
INTRODUCTION
Skeletal muscle is a heterogeneous tissue composed of a large variety of fiber types classified according to their myosin heavy chain (MyHC) isoforms. There are four different types of muscle fibers: type I (expressing slow MyHC, namely MyHC I), type IIa (expressing MyHC IIa), type IIb (expressing MyHC IIb), and type IIx (expressing MyHC IIx). Type I fiber, also known as slow oxidative muscle fiber, is believed to be positively associated with meat quality [1, 2]. The intramuscular fat (IMF) content (marbling) is also associated with a higher content of oxidative fibers in skeletal muscle [3].
Akirin2 is a highly conserved nuclear protein [4]. Numerous reports have described the involvement of Akirin2 in the embryonic development and immune defense function [4–7]. However, the role of Akirin2 in skeletal muscle is far less understood. Currently, Akirin2 has been reported to be located within genomic region of a quantitative trait locus for marbling, and has been shown to be differentially expressed in musculus longissimus muscle of low-marbled and high-marbled steer groups [8]. A number of studies showed that Akirin2 is associated with marbling based on single nucleotide polymorphisms analysis and may be considered as a positional functional candidate for the gene responsible for marbling [9–12].
Little research has been conducted on the role of porcine Akirin2 (pAkirin2). The pAkirin2 gene was cloned and its tissue distribution in pig was examined [13]. We also proved that recombinant pAkirin2 significantly increased the proliferation of C2C12 myoblasts [14]. In C2C12 skeletal muscle cells, we found that Akirin2 could increase the mRNA expression of MyHC I and NFATc1 [15]. To further our knowledge of pAkirin2 in skeletal muscle, the aim of this study was to investigate whether Akirin2 modulates MyHC I expression via calcineurin/NFATc1 signaling pathway in porcine skeletal muscle satellite cells.
RESULTS
Endogenous Akirin2 gene expression in skeletal muscle
The endogenous expression of Akirin2 was analyzed in pig skeletal muscle. Western blot analysis with an anti-Akirin2 antibody prepared in our lab showed that Akirin2 protein level in the pig slow oxidative Psoas major muscle (PM) was about six-fold higher than that in the fast glycolytic tibialis anterior muscle (TA) (Figure 1), suggesting that Akirin2 may play a role in muscle fiber typing.
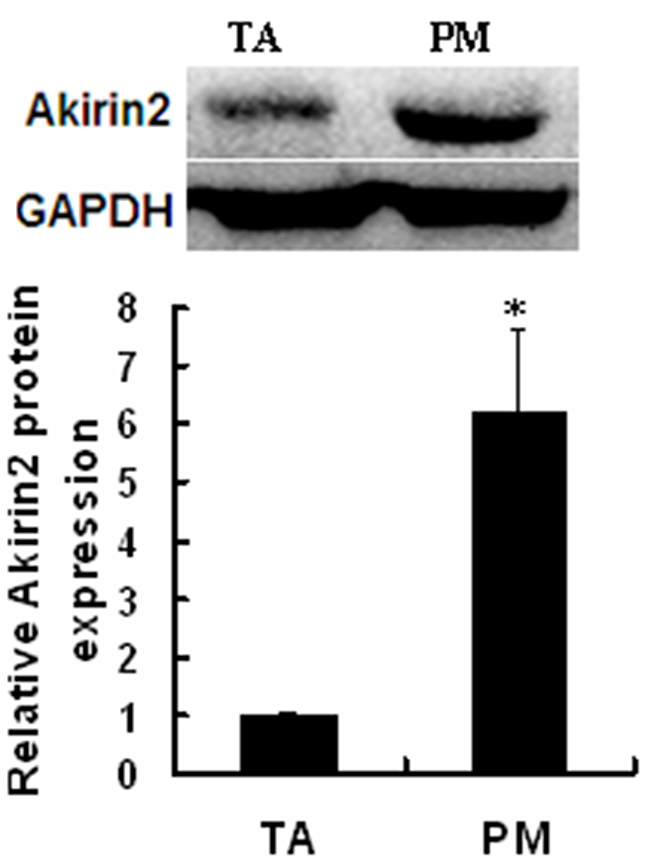
Figure 1: Expression of endogenous Akirin2 gene. Western blot was used to analyze the Akirin2 protein expression in pig slow oxidative Psoas major (PM) muscle and fast glycolytic tibialis anterior (TA) muscle. Data were presented as means ± SE (n=3). *P < 0.05.
