
INTRODUCTION
Ovarian cancer is the leading cause of deaths among female gynecological malignancies. Due to the indolence at the early stage and lack of early diagnostic methods, approximately 70% of patients are diagnosed at advanced stage (stage III and IV) [1–2]. Cytoreductive surgery, followed by systematic chemotherapy of platinum and paclitaxel, is adapted as the standard treatment for advanced epithelial ovarian cancer. Even though the treatment is initially effective to 80% patients, recurrent cancer with paclitaxel resistance is inevitable in many cases, resulting in subsequent failure of chemotherapy [3–4]. Moreover, available evidence indicates that the effect of second-line chemotherapy is compromised due to heterogeneity in populations [5]. Therefore, exploring new anticancer drugs and reversing drug resistance are particularly important.
Natural somatostatin (SST), also known as somatotropin release-inhibiting factor (SRIF), is a cyclic peptide, with a wide range of cellular functions, such as inhibiting the secretion of various hormones, blocking cell proliferation and promoting apoptosis. As one of the short peptides, somatostatin is an optional agent in receptor targeting radiotherapy and chemotherapy. This is based on its ease of synthesis and optimization, which relatively small molecular weight, fast cycling clearance, and good tissue penetration ability [6–8]. The biological effects of somatostatin are mediated by targeting the five somatostatin receptors (SSTR) expressed on cell membranes [9–10]. Somatostatin receptors, which are a group of mediators for somatostatin function, are distributed extensively in neuroendocrine tumors as well as carcinomas such as breast cancer, colon cancer, non-small cell lung cancer, ovarian cancer and cervical cancer [11–12]. These observations raise the possibility of SST and its analogue as potential tumor therapeutic molecules. Compared to natural somatostatin, synthetic somatostatin analogues (SSTA) like lanreotide, octreotide and vapreotide show more advantages including longer half-life, metabolic stability, more specificity and efficiency [13–15]. Studies have shown that SSTAs not only inhibit tumor cell proliferation in vitro, but also suppress the growth of solid tumors in vivo.
One of the five somatostatin receptors, SSTR2, can be detected to be expressed in tumor, and SSTA has much higher binding affinity to SSTR2 in tumor tissues than in normal tissues [16–17]. SSTR2 can mediate the biological effects of somatostatin as the latter can block the cell proliferation, promote apoptosis and inhibit the secretion of various hormones [9–10]. Furthermore, recent studies have found that the expression of SSTR2 in ovarian cancer tissue is quite high and SSTA has the capacity to inhibit and reverse paclitaxel resistance from multiple pathways [18–24]. Some researchers conjugated SSTA (RC121, RC160) with paclitaxel into different targeted drugs and found that the conjugate increased intracellular concentration of paclitaxel, which was cytotoxic to breast cancer cells and its stem cells [25].
Octreotide (OCT) is the most widely applied representative of SSTA, and can inhibit the growth of various carcinomas including gastrointestinal tumors, breast cancer, besides neuroendocrine tumors and leukemia, through targeted binding to somatostatin receptor on tumor cells [26–28]. OCT also displays a high affinity for SSTR2, induces the activation of tyrosine phosphatase and inhibits the proliferation of SSTR2 expressing cells [29]. Our previous studies confirmed that OCT, in combination with cisplatin, can effectively inhibit the growth of SKOV3/DDP cisplatin-resistant human ovarian cancer, promote its apoptosis and reverse its resistance to cisplatin [30–31]. We have also detected that A2780/Taxol paclitaxel-resistant human ovarian cancer cell expresses SSTR2 and therefore, the synthetic POC could exert its functions in a dose and time-dependent manner via its specific binding to cell surface [32].
Some other studies indicate that chemotherapy based on SSTR-targeting has higher efficiency and lower toxicity in comparison to traditional chemotherapy. Some scholars conjugated doxorubicin or gemcitabine with SSTA and discovered that the conjugates had cytotoxic effect specifically on tumor tissues which express SSTR but lower toxicity to normal tissues in animal models. Furthermore, even in tumors with relatively low densities of SSTR expression, the conjugates are more effective and less toxic [33–35].
Based on the previous findings, this study aims to investigate the targeted effectiveness of POC and its mechanism on reversing the resistance of paclitaxel in A2780/Taxol ovarian cancer xenografted in the animal model.
RESULTS
Inhibition of tumor growth
All the xenografted tumors grew up to about 1cm3 in size after 10 days following subcutaneous injection of A2780/Taxol ovarian cancer cells. At this time, no symptoms of apathy, diarrhea or poor vitality was observed in all the tested mice. After four weeks following the first administration, the average tumor weight and size in any treatment group were significantly lower than those in the control group (P <0.05). In comparison with the octreotide group and paclitaxel group, the average weight and size of tumor in PO group (0.86 gm, 1.12 cm3) were even lower and smaller (P<0.05). In addition, POC presented the perfect efficacy in tumor growth inhibition with a significantly decreased average weight and size of tumor (0.59 gm, 0.64 cm3) than any other treatments (P<0.05; Table 1, Figure 1).
Table 1: Average weight and size of tumors in different groups
Groups |
weight(g) |
size(cm3) |
inhibition rate(%) |
|---|---|---|---|
Control |
1.70±0.24 |
3.40±0.26 |
— |
Octreotidea |
1.38±0.27 |
2.08±0.19 |
39% |
Paclitaxela,b |
1.11±0.26 |
1.61±0.18 |
53% |
POa,b,c |
0.86±0.26 |
1.12±0.23 |
67% |
POCa,b,c,d |
0.59±0.18 |
0.64±0.24 |
81% |
aP<0.05 vs Control; bP<0.05 vs Octreotide group; cP<0.05 vs Paclitaxel group; P<0.05 vs PO group.
