
INTRODUCTION
Aflatoxins are probably the most intensively researched mycotoxins in the world as 4.5 billion people worldwide suffer from intemperate exposure to aflatoxins which causes 4.6–28.2% of all global hepatocellular carcinoma cases [1, 2]. These fungal metabolites are mainly produced by Aspergillus flavus and Aspergillus parasiticus. The most common aflatoxins are B1, B2, G1 and G2 which are naturally present in many food products and affect more than one organ system simultaneously, therefore producing cascade of responses in the affected organism [3, 4]. Among the major aflatoxins, aflatoxin B1 (AFB1) is the most widely known carcinogen [5] having highly hepatotoxic, genotoxic, immunotoxic, and other adverse health effects on humans and several other animal species [6]. International Agency for Research on Cancer (IARC) have produced sufficient evidences of carcinogenicity of AFB1 and classified it as a Group 1 human carcinogen and intended no safe dose [7].
As the major drug metabolizing and detoxifying organ in the body, liver is primarily affected followed by ingestion of the aflatoxins [8, 9]. AFB1 is potent hepatocarcinogen in humans and exposure to AFB1 is known to cause both chronic and acute hepatocellular injury. AFB1 action mechanism involves DNA adducts formation and mutation at codon 249 of the p53 tumor suppressor gene leading to carcinogenicity in humans [10–12]. Although AFB1 are hepatotoxic, causing pallor discoloration, enlargement, congestion and necrosis of liver along with triggering proliferation of bile duct and infiltration of mononuclear and heterophilic cells in many livestock and laboratory animals, yet susceptibility varies with breed, species, age, dose, length of exposure and nutritional status [13–15].
Apoptosis is a programmed form of cell death [16, 17] and many studies have demonstrated its role in implicating pathogenesis of multiple diseases in humans and animals [18–20]. It is also well documented in different model systems that aflatoxins react antagonistically with different cell proteins, leading to inhibition of carbohydrate and lipid metabolism and protein synthesis, which can induce apoptosis [21]. Liao et al., 2014, investigated that AFB1 can provoke liver dysfunctioning by promoting hepatocytes apoptosis and disturbing cellular enzymatic activities. AFB1 induced apoptosis can be initiated by two main apoptotic pathways: the extrinsic and the intrinsic. Although intrinsic pathway is also involved in different tissue apoptosis [22, 23], yet in liver cells due to high level of death receptor expression, apoptosis predominantly occurs via the extrinsic pathway [24] and contributes to the development of a number of liver diseases including liver cirrhosis, cholestasis, viral hepatitis and hepatocellular carcinoma [25, 26]. Our previous studies have revealed that AFB1 negatively affect the spleen, thymus, jejunum and ilium and elucidated the mechanisms by which AFB1 induces apoptosis in these organs and tissues in chicken [27–31]. Looking at the earlier reports in different organ systems, it would be enthralling to find out how liver is affected at cellular and molecular level, by AFB1. In order to understand this, the present study is designed to explore the apoptotic mechanisms primed by death receptor molecules in AFB1 induced liver apoptosis by analyzing histological, ultra structural, biochemical, flow cytometrical and relative expression changes in apoptosis-regulating genes in hepatocytes using chicken as a research model, considering the fact that chicks are susceptible to AFB1 exposure [32]. As cell proliferation and apoptosis are directly related to carcinogenicity [33], the outcomes from present study could facilitate the understanding of AFB1 induced carcinogenicity and may be helpful in bringing down the toxigenic potential of AFB1.
RESULTS
Histological and ultrastructural analysis
Microscopically, degenerative reversible lesions were seen, from mildest to severest degree with various distributions in AFB1 group. Compared to control group (Figure 1A), slight to moderate hydropic degeneration, fatty vacuolar degeneration and proliferation of bile duct in hepatocytes were seen in AFB1 group. Moreover pyknotic and fragmented nuclei were also visible in AFB1 group (Figure 1B). Results of ultrastructural observation revealed irregular, fragmented and condensed nucleus, swollen mitochondria with reduced number of cristae and swollen endoplasm reticulum in AFB1 group ( Figure 1D-1F) when compared to control (Figure 1C). AFB1 induced histopathological lesions were observed in chicken liver. The incidence of liver congestion, vacuolar or fatty degeneration, proliferation of bile duct and nuclear fragmentation were seen going up from day 7 to day 21 in AFB1 intoxicated chickens whereas no lesions were observed in the control group (Table 1).
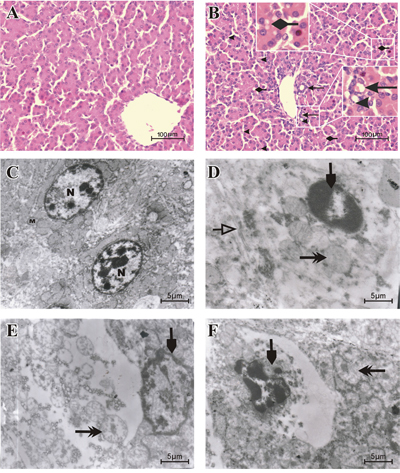
Figure 1: Histopathological and Ultrastructural changes displayed by liver cells when exposed to AFB1. Histological examination of H & E stained liver tissues from 21 days old chickens fed on control diet A. and AFB1 B. (Bar = 100 μm). Proliferation of bile duct . Fragmented nuclei
. Fragmented nuclei . Hydropic and vacuolar fatty degeneration
. Hydropic and vacuolar fatty degeneration . Ultrastructural examination of uranyl acetate and lead citrate stained liver tissue from the control group C. with normal nucleus (N), normal mitochondria (M) and other normal organelles and the AFB1 group D, E and F (Bar = 5 μm). Condensed, irregular or fragmented nucleus (
. Ultrastructural examination of uranyl acetate and lead citrate stained liver tissue from the control group C. with normal nucleus (N), normal mitochondria (M) and other normal organelles and the AFB1 group D, E and F (Bar = 5 μm). Condensed, irregular or fragmented nucleus ( , D, E and F), swollen mitochondria (
, D, E and F), swollen mitochondria ( , D, E and F) with decreased cristae (
, D, E and F) with decreased cristae ( , E) and swollen endoplasmic reticulum (, D).