Akirin2 promotes MyHC I expression in porcine skeletal muscle satellite cells
To test whether Akirin2 affects MyHC I expression in porcine skeletal muscle satellite cells, RNA interference and gene overexpression technologies were employed. Western blot analysis showed that knockdown of Akirin2 by siRNA significantly repressed the protein expressions of Akirin2 (Figure 2A) and MyHC I (Figure 2C). However, Akirin2 overexpression presented an increase in Akirin2 (Figure 2B) and MyHC I protein expressions (Figure 2D), suggesting that Akirin2 promotes MyHC I expression.
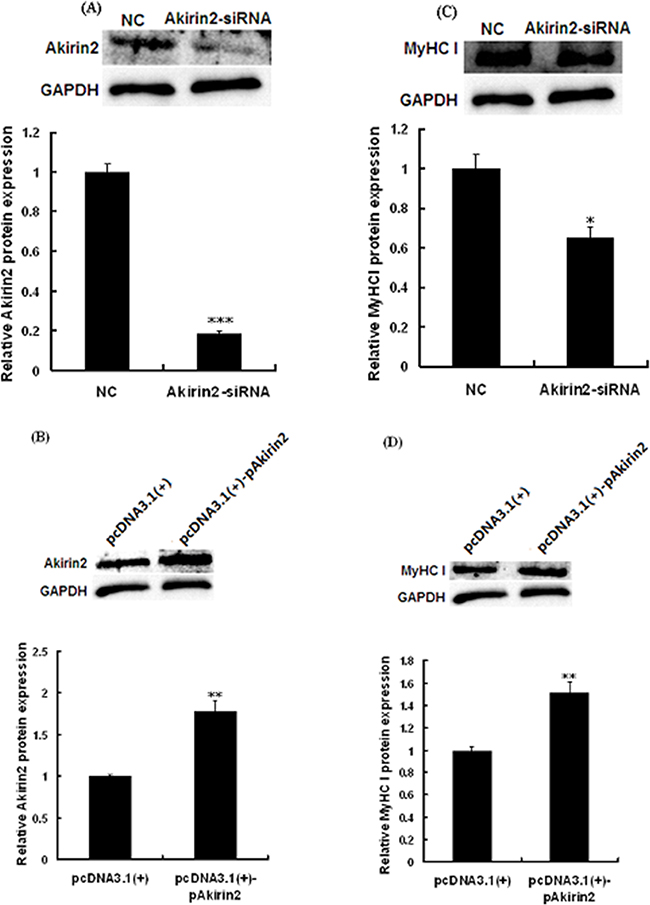
Figure 2: Akirin2 regulates the expression of MyHC I in porcine skeletal muscle satellite cells. Porcine skeletal muscle satellite cells were transfected with negative control siRNA (NC), Akirin2 siRNA, pcDNA3.1(+) or pcDNA3.1(+)-pAkirin2 when cells reached about 80% confluence and induced to differentiate for 6 days before analysis. Western blot analysis was used to analyzed the Akirin2 A-B. and MyHC I C-D. protein expression in porcine skeletal muscle satellite cells. Data were presented as means ± SE (n=3). *P < 0.05 and **P < 0.01 as compared with control.
Effect of Akirin2 on the protein expression of NFATc1 and CaN activity in porcine skeletal muscle satellite cells
As shown in Figure 3A and 3B, silencing of Akirin2 by siRNA and overexpression of Akirin2 significantly decreased and increased modulatory calcineurin interacting protein exon 4 isoform (MCIP1.4, a NFATc1 target) protein expression in porcine skeletal muscle satellite cells, respectively. Analysis also indicated that Akirin2 affected NFATc1 protein expression (Figure 3C-3D), suggesting that NFATc1 is downstream of Akirin2.
