
INTRODUCTION
Ovarian cancer is the most deadly gynaecological cancer, with approximately 200,000 new cases diagnosed globally each year and more than 150,000 deaths due to the disease annually [1]. Epithelial ovarian cancer (EOC) accounts for 90 % of all ovarian cancers and typically presents in post-menopausal women [2]. The pathologic type of EOC includes clear cell carcinoma, mucinous carcinoma, endometrioid carcinoma, low-grade serous ovarian carcinoma and high-grade serous carcinoma [3]. Most patients are not diagnosed until the cancer is at an advanced stage. Standard treatment for EOC involves cytoreductive surgery followed by platinum-based chemotherapy. Nevertheless, recurrence is frequent (around 70%) and prognosis is globally poor [4].
Notch signaling pathway is an evolutionarily conserved signaling pathway, which is composed of receptors, ligands and intracellular domain. In canonical Notch signaling, a Notch transmembrane receptor interacts extracellularly with a canonical Notch transmembrane ligand on a contacting cell, initiating proteolytic cleavage of the receptor by γ-secretase and the subsequent release of the intracellular domain (ICD) of the receptor. Notch intracellular domain (NICD) then translocates to the nucleus and plays its biological functions. Notch pathway plays essential roles in regulating cell differentiation, cell-cell communication, organ development and so on [5].
Notch1 and NICD were frequently expressed in ovarian cancer cell lines and specimens, concluding that Notch1 plays a role in ovarian cancer proliferation [6]. High expression of Notch1 and Jagged1 in breast cancer is linked to poor survival rates, and Jagged 1 is highly expressed in metastatic prostate cancer as compared to localized or benign prostatic tissue [7–9]. Nevertheless, the mechanism of how Notch receptor, ligand and NICD function in carcinogenesis of ovarian cancer is still unclear.
Gamma-secretase inhibitors have been developed to block Notch signaling and have entered clinical trials. These compounds inhibit γ-secretases that cleave Notch and additional proteins, inhibit the proteasome and can elicit endoplasmic reticulum stress [10–14]. Siemers ER et al first reported the effects of a γ-secretase inhibitor in a randomized study of patients with Alzheimer disease [15]. A multinational phase III clinical trial of LY450139 (a γ-secretase inhibitor) is currently under development as a disease-modifying therapy for Alzheimer disease [16]. Gamma-secretase inhibitor is also considered as a promising medicine for cancer therapy [17]. N-[N-(3,5-Difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) is a γ-secretase inhibitor and commonly used to block Notch signaling [14]. Nethertheless, the enzymatic activity of γ-secretase in Notch1 signaling pathway and the mechanism of how γ-secretase inhibitor works on cancer cell are still unknown.
Here, we studied the expression of Notch1, Jagged1 and NICD in epithelial ovarian carcinoma tissues, analyzed the clinical significance and explored the potential anti-tumour effect of γ-secretase in epithelial ovarian carcinoma cell lines.
RESULTS
Jagged1 and NICD are highly expressed in ovarian carcinoma
We investigated the Notch1, Jagged1 and NICD immunohistochemical expression in all the 43 human ovarian cancer tissue specimens and 11 benign ovarian tumour tissue specimens. The percentage of expression of Notch1, Jagged1 and NICD in ovarian cancer specimens was 100% (43/43), 97.7% (42/43) and 100% (43/43) respectively, while the expression of Notch1, Jagged1 and NICD in benign ovarian tumour was detected in 10 (90.9%) specimens, 6 (54.5%) specimens and 1 (9.1%) specimen, respectively. There was not any significant different in the immunohistochemical scoring index (ICS) of Notch1 between ovarian cancer and benign ovarian tumour. Strikingly, the ICSs of Jagged1 and NICD in ovarian cancer were higher than in benign ovarian tumour (P < 0.01) (Table 1). Notch1 immunohistochemical expression was mostly confined to the cell membrane. Jagged1 immunohistochemical expression was mostly confined to the cell membrane and cytoplasm. NICD immunohistochemical expression was mostly confined to cytoplasm and nucleus (Figure 1).
Table 1: ICSs of Notch1, Jagged1 and NICD between ovarian cancer and benign ovarian tumour
Types |
Number |
Notch1 |
Jagged1 |
NICD |
|---|---|---|---|---|
Ovarian cancer |
43 |
6.7±2.2 |
5.3±2.4 |
5.3±2.3 |
Benign ovarian tumour |
11 |
5.4±2.7 |
1.6±1.4 |
3.1±1.7 |
P |
0.153a |
<0.01a |
<0.01a |
a Mann–Whitney test.