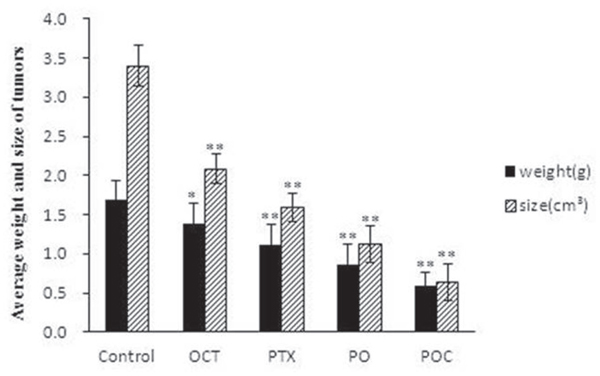
Figure 1: Average weight and size of xenografted tumors. Mice were treated with paclitaxel (PTX), octreotide (OCT), paclitaxel combined with octreotide (PO), paclitaxel-octreotide conjugate (POC) or saline only at the indicated concentration. Compared to the other treatment, POC exhibited an enhanced inhibitory effect on tumor growth (P<0.05) (* P< 0.05 vs Control; ** P< 0.01 vs Control).
Targeting ability of POC in vivo
When the xenografted tumor grew up to about 1cm3, intravenous injection of fluorescence labeled paclitaxel-octreotide conjugate (FITC-POC) was performed in selected nude mice. 30 minutes after the administration, the fluorescent signals were initially observed in tumor site, and the fluorescence intensity was gradually increased during the prolonged observation (Figure 2A). A significant aggregation of fluorescence into the xenografted tumor at 8 hours reaching the peak at 24 hours indicated a favorable targeting of POC. After administration for 72hours, part of the fluorescence was still visible within the tumor site. Besides, mild fluorescent signals could be found aggregated in liver and kidney at each time point. These findings suggested that POC presented specific targeting to paclitaxel-resistant human ovarian cancer cells in nude mice and was probably metabolized by the liver and excreted by the kidneys.
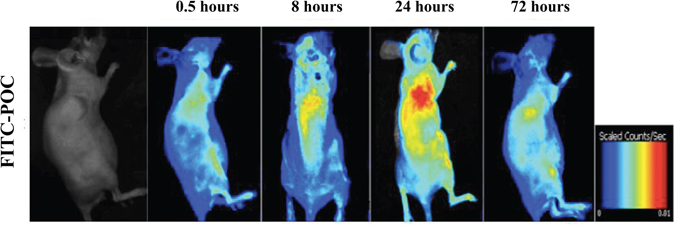
Figure 2: Analysis of specific binding of POC in xenografted tumor. The targeting ability analysis of POC was performed by administrating of fluorescein-labeled paclitaxel-octreotide conjugate (FITC-POC). A significant aggregation of fluorescence into the xenografted tumor was observed at 8 hours with reaching the peak at 24 hours. After administration for 72 hours, fluorescence was still visible in tumor site.
Histology and SSTR2 expression in xenografted tumor
The tumors showed the same morphology as the original serous ovarian cancer by using HE stain. POC group and PO group displayed more necrosis of tumor tissues than paclitaxel group and octreotide group (Figure 3A). In addition, these morphology effects of chemotherapy were not observed in the non-targeted organs, such as liver, kidney, spleen and the tails of the treated mice (Supplemental figure 2). Therefore, it is suggested POC had lower or no toxicity to tissues outside of xenografts. SSTR2 expression in tumor tissues was detected by IHC using the monoclonal antibody against SSTR2. SSTR2 showed cell membrane expression in our study, as described in previous study (Figure 3B).