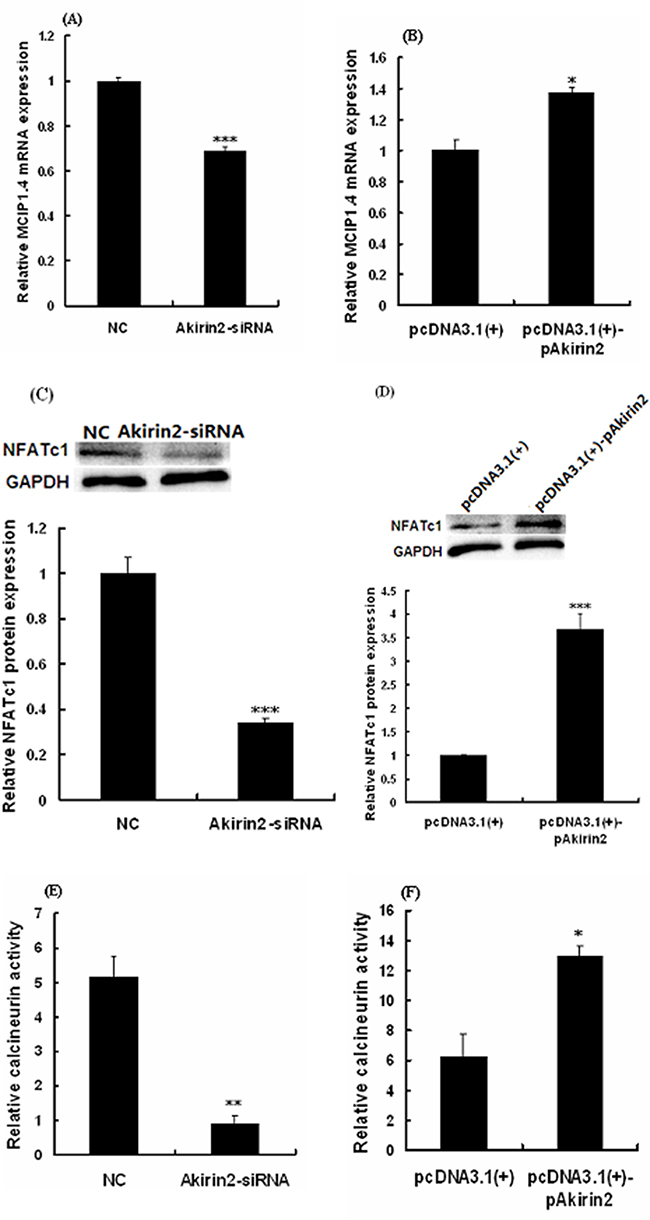
Figure 3: Effects of Akirin2 on the NFATc1 expression and CaN activity. Porcine skeletal muscle satellite cells were cultured, transfected and induced to differentiate as described in Figure 2. Real-time quantitative PCR was used to analyze the MCIP1.4 A-B. mRNA expression in porcine skeletal muscle satellite cells. Western blot was used to analyze to the protein expression of NFATc1 C-D. in porcine skeletal muscle satellite cells. Calcineurin activity was analyzed by Calcineurin Cellular Activity Assay Kit E-F. Data were presented as means ± SE (n=3). *P < 0.05 and ***P < 0.001 as compared with control.
To further determine the relationship between Akirin2 expression and CaN activity, we examined the effects of Akirin2 silencing and overexpression on the CaN activity. The results showed that silencing of Akirin2 significantly decreased the CaN activity (Figure 3E), while overexpression of Akirin2 got the opposite result (Figure 3F).
Akirin2 promotes MyHC I expression via CaN/NFATc1 signaling
To test if NFATc1 was involved in Akirin2-induced promotion ofMyHC I expression, NFATc1 was knocked down by siRNA in porcine skeletal muscle satellite cells, and the expression of MyHC I was analyzed. As shown in Figure 4A and 4B, both the mRNA and protein expression levels of NFATc1 were significantly decreased. As shown in Figure 4C, knockdown of NFATc1 abolished Akirin2 overexpression-induced upregulation of MyHC I.
