
INTRODUCTION
One of the major problems in the current cancer therapies is poor tumor targeting specificity. To address this limitation, bacterial cancer therapy was developed. Salmonella enterica serovar Typhimurium was reported to preferentially accumulate in tumors and inhibit tumor growth [1]. However, wild-type S. enterica is highly pathogenic for humans. As such, avirulent S. enterica A1-R and VNP20009 were developed for therapeutic applications. A1-R is auxotrophic for leucine and arginine. And it has been proven to eradicate primary and metastatic tumors in nude mouse models of prostate, breast, ovarian and pancreatic cancer, etc. [2–7]. VNP20009, with double mutations in msbB and purI, is another attenuated S. enterica and has been proven safe in phase I clinical trials. Furthermore, VNP20009 was most wildly used to deliver anticancer agents and carry shRNA-expressing plasmids [8–12]. However, VNP20009 failed to target tumors and inhibit tumor growth in the phase I study. Therefore, additional studies are required to study the factors affecting tumor colonization and anti-tumor effects. To investigate this, a high-throughput method was used to screen for S. enterica promoters and genes that expressed in tumors [13, 14]. Furthermore, in vivo gene mutation assay showed that SPI-2, invA and flagella (fla) are essential for the anticancer ability of S. enterica [15, 16].
Macrophages play a pivotal role in bacterial clearance during an infection. After being phagocytosed by macrophages, S. enterica modify the vacuoles where they reside and subsequently disseminate throughout the body [17]. Fileds and his colleagues showed that the mutants that cannot survive within macrophages are avirulent [18]. As such, the capacity of S. enterica to cause systematic infection relies on its ability to survive and replicate within macrophages. However, the anticancer ability of the mutants lacking macrophage survival ability has not been well studied.
In this study, we knocked out three genes related to VNP20009 survival within macrophages. The gene slyA and STM3120 are transcriptional regulators and HtrA is a stress protein for degrading the damaged and misfolded proteins. These genes perform different functions for bacterial survival within macrophages. Although ΔslyA, ΔSTM3120 and ΔhtrA displayed reduced fitness in spleen and unchanged fitness in tumors, their anticancer capacities were partially or completely abrogated. Therefore, we further examined the immune responses by comparing immune cells and cytokines in tumor infected by VNP20009 with the mutants, attempting to elucidate the mechanism of the mutants with reduced anti-cancer capacity.
RESULTS
In vitro characterization of ΔslyA, ΔSTM3120 and ΔhtrA
Because the genes slyA, STM3120 and htrA were reported to play vital roles in macrophage survival, each of the mutants was characterized using a gentamicin protection assay. As shown in Figure 1C, the uptake of bacteria was not impaired in the mutant groups compared with that in the VNP20009 group. However, in contrast to VNP20009, the replication rates of all mutants were decreased over five hours. VNP20009 displayed a 2.5- to 10- fold increase in replication rate in macrophages at five hour post infection compared with the mutant groups (Figure 1D).
These mutants were then characterized by probing their growth curve in vitro, and the growth of mutants in LB showed no defects (Figure 1A). In addition, both the mutants and VNP20009 were able to infect B16F10 cells, and there were no significant differences between the mutants and VNP20009 with respect to replication rates (Figure 1B). Collectively, these results suggested that the genes slyA, STM3120 and htrA were not required for VNP20009 replication in melanoma cells, but were vital for VNP20009 to survive within macrophages.
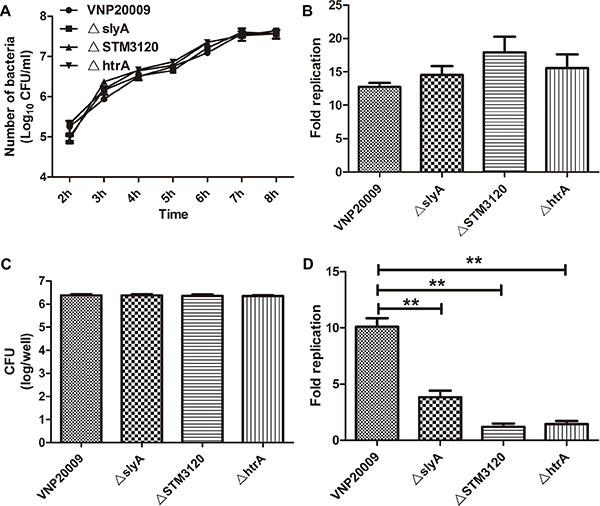
Figure 1: Characterization of VNP20009 and its mutants. (A) Growth rates of VNP20009 and its mutants in vitro. Date are shown in mean ± SEM. (B) Replication rates of VNP20009 and its mutants in B16F10 cells. Date are shown in mean ± SEM. (C) The number of intracellular bacteria in RAW 264.7 cell after invasion by VNP20009 and its mutants. Date are shown in mean ± SEM. (D) Replication rates of VNP20009 and its mutants within RAW 264.7 cell. Date are shown in mean ± SEM, **P < 0.01.