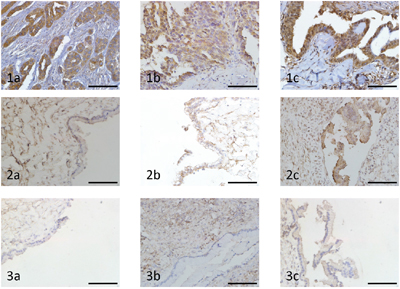
Figure 1: Immunohistochemical analysis of Notch1, Jagged1 and NICD expression in ovarian carcinoma and benign ovarian tumour (original magnification ×400, scale bar shows 50 μm). 1a. High expression of Notch1 in ovarian cancer tissue. 1b. High expression of Jagged1 in ovarian cancer tissue. 1c. High expression of NICD in ovarian cancer tissue. 2a. Low expression of Notch1 in benign ovarian tumour tissue. 2b. Low expression of Jagged1 in benign ovarian tumour tissue. 2c. Low expression of NICD in benign ovarian tumour tissue. 3a. Negative expression of Notch1 in benign ovarian tumour tissue. 3b. Negative expression of Jagged1 in benign ovarian tumour tissue. 3c. Negative expression of NICD in benign ovarian tumour tissue.
We next analyzed the relationship between the expression of Notch1 pathway and clinicopathologic factors of ovarian cancer. The results presented that the expressions of Notch1, Jagged1 and NICD in ovarian cancers were not correlated with age, family history, ascites, serum CA125, size of tumour, FIGO stage, differentiation and pathology (P > 0.05) (Table 2).
Table 2: Relationship between the expression of Notch1, Jagged1 and NICD and clinicopathologic factors of ovarian cancer
Number |
Notch1 positive expression |
P |
Jagged1 positive expression |
P |
NICD positive expression |
P |
||||
|---|---|---|---|---|---|---|---|---|---|---|
Number |
Percentage (%) |
Number |
Percentage (%) |
Number |
Percentage (%) |
|||||
Age |
0.688 |
0.565 |
0.775a |
|||||||
<55 |
30 |
30 |
100 |
30 |
100 |
30 |
100 |
|||
≥55 |
13 |
13 |
13/13c |
12 |
12/13c |
13 |
13/13c |
|||
Malignant cancer family historya |
0.414 |
0.392 |
0.869a |
|||||||
Positive |
32 |
32 |
100 |
31 |
97 |
32 |
100 |
|||
Negative |
11 |
11 |
11/11c |
11 |
11/11c |
11 |
11/11c |
|||
Ascites (ml) |
0.306 |
0.968 |
0.660a |
|||||||
<1 000 |
28 |
28 |
100 |
27 |
96 |
28 |
100 |
|||
≥1 000 |
15 |
15 |
15/15c |
15 |
15/15c |
15 |
15/15c |
|||
Serum CA125 (kU/L) |
0.520 |
0.050 |
0.524a |
|||||||
<1 000 |
32 |
32 |
100 |
32 |
100 |
32 |
100 |
|||
≥1 000 |
11 |
11 |
11/11c |
10 |
10/11c |
11 |
11/11c |
|||
Size of tumour (cm) |
0.731 |
0.553 |
0.608a |
|||||||
<10 |
23 |
23 |
100 |
23 |
100 |
23 |
100 |
|||
≥10 |
20 |
20 |
100 |
19 |
95 |
20 |
100 |
|||
FIGO stage |
0.095 |
0.861 |
0.772b |
|||||||
I |
7 |
7 |
7/7c |
6 |
6/7c |
7 |
7/7c |
|||
II |
4 |
4 |
4/4c |
4 |
4/4c |
4 |
4/4c |
|||
III |
28 |
28 |
100 |
28 |
100 |
28 |
100 |
|||
IV |
4 |
4 |
4/4c |
4 |
4/4c |
4 |
4/4c |
|||
Histology |
0.812 |
0.734 |
0.556b |
|||||||
Poorly differentiated |
10 |
10 |
10/10c |
10 |
10/10c |
10 |
10/10c |
|||
Moderately differentiated |
25 |
25 |
100 |
25 |
100 |
25 |
100 |
|||
Highly differentiated |
8 |
8 |
8/8c |
7 |
7/8c |
8 |
8/8c |
|||
Pathology subtypes |
0.757 |
0.205 |
0.186b |
|||||||
Serous carcinoma |
31 |
31 |
100 |
30 |
97 |
31 |
100 |
|||
Mucinous carcinoma |
4 |
4 |
4/4c |
4 |
4/4c |
4 |
4/4c |
|||
Endometrioid carcinoma |
5 |
5 |
5/5c |
5 |
5/5c |
5 |
5/5c |
|||
Clear cell carcinoma |
3 |
3 |
3/3c |
3 |
3/3c |
3 |
3/3c |
|||
a Mann–Whitney test.
b Kruskal–Wallis test.
c Number less than 20, not shown as percentage.