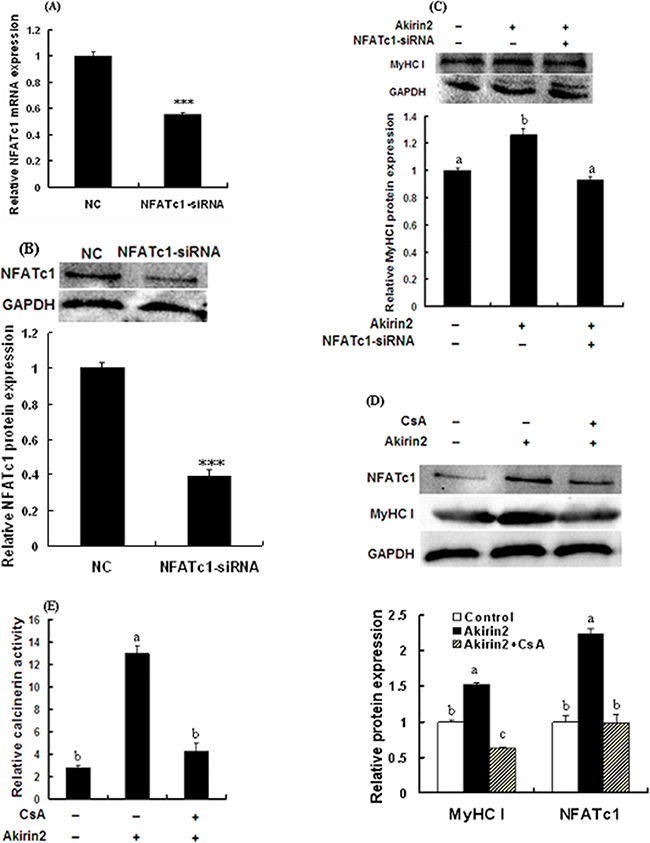
Figure 4: Akirin2 regulates the expression of MyHC I via CaN/NFATc1 signaling in porcine skeletal muscle satellite cells. Cells were cultured, transfected and induced to differentiate as described in Figure 2. Real-time quantitative PCR was used to analyze the NFATc1 mRNA expression in porcine skeletal muscle satellite cells. Western blot was used to analyze to the protein expression of NFATc1 and MyHC I in porcine skeletal muscle satellite cells. Real-time quantitative PCR analysis indicated that the siRNA induced a 45% downregulation of NFATc1 mRNA expression A. Western blot analysis indicated that the protein expression level of NFATc1 was also significantly decreased B. Silencing of NFATc1 by siRNA inhibited Akirin2 overexpression-induced upregulation of MyHC I C. Inhibition of CaN by CsA abolished Akirin2 overexpression-induced upregulation of MyHC I and NFATc1 protein expressions D. and increase in the CaN activity E. Data were presented as means ± SE (n=3). ***P < 0.001. Value with different letters indicates significantly difference (P < 0.05).
As shown in Figure 4D and 4E, CaN inhibitor CsA significantly attenuated Akirin2 overexpression-induced upregulation of NFATc1 and MyHC I expressions and increase in the CaN activity. Taken together, these results showed that Akirin2-mediated upreguation of MyHC I is dependent on CaN/NFATc1 signaling.
DISCUSSION
Meat quality, one of the most important economic traits in farm animals, is controlled by multiple genes and affected by many factors. One of the main factors is the muscle fiber characteristics because skeletal muscles mainly consist of muscle fibers [16]. There is a lot of evidence to suggest that increasing the percentage of type I fibers is related to meat quality improvement [1, 2, 17]. Therefore, it is necessary to identify novel candidate genes for improving meat quality by regulating muscle type I fiber expression.
TA muscle is a typical fast muscle, while PM muscle is a typical slow muscle [18]. We found that higher levels of Akirin2 protein were detected in slow oxidative PM muscle compared to fast glycolytic TA muscle (Figure 1). In the present study, we first evaluated the effect of Akirin2 on MyHC I expression in porcine skeletal muscle satellite cells. The data showed that Akirin2 affected MyHC I in both gene silencing and overexpression of Akirin2 studies (Figure 2). Additional molecular mechanism experiments revealed that Akirin2 acted through CaN/NFATc1 signaling (Figure 3).