, E) and swollen endoplasmic reticulum (, D).
Table 1: Incidence of major histopathological lesions of chicken liver
Pathological lesions |
Time |
Control group |
AFB1 group |
|---|---|---|---|
7 days |
0/6 |
1/6 |
|
Liver congestion |
14 days |
0/6 |
2/6 |
21 days |
0/6 |
4/6 |
|
7 days |
0/6 |
1/6 |
|
Vacuolar or fatty degeneration |
14 days |
0/6 |
3/6 |
21 days |
0/6 |
5/6 |
|
7 days |
0/6 |
2/6 |
|
Proliferation of bile duct |
14 days |
0/6 |
4/6 |
21 days |
0/6 |
6/6 |
|
7 days |
0/6 |
1/6 |
|
Increased nuclear fragmentation |
14 days |
0/6 |
2/6 |
21 days |
0/6 |
5/6 |
Incidence of histopathological lesions in the liver among chickens from different experimental groups (n=6)
Apoptotic percentage by flow cytometer
FITC Annexin V was used to quantitatively determine the percentage of cells within groups that were actively undergoing apoptosis. Apoptotic cell counts were determined by examining the total percentage of early (Annexin-V positive and PI negative) and late (both Annexin-V and PI positive) apoptotic cells. Figure 2A, 2C, 2E and 2B, 2D, 2F shows scattered analysis of early and late hepatocytes apoptosis at 7, 14 and 21 days in the control and AFB1 groups respectively. When compared to the control, the AFB1 group at 7, 14 and 21 days displayed a significantly increased percentage of apoptotic cells (p<0.05 or p<0.01) (Figure 2G).
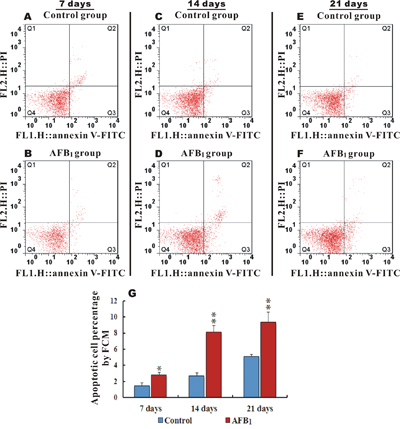
Figure 2: Apoptotic hepatocytes percentage and scattered analysis of early and late AFB1 induced apoptosis in chicken hepatocytes. Figure A, C, E and B, D, F. represents scattered analysis of early (Annexin-V positive and PI negative) and late (Annexin-V and PI positive) hepatocytes apoptosis in the control and AFB1 groups at 7, 14 and 21 days respectively, while G. is a bar graph showing apoptotic percentage rate in the control and AFB1 group at day 7, 14 and 21 by flow cytometer (number of chickens, n=6). *p<0.05, or **p<0.01, when compared to the control.
Biochemical analysis
Compared to the control group, the activities of CAT (Catalase) and GSH-Px (Glutathione Peroxidase) in the AFB1 group were observed significantly downregulated (p<0.05 or p<0.01) throughout the experiment at different time points. Activity of SOD (Superoxide Dismutase) and Hydroxyl free radical scavenging were also seen remarkably down (p<0.05 or p<0.01) in the AFB1 group compared to the control at 14 and 21 days. The contents of MDA (Malondialdehyde) were observed significantly upregulated at 7, 14 and 21 days (p<0.01), while GSH (glutathione) contents were seen reduced (p<0.05 or p<0.01) at 14 and 21 days in AFB1 group when compared to the control group (Figure 3).
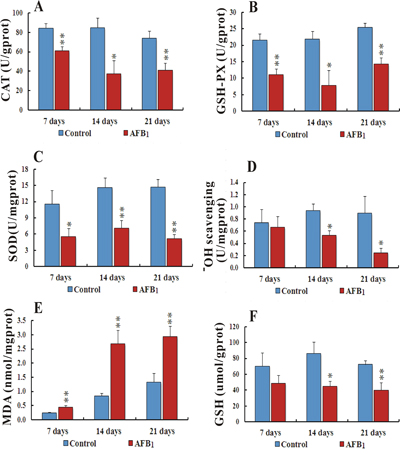
Figure 3: Biochemical analyses of activities of CAT, GSH-Px, SOD, Hydroxyl ion scavenging and contents of MDA and GSH in liver cells intoxicated with AFB1. Figure 3A-3C, represents activities of CAT, GSH-Px and SOD respectively. Bar graph D. shows Hydroxyl free radical scavenging while E and F. bar graphs display contents of MDA and GSH in chicken hepatocytes (number of chickens, n = 6). *p<0.05, or **p<0.01, when compared to the control.