The biodistribution of the mutants in tumor bearing mice
To determine the biodistribution of the mutants, 1 × 105 colony forming unit (CFU) S. enterica were inoculated into tumor-bearing mice when the tumor size reached 150 mm3. VNP20009 and all of the mutants targeted the tumor at similar levels, reaching above 109 CFU/g in tumor 3 days post infection (P > 0.05) (Figure 2A). However, the bacterial loads of the mutants were significantly lower than that of VNP20009 in spleen, which is the main target of bacteria (P < 0.05) (Figure 2B). Additional calculations were performed to determine the ratio between the bacterial loads in tumor and spleen. The ratios of the ΔslyA, ΔSTM3120 and ΔhtrA mutants were ten times higher than that of VNP20009 (Figure 2C). Overall, ΔslyA, ΔSTM3120 and ΔhtrA displayed greater therapeutic potentials, as the mutants showed reduced bacterial accumulation in normal tissues and unchanged tumor fitness.
The anticancer capacities of the mutants
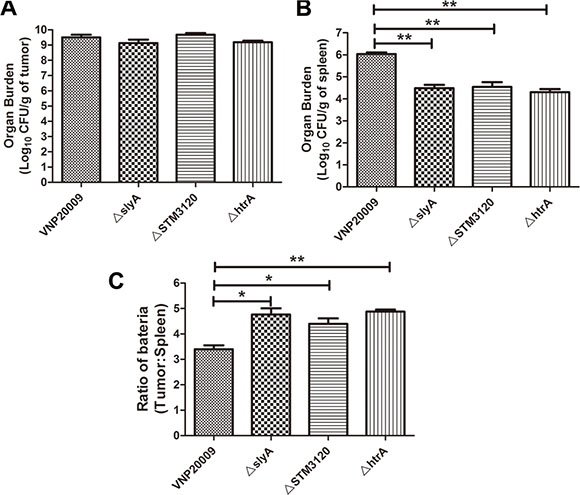
Figure 2: Biodistribution of VNP20009 and its mutants in tumor-bearing mice. VNP20009 and its mutants were intraperitoneally inoculated into the tumor-bearing mice. The bacterial titer in spleen and tumor were determined on the third day post infection. (A) Bacterial burden of ΔslyA, ΔSTM3120 and ΔhtrA mutants in tumor compared with VNP20009. Date are shown in mean ± SEM. (B) Bacterial burden of ΔslyA, ΔSTM3120 and ΔhtrA mutants in spleen compared with VNP20009. Date are shown in mean ± SEM, **P < 0.01. (C) Tumor/spleen ratio of VNP20009 and the mutants in the tumor bearing mice. Date are shown in mean ± SEM, *P < 0.05, **P < 0.01.
Because the mutants displayed a higher tumor to spleen ratio than VNP20009, their anticancer activities were further investigated. The tumor suppression capacities of the mutants were partially or completely eliminated (P < 0.05) (Figure 3). On the 9th day of infection, the tumor volumes of mice treated with VNP20009 were significantly lower than those in the PBS group (Figure 3A). Tumor doubling time and tumor growth delay were significantly increased in the mice treated with VNP20009 than those in the PBS group (P < 0.05) (Figure 3B and 3C). However, the mice infected with ΔslyA and ΔSTM3120 showed only a partial tumor growth delay compared with those infected with VNP20009 (P < 0.05). More importantly, ΔhtrA failed to suppress tumor growth. The tumor doubling time was remarkably decreased from 2.69 days (CI, 2.78 d to 2.51 d) in the VNP20009 group to 2.4 days (CI, 2.52 d to 2.28 d) in the ΔhtrA group (Figure 3C). The tumor growth delay was significantly decreased from 23.67 days (CI, 22.99 d to 24.47 d) in the VNP20009 group to 17.69 days (CI, 17.59 d to 17.70 d) in the ΔhtrA group (Figure 3B).
In order to definitively link the genes slyA, STM3120 and htrA to the diminished antitumor activities, the anticancer activities of ΔslyA, ΔSTM3120 and ΔhtrA mutants were studied after complementation. Intact copies of the genes were cloned into pHQ-003-F, and the complementing plasmids were transformed into the corresponding mutants, respectively. After confirming the proteins were precisely expressed, we next assessed their antitumor properties (Figure 3D). As shown in Figure 3E, 3F and 3G, the anticancer capacities of the mutants were partially or completely reversed when the mutants were complemented with a copy of the corresponding gene. The presence of the vector alone did not improve the anticancer ability. These results indicated that the genes slyA, STM3120 and htrA are vital for the anticancer ability of VNP20009.
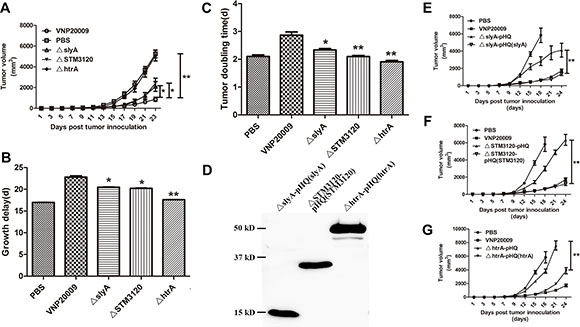
Figure 3: Antitumor effects of VNP20009 and its mutants. S. enterica (1 × 105 CFU/mouse) were injected intraperitoneally into the tumor-bearing mice when the tumor size reached 150 mm3. Tumor volumes of different groups were compared. (A) Tumor volumes of the mice treated with ΔslyA, ΔSTM3120 and ΔhtrA compared with that treated with VNP20009. Date are shown in mean ± SEM, *P < 0.05, **P < 0.01. (B) Tumor growth delay of the mice treated with ΔslyA, ΔSTM3120 and ΔhtrA versus VNP20009. Date are shown in mean ± SEM, *P < 0.05, **P < 0.01. (C) Tumor doubling time of the mice treated with ΔslyA, ΔSTM3120 and ΔhtrA compared with that with VNP20009. Date are shown in mean ± SEM, *P < 0.05, **P < 0.01. (D) FLAG expressions from the mutants carrying pHQ003-F-slyA, pHQ003-F-STM3120 and pHQ003-F-htrA was detected by western blotting. (E) Tumor volumes of the mice treated with ΔslyA-pHQ(slyA) compared with that treated with ΔslyA-pHQ. Date are shown in mean ± SEM, **P < 0.01. (F) Tumor volumes of the mice treated with ΔSTM3120-pHQ(STM3120) compared with that treated with ΔSTM3120-pHQ. Date are shown in mean ± SEM, **P < 0.01. (G) Tumor volumes of the mice treated with ΔhtrA-pHQ(htrA) compared with that treated with ΔhtrA-pHQ. Date are shown in mean ± SEM, **P < 0.01.