Previous studies have described calcineurin-NFAT signaling pathway regulating oxidative muscle fiber type transition in skeletal muscle [19–22]. Calcineurin is activated by calcium/calmodulin and promotes NFAT nuclear translocation where NFAT induces the slow-twitch phenotype in skeletal muscle [22, 23]. The NFAT transcription factor family consists of five members NFATc1, NFATc2, NFATc3, NFATc4 and NFATc5. NFATc1 through NFATc4 are activated by calcineurin and expressed in skeletal muscle [24]. NFATc1 may be control fiber type composition and plays an important role in fast-to-slow fiber type switching [19, 25]. In this study, we present the first evidence that Akirin2 mediated the MyHC I expression via NFATc1 signaling since knockdown of Akirin2 not only downregulated MyHC I, but also decreased the expressions of NFATc1 and its downstream target gene MCIP1.4, whereas overexpression of Akirin2 got the opposite results (Figure 3). The expression level of MCIP1.4 reflects the calcineurin activity and the promoter of MCIP1.4 includes a cluster of NFAT-binding sites, and hence MCIP1.4 appears a direct downstream target of the calcineurin/NFAT pathway [26–28]. Furthermore, an increase in MCIP1.4 mRNA level reflects the switch from fast to slow-type MyHC isoforms [28]. We therefore measured the level of MCIP1.4 mRNA and found overexpression of Akirin2 significantly increased MCIP1.4 mRNA level (Figure 3B), whereas knockdown of Akirin2 got the opposite result (Figure 3A). These results suggested that the MCIP1.4 mRNA level may be mirror the calcineurin activity and MCIP1.4 plays an important role in affecting slow type fiber gene expression. Furthermore, silencing of NFATc1 by siRNA inhibited the effect of Akirin2 overexpression on MyHC I expression (Figure 4C). CaN activity assay and Western blot analysis further indicated that Akirin2 promotes MyHC I expression via CaN/NFATc1 signaling pathway.
In conclusion, we reported that Akirin2 promotes MyHC I expression in porcine skeletal muscle satellite cells. This function is dependent on CaN/NFATc1 signaling. The present study shed new light on the role of Akirin2 in pigs.
MATERIALS AND METHODS
Reagents
The calcineurin inhibitor cyclosporin A (CsA) was obtained from Amresco (Solon, OH, USA) and resolved in DMSO, then used at 5 μM. NFATc1 antibody was obtained from Cell Signaling Technology (Danvers, MA, USA). GAPDH antibody was obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). MyHC I antibody (BA-F8) was purchased from the Developmental Studies Hybridoma Bank (DSHB, Iowa City, IA, USA). Calcineurin Cellular Activity Assay Kit was purchased from Solarbio (Beijing, China).
Animals and tissue sample collection
Three 10-week-old female Duroc × Landrace × Yorkshire (DLY) pigs were slaughtered at body weight of 31-31.6 kg in a humane manner according to protocols approved by the Animal Care Advisory Committee of Sichuan Agricultural University. The psoas major (PM) muscle and the tibialis anterior (TA) muscle were removed and immediately snap frozen in liquid nitrogen before being stored at −80 °C for protein extraction.
Isolation of porcine skeletal muscle satellite cells and cell culture
Porcine skeletal muscle satellite cells were isolated from longissimus lumborum muscle of 3-day-old male DLY pigs. The pigs obtained from Sichuan Zhengyuan swine industry Co., Ltd. (Chengdu, Sichuan, China) were slaughtered according to protocols approved by the Animal Care Advisory Committee of Sichuan Agricultural University. Briefly, muscle was digested with 0.2% collagenase type II (Sigma, St. Louis, MO, USA) and then filtered successively through 200-mesh and 400-mesh cell sieves. The collected cells were purified by differential adhesion method. The resulting cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM)/F12 (Invitrogen, Carlsbad, CA, USA) comtaining 20% fetal bovine serum (FBS), 100 U/mL penicillin and 100 μg/μL streptomycin at 37 ºC in a humidified 5% CO2 atmosphere. The cells were identified by immunofluorescence staining with Pax7 antibody (DSHB).