qRT-PCR analysis of relative expression of genes involved in death receptors induced apoptosis
The death receptor genes FAS (Fatty acid synthetase receptor) and TNF R1 (Tumor necrosis factor receptor 1), and the genes involved in DISC (death-inducing signaling complex) formation i-e FADD (FAS-associated death domain), TRADD (TNF receptor-associated death domain), and TRAF2 (TNF receptor-associated factor 2) were seen significantly upregulated (p<0.05 or p<0.01) at 7, 14, & 21 days with an exception of TRAF2 which was not significantly upregulated at day 7 in AFB1 group when compared with the control group. The mRNA levels of CASPASE (cysteine-aspartic protease) family genes, i-e. CASPASE 3, CASPASE 8, CASPASE 9, and CASPASE 10 were seen significantly raised at 14 and 21 days (p<0.05 or p<0.01), also CASPASE 3 was seen significantly upregulated at day 7 in AFB1 group compared to the control. BCL-2 (B-cell lymphoma 2) and XIAP (X-linked inhibitor of apoptosis protein) genes displayed notable downregulation (p<0.05 or p<0.01) at day 14 and 21, while mRNA levels of IKK (IκB kinase) did not change throughout 7, 14 and 21 days, although it was seen increasing at day 21 but not significant enough (p>0.05) (Figure 4).
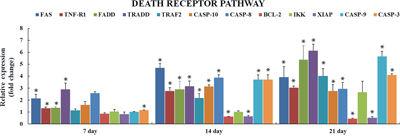
Figure 4: Relative expression of genes involved in death receptor induced apoptosis in liver of chickens exposed to AFB1. Figure 4 represents the mRNA levels of FAS, TNF-α, FADD, TRADD, TRAF2, CASPASE 10, CASPASE 8, BCL-2, IKK, XIAP, CASPASE 9 and CASPASE 3 in liver of the AFB1-fed chickens and expressed as fold change relative to the control group (number of chickens, n = 6). *p<0.05, or **p<0.01, when compared to the control.
DISCUSSION
In the current study we scrutinized the mechanisms of AFB1 induced hepatocyte apoptosis by judging the involvement of apoptosis- associated genes and oxidant situation. The histopathological changes in AFB1 intoxicated livers were in general agreement with previous reports [34, 35]. Slight to moderate hydropic degeneration, fatty vacuolar degeneration, proliferation of bile duct, massive congestion in midzonal areas, pyknotic and fragmented nuclei within hepatocytes were seen in AFB1 intoxicated chickens. Furthermore various electron microscopically detectable changes characterized apoptosis, such as condensed and hyperchromatic chromatin, fragmented nuclei, swollen mitochondria and swollen endoplasmic reticulum, which were in concordance with previously published work by several research groups [16, 19, 36]. The flow cytometric analysis revealed increased hepatocyte apoptotic percentage in AFB1 group, which was similar to our previous findings in thymus, spleen, intestine and bursa of fabricius [23, 30, 31, 37] and this increased hepatocyte apoptosis can possibly be associated with AFB1 induced carcinogenicity since the rates of cell proliferation and cell death are directly related to cancer [32]. To understand the involvement of apoptosis associated genes in liver cells, we further explored the expression of death receptor signaling pathway involved in hepatocyte apoptosis.
Induction of apoptosis can be triggered by the activation of death receptors including FAS, TNF-α, followed by DISC (death-inducing signaling complex) formation, which consists of CASPASE 8, and associated proteins such as FADD, TRADD and TRAF2 [17]. The present study revealed over expression of FAS and TNF-R1 receptors along with their associated genes i-e CASPASE 8, CASPASE 10, FADD, TRADD and TRAF2, which was consistent with earlier reports in human hepatocytes [38, 39] and with our recent study on thymocytes apoptosis [23]. IKK is crucial in activation of NF-kB based intracellular survival signaling and can activate the expression of several anti-apoptotic proteins [40, 41]. XIAP is apoptotic inhibitor that can plug up the process of cell death induced by any foreign stimuli [42]. In our present study mRNA levels of IKK were not seen significantly increased throughout the experiment at different time points and XIAP expression was significantly decreased at 14 and 21 days and the expression of FADD was much higher than TRAF2. Taken all together present study insinuated that TNF1-TRADD mediated signaling is originating from TRADD, supported by ability of FADD to activate apoptosis rather than activating NF-kB cell survival signaling.
The CASPASE-cascade system plays vital roles in the induction, transduction and amplification of intracellular apoptotic signals [43] and overexpression of CASPASE family genes can force cells to undergo apoptosis. In present study, AFB1 administration led to overexpression of CASPASE family genes (CASPASE 10, 8, and 3), suggesting that apoptosis induced by FAS and TNFR1 activates CASPASE 8, which contains an N-terminus with FADD, so providing a direct link between cell death receptors and the Caspases. In addition overexpression of CASPASE 9 and down-regulation of BCL-2 genes indicated that mitochondrial pathway was also involved in hepatocyte apoptosis induced by AFB1 (Figure 5). These results were in agreement with previous studies in human and rat hepatocytes [44–47] and our recent report in thymus apoptosis induced by AFB1 [23].