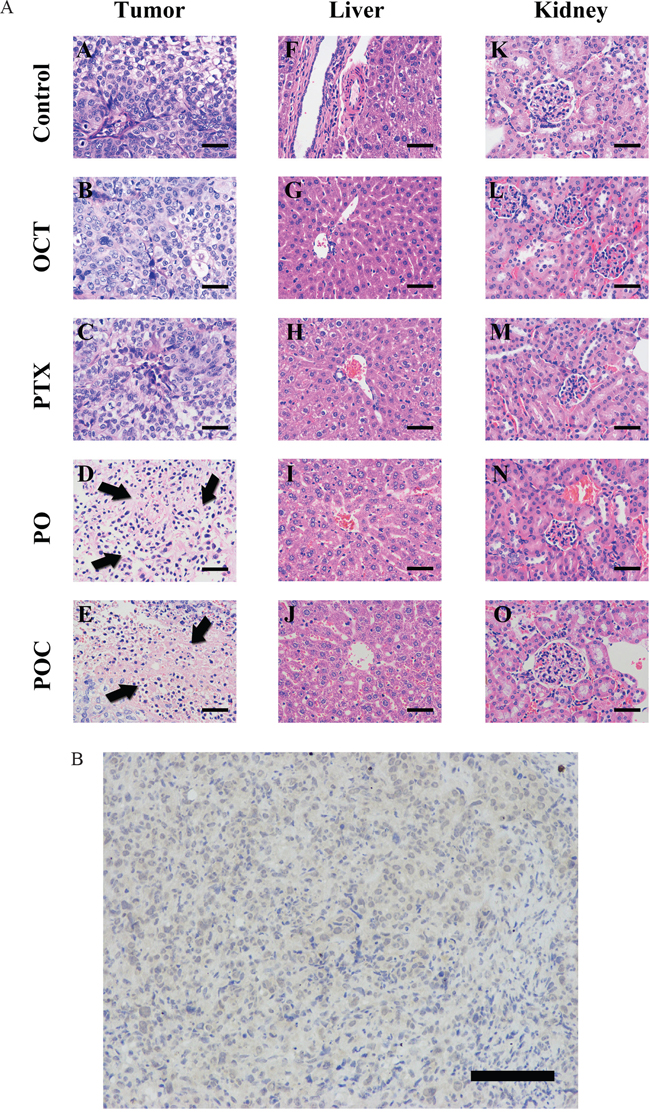
Figure 3: A. Histolog of tumor, liver, kidney by HE staining. Pathological examination was performed by HE staining. Various degrees of necrosis, inflammatory cell infiltration and fibrosis, as well as cell apoptosis, evidenced by karyorrhexis and pyknoses were observed in PO group and POC group (A, B, C, D and E; arrowheads). Tissue injury of liver (F, G, H, I and J) and kidney (K, L, M, N and O) tissues were not observed in all treated mice (magnification ×400, scale bar =200μm). B. Expression of SSTR2 in xenografted tumor. Immunohistochemistry with sections of xenografted tumor was detected under high power microscopic view. Brown granular particles were observed gathering on cell membrane surface and surrounding the cytoplasm, indicating SSTR2 expression of xenografted tumor was positive (magnification ×200, scale bar =400μm).
Expression of SSTR2, MDR1, VEGF, MMP-9, Tubα1a and Tubβ3 mRNA
To investigate the resistance reversal mechanisms of POC, real-time PCR was performed to measure the mRNA expression levels in each treatment group. SSTR2 mRNA expression was detected in tumor tissues in all experimental groups and was higher in each treatment group than in the control group. Moreover, SSTR2 mRNA expression level in POC group was significantly higher than in the other treatment groups (P<0.05, Figure 4). However, the mRNA expression levels of MDR1, VEGF, MMP-9, Tubα1a and Tubβ3 in treatment groups were lower than those in the control group, and POC group has a significant lower expression than the other treatment groups (P<0.05). In addition, Tubα1a and Tubβ3 mRNA expression in octreotide group was higher than in paclitaxel group, but MDR1, VEGF, MMP-9 mRNA expression was lower than in paclitaxel group. The differences between each group were statistically significant (P <0.05).
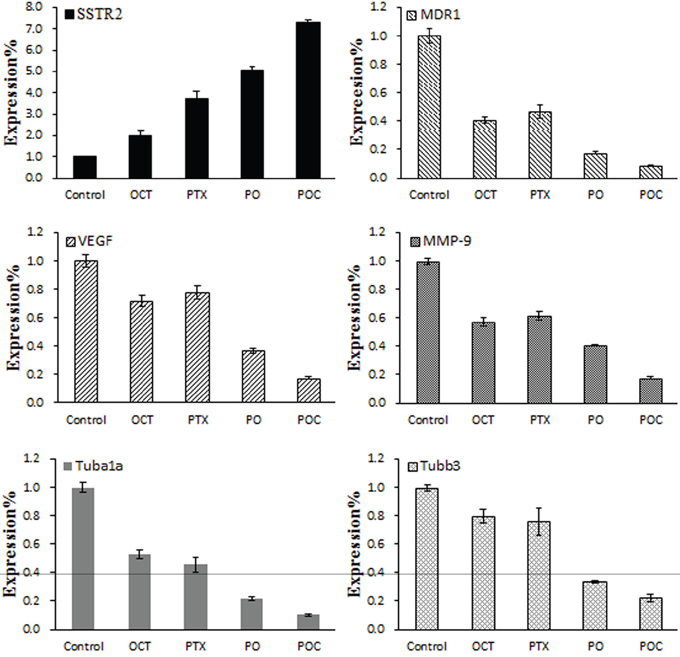
Figure 4: Effect of POC on the mRNA expression of SSTR2, MDR1, VEGF, MMP-9, Tubα1a and Tubβ3 in tumor tissues. SSTR2, MDR1, VEGF, MMP-9, Tubα1a and Tubβ3 mRNA expressions were determined following each treatment by real-time PCR. SSTR2 mRNA expression was higher, but MDR1, VEGF, MMP-9, Tubα1a and Tubβ3 mRNA expression were lower in POC group than in the other treatment groups (P<0.05).
Expression of SSTR2, MDR1, VEGF, MMP-9, α-tubulin and βIII-tubulin protein
Western blotting was applied to determine the protein expression levels of SSTR2, MDR1, VEGF, MMP-9, α-tubulin and βIII-tubulin after each treatment. The protein level of SSTR2 was in accordance with the mRNA expression. Namely, SSTR2 protein expressions in all treatment groups were higher than in the control group and POC group was significantly higher than the other treatment groups. Furthermore, MDR1, VEGF, MMP-9, α-tubulin and βIII-tubulin protein expressions in each treatment group were lower in comparison with those in the control group, and levels of POC group were significantly lower than those from the other treatment groups (P <0.05). It is worth noting that, the protein expression levels of MDR1, VEGF, MMP-9 in octreotide group were lower than those in paclitaxel group, and α-tubulin, βIII-tubulin were still higher than paclitaxel group. The differences between each group were statistically significant (P <0.05, Figure 5).