The siRNA and plasmid transfection
In this study, the siRNA sequence targeting NFATc1 was designed and synthesized by GenePharm (Shanghai, China). The sense strand of the NFATc1 siRNA was 5’-GGACCAGGAGUUCGACUUUTT-3’, and the antisense strand was 5’-AAAGUCGAACUC CUGGUCCTT-3’; The sense strand of the negative control siRNA was 5’-UUCUCCGAACGUGUCACGUTT-3’, and the antisense strand was 5’-ACGUGACACGUU CGGAGAATT-3’. Porcine skeletal muscle satellite cells were transfected with 50 nM siRNA for NFATc1, 50 nM negative control siRNA, 2 μg pcDNA3.1(+)-Akirin2 plasmid (Chen at al., 2012) or 2 μg empty pcDNA3.1(+) vector by using Lipofectamine 3000 (Invitrogen) according to the manufacturer’s instructions.
Cell treatment
To determine whether the Akirin2 affects the expression of MyHC I via the calcineurin/NFATc1 signaling pathway, we treated the porcine skeletal muscle satellite cells with 5 μM CsA or the siRNA for NFATc1, and analyzed the protein expressions of NFATc1 and MyHC I by Western blot.
RNA extraction and real-time quantitative PCR
Total RNA was extracted from the cells using the RNAiso Plus reagent (TaKaRa, Dalian, China) according to the manufacture’s instructions. One microgram of total RNA was transcribed into a single strand cDNA by using PrimeScript® RT reagent Kit with gDNA Eraser (TaKaRa). For real-time quantitative PCR, the SYBR select Master Mix (Applied Biosystems, Foster, CA, USA) was used in an ABI 7900HT Real-time PCR system. The gene specific primers used are as follows: porcine MCIP1.4 (forward) 5’-GGGCCAAATTTGAATCCC-3’ and (reverse) 5’-GGGTTGCTGAAGTTGATTCTG-3’; porcine GAPDH (forward) 5’-ACACTGAGGACCAGGTTGTG-3’ and (reverse) 5’-GACGAAGTGGTCGTTGAGGG-3’. The PCR cycling conditions used were: 45 cycles at 95 °C for 15 s and 60 °C for 30 s. Relative gene expression was calculated by the comparative Ct method [29]. GAPDH served as an internal standard.
Western blot
Western blot analysis was performed as previously described by Chen et al. [30]. Briefly, equal amounts (30 μg) of protein of each sample were subjected to 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Following transfer of the proteins to a nitrocellulose membrane, the membrane was incubated in 3% non-fat milk in TBS-0.1% Tween-20 and then incubated overnight with primary antibody at 4 ºC, followed by horseradish peroxidase-linked secondary antibodies (Santa Cruz Biotechnology). The proteins were visualized by ClarityTM Western ECL Substrate (Bio-Rad, Hercules, CA, USA). GAPDH is considered as a housekeeping protein. The density of protein bands was measured using Gel-Pro Analyzer 4.2 software (Media Cybernetics, Rockville, MD, USA).
Calcineurin activity analysis
Cells were harvested and rinsed three times using PBS. Cells were lysed in ice-cold lysis buffer (1% Triton X-100) for 30 min, and the lysates were used immediately for calcineurin (CaN) activity assay. CaN activity was measured with the Calcineurin Cellular Activity Assay Kit according to the manufacturer’s instructions. Specific CaN activity was calculated by normalization to the total protein content. Data are expressed as U per mg protein.
Statistical analysis
All data are presented as mean ± SE. The data were subjected to one-way ANOVA analysis and Tukey’s tests using SPSS 11.0 software. Differences were accepted as significant at P < 0.05.
ACKNOWLEDGMENTS
We would like to thank the staff at our laboratory for the assistance they have provided in this study.
CONFLICTS OF INTEREST
None declared.