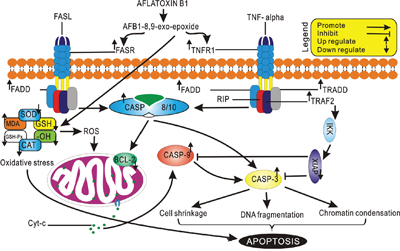
Figure 5: Schematic representation of Aflatoxin B1 induced apoptosis through Death Receptor Signaling.
Recently many studies have focused on revealing the pivotal role of ROS as well as resulting oxidative stress in inducing apoptosis [48, 49]. It’s also previously been demonstrated that disequilibrium between ROS formation and antioxidant defense system may cause DNA damage and mitochondrial disintegration by producing oxidative stress which could lead to apoptosis [50–52]. Oxidative damage is relieved and free radicals are removed from the system by crucial catalyst, such as SOD, CAT and GSH-Px. O2 is converted in to H2O2 by SOD, which is further converted in to H2O by GSH-Px [53]. MDA is an important index of antioxidant ability, since it is the main product of lipid peroxidation [54]. In present study, activities of CAT, GSH-Px and SOD were observed significantly decreased in AFB1 treated group along with hydroxyl ion free radical scavenging. A significantly increased contents of MDA and reduced contents of GSH were seen in AFB1 treated group. These results were constant with previous studies in multiple organs in different species [23, 30, 50, 55–57]. These results clearly demonstrated that AFB1 could cause imbalance between ROS formation and antioxidant defense by accumulating ROS and producing oxidative stress that could play an important role in mediating hepatocytes apoptosis.
MATERIALS AND METHODS
All the chickens and AFB1 experiments have been conducted in accordance with the ethical standards, declaration of Helsinki and national and international guidelines have been approved by Sichuan Agricultural University Animal Care and Use Committee (Approval No: 2012-024).
Chickens and diets
One hundred and fifty-six one-day-old healthy Cobb-500 broilers were purchased from Chia Tai Group (Wenjiang, Sichuan, China), and were randomly divided into two groups which are the control group (0 mgAFB1/kg of basal diet) and AFB1 group (0.6 mg AFB1/kg of basal diet) with 26 chickens in each group and each group in triplicates. The basal diet, which was control diet, was formulated according to National Research Council (NRC, 1994) [58] and Chinese Feeding Standard of Chicken (NY/T33-2004) recommendations. The AFB1-contaminated diet was prepared, using the method described by Kaoud [59]. Briefly, 27 mg AFB1 (A6636, Sigma-Aldrich, USA) was dissolved into 30 ml methanol, and the solution was mixed with 45 kg corn-soybean basal diet to formulate the AFB1-contaminated diet. The control diet was prepared using the same constituents except AFB1. Then the methanol of diets was evaporated at 98 °F (37 °C). The AFB1 concentrations were analyzed by HPLC (Waters, Milford, MA, USA) with fluorescence detection (Waters, Model 2475, Milford, MA, USA), and the AFB1 concentration was determined as <0.001 mg/kg and 0.601 mg/kg respectively in the control diet and AFB1 diet. chickens were housed in cages with electrically heated units and provided with water as well as aforementioned diet ad libitum for 21 days.
Histopathological and ultrastructural examination
At the age of 7, 14 and 21 days, six chickens in each group were euthanized and the livers were fixed in 4% paraformaldehyde (PFA) and routinely processed in paraffin. Thin sections (5 μm) of tissue were sliced, mounted on glass slides and stained with hematoxylin and eosin Y. The histological organization of the tissues were contemplated and snapped with a digital camera (Nikon, eclipse 50i, Japan).
In each group one chick per replicate, was euthanized and then immediately necropsied at the end of the trial. Small pieces of liver tissues were immediately fixed with 2.5% glutaraldehyde and post-fixed in 2% Veronal acetate-buffered OsO4. The tissues were embedded in Araldite after dehydrating in alcohol gradient. The blocks were sectioned in 65-75 nm thick sections in a microtome with a glass knife and placed in uncoated copper grids. The sections were stained with uranyl acetate, and post-stained with 0.2% lead citrate. The subcellular architecture of liver was examined with a Hitachi H-600 transmission electron microscope (Japan).
Annexin v apoptosis detection by flow cytometry
At 7, 14, and 21 days of the experiment, six chickens in each group were euthanized, and livers were sampled from each chick to determine the percentage of apoptotic cells by flow cytometer, using the method by Chen et al. 2011 [60]. Briefly, the dissected livers were thereupon homogenized to form a cell suspension and filtered, and then the cells were washed and resuspended in phosphate buffer at a concentration of 1x 106 cells/mL. 5 μL Annexin V-Fluorescein isothiocyanate (V-FITC) and 5 μL propidium iodide (PI) were added into 100 μL cell suspension, and incubated at 25 ºC for 15 min in the dark. 400 μL 1 x Annexin binding buffer was added to the mixture, and then the apoptotic cells were assayed by flow cytometer (BD FACSCalibur) within 1 h. The annexin V-FITC Kit was obtained from BD Pharmingen (USA, 556547).