Expressions of Notch1, Jagged1 and NICD do not correlate with outcome in ovarian cancer
We tested for the relationship between the expression of Notch1, Jagged1 and NICD and patient survival. Table 3 summarizes the association of Notch1, Jagged1 and NICD expression data with the patient’s hazard ratio of survival. Patients with tumors expressing a high level of Notch1, Jagged1 and NICD were not more likely to die than patients with tumors expressing a moderate and low level of Notch1, Jagged1 and NICD (hazard ratio was 0.935, 1.236, and 1.104 respectively).
Table 3: Survival analysis of Notch1Hi/Notch1Mo+Lo, Jagged1Hi/Jagged1Mo+Lo and NICDHi/NICDMo+Lo
Expression |
n |
Hazard ratio (95% CI) |
Pa |
Notch1 |
43 |
0.935 (0.438-1.999) |
0.863 |
Jagged1 |
42 |
1.236 (0.497-3.074) |
0.656 |
NICD |
43 |
1.104 (0.419-2.910) |
0.842 |
95% CI 95% confidence interval, lo low, mo moderate, hi high.
a Cox proportional hazard model.
We next compared overall survival in patients whose tumors expressed high levels of Notch1, Jagged1 and NICD (NoHi, JaHi and NIHi) to patients with tumors expressing moderate and low levels of Notch1, Jagged1 and NICD (NoMo+Lo, JaMo+Lo and NIMo+Lo). Patients harboring tumors with NoHi, JaHi and NIHi had similar 5-year survival rates compared with tumors with NoMo+Lo, JaMo+Lo and NIMo+Lo (P > 0.05). (Table 4 and Figure 2).
Table 4: Comparison of tumors expressing high and moderate+low levels of Notch1, Jagged1 and NICD
Expression |
n |
5-year survival (%) |
Mean survival time (mo) (95%CI) |
Pa |
|---|---|---|---|---|
Notch1 |
||||
NoMo+Lo |
26 |
46.15 |
40.5(24.3-.) |
0.863 |
NoHi |
17 |
41.18 |
34.9(23.1-.) |
|
Jagged1 |
||||
JaMo+Lo |
33 |
45.45 |
42.1(27.1-.) |
0.649 |
JaHi |
9 |
33.33 |
31.5(12.7-.) |
|
NICD |
||||
NIMo+Lo |
35 |
45.71 |
42.1(28.8-78.7) |
0.842 |
NIHi |
8 |
37.50 |
28.6(10.2-.) |
95% CI 95% confidence interval, lo low, mo moderate, hi high.
a Log rank test.
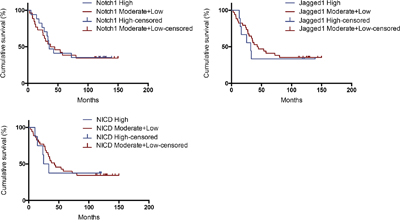
Figure 2: Kaplan–Meier analysis of patients with high vs. moderate+low expression of Notch1, Jagged1 and NICD.
The enzymatic activity of γ-secretase in ovarian cancer cell lines is significantly higher than in normal ovarian epithelial cell line
In order to confirm if γ-secretase in Notch1 pathway plays a role in the carcinogenesis of ovarian cancer, we detected the relative enzymatic activity of γ-secretase by dual-luciferase reporter assay system. The activity of γ-secretase in T29 was supposed as 1.00. The relative activity of γ-secretase in SKOV3, CAOV3 and ES2 was 12.60, 13.80 and 15.73, respectively, which was significantly higher than the activity of γ s-secretase in T29. (Figure 3)
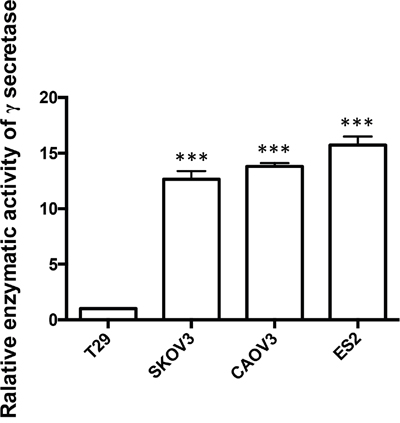
Figure 3: The relative enzymatic activity of γ-secretase in SKOV3, CAOV3, ES2 and T29 cell lines. The relative enzymatic activity of γ-secretase of T29 was set as 1. *** p < 0.001.