The analysis of the tumor immune-environment in mice
The anti-cancer ability of S. enterica relies on its ability to recruit immune cells into the tumor microenvironment [19]. Therefore, whether the mutants can promote proper immune responses in tumor microenvironment remains unclear. We analyzed CD4+ T cells, macrophages and granulocytes in tumor, which are reported to be responsible for the anticancer ability of S. enterica [19–21]. The results indicated that in contrast to the mutants, VNP20009 facilitated the migration of CD4+ T cells, macrophages and granulocytes into the tumor (Figure 4A, 4B, 4C). The ΔhtrA mutant, which completely lost its tumor suppressive capacity, induced 1.2% of CD4+ T cells on the 3rd day post infection. This level was only one fifth of that induced by VNP20009 (P < 0.05) (Figure 4A). The macrophages and granulocytes in the tumor of ΔhtrA group were decreased by more than 40% compared with that of VNP20009 group (P < 0.05) (Figure 4B, 4C). In addition, the ΔslyA and ΔSTM3120 treatments led a significant reduction of immune cells compared with the VNP20009 treatment (P < 0.05) (Figure 4A, 4B, 4C). Generally, the ability of the mutants to recruit immune cells into the tumor was reduced.
Previous studies showed that TNF-α and IL-1β are essential for bacterial cancer therapy. Thus, we further analyzed the expression of TNF-α and IL-1β in the tumors of bacteria-treated mice. The results showed that the mRNA levels of TNF-α and IL-1β in the mutant groups were significantly down-regulated compared with those in the VNP20009 group (P < 0.05) (Figure 4D, 4E). The expression of TNF-α in the VNP20009 group was increased 238-fold compared with that in the PBS control. However, the mice treated with ΔslyA, ΔSTM3120 and ΔhtrA mutants exhibited only 93-, 128- and 28- fold increase in TNF-α expression, respectively, compared with the mice treated with PBS (Figure 4D). Although IL-1β was significantly increased after bacterial therapy, the expression of IL-1β in the VNP20009 group was increased greater than 18- fold compared with the mutant groups (P < 0.05) (Figure 4E). These results indicated that the reduced anticancer activities of the mutants, which were deleted the genes slyA, STM3120 and htrA in the VNP20009 background, might be ascribed to the inability to activate immune-mediated antitumor responses.
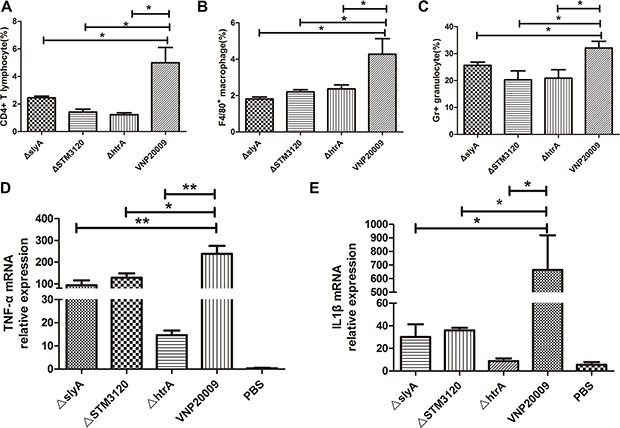
Figure 4: Subpopulation of immune cells and expression of TNF-α and IL-1β in the tumor. VNP20009 and its mutants (1 × 105 CFU/mouse) were injected intraperitoneally into the tumor-bearing mice. The immune cells in the tumor were determined on the third day post infection. (A) CD4+ T cells in the tumors of mice treated with ΔslyA, ΔSTM3120, ΔhtrA and VNP20009. Date are shown in mean ± SEM, *P < 0.05 for VNP20009 versus individual mutant. (B) Macrophages in the tumors of mice treated with ΔslyA, ΔSTM3120, ΔhtrA and VNP20009. Date are shown in mean ± SEM, *P < 0.05 for VNP20009 versus individual mutant. (C) Granulocytes in the tumors of mice treated with ΔslyA, ΔSTM3120, ΔhtrA and VNP20009. Date are shown in mean ± SEM, *P < 0.05 for VNP20009 versus individual mutant. (D) TNF-α mRNA expression levels in the tumors colonized by ΔslyA, ΔSTM3120, ΔhtrA and VNP20009. Date are shown in mean ± SEM, *P < 0.05, **P < 0.01 for VNP20009 versus individual mutant. (E) IL-1β mRNA expression levels in the tumors colonized by ΔslyA, ΔSTM3120, ΔhtrA and VNP20009. Date are shown in mean ± SEM, *P < 0.05 for VNP20009 versus individual mutant.