FUNDING
This work was supported by the National Natural Science Foundation of China (No. 31472108) and the Specific Research Supporting Program for Discipline Construction in Sichuan Agricultural University.
REFERENCES
1. Klont RE, Brocks L, Eikelenboom G. Muscle fibre type and meat quality. Meat Science. 1998; 49: S219-S229.
2. Lefaucheur L, Lebret B, Ecolan P, Louveau I, Damon M, Prunier A, Billon Y, Sellier P, Gilbert H. Muscle characteristics and meat quality traits are affected by divergent selection on residual feed intake in pigs. Journal of Animal Science. 2011; 89: 996-1010.
3. Raj S, Skiba G, Weremko D, Fandrejewski H, Migdal W, Borowiec F, Polawska E. The relationship between the chemical composition of the carcass and the fatty acid composition of intramuscular fat and backfat of several pig breeds slaughtered at different weights. Meat Science. 2010; 86: 324-330.
4. Goto A, Matsushita K, Gesellchen V, Chamy LE, Kuttenkeuler D, Takeuchi O, Hoffmann JA, Akira S, Boutros M, Reichhart J. Akirins are highly conserved nuclear proteins required for NF-κB-dependent gene expression in drosophila and mice. Nature Immunology. 2008; 9: 97-104.
5. Akiyama H, Iwahana Y, Suda M, Yoshimura A, Kogai H, Nagashima A, Ohtsuka H, Komiya Y, Tashiro F. The FBI1/Akirin2 target gene, BCAM, acts as a suppressive oncogene. PLoS One. 2013; 8: e78716.
6. Qu FF, Xiang ZM, Zhang Y, Li J, Zhang YH, Yu ZN. The identification of the first molluscan Akirin2 with immune defense function in the Hong Kong oyster Crassostrea hongkongensis. Fish & Shellfish Immunology. 2014; 41: 455-465.
7. Tartey S, Matsushita K, Vandenbon A, Ori D, Imamura T, Mino T, Standley DM, Hoffmann JA, Reichhart J, Akira S, Takeuchi O. Akirin2 is critical for inducing inflammatory genes by bridging IκB-ζ and the SWI/SNF complex. EMBO Journal. 2014; 16: 2332-2348.
8. Sasaki Y, Nagai K, Nagata Y, Doronbekov K, Nishimura S, Yoshioka S, Fujita T, Shiga K, Miyake T, Taniguchi Y, Yamada T. Exploration of genes showing intramuscular fat deposition-associated changes in musculus longissimus muscle. Animal Genetics. 2006; 37: 40-46.
9. Sasaki S, Yamada T, Sukegawa S, Miyake T, Fujita T, Morita M, Ohta T, Takahagi Y, Murakami H, Morimatsu F, Sasaki Y. Association of a single nucleotide polymorphism in Akirin2 gene with marbling in Japanese Black beef cattle. BMC Research Notes. 2009; 2: 131-135.
10. Watanabe N, Satoh Y, Fujita T, Ohta T, Kose H, Muramatsu Y, Yamamoto T, Yamada T. Distribution of allele frequencies at TTN g.231054C>T, RPL27A g.3109537C>T and Akirin2 c.*188G>A between Japanese black and four other cattle breeds with differing historical selection for marbling. BMC Research Notes. 2011; 4: 10.
11. Kim H, Lee SK, Hong MW, Park SR, Lee YS, Kim JW, Lee HK, Jeong DK, Song YH, Lee SJ. Association of a single nucleotide polymorphism in the Akirin2 gene with economically important traits in Korean native cattle. Animal Genetics. 2013; 44: 750-753.
12. Sukegawa S, Miyake T, Ibi T, Takahagi Y, Murakami H, Morimatsu F, Yamada T. Multiple marker effects of single nucleotide polymorphisms in three genes, AKIRIN2, EDG1 and RPL27A, for marbling development in Japanese Black cattle. Animal Science Journal. 2014; 85: 193-197.
13. Chen XL, Huang ZQ, Jia G, Wu XQ, Wu CM. Molecular cloning, tissue distribution, and functional analysis of porcine Akirin2. Animal Biotechnology. 2012; 23: 124-131.