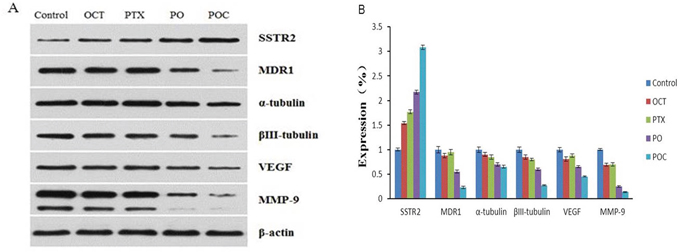
Figure 5: Effect of POC on the protein expression of SSTR2, MDR1, VEGF, MMP-9, α-tubulin and βIII-tubulin in tumor tissues. SSTR2, MDR1, VEGF, MMP-9, α-tubulin and βIII-tubulin protein expressions by Western blot indicated that SSTR2 protein increased but MDR1, VEGF, MMP-9, α-tubulin and βIII-tubulin proteins decreased following treatment with POC (P<0.05) A,B.
DISCUSSION
Chemotherapeutic regimens based on paclitaxel and platinum combination are currently used for the treatment of patients with advanced ovarian cancer. However, due to the emergence of drug resistance, tumor relapse happens even with chemotherapy and usually leads to treatment failure. More investigations thus emphasize on the paclitaxel resistance and the development of new therapeutic agents that can overcome this resistance.
Peptide receptors are potential targets for the molecular therapies, as represented by somatostatin receptor [36]. The synthetic somatostatin analogue, octreotide, has a longer half-life than native somatostatin, and is effective in inhibition of multiple tumors such as neuroendocrine tumors, gastrointestinal cancer, breast cancer and leukemia [26–28].
Our previous study in vitro using the human ovarian cancer cell line found that paclitaxel-resistant cancer cells express high levels of SSTR2, and POC treatment could lead to more cell apoptosis when compared to cells treated with paclitaxel alone. Underlying cause for the difference is possibly the specific targeting of octreotide to SSTR2, which is distributed at high density on tumor cell surface [32]. Here we reported the first in vivo study using the nude mice. As in previous study, the high level expression of SSTR2 in xenografted paclitaxel-resistant ovarian tumor was confirmed by IHC, RT-PCR and western blot.
Previous in vitro research found that POC reversed chemotherapy paclitaxel-resistance through reducing the protein levels of SSTR2. However, the in vivo results represented in this article suggested that SSTR2 expression in POC treatment was significantly higher than in other treatment at both mRNA and protein levels. These observations may indicate that the reversal effect of POC might be associated with the increased the expression of SSTR2. The discrepancy of SSTR2 observed in Hua group between short-term and long-term treatment of Hepatocellular carcinoma using octreotide can explain the different SSTR2 expression levels in our in vivo and in vitro studies. This discrepancy could have resulted from the desensitization and internalization of SSTR2 after short-term octreotide application, and may be related to drug concentration, treatment time and microenvironment [37].
In vivo imaging results further showed that POC could combine with xenografted tumor specifically and POC was accumulated in tumor significantly during the prolonged observation in 24 hours. However, other organs such as the liver and the kidneys have no obvious uptake of POC. The inhomogeneous POC distribution indicated that POC can specifically target at paclitaxel-resistant human ovarian cancer cells in nude mice. In addition, the tumor weight and size of POC treatment were significantly lower than those in the other treatment group. Histologic examination also revealed that the degree of tumor necrosis and chemotherapy morphologic changes was more significant in POC group. A large lightly stained area with unclear boundary was surrounded by different amount of inflammatory cell infiltration and irregularly divided tumor cells. However, other organs showed no remarkable pathological changes. In line with our previous studies, these results further illustrated that POC could not only inhibit the proliferation of tumor cells in vitro, but also reverse the resistance of human ovarian cancer cell to paclitaxel and induce the better chemotherapy response of solid tumors in vivo. These therapeutic effects were even further optimized as reduced cytotoxicity of POC to SSTR-negative cells was observed simultaneously.
Multidrug resistance gene 1 (MDR1) is known as one of the important mechanisms causing paclitaxel resistance. Overexpression of MDR1 leads to membrane p-glycoprotein (p-gp) overexpression, which could decrease the intracellular paclitaxel concentration, and results in the reduced inhibitory effect on tumor cells by paclitaxel [31–42]. Studies have shown that, SSTA can suppress MDR1 expression and reverse drug resistance by inhibiting the PI3K/Akt signal pathway when binding to SSTR1A, SSTRR1B and SSTR2 [21]. In our study, POC treatment significantly decreased the mRNA and protein expressions of MDR1 if compared with other treatment group which was consistent with our previous study in vitro. Furthermore, the MDR1 expression was lower in mRNA and protein levels by octreotide treatment than paclitaxel treatment. In combination, these results suggested that the reversal mechanism of POC to paclitaxel-resistance in human ovarian cancer was possibly by reducing MDR1 expression.