DAPT reduces the enzymatic activity of γ-secretase in ovarian cancer cell lines
In order to determine if DAPT can reduce the enzymatic activity of γ-secretase in ovarian cancer cells and benign ovarian epithelial cell, we tested the enzymatic activity of γ-secretase in SKOV3, CAOV3, ES2 and T29 after DAPT treatment with different concentrations and different time. Interestingly, after 25–100 μM of DAPT was added for 24 h or 48 h, the enzymatic activity of γ-secretase in SKOV3, CAOV3 and ES2 cell lines declined significantly in a dose- and time-dependent manner (P < 0.01). Also, the enzymatic activity of γ-secretase in T29 cell line declined after DAPT treatment. (Figure 4)
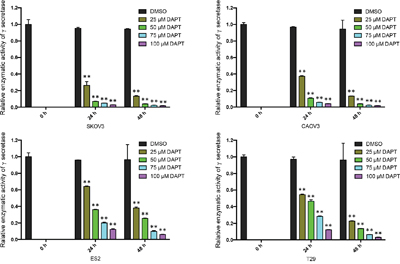
Figure 4: The relative enzymatic activity of γ-secretase in ovarian cancer cell lines and normal ovarian epithelial cell line after treatment of DAPT. The relative enzymatic activity of γ-secretase at 0 h was set as 1 in each group. ** p < 0.01.
DAPT inhibits the proliferation of ovarian cancer cell lines
Next, in order to determine if DAPT exhibits inhibitory effect on ovarian cancer cell proliferation, we investigated the role of DAPT in proliferation of ovarian cancer cell lines. After SKOV3, CAOV3 and ES2 cells were treated with 25–100 μM DAPT for 24 h or 48 h, an MTT assay was performed to detect cell proliferation. MTT data showed that DAPT (50 μM, 75 μM and 100 μM) inhibited the proliferation of SKOV3 and CAOV3 cell lines 24 h and 48 h after DAPT treatment compared with control group (P < 0.05). DAPT (75 μM and 100 μM) 24 h and DAPT (25 μM, 50 μM, 75 μM and 100 μM) 48 h after treatment inhibited the proliferation of ES2 cell line compared with control group (P < 0.05). DAPT (25 μM) inhibited the proliferation of SKOV3, CAOV3 and ES2 cell lines only 48 h after DAPT treatment (P < 0.05). However, DAPT (25–100 μM) showed no inhibitory effect on the proliferation of T29 cell line at 24 h or 48 h after DAPT treatment (P > 0.05). (Figure 5)
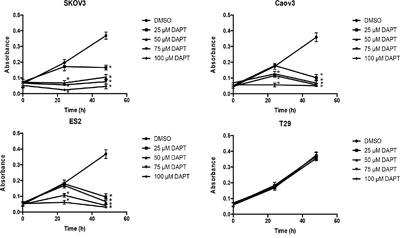
Figure 5: Effect of different concentrations of DAPT on the proliferation of ovarian cancer cell lines and normal cell line. * p < 0.05.
DAPT increases the apoptosis of ovarian cancer cell lines
To test the effect of DAPT on ovarian cancer cell apoptosis, flow cytometry was performed. It is found that DAPT decreased the PI and SPF and increased the AI in SKOV3, CAOV and ES2 cell lines as concentration of DAPT increased and time after DAPT treatment prolonged. The initiation concentration of DAPT decreased the PI and SPF in SKOV3, CAOV3 and ES2 cell lines was 50 μM, 50 μM and 75 μM at 24 h after DAPT treatment (P < 0.05). The initiation concentration of DAPT increased the AI in SKOV3, CAOV3 and ES2 cell lines was all 25 μM at 24 h after DAPT treatment (P < 0.05). However, DAPT (25–100 μM) did not change the PI, SPF and AI in T29 cell line at 24 h or 48 h after DAPT treatment (P > 0.05). (Figure 6)
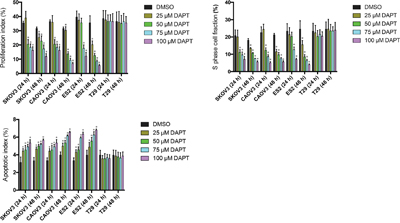
Figure 6: Effect of different concentrations of DAPT on the PI, SPF and AI of ovarian cancer cell lines and normal cell line. * p < 0.05.