Quantitative real-time PCR (qRT-PCR)
Livers from six chickens in each group were removed at 7, 14, and 21 days of age, and instantly stored in liquid nitrogen. The livers samples were homogenized in liquid nitrogen, by crushing with a mortar and pestle and the powdered tissues were collected into eppendorf tubes and stored at -80°C. Total RNA was extracted using TriPure Isolation Reagent kit (Cat No. 11667165001, Roche Applied Science, Germany) following manufacturer’s protocol. The quality and quantity of total RNA was measured spectrophotometrically. Extracted RNA was forthwith reverse-transcribed into cDNA using Transcriptor First Strand cDNA Synthesis Kit (Cat No: 04897030001, Roche Applied Science, Germany). qRT-PCR reactions were performed in a total volume of 20 μL using FAStStart Universal SYBR Green Master mix (Cat No: 04913914001, Roche Applied Science, Germany), at the following thermocycler program; Initial denaturation at 95 °C for 10 min, followed by 44 cycles of “10 s at 95 °C and 30 s at melting temperature (Tm) of a specific primer pair”, and melt curve analysis by 10 s at 95 °C, and 72 °C for 10 s, using Thermal Cycler (Step One Plus, Applied BioSystems, USA). β-actin was used as an internal control [61, 62]. Primers information is provided in Table 2. The qRT-PCR data were analyzed and fold change in expressions were calculated using 2-ΔΔCt calculation method described by [63].
Table 2: List of oligonucleotides used as primers in qRT-PCR analysis
Gene symbol |
RefSeq mRNA number |
Forward primer |
Reverse primer |
Amplicon length (bp) |
|---|---|---|---|---|
FAS |
NM_001199487 |
TCCACCTGCTCCTCGTCATT |
GTGCAGTGTGTGTGGGAACT |
78 |
TNF-R1 |
NM_001030779 |
CCTGTCTGTCTTCCCTGTCC |
GGTGCATGGGGTCTTTTCTA |
120 |
TRADD |
XM_414067 |
CTAGAGCCCAAAGGAAGTCGAT |
TGGCTGCTTCTCTGTGACAT |
100 |
FADD |
XM_421073 |
GGGGTAAAGAGGCTGAACTCTTA |
TGAGTCCTATTGCACTGCTGTC |
163 |
TRAF2 |
XM_015279623 |
CGTGGTGATGAAAGGACCCA |
AATGATGTGCTCCCGGTTGT |
100 |
Casp-10 |
XM_421936 |
CTGGGGGCTCCAAAAGTCC |
AAAGGGGGACAAAGCCAACA |
204 |
Casp-9 |
AY057940 |
CCAACCTGAGAGTGAGCGATT |
GTACACCAGTCTGTGGGTCGG |
87 |
Casp-8 |
NM_204592 |
GTCTCCGTTCAGGTATCTGCT |
TCTCAATGAAAACGTCCGGC |
143 |
Casp-3 |
NM_204725 |
TGGCCCTCTTGAACTGAAAG |
TCCACTGTCTGCTTCAATACC |
139 |
IKK (IKBIP) |
XM_001232182 |
GGCTTGGTTTTGGCAGTGAG |
CGGCTTTGACGTTTGCTGAA |
144 |
XIAP |
NM_204588 |
GCAGAATATGAGAGGCGGATAC |
TCCTTCCACTCTTGCAATCC |
149 |
BCL-2 |
NM_205339 |
TGTTTCTCAAACCAGACACCAA |
CAGTAGGCACCTGTGAGATCG |
205 |
β-actin |
L08165 |
TGCTGTGTTCCCATCTATCG |
TTGGTGACAATACCGTGTTCA |
178 |
Biochemical analysis
Six chickens in each group were euthanized and immediately necropsied at 7, 14 and 21 days of age. Then livers were immediately taken out and chilled to 0°C in 0.85% NaCl, and then dried, weighed and homogenized in 9 vol of icecold 0.85% NaCl in a chilled homogenizer and centrifuged at 3500 × g at 4°C for 10 min. Total protein was measured by the method of Bradford [33] The commercial kits were purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China) and used to detect total protein (Total protein quantification kit No.A045), activities of SOD (Superoxide dismutase detection kit No.A001-1), CAT (Catalase detection kit No.A007), GSH-Px (Glutathione peroxidase detection kit No.A005) and contents of GSH (Glutathione detection kit No.A006) and MDA (Malonaldehyde detection kit No.A003-1), and −OH radical (Hydroxyl free radical detection kit No.A018) in the supernatant, according to the manufacturer’s instructions.
Statistical analysis
The significance of difference between two groups was analyzed by variance analysis, and results are expressed as the mean value with deviation. The analysis was performed using the independent sample t test of SPSS software for Mac v.20.0 (IBM Corp, Armonk, NY, USA) and a value of p<0.05 was considered significant, while p<0.01 was considered markedly significant.
CONCLUSION
Tissue specific apoptosis caused by AFB1 induced toxicity has been studied in much detail, yet the mechanisms of death receptor induced apoptosis in hepatocytes remain least explored. In present study, we have delineated the mechanisms involved in death receptor provoked apoptosis using chicken liver as a model system. The results obtained in this study could be further used to understand involvement of death receptors in apoptosis and could help in amending the toxic effect provoked by AFB1.
Abbreviations
AFB1, aflatoxin B1; CAT, Catalase; GSH-Px, Glutathione Peroxidase; SOD, Superoxide Dismutase; MDA, Malondialdehyde; GSH, glutathione; FAS, Fatty acid synthetase receptor; TNF R1, Tumor necrosis factor receptor 1; DISC, death-inducing signaling complex; FADD, FAS-associated death domain; TRAF2, TNF receptor-associated factor 2; CASPASE, cysteine-aspartic protease; BCL-2, B-cell lymphoma 2; XIAP, X-linked inhibitor of apoptosis protein); IKK, IκB kinase.