DISCUSSION
VNP20009 is a live anti-cancer agent and has some unique properties to make it better than traditional methods, including chemotherapy and radiation. For example, VNP20009 can specifically target tumors, sense the local environment and respond to external signals and can be engineered at genetic levels [1]. However, it failed in phase I clinical trials because of no antitumor effects [22]. Thus, tremendous efforts, including whole genome sequencing, were performed to improve the safety profile and tumor targeting ability [23]. However, the underlying mechanism of the anticancer capacity of VNP20009 still remains unclear. In this study, we knocked out the genes slyA, STM3120 and htrA, which are responsible for bacterial survival within macrophages, in the background of VNP20009 to study the relationship between bacterial intramacrophage survival ability and antitumor capacity. We found that all the mutants partially or completely lost their antitumor capacities, although they preferentially targeted and colonized tumor. The result indicates that in contrast to the former assumption, the bacterial burden in tumor does not play a decisive role in bacterial anticancer ability, which is consistent with previous observations [24, 25].
To establish a systematic infection, S. enterica must survive and replicate within the host macrophages where it is exposed to poor nutrients and microbicidal environments [26]. Production of reactive oxygen species (ROS) and nitric oxide is an important feature of intracellular bactericidal activities, which can be prevented by expressing slyA, STM3120 and htrA genes. HtrA possesses protease and chaperone activities, which are temperature dependent in vitro [27, 28]. The chaperone activity of HtrA predominates at low temperatures, whereas protease activity is present at high temperatures. In the present study, the growth rate of ΔhtrA in vitro was not impaired at 37°C, although the growth of ΔhtrA was reported to be inhibited at 46°C or higher [27]. In addition, Mutunga et al. found that ΔhtrA was attenuated because of increased sensitivity to oxidative stress [29]. Thus, ΔhtrA mutant was evaluated as a live vaccine and used for delivering recombinant antigens, such as CVD 908-htrA [30]. SlyA, a member of the MarR family of transcription factors, is a transcription factor that regulates a large number of genes, including S. enterica pathogenicity island (SPI)-2 [31, 32]. Because the genes within SPI-2 were reported to be involved in S. enterica virulence by preventing killing by mechanisms involving oxidative mechanism and because SPI-2 is required for S. enterica survival within macrophages, ΔslyA is sensitive to oxidative products of the respiratory burst [33, 34]. STM3120 is a predicted citE (citrate lyase) homolog and is important during systemic infection in BALB/c mice. Santiviago et al. suggested that STM3120 could decrease the level of nitric oxide in macrophages [35]. Consistent with these findings, our results revealed that the deletion of the genes slyA, STM3120 and htrA in the background of VNP20009 made the mutants more vulnerable in macrophages, although their growth rates in vitro and replications in B16F10 cells were not impaired.
The results showed that ΔslyA, ΔSTM3120 and ΔhtrA exhibited the highest tumor to spleen ratio, indicating that the tumor speci ficity of S. enterica was closely related to survival capacity within macrophages. When S. enterica enters the blood, the immune system is activated to deplete bacteria from the circulation system and macrophage is probably the most important. Given that S. enterica is rapidly subjected to phagocytosis by macrophages, only those bacteria capable of replicating within macrophages can cause a systematic infection. Thus, S. enterica that is incapable of replicating intracellularly is highly attenuated. In the present study, ΔslyA, ΔSTM3120 and ΔhtrA were efficiently cleared by macrophages, which could explain the low bacterial burden in spleen.
By contrast, S. enterica in tumors are protected from the immune system, and all the mutants accumulated in the tumor to the same extent as VNP20009. Although the process of bacterial tumor targeting is not entirely clear, macrophage was reported not to be a transporter of bacteria to tumor, and depleting macrophages did not affect tumor colonization [36]. Our results further confirmed this finding because the mutants with reduced survival within macrophages reached the same accumulation level in tumors as VNP20009. In addition, S. enterica was found both intracellular and extracellular in the tumor, where immune surveillance fails to function [36]. Three days post infection, when the bacterial titer reached the highest level in the tumor, all the mutants reached the same accumulation level in tumor as VNP20009 because the growth and replication rates of the mutants within melanoma cells were not impaired [37]. These results further reinforce that the low accumulation of mutant strains in spleen is due to impaired survival ability in macrophages. Although further investigations are necessary, the results showed that the bacterial tumor selectivity depends on macrophage-mediated clearance from the systemic organs and persistence of bacteria in the tumor microenvironment, which also partially explain Hoffman’s discovery, which showed mutants in STM3120 and htrA displayed reduced fitness in normal tissue and unchanged fitness in tumor [13].