DISCUSSION
In 1991, Ellen LW et al found that Notch gene was broken by chromosomal translocation in T lymphoblastic neoplasms, which opened a new era of Notch signaling and malignancies [18]. Rose SL et al found abundant NICD expression in ovarian cancer cell lines, as well as in 16 of 21 (76%) human ovarian cancer samples. The expression of NICD was greatly reduced in ovarian cancer cell lines after treatment with Notch1 siRNA. Furthermore, depletion of Notch1 led to growth inhibition of ovarian cancer cell lines, which provide evidence for target therapy of ovarian cancer by inhibiting Notch pathway [6]. However, Hopfer O et al concluded that Notch1 pathway was expressed in ovarian adenomas and cancers and the transcription factor hairy and enhancer of split 1 (HES1) was strongly expressed in ovarian cancers [19]. In this research, we found that Notch1 was widely expressed in both epithelial ovarian cancers and benign tumours, while the ICSs of Jagged1 and NICD in ovarian cancer were higher than in benign ovarian tumour. A possible reason is that Notch signaling pathway is an evolutionarily conserved signaling pathway. The combination of ligand and receptor activated Notch signaling and receptor was cleaved into NICD by γ-secreatase. NICD interacted with HES1 or other signaling such as Ras and PI3K/Akt and played an oncogenic role in ovarian cancer [20, 21]. Notch pathway cannot be triggered as carcinogenesis without the combination of ligand and Notch receptor.
Parr C et al found that Notch1 expression was low in grade 1 breast cancers and increased in poorly-differentiated breast cancers [22]. Wang M et al found that expression of Notch1 increased gradually with the poor differentiating of cancer tissues and the increasing of FIGO stage in ovarian cancer tissues [23]. In this research, we found the expressions of Notch1, Jagged1 and NICD in ovarian cancers were not correlated with clinicopathologic factors, which needs more specimens to be demonstrated.
Lin JT et al found that high-level coexpression of Notch1 and Jagged1 was associated with poor overall survival in patients with head and neck cancer [24]. Wu K et al found that high Jagged1 expression was statistically linked to reduced overall and disease-free survival in clear cell renal cell carcinoma patients, especially at the early stage [25]. Xu X et al concluded that the activation of Notch1 pathway might indicate a poor prognosis in acute myeloid leukemia. Especially, Notch1, Jagged1 and Delta1 expression might be relevant prognostic markers in intermediate risk acute myeloid leukemia [26]. In our research, we did not find a correlation between expressions of Notch1, Jagged1 and NICD and survival of ovarian cancer patients, although there seems to be a tendency that high expression of Jagged1 may have a poor survival (Figure 2). A possible reason is that the number of ovarian cancer patients included in this research is low (n = 43). In future study, in order to obtain more significant result, the number of cancer patients should be increased and the time of follow-up should be prolonged.
Gamma secretase is the key element in Notch pathway and the oncogenic role of Notch signaling relies on the cleavage of Notch receptor by γ-secretase. We haven’t found previous report on detecting the enzymatic activity of γ-secretase in Notch pathway related ontogenesis. In this research, the enzymatic activity of γ-secretase was detected by Dual Luciferase Reporter Assay. Plasmid Notch1 ΔE-GVP expresses Notch1 plus transcription activator Gal4-VP16. Plasmid MH100 expresses luciferase and is used as a reporter. In the presence of γ-secretase, Notch1 ΔE-GVP plus Gal4-VP16 is cleaved into intracellular domain plus Gal4-VP16 and translocates to the nucleus and activates transcription from a UAS promoter element. Luciferase expressed by MH100 is activated by USA promoter element [27–29]. We found that the enzymatic activity of γ-secretase in ovarian cancer cell was significantly higher than in normal ovarian epithelial cell, which demonstrates γ-secretase gets involved in the carcinogenesis of ovarian cancer. We firstly used Dual Luciferase Reporter Assay to detect enzymatic activity of γ-secretase, which could be widely used in cancer research. The therapeutic effect of γ-secretase inhibitor was also demonstrated on DAPT treated ovarian cancer cells. Chen X et al reported sequential combination therapy of ovarian cancer with cisplatin and γ-secretase inhibitor MK-0752 in cell and mice models [30]. In this research, we found that DAPT had therapeutic effect by inhibiting proliferation and increasing apoptosis of ovarian cancer cells. Interestingly, DAPT showed no effect on proliferation and apoptosis of the normal ovarian surface epithelial cell line T29 although DAPT reduced its enzymatic activity of γ-secretase, possibly because Notch pathway may not be essential for normal ovarian epithelial cells. Presumably, DAPT may minimize the side effect on normal ovarian tissue. Therefore, γ-secretase inhibitor may become a highly promising novel experimental therapeutic strategy against ovarian cancer in the field of precision medicine.
MATERIALS AND METHODS
Patients and specimens
Forty-three human ovarian tissue specimens were obtained from patients who underwent surgical resection of ovarian from the Department of Obstetrics and Gynecology of Peking University First Hospital, Beijing, China, between March 2004 and July 2007. At the time of surgery, no patient had received chemotherapy or radiotherapy. Histomorphology of all specimens was confirmed by the Department of Pathology, Peking University First Hospital. Tumours were staged according to the Federation International of Gynecology and Obstetrics (FIGO) staging of ovarian cancer in 2014. Benign epithelia ovarian tumour tissues were obtained from patients undergoing surgery for benign ovarian cyst. A total of 54 specimens were analyzed, including 43 ovarian epithelial carcinoma tissues, and 11 benign ovarian epithelial tumour tissues. The ovarian cancer patients were followed up for a median 63.8 months (range 2.9–250.1 months) and ended at July 5 2016.