ACKNOWLEDGMENTS
This work was supported by the program for Changjiang scholars, the University Innovative Research Team (IRT 0848) and the Education Department of Sichuan Province (2012FZ0066) and (2013FZ0072).
CONFLICTS OF INTEREST
The authors declare no competing financial interests.
REFERENCES
1. Liu Y, Wu F. Global burden of aflatoxin-induced hepatocellular carcinoma: a risk assessment. Environmental health perspectives. 2010; 118:818. doi: 10.1289/ehp.0901388.
2. Abrar M, Anjum FM, Butt MS, Pasha I, Randhawa MA, Saeed F, Waqas K. Aflatoxins: biosynthesis, occurrence, toxicity, and remedies. Critical reviews in food science and nutrition. 2013; 53:862-74. doi: 10.1080/10408398.2011.563154.
3. Ferenčík M, Ebringer L. Modulatory effects of selenium and zinc on the immune system. Folia microbiologica. 2003; 48:417-26. doi: 10.1007/BF02931378.
4. Herzallah SM. Aflatoxin B1 residues in eggs and flesh of laying hens fed aflatoxin B1 contaminated diet. American Journal of Agricultural & Biological Science. 2013; 8:156-61. doi: 10.3844/ajabssp.2013.156.161.
5. Stettler P, Sengstag C. Liver carcinogen aflatoxin B1 as an inducer of mitotic recombination in a human cell line. Molecular Carcinogenesis, 2001, 31:125-138. doi: 10.1002/mc.1047.
6. Kalpana S, Aggarwal M, Rao GS, Malik JK. Effects of aflatoxin B1 on tissue residues of enrofloxacin and its metabolite ciprofloxacin in broiler chickens. Environmental toxicology and pharmacology. 2012; 33:121-6. doi: 10.1016/j.etap.2011.11.005.
7. Saini SS, Kaur A. Aflatoxin B1: Toxicity, characteristics and analysis: Mini review. Global Advanced Research Journal of Chemistry and Material Science. 2012; 1:063-70.
8. Grant D, Mendicino M, Levy G. Xenotransplantation: Just around the corner? Surgery. 2001; 129:243-7. doi: 10.1067/msy.2001.118380.
9. Rawal S, Ji EK, Coulombe R. Aflatoxin B 1 in poultry: Toxicology, metabolism and prevention. Research in veterinary science. 2010; 89:325-31. doi: 10.1016/j.rvsc.2010.04.011.
10. Preisler V, Caspary WJ, Hoppe F, Hagen R, Stopper H. Aflatoxin B1-induced mitotic recombination in L5178Y mouse lymphoma cells. Mutagenesis. 2000; 15:91-7. doi: 10.1093/mutage/15.1.91.
11. Aguilar F, Hussain SP, Cerutti P. Aflatoxin B1 induces the transversion of G--> T in codon 249 of the p53 tumor suppressor gene in human hepatocytes. Proceedings of the National Academy of Sciences. 1993; 90:8586-90.
12. Aguilar F, Harris C, Sun T, Hollstein M, Cerutti P. Geographic variation of p53 mutational profile in nonmalignant human liver. Science. 1994; 264:1317-9. doi: 10.1126/science.8191284.
13. Hussain I, Anwar J. A study on contamination of aflatoxin M 1 in raw milk in the Punjab province of Pakistan. Food Control. 2008; 19:393-5. doi: 10.1016/j.foodcont.2007.04.019.
14. Newberne PM, Butler WH. Acute and chronic effects of aflatoxin on the liver of domestic and laboratory animals: a review. Cancer Research. 1969; 29:236-50.
15. Richard JL. Some major mycotoxins and their mycotoxicoses—An overview. International journal of food microbiology. 2007; 119:3-10. doi: 10.1016/j.ijfoodmicro.2007.07.019.
16. Saraste A, Pulkki K. Morphologic and biochemical hallmarks of apoptosis. Cardiovascular research. 2000; 45:528-37. doi: 10.1016/S0008-6363(99)00384-3.
17. Hirsova P, Gores GJ. Death receptor-mediated cell death and proinflammatory signaling in nonalcoholic steatohepatitis. CMGH Cellular and Molecular Gastroenterology and Hepatology. 2015; 1:17-27. doi: 10.1016/j.jcmgh.2014.11.005.
18. Thompson CB. Apoptosis in the pathogenesis and treatment of disease. Science. 1995; 267:1456. doi: 10.1126/science.7878464.
19. Cummings M, Winterford C, Walker N, Sternberg S. Histology for pathologists. Apoptosis. 1997:3Á9.
20. Choi AM, Ryter SW, Levine B. Autophagy in human health and disease. New England Journal of Medicine. 2013; 368:651-62. doi: 10.1056/NEJMra1205406.
21. Bbosa GS, Kitya D, Odda J, Ogwal-Okeng J. Aflatoxins metabolism, effects on epigenetic mechanisms and their role in carcinogenesis. Health. 2013; 2013. doi: 10.4236/health.2013.510A1003.
22. Petit PX, Zamzami N, Vayssiere J, Mignotte B, Kroemer G, Castedo M. Implication of mitochondria in apoptosis. Molecular and Cellular Biochemistry. 1997; 174:185-8. doi: 10.1007/978-1-4615-6111-8_28.