Given that ΔslyA, ΔSTM3120 and ΔhtrA exhibited therapeutic potentials with reduced colonization in normal organs and unchanged colonization in tumors, they were thus validated for their anticancer activities. All of the mutants partially or completely lost their anticancer abilities. When the mutants were complemented with respective genes, their antitumor activities were partially or completely restored. In addition, the immune cells in the tumors of mice infected with the mutants were significantly decreased. Previous studies showed that the anti-tumor effects of S. enterica depend on the functional role of the MyD88-TLR pathway, and S. enterica enhances anti-tumor immunity by inhibiting tumor indoleamine 2, 3-dioxygenase 1 expression and inducing gap junctions in tumors [20, 38, 39]. Our results indicated the genes slyA, STM3120 and htrA, which are indispensable for the anticancer activity of VNP20009, are required for VNP20009 to induce the immune cells and exert an anti-tumor effect.
Furthermore, TNF-α and IL-1β are important for the tumor suppressive effects of S. enterica. Kim et al. found that the co-administration of recombinant TNF-α could prolong the tumor-suppressive effects of ΔppGpp Salmonella [25]. In our study, TNF-α was significantly decreased in the mice treated with the mutants. In addition, the mutants with impaired anticancer ability induced far less IL-1β than VNP20009. A recent study showed that the impaired anticancer ability of MG1655 (LPS-competent E.coli), which displayed similar tumor targeting capacity as S. enterica, is due to reduced IL-1β expression [25]. The result may also explain why the S. enterica mutants with unchanged tumor targeting ability failed to suppress tumor growth.
In conclusion, we created ΔslyA, ΔSTM3120 and ΔhtrA mutants that had similar tumor targeting abilities, but reduced anticancer capacities compared to VNP20009. We found that the survival ability within macrophages can increase the bacterial burden in normal organs without affecting the bacterial tumor targeting ability. In addition to this finding, immune cells and cytokines within tumors, rather than the bacterial burden in tumors, are responsible for the anticancer activity of VNP20009. The current study is the first to show the relationship between macrophage intracellular survival and anticancer activity and thus provides novel insights into bacterial cancer therapy.
MATERIALS AND METHODS
Bacterial strains, cell lines, and mice
VNP20009 is a purine auxotrophy, which deletes msbB from S. enterica ATCC 14028S [8]. All the mutants, namely ΔslyA, ΔSTM3120 and ΔhtrA with a deletion of slyA, STM3120 or htrA in VNP20009 were gifts from Prof. Shi’s lab. All the strains of S. enterica were cultured in Luria Broth (LB) or on agar plates using standard procedures. B16F10 melanoma cells were obtained from American Type Culture Collection (ATCC, USA) and were cultured in 5% CO2 in a humidified atmosphere in Dulbecco’s modified Eagle’s media (DMEM) supplemented with 10% fetal bovine serum (FBS). Six-week-old female C57BL/6 mice were purchased from the Comparative Medicine Center of Yangzhou University and maintained in pathogen-free conditions for one week before treatments.
Quantification of S. enterica in tumor
Briefly, 1 × 105 B16F10 cells were subcutaneously inoculated into the mid-right flank of C57BL/6 mice. When the tumor size reached 150 mm3, mice were randomly divided into VNP20009 group, ΔslyA group, ΔSTM3120 group and ΔhtrA group, with each group containing of at least 4 mice. S. enterica was grown overnight in LB and was diluted at a ratio of 1:100 into fresh LB. When optical density (OD) at 600nm was between 0.7 and 1.0, S. enterica with 1 × 108 CFU were obtained through centrifuging and suspended in phosphate buffered saline (PBS). S. enterica with 1 × 105 CFU were obtained by a serial dilution and intraperitoneally injected into tumor-bearing mice. The mice were sacrificed 3 days after S. enterica injection. Spleen and tumor were homogenized in 2 ml of PBS to release S. enterica and then plated on LB agar by a serial dilution. After 16 h of culture at 37°C, the titer of S. enterica was determined by counting colonies and dividing them by the weight of the tissue (CFU/g) [40].
Anti-tumor assay in vivo
1 × 105 B16F10 cells were subcutaneously inoculated into the mid-right flank of C57BL/6 mice. When the tumor size reached 150 mm3, tumor bearing mice were randomly divided into groups, with each group containing 8 mice. For comparing the antitumor activities of the mutants and VNP20009, 1 × 105 CFU S. enterica were intraperitoneally inoculated into the tumor-bearing mice. For complementation assay, the mutants carrying corresponding plasmids and the mutants were intraperitoneally inoculated into the tumor-bearing mice, respectively. The length and width of the tumors were measured every two days by using a Vernier caliper (Mytutoyo Co., Japan) across two perpendicular diameters. Tumor volume was calculated using the following formula: tumor volume = length × width2 × 0.52.
Flow cytometry
As mentioned above, tumor bearing mice were randomly divided into 5 groups, with each group containing 4 mice. When the tumor size reached 150 mm3, 1 × 105 S. enterica were intraperitoneally inoculated into the tumor-bearing mice. The tumor-bearing mice were sacrificed on the third day. According to Bayne and Vonderheide’s work, tumors were collected post infection for subpopulation analysis of immune cells [41]. The tumor was digested by collagenase IV. A single cell suspension, which was obtained using a 40-μm cell falcon, was incubated in red blood cell lysis buffer for three to five minutes to exclude red blood cells. The cells were then washed twice in DMEM medium. To determine the percentage of immune cells in the tumor, 10,000 cells suspended in 100 μl PBS containing 10% fetal calf serum (FCS) were stained for 30 min at 4°C by using appropriate isotype controls and phycoerythrin-conjugated anti-CD4, anti-F4/80, FITC-conjugated anti-Gr (BD Biosciences). After washing in PBS, the cells were analyzed on a FACSCalibur using CELLQUEST software (BD Biosciences).