Another mechanism of paclitaxel resistance may be related to abnormal expression of vascular endothelial growth factor (VEGFA, VEGF), a growth factor promoting angiogenesis and playing the critical role in tumor expansion [43]. The VEGF released by tumor tissues leads to an upregulation of MDR1 through VEGF-VEGFR signal pathway, and VEGFR antagonist can significantly inhibit the ability of drug resistance and invasion of tumors [44, 45]. Studies have demonstrated that the microvascular endothelial cells of tumor tissues sometimes express SSTR and somatostatin. These expressions could induce vasoconstriction and consequently cause circulatory disturbance to inhibit tumor growth [22–23]. This study showed that both mRNA and protein expression of VEGF after POC treatment were down-regulated, which was consistent with the result in vitro [32]. In addition, octreotide treatment led to a lower mRNA and protein expressions of VEGF in comparison to paclitaxel treatment. The down-regulating of VEGF expression was probably the reversal mechanisms mediated by POC.
Matrix metalloproteinases (MMPs) are demanding molecules in tumor cell invasion by degrading extracellular matrix (ECM). MMP-2 and -9 are particularly associated with the invasion of multiple gynecological maliginancies, such as cervical cancer, endometrial cancer and ovarian cancer [46]. Bergers found that MMP-9 could promote tumor angiogenesis by increasing the expression of VEGF and its receptor (VEGFR) [47]. When angiogenesis was inhibited, MMP-2 and -9 expressions are up-regulated. Inhibitors of MMPs can weaken the MMPs roles in regulation of angiogenesis by decreasing MMP-9 expression, and consequently the inhibition of tumor growth. Some other studies also revealed that up-regulation of MMP-9 expression enhanced the chemotherapeutic resistance, and SSTR2 could suppress tumor growth and metastasis by inhibition of MMP-2 and -9 expressions indirectly [24, 48]. Here we found that MMP-9 mRNA and protein expressions in tumor tissues were significantly decreased following POC treatment as compared to the other treatment and control group. Interestingly, the mRNA and protein levels of MMP-9 by octreotide treatment alone were both lower than by paclitaxel treatment alone. In summary, these findings suggested that POC reversal of the resistance of paclitaxel probably by inhibiting MMP-9 expressions in both mRNA and protein levels, and it seems that octreotide maybe more relevant to regulate the MMPs.
The anti-tumor activity of paclitaxel relies on its capability of stabilizing microtubules and the resultant interference with the G2-M transition of the cell cycle [49–50]. Thus, the altered structure of microtubules is one of the reasons for paclitaxel resistance. Fluctuation of microtubule composition expression may lead to dynamic changes in microtubule and eventually alter the tumor response to paclitaxel. The increased expression levels of α-tubulin and βIII-tubulin, mutation of βI-tubulin or down-regulation of microtubule associated proteins (MAPs) can undermine the inherent stability of microtubules and reduce the efficacy of paclitaxel [51–52]. In this study, decreased mRNA and protein expressions of α-tubulin and βIII-tubulin in tumor tissues were detected in each treatment group except the control group. Octreotide treatment showed higher α-tubulin and βIII-tubulin expression in both mRNA and protein levels than paclitaxel treatment, which was higher than octreotide combined with paclitaxel treatment. Additionally, POC treatment had the lowest expressions of α-tubulin and βIII-tubulin in all experimental groups. The alteration of tubulin expression levels may be explained by POC increasing the intracellular concentration of paclitaxel through specifically targeting octreotide on cell membrane followed by endocytosis. The endocytosis of the conjugate then concentrates the intracellular paclitaxel and promotes its inhibitory effect on α-tubulin and βIII-tubulin.
Paclitaxel-octreotide conjugate presents better effects on targeting and inhibiting paclitaxel-resistant human ovarian cancer cells in nude mice. The inhibition of POC on tumors could probably result from a series of mechanisms, including specific binding of octreotide to the membrance receptor of SSTR2, up-regulated expression of SSTR2 as well as down-regulated expressions of MDR1, α-tubulin, βIII-tubulin, VEGF and MMP-9.
MATERIALS AND METHODS
Peptide and chemicals
Paclitaxel (Taiji Pharmaceutical Co.Ltd., China) – octreotide (Zineng Pharmaceutical Co.Ltd., China) conjugate was synthesized in our lab, using the following steps: (1) Preparation of SDPP (N-Hydroxysuccinimido diphenyl phosphate) and paclitaxel 2’ -succinate. (2) Synthesis of paclitaxel 2’-succinyl-NHS by the reaction of paclitaxel 2’-succinate with SDPP. (3) Paclitaxel - octreotide conjugate was obtained by the coupling reaction of paclitaxel 2’-succinyl-NHS with octreotide. The structure of paclitaxel - octreotide conjugate was confirmed by ESI-MS (as shown in supplemental figure 1). Fluorescein isothiocyanate (FITC) (Sigma Chemical Co.St.Louis, MO, USA) was labeled to paclitaxel - octreotide conjugate by our laboratory.
Cell culture
Paclitaxel-resistant human ovarian cancer cell line A2780/Taxol (BogooBioTECH, Co.Ltd., China) was cultured in RPMI1640 medium (Gibco BRL Co.Ltd., USA) with 10% fetal bovine serum (FBS) (Invitrogen, USA) at 37°C in 5% CO2 atmosphere and 90% humidity. The log-phase cells were collected by using 0.25% trypsin (Gibco BRL Co.Ltd., USA) and then xenografted into nude mice simultaneously.