23. Peng X, Yu Z, Liang N, Chi X, Li X, Jiang M, Fang J, Cui H, Lai W, Zhou Y. The mitochondrial and death receptor pathways involved in the thymocytes apoptosis induced by aflatoxin B1. Oncotarget. 2016; 7:12222-34. doi: 10.18632/oncotarget.7731.
24. Chen T, Cui H, Cui Y, Bai C, Gong T, Peng X. Cell-cycle blockage associated with increased apoptotic cells in the thymus of chickens fed on diets high in fluorine. Human & Experimental Toxicology. 2011; 30:685-92. doi: 10.1177/0960327110379022.
25. Wang K. Molecular mechanisms of hepatic apoptosis. Cell death & disease. 2014; 5:e996. doi: 10.1038/cddis.2013.499.
26. Jung-Hwan Y, Gores GJ. Death receptor-mediated apoptosis and the liver. Journal of Hepatology. 2002; 36:400. doi: 10.1016/S0168-8278(02)00209-X.
27. Chen K, Shu G, Peng X, Fang J, Cui H, Chen J, Wang F, Chen Z, Zuo Z, Deng J. Protective role of sodium selenite on histopathological lesions, decreased T-cell subsets and increased apoptosis of thymus in broilers intoxicated with aflatoxin B 1. Food and chemical toxicology. 2013; 59:446-54. doi: 10.1016/j.fct.2013.06.032.
28. Chen K, Peng X, Fang J, Cui H, Zuo Z, Deng J, Chen Z, Geng Y, Lai W, Tang L. Effects of dietary selenium on histopathological changes and T cells of spleen in broilers exposed to aflatoxin B1. International journal of environmental research and public health. 2014; 11:1904-13. doi: 10.3390/ijerph110201904.
29. Peng X, Zhang S, Fang J, Cui H, Zuo Z, Deng J. Protective Roles of Sodium Selenite against Aflatoxin B1-Induced Apoptosis of Jejunum in Broilers. International journal of environmental research and public health. 2014; 11:13130-43. doi: 10.3390/ijerph111213130.
30. Wang F, Shu G, Peng X, Fang J, Chen K, Cui H, Chen Z, Zuo Z, Deng J, Geng Y. Protective effects of sodium selenite against aflatoxin B1-induced oxidative stress and apoptosis in broiler spleen. International journal of environmental research and public health. 2013; 10:2834-44. doi: 10.3390/ijerph10072834.
31. He Y, Fang J, Peng X, Cui H, Zuo Z, Deng J, Chen Z, Lai W, Shu G, Tang L. Effects of Sodium Selenite on Aflatoxin B1-Induced Decrease of Ileac T cell and the mRNA Contents of IL-2, IL-6, and TNF-α in Broilers. Biological trace element research. 2014; 159:167-73. doi: 10.1007/s12011-014-9999-2.
32. Yang P, Hao Y, Feng J, Lin H, Feng Y, Wu X, Yang X, Gu X. The Expression of Carnosine and Its Effect on the Antioxidant Capacity of Longissimus dorsi Muscle in Finishing Pigs Exposed to Constant Heat Stress. Asian Australasian Journal of Animal Sciences. 2014; 27:1763-72. doi: 10.5713/ajas.2014.14063.
33. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 2015; 72:248-54.
34. Kumar R, Balachandran C. Histopathological changes in broiler chickens fed aflatoxin and cyclopiazonic acid. Vet Arhiv. 2009; 79:31-40.
35. Ibrahim Q. HISTOPATHOLOGICAL STUDY OF QUAILS LIVER EXPERIMENTALLY INDUCED BY AFLATOXIN. Basrah journal of veterinary research. 2013; 12:116-127.
36. Kerr JF, Wyllie AH, Currie AR. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. British journal of cancer. 1972; 26:239. doi: 10.1038/bjc.1972.33.
37. Peng X, Chen K, Chen J, Fang J, Cui H, Zuo Z, Deng J, Chen Z, Geng Y, Lai W. Aflatoxin B1 affects apoptosis and expression of Bax, Bcl-2, and CASPASE-3 in thymus and bursa of fabricius in broiler chickens. Environmental toxicology. 2015. doi: 10.1002/tox.22120.
38. Josse R, Dumont J, Fautrel A, Robin M-A, Guillouzo A. Identification of early target genes of aflatoxin B1 in human hepatocytes, inter-individual variability and comparison with other genotoxic compounds. Toxicology and applied pharmacology. 2012; 258:176-87. doi: 10.1016/j.taap.2011.10.019.
39. Hsu H, Shu H-B, Pan M-G, Goeddel DV. TRADD-TRAF2 and TRADD-FADD interactions define two distinct TNF receptor 1 signal transduction pathways. Cell. 1996; 84:299-308. doi: 10.1016/S0092-8674(00)80984-8.
40. Régnier CH, Song HY, Gao X, Goeddel DV, Cao Z, Rothe M. Identification and characterization of an IκB kinase. Cell. 1997; 90:373-83. doi: 10.1016/S0092-8674(00)80344-X.
41. Hacker H, Karin M. Regulation and function of IKK and IKK-related kinases. Sci Stke. 2006; 357:12-4. doi: 10.1126/stke.3572006re13.
42. Duckett CS, Li F, Wang Y, Tomaselli KJ, Thompson CB, Armstrong RC. Human IAP-like protein regulates programmed cell death downstream of Bcl-xL and cytochrome c. Molecular and cellular biology. 1998; 18:608-15.