Bacterial invasion and amplification assay in vitro
The day before infection, 1 × 106 B16F10 or RAW264.7 cells were seeded into 12-well plate with 500 μl appropriate cell culture medium. The cells were incubated at 37°C in 5% CO2. The cell culture medium was replaced with fresh medium one hour prior to infection. 1 × 107 CFU bacteria were added into each well. After infection, the plates were centrifuged at 250 g for 5 minutes in order to bring cells and bacteria in contact and initiate infection. The plates were then incubated for one hour at 37°C in 5% CO2. Gentamicin (100 μg/ml) was added to the plates to kill extracellular bacteria one hour after infection. The gentamicin concentration was reduced to 10 μg/ml two hours later. At designated time points, cells were washed with PBS and lysed by 0.1% Triton X-100. Bacteria were released and plated onto LB agar plates by a serial dilution. CFUs on the plates were counted after overnight incubation at 37°C.
Real-time quantitative PCR assay
As mentioned above, tumor bearing mice were randomly divided into 5 groups, with each group containing 3 mice. Briefly, 1 × 105 S. enterica were intraperitoneally inoculated into the tumor-bearing mice. 7 days post infection, the mice were sacrificed and total RNA was isolated from tumor tissues with TRIzol reagent (Invitrogen). cDNA was synthesized using ReverTra Ace® qPCR RT Kit (Toyobo). Real-time quantitative PCR was performed using AceQ qPCR SYBR Green Master Mix (Vazyme) to determine TNF-α and IL-1β expression. β-actin was used as a normalizing control.
Construction of complementing plasmids
The genes slyA, STM3120 and htrA were amplified for complementation using the primer sets as listed in Table 1. The resulting PCR products were purified using agarose gel electrophoresis and extracted from the gel using the gel extraction kit. The purified PCR products were inserted into pHQ003-F, a derivative of pET28a vector by ClonExpress II One Step Cloning Kit (Vazyme). The expression of the gene is controlled by a constitutive J23100 promoter and the FLAG tag is attached to the C terminus of complement protein. E.coli DH5α was transformed with the complement plasmid, and the positive clones were selected on LB plates containing Kanamycin. Plasmids bearing the correct insert were electroporated into the mutants, namely ΔslyA-pHQ(slyA), ΔSTM3120-pHQ(STM3120) and ΔhtrA-pHQ(htrA), respectively. Plasmid-bearing mutants were selected on LB Kna plates.
Table 1: Primer sequence used to construct complementing plasmids
Name |
Sequence (5′–3′) |
|
|---|---|---|
slyA |
slyA-F |
gaggagaaatactagttggaatcgccactaggttc |
slyA-R |
tttgtagtcaagcttatcgtgagagtgcaattcca |
|
STM3120 |
STM3120-F |
gaggagaaatactagatgcccgatatttctcatac |
STM3120-R |
tttgtagtcaagcttggaaaagatccctgcccggg |
|
htrA |
htrA-F |
gaggagaaatactagatgaaaaaaaccacattagc |
htrA-R |
tttgtagtcaagcttctgcatcagcaaataaatag |
Western bolt analysis
ΔslyA-pHQ(slyA), ΔSTM3120-pHQ(STM3120) and ΔhtrA-pHQ(htrA) were grown overnight in LB broth containing Kanamycin. Bacterial pellets were boiled in SDS loading buffer for 15 min. Blots were probed using monoclonal mouse antibody (F1804) against FLAG (Sigma).
Statistical analysis
The results are presented as mean ± SEM using Graphpad software. Statistical significance for comparisons among two was analyzed using the student t test by SPSS. The level of significance was set at P < 0.05.
ACKNOWLEDGMENTS
The authors are grateful to grants from the Chinese National Nature Sciences Foundation (81573338), Doctoral Station Science Foundation from the Chinese Ministry of Education (20130091130003), the Ministry of Science and Technology (2012AA020809), the Jiangsu Provincial Nature Science Foundation (BE2013630), the Open Project Programs from State Key Laboratory of Natural Medicines of China Pharmaceutical University (G140014) and the Bureau of Science and Technology of Changzhou (CZ20130011, CE20135013).
CONFLICTS OF INTEREST
The authors disclose no conflicts.
REFERENCES
1. Forbes NS. Engineering the perfect (bacterial) cancer therapy. Nat Rev Cancer. 2010; 10:785–794.
2. Zhao M, Geller J, Ma H, Yang M, Penman S, Hoffman RM. Monotherapy with a tumor-targeting mutant of Salmonella typhimurium cures orthotopic metastatic mouse models of human prostate cancer. Proc Natl Acad Sci USA. 2007; 104:10170–10174.
3. Zhao M, Yang M, Ma H, Li X, Tan X, Li S, Yang Z, Hoffman RM. Targeted therapy with a Salmonella typhimurium leucine-arginine auxotroph cures orthotopic human breast tumors in nude mice. Cancer Res. 2006; 66:7647–7652.
4. Zhang Y, Miwa S, Zhang N, Hoffman RM, Zhao M. Tumor-targeting Salmonella typhimurium A1-R arrests growth of breast-cancer brain metastasis. Oncotarget. 2015; 6:2615–2622. doi: 10.18632/oncotarget.2811.