Animals and xenografts
Female athymic nude mice (BALB/c-nu/nu) (Experimental Animal Center of Chinese Academy of Science), approximately 6-8 weeks old and weighing 18-20g were breeding in specific-pathogen-free (SPF) conditions. Xenograft models were established by subcutaneous injection of 5×106 A2780/Taxol ovarian cancer cells into the right armpit of forty nude mice, and mice behavior and tumor growth were observed daily. All animal experiments were approved by Experimental Animal Ethics Committee of Southeast University.
Growth inhibition of xenografted tumor
When the size of xenografted tumor grow up to 1cm3, forty healthy nude mice were chosen and averagely divided into five groups: The paclitaxel (150 nmol/kg), octreotide (150 nmol/kg), paclitaxel combined with octreotide (150 nmol/kg), paclitaxel-octreotide conjugate (150 nmol/kg) or saline only were intravenously administrated on days 1,8 and 15. Four weeks after the first administration, the mice were sacrificed to measure the size of tumors according to Steel formula tumor volume: V = ab2/2 (cm3) (a: short-track; b: long-track). The tumors were weighed in grams after stripping out the tumors, removing adjacent non-tumor soft tissue. Then the inhibitory rate of each drugs were calculated as follows: inhibitory rate = (Vcontrol group -Vexperimental group)/Vcontrol group × 100 %.
Targeting ability studies
To evaluate the targeting ability of POC, we used nude mice with the tumor size larger than 1cm3 for the imaging experiments. The mice were intravenously injected with fluorescein-labeled paclitaxel-octreotide conjugate (FITC-POC) (150 nmol/kg). After being anesthetized with isoflurane (2-3% concentration), all mice were observed by using the automated small-animal in vivo imaging systems (Maestro™ Automated In-Vivo Imaging, USA) at multiple time points (0.5 and 8, 24 and 72hours).
Histopathology and immunohistochemistry assay
To perform pathological examination, the tumor tissue of mice in each treated group was fixed with 4% formaldehyde solution, dehydrated, embedded in paraffin and cut into 5μm-thick serial sections. Selected sections were then stained with hematoxylin–eosin staining (HE stain). Morphological changes of tumor tissues and non-targeted organs were observed under light microscope and confocal microscope (Olympus FluoView ™ FV1000, Japan). The primary rabbit anti-mouse SSTR2 monoclonal antibody (ab134152, diluted 1:100; Abcam) was used to investigate whether SSTR2 was expressed in tumor tissues.
Quantitative realtime-PCR and western blot analysis
The tumor tissues were cut into small pieces and homogenized into powder in Trizol reagent until the slurry was without transparent particles (100 mg: 1ml). The cell lysate was transferred into a centrifuge tube and placed at room temperature for 5min. After centrifuging at 12,000rpm, at 4°C for 10min, supernatant was carefully transferred into a new centrifuge tube. The mRNA and protein expressions of MDR1/p-gp, α-tubulin, βIII-tubulin, VEGF and MMP-9 were detected as described in manual from RT-PCR kit (Takara Biotechnology Co.Ltd., Dalian, China) and BeyoECL star (Beyotime Biotechnology, Co.Ltd., China), respectively. Specific primary antibodies (SSTR2(1:2000), MDR1(1:1000), VEGF(1:1000), MMP-9(1:1000), α-tubulin(1:1000), βIII-tubulin(1:1500), β – actin(1:1000)) were applied.
Statistical analysis
Statistical analysis was performed using SPSS 19.0 statistical software. The significance between groups were analysed by variance (ANOVA) and q test (Student-Newman-Keuls). Data were expressed as mean ± standard deviation (SD). A P value of <0.05 was considered statistically significant.
ACKNOWLEDGMENTS
This study was supported by the Youth Projects of Jiangsu Provincial Health Department (No. Q201305), the Science and Technology Research Project of Nanjing City (No. 201201054), a Pre-research Project for National Natural Science Foundation of Southeast University (No. KJ2010493), the Scientific Research Project of Southeast University (No. 3290003101).
CONFLICTS OF INTEREST
The authors confirm that this article content has no conflicts of interest.
REFERENCES
1. Syrios J, Banerjee S, Kaye SB. Advanced epithelial ovarian cancer: from standard chemotherapy to promising molecular pathway targets--where are we now? Anticancer research. 2014; 34:2069-2077.
2. McGuire WP, Hoskins WJ, Brady MF, Kucera PR, Partridge EE, Look KY, Clarke-Pearson DL, Davidson M. Cyclophosphamide and cisplatin compared with paclitaxel and cisplatin in patients with stage III and stage IV ovarian cancer. The New England journal of medicine. 1996; 334:1-6.
3. Harter P, Hilpert F, Mahner S, Heitz F, Pfisterer J and du Bois A. Systemic therapy in recurrent ovarian cancer: current treatment options and new drugs. Expert review of anticancer therapy. 2010; 10:81-88.
4. Bookman MA. First-line chemotherapy in epithelial ovarian cancer. Clinical obstetrics and gynecology. 2012; 55:96-113.
5. Markman M, Bookman MA. Second-line treatment of ovarian cancer. The oncologist. 2000; 5:26-35.
6. Mezo G, Manea M. Receptor-mediated tumor targeting based on peptide hormones. Expert opinion on drug delivery. 2010; 7:79-96.
7. Sun LC, Coy DH. Cytotoxic conjugates of peptide hormones for cancer chemotherapy. Drugs Future. 2008; 33:217-223.
8. Reubi JC. Peptide receptors as molecular targets for cancer diagnosis and therapy. Endocrine reviews. 2003; 24:389-427.