14. Chen XL, Huang ZQ, Zhou B, Wang H, Jia G, Qiao JY. Expression and purification of porcine Akirin2 in Escherichia coli. Turkish Journal of Biology. 2014; 38: 339-345.
15. Chen XL, Luo YL, Zhou B, Huang ZQ, Jia G, Liu GM, Zhao H, Yang ZP, Zhang RN. Effect of porcine Akirin2 on skeletal myosin heavy chain isoform expression. International Journal of Molecular Sciences. 2015; 16: 3996-4006.
16. Joo ST, Kim GD, Hwang YH, Ryu YC. Control of fresh meat quality through manipulation of muscle fiber characteristics. Meat Science. 2013; 95: 828-836.
17. Shen LY, Lei HG, Zhang SH, Li XW, Li MZ, Jiang XB, Zhu KP, Zhu L. The comparison of energy metabolism and meat quality among three pig breeds. Animal Science Journal. 2014; 85: 770-779.
18. Liu Y, Li M, Ma J, Zhang J, Zhou C, Wang T, Gao X, Li X. Identification of differences in microRNA transcriptomes between porcine oxidative and glycolytic skeletal muscles. BMC Molecular Biology. 2013; 14: 7.
19. Chin ER, Olson EN, Richardson JA, Yang Q, Humphries C, Shelton JM, Wu H, Zhu W, Bassel-Duby R, Williams RS. A calcineurin-dependent transcriptional pathway controls skeletal muscle fiber type. Genes & Development. 1998; 12: 2499-2509.
20. Swoap SJ, Hunter RB, Stevenson EJ, Felton HM, Kansagra NV, Lang JM, Esser KA, Kandarian SC. The calcineurin-NFAT pathway and muscle fiber-type gene expression. American Journal of Physiology-Cell Physiology. 2000; 279: C915-C924.
21. Kubis HP, Hanke N, Scheibe RJ, Meissner JD, Gros G. Ca2+ transients activate calcineurin/NFATc1 and intiate fast-to-slow transformation in a primary skeletal muscle culture. American Journal of Physiology-Cell Physiology. 2003; 285: C56-C63.
22. Schiaffino S, Reggiani C. Fiber types in mammalian skeletal muscles. Physiological Reviews. 2011; 91: 1447-1531.
23. Calabria E, Ciciliot S, Moretti I, Garcia M, Picard A, Dyar KA, Pallafacchina G, Tothova J, Schiaffino S, Murgia M. NFAT isoforms control activity-dependent muscle fiber type specification. Proceedings of the National Academy of Sciences of the United States of America. 2009; 106: 13335-13340.
24. Macian F. NFAT proteins: key regulators of T-cell development and function. Nature Reviews Immunology. 2005; 5: 472-484.
25. Ehlers ML, Celona B, Black BL. NFATc1 controls skeletal muscle fiber type and is a negative regulator of MyoD activity. Cell Reports. 2014; 8: 1639-1648.
26. Yang J, Rothermel B, Vega RB, Frey N, McKinsey TA, Olson EN, Bassel-Duby R, Williams RS. Independent signals control expression of the calcineurin inhibitory proteins MCIP1 and MCIP2 in striated muscles. Circulation Research. 2000; 87: E61-E68.
27. Rothermel BA, Vega RB, Williams RS. The role of modulatory calcineurin-interacting proteins in calcineurin signaling. Trends in Cardiovascular Medicine. 2003; 13: 15-21.
28. Fenyvesi R, Rábor G, Wuytack F, Zádor E. The calcineurin activity and MCIP1.4 mRNA levels are increased by innervation in regenerating soleus muscle. Biochemical and Biophysical Research Communications. 2004; 320: 599-605.
29. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔCt method. Methods. 2001; 25: 402–408.
30. Chen XL, Zhou B, Luo YL, Huang ZQ, Jia G, Liu GM, Zhao H. Tissue distribution of porcine FTO and its effect on porcine intramuscular preadipocytes proliferation and differentiation. PLoS One. 2016; 11: e0151056.