5. Matsumoto Y, Miwa S, Zhang Y, Zhao M, Yano S, Uehara F, Yamamoto M, Hiroshima Y, Toneri M, Bouvet M, Matsubara H, Tsuchiya H, Hoffman RM. Intraperitoneal administration of tumor-targeting Salmonella typhimurium A1-R inhibits disseminated human ovarian cancer and extends survival in nude mice. Oncotarget. 2015; 6:11369–11377. doi: 10.18632/oncotarget.3607.
6. Toneri M, Miwa S, Zhang Y, Hu C, Yano S, Matsumoto Y, Bouvet M, Nakanishi H, Hoffman RM, Zhao M. Tumor-targeting Salmonella typhimurium A1-R inhibits human prostate cancer experimental bone metastasis in mouse models. Oncotarget. 2015; 6:31335–31343. doi: 10.18632/oncotarget.5866.
7. Nagakura C, Hayashi K, Zhao M, Yamauchi K, Yamamoto N, Tsuchiya H, Tomita K, Bouvet M, Hoffman RM. Efficacy of a genetically-modified Salmonella typhimurium in an orthotopic human pancreatic cancer in nude mice. Anticancer Res. 2009; 29:1873–1878.
8. Low KB, Ittensohn M, Le T, Platt J, Sodi S, Amoss M, Ash O, Carmichael E, Chakraborty A, Fischer J, Lin SL, Luo X, Miller SI, et al. Lipid A mutant Salmonella with suppressed virulence and TNF alpha induction retain tumor-targeting in vivo. Nat Biotechnol. 1999; 17:37–41.
9. Blache CA, Manuel ER, Kaltcheva TI, Wong AN, Ellenhorn JD, Blazar BR, Diamond DJ. Systemic delivery of Salmonella typhimurium transformed with IDO shRNA enhances intratumoral vector colonization and suppresses tumor growth. Cancer Res. 2012; 72:6447–6456.
10. Cheng X, Zhang X, Cheng W, Chen J, Ma C, Yang B, Hua ZC. Tumor-specific delivery of histidine-rich glycoprotein suppresses tumor growth and metastasis by anti-angiogenesis and vessel normalization. Curr Gene Ther. 2014; 14:75–85.
11. Zhang X, Cheng X, Lai Y, Zhou Y, Cao W, Hua ZC. Salmonella VNP20009-mediated RNA interference of ABCB5 moderated chemoresistance of melanoma stem cell and suppressed tumor growth more potently. Oncotarget. 2016. 7:14940–14950. doi: 10.18632/oncotarget.7496.
12. Friedlos F, Lehouritis P, Ogilvie L, Hedley D, Davies L, Bermudes D, King I, Martin J, Marais R, Springer CJ. Attenuated Salmonella targets prodrug activating enzyme carboxypeptidase G2 to mouse melanoma and human breast and colon carcinomas for effective suicide gene therapy. Clin Cancer Res. 2008; 14:4259–4266.
13. Arrach N, Cheng P, Zhao M, Santiviago CA, Hoffman RM, McClelland M. High-throughput screening for salmonella avirulent mutants that retain targeting of solid tumors. Cancer Res. 2010; 70:2165–2170.
14. Arrach N, Zhao M, Porwollik S, Hoffman RM, McClelland M. Salmonella promoters preferentially activated inside tumors. Cancer Res. 2008; 68:4827–4832.
15. Kasinskas RW, Forbes NS. Salmonella typhimurium lacking ribose chemoreceptors localize in tumor quiescence and induce apoptosis. Cancer Res. 2007; 67:3201–3209.
16. Pawelek JM, Sodi S, Chakraborty AK, Platt JT, Miller S, Holden DW, Hensel M, Low KB. Salmonella pathogenicity island-2 and anticancer activity in mice. Cancer Gene Ther. 2002; 9:813–818.
17. Linehan SA, Holden DW. The interplay between Salmonella typhimurium and its macrophage host—what can it teach us about innate immunity? Immunology letters. 2003; 85:183–192.
18. Fields PI, Swanson RV, Haidaris CG, Heffron F. Mutants of Salmonella typhimurium that cannot survive within the macrophage are avirulent. Proc Natl Acad Sci USA. 1986; 83:5189–5193.
19. Avogadri F, Martinoli C, Petrovska L, Chiodoni C, Transidico P, Bronte V, Longhi R, Colombo MP, Dougan G, Rescigno M. Cancer immunotherapy based on killing of Salmonella-infected tumor cells. Cancer Res. 2005; 65:3920–3927.
20. Kaimala S, Mohamed YA, Nader N, Issac J, Elkord E, Chouaib S, Fernandez-Cabezudo MJ, Al-Ramadi BK. Salmonella-mediated tumor regression involves targeting of tumor myeloid suppressor cells causing a shift to M1-like phenotype and reduction in suppressive capacity. Cancer Immunol Immunother. 2014; 63:587–599.
21. Vendrell A, Gravisaco MJ, Pasetti MF, Croci M, Colombo L, Rodriguez C, Mongini C, Waldner CI. A novel Salmonella Typhi-based immunotherapy promotes tumor killing via an antitumor Th1-type cellular immune response and neutrophil activation in a mouse model of breast cancer. Vaccine. 2011; 29:728–736.