Immunohistochemistry
Expression of Notch1, Jagged1 and Notch1 ICD in the pathologic sections was detected by an immunoperoxidase method. The paraffin-embedded ovarian tissue sections were deparaffinized with xylene. Peroxidase activity was quenched using 3.0% hydrogen peroxide. The antigen retrieval was by microwaving sections in 0.01 M sodium citrate, pH 6.0. Sections were incubated overnight with goat anti-human Notch1 (1:100; Santa Cruz), goat anti-human Jagged1 (1:100; Santa Cruz) and rabbit anti-human NICD (1:100; Millipore) at 4°C. The color development was using ABC kit.
ICS was then calculated by multiplying the intensity and percentage scores to determine the following results: 0, 1, 2, 3, 4, 6, and 9. In statistical analyses, ICS of 0 and 1 were defined as negative (-), ICS of 2 and 3 were defined as low expression (+), ICS of 4 and 6 were defined as mediate expression (++), and ICS of 9 was defined as high expression (+++) [31, 32].
Cell culture
Human ovarian adenocarcinoma cell lines cell line SKOV3 and human clear ovarian cancer cell line ES2 were cultured in RPMI1640 medium with 10 % fetal bovine serum. Human ovarian adenocarcinoma cell line CAOV3 was cultured in DMEM with 10 % fetal bovine serum. Human immortalized ovarian surface epithelial cell line T29 was cultured in Medium 199 and MCDB 105 Medium. All media contained 1% penicillin and streptomycin. All cell lines were kindly provided by Center of Gynecologic Oncology, Peking University People’s Hospital, Beijing, China.
Dual luciferase reporter assay system
A Dual-Luciferase Reporter Assay (Promega) was used to detect the relative enzymatic activity of γ-secretase. SKOV3, CAOV, ES2 and T29 cells were seeded in 96-well plates at 105 cells per well. Then the cells were transfected respectively with 200ng MH100, 100ng Notch1 ΔE-GVP and 2ng pRL-CMV using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol. The experiment was independently repeated at least three times. The transfected cells were collected in lysis buffer. Luciferase activity in triplicate samples for each condition was measured with a luminometer. Renilla luciferase activity was used to normalize the firefly luciferase activity.
MTT assay
An MTT assay was performed to assess the effect of DAPT on cell proliferation. Briefly, the cells were added to 96-well plates and then treated with DAPT (25–100 μM, Sigma-Aldrich) and DMSO (Sigma-Aldrich) for 24–48 h. Next, the MTT reagent was introduced to each well, and the supernatants were removed 4 h later. A total of 150 μL DMSO was used to dissolve the resultant formazan crystals. The absorption was read at 490 nm using a spectrophotometer.
Flow cytometry
Cells were added treated with DAPT (25–100 μM) and DMSO for 24–48 h respectively and analyzed by flow cytometry. S phase cell fraction (SPF) was calculated according to the equation: SPF=S/(S+G1+G2)×100%. Proliferation index (PI) was calculated according to the equation: PI=(S+G2)/ (S+G1+G2)×100%. Apoptosis was presented as apoptotic index (AI), which is expressed as the ratio of apoptotic cells to all the cells checked as a percentage.
Statistical analysis
A comparison of protein expression was evaluated with Mann-Whitney test. A comparison of clinicopathologic characteristics was evaluated with the Mann-Whitney test and Kruskal-Wallis test. The survival rate was calculated by the Kaplan-Meier method. The Cox proportional hazard regression model was used to determine the joint effects of several variables on survival. The differences between the survival curves were tested by the log rank test. The activity of enzymatic activity of γ-secretase, cell proliferation and apoptosis were analyzed by t test. All statistical analyses were performed with SPSS for Windows version 22.0 (SPSS, Chicago, IL, USA).
ACKNOWLEDGMENTS
We thank Prof. Heng Cui of Center of Gynecologic Oncology, Peking University People’s Hospital, Beijing, China, for providing us all the cells used in this research. We also thank Prof. Helena Karlström of Center for Alzheimer Research, Division of Neurogeriatrics, Department of Neurobiology, Care Sciences and Society, Karolinska Institutet, Stockholm, Sweden, for providing us plasmid Notch1 ΔE-GVP and MH100 (UAS-luc). We thank Prof. Jun Xie of Department of Biochemistry and Molecular Biology, Shanxi Medical University, Shanxi, China, for his guidance on testing enzymatic activity of γ-secretase. We thank Thomas C. Raissi from University of Western Ontario for his linguistic advice during the preparation of this manuscript.