9. Theodoropoulou M, Stalla GK. Somatostatin receptors: from signaling to clinical practice. Frontiers in neuroendocrinology. 2013; 34:228-252.
10. Watt HL, Kharmate G, Kumar U. Biology of somatostatin in breast cancer. Molecular and cellular endocrinology. 2008; 286:251-261.
11. Hejna M, Schmidinger M, Raderer M. The clinical role of somatostatin analogues as antineoplastic agents: much ado about nothing? Annals of oncology. 2002; 13:653-668.
12. Grimberg A. Somatostatin and cancer: applying endocrinology to oncology. Cancer biology & therapy. 2004; 3:731-733.
13. Grozinsky-Glasberg S, Shimon I, Korbonits M, Grossman AB. Somatostatin analogues in the control of neuroendocrine tumours: efficacy and mechanisms. Endocrine-related cancer. 2008; 15:701-720.
14. Gustafsson BI, Kidd M, Modlin IM. Neuroendocrine tumors of the diffuse neuroendocrine system. Current opinion in oncology. 2008; 20:1-12.
15. Borbath I, Leclercq IA, Sempoux C, Abarca-Quinones J, Desaeger C, Horsmans Y. Efficacy of lanreotide in preventing the occurrence of chemically induced hepatocellular carcinoma in rats. Chemico-biological interactions. 2010; 183:238-248.
16. Plonowski A, Schally AV, Koppan M, Nagy A, Arencibia JM, Csernus B, Halmos G. Inhibition of the UCI-107 human ovarian carcinoma cell line by a targeted cytotoxic analog of somatostatin, AN-238. Cancer. 2001; 92:1168-1176.
17. Pozsgai E, Schally AV, Halmos G, Rick F, Bellyei S. The inhibitory effect of a novel cytotoxic somatostatin analogue AN-162 on experimental glioblastoma. Hormone and metabolic research. 2010; 42:781-786.
18. Halmos G, Sun B, Schally AV, Hebert F, Nagy A. Human ovarian cancers express somatostatin receptors. The Journal of clinical endocrinology and metabolism. 2000; 85:3509-3512.
19. Lee S, Choi EJ, Jin C, Kim DH. Activation of PI3K/Akt pathway by PTEN reduction and PIK3CA mRNA amplification contributes to cisplatin resistance in an ovarian cancer cell line. Gynecologic oncology. 2005; 97:26-34.
20. Fraser M, Leung BM, Yan X, Dan HC, Cheng JQ, Tsang BK. p53 is a determinant of X-linked inhibitor of apoptosis protein/Akt-mediated chemoresistance in human ovarian cancer cells. Cancer research. 2003; 63:7081-7088.
21. Hagemeister AL, Kittilson JD, Bergan HE, Sheridan MA. Rainbow trout somatostatin receptor subtypes SSTR1A, SSTR1B, and SSTR2 differentially activate the extracellular signal-regulated kinase and phosphatidylinositol 3-kinase signaling pathways in transfected cells. Journal of molecular endocrinology. 2010; 45:317-327.
22. Mei S, Cammalleri M, Azara D, Casini G, Bagnoli P, Dal Monte M. Mechanisms underlying somatostatin receptor 2 down-regulation of vascular endothelial growth factor expression in response to hypoxia in mouse retinal explants. The Journal of pathology. 2012; 226:519-533.
23. Wang C, Tang C. [Inhibition of human gastric cancer metastasis by ocreotide in vitro and in vivo]. Zhonghua yi xue za zhi. 2002; 82:19-22.
24. Huang MT, Chen ZX, Wei B, Zhang B, Wang CH, Huang MH, Tang CW. [The inhibitive effects of celecoxib combined with octreotide on the metastasis of human gastric cancer in vivo]. Zhonghua yi xue za zhi. 2006; 86:3255-3259.
25. Zhang Y, Zhang H, Wang X, Wang J, Zhang X, Zhang Q. The eradication of breast cancer and cancer stem cells using octreotide modified paclitaxel active targeting micelles and salinomycin passive targeting micelles. Biomaterials. 2012; 33:679-691.
26. Huang MT, Chen ZX, Wei B, Zhang B, Wang CH, Huang MH, Liu R, Tang CW. Preoperative growth inhibition of human gastric adenocarcinoma treated with a combination of celecoxib and octreotide. Acta pharmacologica Sinica. 2007; 28:1842-1850.
27. Dimitroulopoulos D, Xinopoulos D, Tsamakidis K, Zisimopoulos A, Andriotis E, Panagiotakos D, Fotopoulou A, Chrysohoou C, Bazinis A, Daskalopoulou D, Paraskevas E. Long acting octreotide in the treatment of advanced hepatocellular cancer and overexpression of somatostatin receptors: randomized placebo-controlled trial. World journal of gastroenterology. 2007; 13:3164-3170.
28. Tjomsland V, El-Salhy M. Anti-tumour effects of triple therapy with octreotide, galanin and serotonin in comparison with those of 5-fluorouracil/leukovorin on human colon cancer. International journal of oncology. 2005; 27:427-432.
29. Schmid HA, Lambertini C, van Vugt HH, Barzaghi-Rinaudo P, Schafer J, Hillenbrand R, Sailer AW, Kaufmann M, Nuciforo P. Monoclonal antibodies against the human somatostatin receptor subtypes 1-5: development and immunohistochemical application in neuroendocrine tumors. Neuroendocrinology. 2012; 95:232-247.