22. Toso JF, Gill VJ, Hwu P, Marincola FM, Restifo NP, Schwartzentruber DJ, Sherry RM, Topalian SL, Yang JC, Stock F. Phase I study of the intravenous administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J Clin Oncol. 2002; 20:142–152.
23. Broadway KM, Modise T, Jensen RV, Scharf BE. Complete genome sequence of Salmonella enterica serovar Typhimurium VNP20009, a strain engineered for tumor targeting. J Biotechnol. 2014; 192:177–178.
24. Frahm M, Felgner S, Kocijancic D, Rohde M, Hensel M, Curtiss R, 3rd, Erhardt M, Weiss S. Efficiency of conditionally attenuated Salmonella enterica serovar Typhimurium in bacterium-mediated tumor therapy. MBio. 2015; 6.
25. Kim JE, Phan TX, Nguyen VH, Dinh-Vu HV, Zheng JH, Yun M, Park SG, Hong Y, Choy HE, Szardenings M, Hwang W, Park JA, Park S, Im SH, Min JJ. Salmonella typhimurium Suppresses Tumor Growth via the Pro-Inflammatory Cytokine Interleukin-1beta. Theranostics. 2015; 5:1328–1342.
26. Ohl ME, Miller SI. Salmonella: a model for bacterial pathogenesis. Annu Rev Med. 2001; 52:259–274.
27. Lewis C, Skovierova H, Rowley G, Rezuchova B, Homerova D, Stevenson A, Spencer J, Farn J, Kormanec J, Roberts M. Salmonella enterica Serovar Typhimurium HtrA: regulation of expression and role of the chaperone and protease activities during infection. Microbiology. 2009; 155:873–881.
28. Spiess C, Beil A, Ehrmann M. A temperature-dependent switch from chaperone to protease in a widely conserved heat shock protein. Cell. 1999; 97:339–347.
29. Mutunga M, Graham S, De Hormaeche RD, Musson JA, Robinson JH, Mastroeni P, Khan CM, Hormaeche CE. Attenuated Salmonella typhimurium htrA mutants cause fatal infections in mice deficient in NADPH oxidase and destroy NADPH oxidase-deficient macrophage monolayers. Vaccine. 2004; 22:4124–4131.
30. Tacket CO, Levine MM. CVD 908, CVD 908-htrA, and CVD 909 live oral typhoid vaccines: a logical progression. Clin Infect Dis. 2007; 45:S20–23.
31. Linehan SA, Rytkonen A, Yu XJ, Liu M, Holden DW. SlyA regulates function of Salmonella pathogenicity island 2 (SPI-2) and expression of SPI-2-associated genes. Infect Immun. 2005; 73:4354–4362.
32. Stapleton MR, Norte VA, Read RC, Green J. Interaction of the Salmonella typhimurium transcription and virulence factor SlyA with target DNA and identification of members of the SlyA regulon. J Biol Chem. 2002; 277:17630–17637.
33. Vazquez-Torres A, Xu Y, Jones-Carson J, Holden DW, Lucia SM, Dinauer MC, Mastroeni P, Fang FC. Salmonella pathogenicity island 2-dependent evasion of the phagocyte NADPH oxidase. Science. 2000; 287:1655–1658.
34. Buchmeier N, Bossie S, Chen CY, Fang FC, Guiney DG, Libby SJ. SlyA, a transcriptional regulator of Salmonella typhimurium, is required for resistance to oxidative stress and is expressed in the intracellular environment of macrophages. Infect Immun. 1997; 65:3725–3730.
35. Santiviago CA, Reynolds MM, Porwollik S, Choi SH, Long F, Andrews-Polymenis HL, McClelland M. Analysis of pools of targeted Salmonella deletion mutants identifies novel genes affecting fitness during competitive infection in mice. PLoS Pathog. 2009; 5:e1000477.
36. Stritzker J, Weibel S, Seubert C, Gotz A, Tresch A, van Rooijen N, Oelschlaeger TA, Hill PJ, Gentschev I, Szalay AA. Enterobacterial tumor colonization in mice depends on bacterial metabolism and macrophages but is independent of chemotaxis and motility. Int J Med Microbiol. 2010; 300:449–456.
37. Crull K, Bumann D, Weiss S. Influence of infection route and virulence factors on colonization of solid tumors by Salmonella enterica serovar Typhimurium. FEMS Immunol Med Microbiol. 2011; 62:75–83.
38. Saccheri F, Pozzi C, Avogadri F, Barozzi S, Faretta M, Fusi P, Rescigno M. Bacteria-induced gap junctions in tumors favor antigen cross-presentation and antitumor immunity. Sci Transl Med. 2010; 2:44ra57.
39. Kuan YD, Lee CH. Salmonella overcomes tumor immune tolerance by inhibition of tumor indoleamine 2, 3-dioxygenase 1 expression. Oncotarget. 2016; 7:374–385. doi: 10.18632/oncotarget.6258.
40. Cheng X, Zhang X, Zhou Y, Zhang C, Hua ZC. A Salmonella Typhimurium mutant strain capable of RNAi delivery: higher tumor-targeting and lower toxicity. Cancer biology & therapy. 2014; 15:1068–1076.
41. Bayne LJ, Vonderheide RH. Multicolor flow cytometric analysis of immune cell subsets in tumor-bearing mice. Cold Spring Harb Protoc. 2013; 2013:955–960.