CONFLICTS OF INTEREST
All the authors have declared no conflicts of interest.
REFERENCES
1. Global Burden of Disease Cancer C, Fitzmaurice C, Dicker D, Pain A, Hamavid H, Moradi-Lakeh M, MacIntyre MF, Allen C, Hansen G, Woodbrook R, Wolfe C, Hamadeh RR, Moore A, et al. The Global Burden of Cancer 2013. JAMA oncology. 2015; 1:505-527.
2. Papa A, Caruso D, Strudel M, Tomao S, Tomao F. Update on Poly-ADP-ribose polymerase inhibition for ovarian cancer treatment. Journal of translational medicine. 2016; 14:267.
3. Prat J, Oncology FCoG. Staging classification for cancer of the ovary, fallopian tube, and peritoneum. International journal of gynaecology and obstetrics. 2014; 124:1-5.
4. Mittica G, Genta S, Aglietta M, Valabrega G. Immune Checkpoint Inhibitors: A New Opportunity in the Treatment of Ovarian Cancer? International journal of molecular sciences. 2016; 17.
5. Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch signaling: cell fate control and signal integration in development. Science. 1999; 284:770-776.
6. Rose SL, Kunnimalaiyaan M, Drenzek J, Seiler N. Notch 1 signaling is active in ovarian cancer. Gynecologic oncology. 2010; 117:130-133.
7. Reedijk M, Odorcic S, Chang L, Zhang H, Miller N, McCready DR, Lockwood G, Egan SE. High-level coexpression of JAG1 and NOTCH1 is observed in human breast cancer and is associated with poor overall survival. Cancer research. 2005; 65:8530-8537.
8. Rizzo P, Miao H, D'Souza G, Osipo C, Song LL, Yun J, Zhao H, Mascarenhas J, Wyatt D, Antico G, Hao L, Yao K, Rajan P, et al. Cross-talk between notch and the estrogen receptor in breast cancer suggests novel therapeutic approaches. Cancer research. 2008; 68:5226-5235.
9. Santagata S, Demichelis F, Riva A, Varambally S, Hofer MD, Kutok JL, Kim R, Tang J, Montie JE, Chinnaiyan AM, Rubin MA, Aster JC. JAGGED1 expression is associated with prostate cancer metastasis and recurrence. Cancer research. 2004; 64:6854-6857.
10. Dovey HF, John V, Anderson JP, Chen LZ, de Saint Andrieu P, Fang LY, Freedman SB, Folmer B, Goldbach E, Holsztynska EJ, Hu KL, Johnson-Wood KL, Kennedy SL, et al. Functional gamma-secretase inhibitors reduce beta-amyloid peptide levels in brain. Journal of neurochemistry. 2001; 76:173-181.
11. Marambaud P, Shioi J, Serban G, Georgakopoulos A, Sarner S, Nagy V, Baki L, Wen P, Efthimiopoulos S, Shao Z, Wisniewski T, Robakis NK. A presenilin-1/gamma-secretase cleavage releases the E-cadherin intracellular domain and regulates disassembly of adherens junctions. The EMBO journal. 2002; 21:1948-1956.
12. Olsauskas-Kuprys R, Zlobin A, Osipo C. Gamma secretase inhibitors of Notch signaling. OncoTargets and therapy. 2013; 6:943-955.
13. Rosati E, Sabatini R, De Falco F, Del Papa B, Falzetti F, Di Ianni M, Cavalli L, Fettucciari K, Bartoli A, Screpanti I, Marconi P. gamma-Secretase inhibitor I induces apoptosis in chronic lymphocytic leukemia cells by proteasome inhibition, endoplasmic reticulum stress increase and notch down-regulation. International journal of cancer. 2013; 132:1940-1953.
14. Dorneburg C, Goss AV, Fischer M, Roels F, Barth TF, Berthold F, Kappler R, Oswald F, Siveke JT, Molenaar JJ, Debatin KM, Beltinger C. gamma-secretase inhibitor I inhibits neuroblastoma cells, with NOTCH and the proteasome among its targets. Oncotarget. 2016; 7:62799-62813. doi: 10.18632/oncotarget.11715.
15. Siemers ER, Quinn JF, Kaye J, Farlow MR, Porsteinsson A, Tariot P, Zoulnouni P, Galvin JE, Holtzman DM, Knopman DS, Satterwhite J, Gonzales C, Dean RA, May PC. Effects of a gamma-secretase inhibitor in a randomized study of patients with Alzheimer disease. Neurology. 2006; 66:602-604.
