamboglia A, Fontani V, Maioli M, Santaniello S, Pigliaru G, Castagna A, Moretti E, Iacoponi F, Rinaldi S, Ventura C. Effects of regenerative radioelectric asymmetric conveyer treatment on human normal and osteoarthritic chondrocytes exposed to il-1beta. A biochemical and morphological study. Clin Interv Aging. 2013; 8:309-16. doi: 10.2147/CIA.S42229.
40. Xiang Y, Masuko-Hongo K, Sekine T, Nakamura H, Yudoh K, Nishioka K, Kato T. Expression of proteinase-activated receptors (par)-2 in articular chondrocytes is modulated by il-1beta, tnf-alpha and tgf-beta. Osteoarthritis Cartilage. 2006; 14: 1163-1173.
41. Su SL, Tsai CD, Lee CH, Salter DM, Lee HS. Expression and regulation of toll-like receptor 2 by il-1beta and fibronectin fragments in human articular chondrocytes. Osteoarthritis Cartilage. 2005; 13: 879-886.
42. Kapoor M, Martel-Pelletier J, Lajeunesse D, Pelletier JP, Fahmi H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat Rev Rheumatol. 2011; 7: 33-42.
43. Daheshia M, Yao JQ. The interleukin 1beta pathway in the pathogenesis of osteoarthritis. J Rheumatol. 2008; 35: 2306-2312.
44. Kobayashi M, Squires GR, Mousa A, Tanzer M, Zukor DJ, Antoniou J, Feige U, Poole AR. Role of interleukin-1 and tumor necrosis factor alpha in matrix degradation of human osteoarthritic cartilage. Arthritis Rheum. 2005; 52: 128-135.
45. Hu Z, Yik JH, Cissell DD, Michelier PV, Athanasiou KA, Haudenschild DR. Inhibition of cdk9 prevents mechanical injury-induced inflammation, apoptosis and matrix degradation in cartilage explants. Eur Cell Mater. 2016; 30: 200-209.
46. Lopez-Armada MJ, Carames B, Lires-Dean M, Cillero-Pastor B, Ruiz-Romero C, Galdo F, Blanco FJ. Cytokines, tumor necrosis factor-alpha and interleukin-1beta, differentially regulate apoptosis in osteoarthritis cultured human chondrocytes. Osteoarthritis Cartilage. 2006; 14: 660-669.
47. Zhu W, Meng L, Jiang C, He X, Hou W, Xu P, Du H, Holmdahl R, Lu S. Arthritis is associated with t-cell-induced upregulation of toll-like receptor 3 on synovial fibroblasts. Arthritis Res Ther. 2011; 13: R103.
48. Jung YO, Cho ML, Kang CM, Jhun JY, Park JS, Oh HJ, Min JK, Park SH, Kim HY. Toll-like receptor 2 and 4 combination engagement upregulate il-15 synergistically in human rheumatoid synovial fibroblasts. Immunol Lett. 2007; 109: 21-27.
49. Brentano F, Schorr O, Gay RE, Gay S, Kyburz D. Rna released from necrotic synovial fluid cells activates rheumatoid arthritis synovial fibroblasts via toll-like receptor 3. Arthritis Rheum. 2005; 52: 2656-2665.
50. Ospelt C, Brentano F, Rengel Y, Stanczyk J, Kolling C, Tak PP, Gay RE, Gay S, Kyburz D. Overexpression of toll-like receptors 3 and 4 in synovial tissue from patients with early rheumatoid arthritis: Toll-like receptor expression in early and longstanding arthritis. Arthritis Rheum. 2008; 58: 3684-3692.
51. Cho ML, Ju JH, Kim HR, Oh HJ, Kang CM, Jhun JY, Lee SY, Park MK, Min JK, Park SH, Lee SH, Kim HY. Toll-like receptor 2 ligand mediates the upregulation of angiogenic factor, vascular endothelial growth factor and interleukin-8/cxcl8 in human rheumatoid synovial fibroblasts. Immunol Lett. 2007; 108: 121-128.