43. Fan T-J, Han L-H, Cong R-S, Liang J. CASPASE family proteases and apoptosis. Acta biochimica et biophysica Sinica. 2005; 37:719-27. doi: 10.1111/j.1745-7270.2005.00108.x.
44. Hu Q, Wu D, Chen W, Yan Z, Yan C, He T, Liang Q, Shi Y. Molecular determinants of CASPASE-9 activation by the Apaf-1 apoptosome. Proceedings of the National Academy of Sciences. 2014; 111:16254-61. doi: 10.1073/pnas.1418000111.
45. Chen N, Chen X, Huang R, Zeng H, Gong J, Meng W, Lu Y, Zhao F, Wang L, Zhou Q. BCL-xL is a target gene regulated by hypoxia-inducible factor-1α. Journal of Biological Chemistry. 2009; 284:10004-12. doi: 10.1074/jbc.M805997200.
46. Meki A-RM, Abdel-Ghaffar SK, El-Gibaly I. Aflatoxin B1 induces apoptosis in rat liver: protective effect of melatonin. Neuroendocrinology Letters. 2001; 22:417-26.
47. Ribeiro DH, Ferreira FL, Da Silva VN, Aquino S, Corrêa B. Effects of aflatoxin B1 and fumonisin B1 on the viability and induction of apoptosis in rat primary hepatocytes. International journal of molecular sciences. 2010; 11:1944-55. doi: 10.3390/ijms11041944.
48. Kannan K, Jain SK. Oxidative stress and apoptosis. Pathophysiology. 2000; 7:153-63. doi: 10.1016/S0928-4680(00)00053-5.
49. Rüdiger von Harsdorf M. Reactive oxygen species and apoptosis. Apoptosis in Cardiac Biology. 2007. doi: 10.1007/978-0-585-38143-5_3.
50. Chen J, Chen K, Yuan S, Peng X, Fang J, Wang F, Cui H, Chen Z, Yuan J, Geng Y. Effects of aflatoxin B1 on oxidative stress markers and apoptosis of spleens in broilers. Toxicology and industrial health. 2013:0748233713500819. doi: 10.1177/0748233713500819.
51. Buttke TM, Sandstrom PA. Oxidative stress as a mediator of apoptosis. Immunology today. 1994; 15:7-10. doi: 10.1016/0167-5699(94)90018-3.
52. Kujoth G, Hiona A, Pugh T, Someya S, Panzer K, Wohlgemuth S, Hofer T, Seo A, Sullivan R, Jobling W. Mitochondrial DNA mutations, oxidative stress, and apoptosis in mammalian aging. Science. 2005; 309:481-4. doi: 10.1126/science.1112125.
53. Yang J, Bai F, Zhang K, Bai S, Peng X, Ding X, Li Y, Zhang J, Zhao L. Effects of feeding corn naturally contaminated with aflatoxin B1 and B2 on hepatic functions of broilers. Poultry science. 2012; 91:2792-801. doi: 10.3382/ps.2012-02544.
54. Wills E. Mechanisms of lipid peroxide formation in animal tissues. Biochemical Journal. 1966; 99:667. doi: 10.1042/bj0990667.
55. Kim Y-S, Kim Y-H, Noh J-R, Cho E-S, Park J-H, Son H-Y. Protective effect of korean red ginseng against aflatoxin B 1-induced hepatotoxicity in rat. Journal of ginseng research. 2011; 35:243-9. doi: 10.5142/jgr.2011.35.2.243.
56. Gowda N, Ledoux D, Rottinghaus G, Bermudez A, Chen Y. Efficacy of turmeric (Curcuma longa), containing a known level of curcumin, and a hydrated sodium calcium aluminosilicate to ameliorate the adverse effects of aflatoxin in broiler chicks. Poultry science. 2008; 87:1125-30. doi: 10.3382/ps.2007-00313.
57. Hou R, Xie P, Zhang M, Feng J, Zheng S, Ma A. Effects of Glucomannan on Liver Biochemical Indices and Hepatic Pathological Observation of Broilers Fed with Dietary Containing Aflatoxin B_1 [J]. Chinese Journal of Animal Nutrition. 2008; 2:007.
58. Council NR. (1994). National research council nutrient requirements of poultry: ninth revised edition. (Washington, DC, USA: National Academies Press).
59. Kaoud HA. Innovative methods for the amelioration of aflatoxin (AFB1) effect in broiler chicks. Sci J Appl Res. 2013; 1:15-9.
60. Chen T, Cui H, Cui Y, Bai C, Gong T, Peng X. Cell-cycle blockage associated with increased apoptotic cells in the thymus of chickens fed on diets high in fluorine. Human & experimental toxicology. 2011; 30:685-92. doi: 10.1177/0960327110379022.
61. Hong YH, Lillehoj HS, Lillehoj EP, Lee SH. Changes in immune-related gene expression and intestinal lymphocyte subpopulations following Eimeria maxima infection of chickens. Vet Immunol Immunopathol. 2006; 114:259-72. doi: 10.1016/j.vetimm.2006.08.006.
62. Shini S, Kaiser P. Effects of stress, mimicked by administration of corticosterone in drinking water, on the expression of chicken cytokine and chemokine genes in lymphocytes. Stress. 2009; 12:388-99. doi: 10.1080/10253890802526894.
63. Livak KJ, Schmittgen TD. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2 −ΔΔ C T Method. Methods. 2001; 25:402-8. doi: 10.1006/meth.2001.1262.