16. Hsu CK, Hsu CC, Lee JY, Kuo YM, Pai MC. Exacerbation of psoriatic skin lesions in a patient with Alzheimer disease receiving gamma-secretase inhibitor. Journal of the American Academy of Dermatology. 2013; 68:e46-48.
17. Lewis HD, Leveridge M, Strack PR, Haldon CD, O'Neil J, Kim H, Madin A, Hannam JC, Look AT, Kohl N, Draetta G, Harrison T, Kerby JA, Shearman MS, Beher D. Apoptosis in T cell acute lymphoblastic leukemia cells after cell cycle arrest induced by pharmacological inhibition of notch signaling. Chemistry & biology. 2007; 14:209-219.
18. Ellisen LW, Bird J, West DC, Soreng AL, Reynolds TC, Smith SD, Sklar J. TAN-1, the human homolog of the Drosophila notch gene, is broken by chromosomal translocations in T lymphoblastic neoplasms. Cell. 1991; 66:649-661.
19. Hopfer O, Zwahlen D, Fey MF, Aebi S. The Notch pathway in ovarian carcinomas and adenomas. British journal of cancer. 2005; 93:709-718.
20. Weijzen S, Rizzo P, Braid M, Vaishnav R, Jonkheer SM, Zlobin A, Osborne BA, Gottipati S, Aster JC, Hahn WC, Rudolf M, Siziopikou K, Kast WM, Miele L. Activation of Notch-1 signaling maintains the neoplastic phenotype in human Ras-transformed cells. Nature medicine. 2002; 8:979-986.
21. Wei Y, Zhang Z, Liao H, Wu L, Wu X, Zhou D, Xi X, Zhu Y, Feng Y. Nuclear estrogen receptor-mediated Notch signaling and GPR30-mediated PI3K/AKT signaling in the regulation of endometrial cancer cell proliferation. Oncology reports. 2012; 27:504-510.
22. Parr C, Watkins G, Jiang WG. The possible correlation of Notch-1 and Notch-2 with clinical outcome and tumour clinicopathological parameters in human breast cancer. International journal of molecular medicine. 2004; 14:779-786.
23. Wang M, Wang J, Wang L, Wu L, Xin X. Notch1 expression correlates with tumor differentiation status in ovarian carcinoma. Medical oncology. 2010; 27:1329-1335.
24. Lin JT, Chen MK, Yeh KT, Chang CS, Chang TH, Lin CY, Wu YC, Su BW, Lee KD, Chang PJ. Association of high levels of Jagged-1 and Notch-1 expression with poor prognosis in head and neck cancer. Annals of surgical oncology. 2010; 17:2976-2983.
25. Wu K, Xu L, Zhang L, Lin Z, Hou J. High Jagged1 expression predicts poor outcome in clear cell renal cell carcinoma. Japanese journal of clinical oncology. 2011; 41:411-416.
26. Xu X, Zhao Y, Xu M, Dai Q, Meng W, Yang J, Qin R. Activation of Notch signal pathway is associated with a poorer prognosis in acute myeloid leukemia. Medical oncology. 2011; 28:S483-489.
27. Karlstrom H, Bergman A, Lendahl U, Naslund J, Lundkvist J. A sensitive and quantitative assay for measuring cleavage of presenilin substrates. The Journal of biological chemistry. 2002; 277:6763-6766.
28. Taniguchi Y, Karlstrom H, Lundkvist J, Mizutani T, Otaka A, Vestling M, Bernstein A, Donoviel D, Lendahl U, Honjo T. Notch receptor cleavage depends on but is not directly executed by presenilins. Proceedings of the National Academy of Sciences of the United States of America. 2002; 99:4014-4019.
29. Saxena MT, Schroeter EH, Mumm JS, Kopan R. Murine notch homologs (N1-4) undergo presenilin-dependent proteolysis. The Journal of biological chemistry. 2001; 276:40268-40273.
30. Chen X, Gong L, Ou R, Zheng Z, Chen J, Xie F, Huang X, Qiu J, Zhang W, Jiang Q, Yang Y, Zhu H, Shi Z, Yan X. Sequential combination therapy of ovarian cancer with cisplatin and gamma-secretase inhibitor MK-0752. Gynecologic oncology. 2016; 140:537-544.
31. Ferrari A, Petterino C, Ratto A, Campanella C, Wurth R, Thellung S, Vito G, Barbieri F, Florio T. CXCR4 expression in feline mammary carcinoma cells: evidence of a proliferative role for the SDF-1/CXCR4 axis. BMC veterinary research. 2012; 8:27.
32. Menczer J, Schreiber L, Sukmanov O, Czernobilsky B, Berger E, Golan A, Levi T. COX-2 expression in nonepithelial ovarian malignancies. International journal of gynecological pathology. 2011; 30:41-45.