52. Kim KW, Cho ML, Lee SH, Oh HJ, Kang CM, Ju JH, Min SY, Cho YG, Park SH, Kim HY. Human rheumatoid synovial fibroblasts promote osteoclastogenic activity by activating rankl via tlr-2 and tlr-4 activation. Immunol Lett. 2007; 110: 54-64.
53. Marshak-Rothstein A. Toll-like receptors in systemic autoimmune disease. Nat Rev Immunol. 2006; 6: 823-835.
54. Iwasaki A, Medzhitov R. Toll-like receptor control of the adaptive immune responses. Nat Immunol. 2004; 5: 987-995.
55. Akhtar N, Miller MJ, Haqqi TM. Effect of a herbal-leucine mix on the il-1beta-induced cartilage degradation and inflammatory gene expression in human chondrocytes. BMC Complement Altern Med. 2011; 11:66. doi: 10.1186/1472-6882-11-66.
56. Pfarr KM, Debrah AY, Specht S, Hoerauf A. Filariasis and lymphoedema. Parasite Immunol. 2009; 31: 664-672.
57. Venugopal PG, Nutman TB, Semnani RT. Activation and regulation of toll-like receptors (tlrs) by helminth parasites. Immunol Res. 2009; 43: 252-263.
58. Hartgers FC, Obeng BB, Voskamp A, Larbi IA, Amoah AS, Luty AJ, Boakye D, Yazdanbakhsh M. Enhanced toll-like receptor responsiveness associated with mitogen-activated protein kinase activation in plasmodium falciparum-infected children. Infect Immun. 2008; 76: 5149-5157.
59. Turner JD, Langley RS, Johnston KL, Gentil K, Ford L, Wu B, Graham M, Sharpley F, Slatko B, Pearlman E, Taylor MJ. Wolbachia lipoprotein stimulates innate and adaptive immunity through toll-like receptors 2 and 6 to induce disease manifestations of filariasis. J Biol Chem. 2009; 284: 22364-22378.
60. Su SL, Yang HY, Lee CH, Huang GS, Salter DM, Lee HS. The (-1486t/c) promoter polymorphism of the tlr-9 gene is associated with end-stage knee osteoarthritis in a chinese population. J Orthop Res. 2012; 30: 9-14.
61. Bhaskar S, Shalini V, Helen A. Quercetin regulates oxidized ldl induced inflammatory changes in human pbmcs by modulating the tlr-nf-kappab signaling pathway. Immunobiology. 2011; 216: 367-373.
62. Marcu KB, Otero M, Olivotto E, Borzi RM, Goldring MB. Nf-kappab signaling: Multiple angles to target oa. Curr Drug Targets. 2010; 11: 599-613.
63. Chen S, Lin G, Lei L, You X, Wu C, Xu W, Huang M, Luo L, Wang Z, Li Y, Zhao X, Yan F. Hyperlipidemia modifies innate immune responses to lipopolysaccharide via the tlr-nf-kappab signaling pathway. Inflammation. 2013; 36: 968-976.
64. Zhu HT, Bian C, Yuan JC, Chu WH, Xiang X, Chen F, Wang CS, Feng H, Lin JK. Curcumin attenuates acute inflammatory injury by inhibiting the tlr4/myd88/nf-kappab signaling pathway in experimental traumatic brain injury. J Neuroinflammation. 2014; 11:59. doi: 10.1186/1742-2094-11-59.
65. Saraux A, Berthelot JM, Chales G, Le Henaff C, Thorel JB, Hoang S, Valls I, Devauchelle V, Martin A, Baron D, Pennec Y, Botton E, Mary JY, et al. Ability of the american college of rheumatology 1987 criteria to predict rheumatoid arthritis in patients with early arthritis and classification of these patients two years later. Arthritis Rheum. 2001; 44: 2485-2491.
66. Mankin HJ, Johnson ME, Lippiello L. Biochemical and metabolic abnormalities in articular cartilage from osteoarthritic human hips. Iii. Distribution and metabolism of amino sugar-containing macromolecules. J Bone Joint Surg Am. 1981; 63: 131-139.