30. Shen Y, Ren ML, Shi YH, Zhang YX, Cai YL. Octreotide is the favorable alternative for cisplatin resistance reversal of ovarian cancer in vitro and in nude mice in vivo. European journal of gynaecological oncology. 2012; 33:584-590.
31. Shen Y, Ren M, Shi Y, Zhang Y, Cai Y. Octreotide enhances the sensitivity of the SKOV3/DDP ovarian cancer cell line to cisplatin chemotherapy in vitro. Experimental and therapeutic medicine. 2011; 2:1171-1176.
32. Shen Y, Zhang XY, Chen X, Ren ML, Cai YL. Octreotide reverses the resistance of A2780/Pacliaxel ovarian cancer cell line to paclitaxel chemotherapy in vitro. Journal of cancer research and therapeutics. 2016; 12:657-662.
33. Hohla F, Buchholz S, Schally AV, Krishan A, Rick FG, Szalontay L, Papadia A, Halmos G, Koster F, Aigner E, Datz C, Seitz S. Targeted cytotoxic somatostatin analog AN-162 inhibits growth of human colon carcinomas and increases sensitivity of doxorubicin resistant murine leukemia cells. Cancer letters. 2010; 294:35-42.
34. Niu J, Su Z, Xiao Y, Huang A, Li H, Bao X, Li S, Chen Y, Sun M, Ping Q. Octreotide-modified and pH-triggering polymeric micelles loaded with doxorubicin for tumor targeting delivery. European journal of pharmaceutical sciences. 2012; 45:216-226.
35. Sun LC, Coy DH. Somatostatin receptor-targeted anti-cancer therapy. Current drug delivery. 2011; 8:2-10.
36. Corleto VD, Nasoni S, Panzuto F, Cassetta S, Delle Fave G. Somatostatin receptor subtypes: basic pharmacology and tissue distribution. Digestive and liver disease. 2004; 36:S8-16.
37. Huang CM, Wu YT, Chen ST. Targeting delivery of paclitaxel into tumor cells via somatostatin receptor endocytosis. Chemistry & biology. 2000; 7:453-461.
38. Hayeshi R, Chinyanga F, Chengedza S, Mukanganyama S. Inhibition of human glutathione transferases by multidrug resistance chemomodulators in vitro. Journal of enzyme inhibition and medicinal chemistry. 2006; 21:581-587.
39. Szakacs G, Paterson JK, Ludwig JA, Booth-Genthe C, Gottesman MM. Targeting multidrug resistance in cancer. Nature reviews Drug discovery. 2006; 5:219-234.
40. Gottesman MM, Fojo T, Bates SE. Multidrug resistance in cancer: role of ATP-dependent transporters. Nature reviews Cancer. 2002; 2:48-58.
41. Lukyanova NY. Characteristics of homocysteine-induced multidrug resistance of human MCF-7 breast cancer cells and human A2780 ovarian cancer cells. Experimental oncology. 2010; 32:10-14.
42. Schondorf T, Kurbacher CM, Gohring UJ, Benz C, Becker M, Sartorius J, Kolhagen H, Mallman P, Neumann R. Induction of MDR1-gene expression by antineoplastic agents in ovarian cancer cell lines. Anticancer research. 2002; 22:2199-2203.
43. Ebos JM, Kerbel RS. Antiangiogenic therapy: impact on invasion, disease progression, and metastasis. Nature reviews Clinical oncology. 2011; 8:210-221.
44. Akiyama K, Ohga N, Hida Y, Kawamoto T, Sadamoto Y, Ishikawa S, Maishi N, Akino T, Kondoh M, Matsuda A, Inoue N, Shindoh M, Hida K. Tumor endothelial cells acquire drug resistance by MDR1 up-regulation via VEGF signaling in tumor microenvironment. The American journal of pathology. 2012; 180:1283-1293.
45. Li L, Jiang AC, Dong P, Wang H, Xu W, Xu C. MDR1/P-gp and VEGF synergistically enhance the invasion of Hep-2 cells with multidrug resistance induced by taxol. Annals of surgical oncology. 2009; 16:1421-1428.
46. Hu X, Li D, Zhang W, Zhou J, Tang B, Li L. Matrix metalloproteinase-9 expression correlates with prognosis and involved in ovarian cancer cell invasion. Archives of gynecology and obstetrics. 2012; 286:1537-1543.
47. Bergers G, Brekken R, McMahon G, Vu TH, Itoh T, Tamaki K, Tanzawa K, Thorpe P, Itohara S, Werb Z, Hanahan D. Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis. Nature cell biology. 2000; 2:737-744.
48. Kato T, Fujita Y, Nakane K, Kojima T, Nozawa Y, Deguchi T, Ito M. ETS1 promotes chemoresistance and invasion of paclitaxel-resistant, hormone-refractory PC3 prostate cancer cells by up-regulating MDR1 and MMP9 expression. Biochemical and biophysical research communications. 2012; 417:966-971.
49. Chu JJ, Chen KD, Lin YL, Fei CY, Chiang AS, Chiang CD, Lai YK. Taxol induces concomitant hyperphosphorylation and reorganization of vimentin intermediate filaments in 9L rat brain tumor cells. Journal of cellular biochemistry. 1998; 68:472-483.
50. Horwitz SB. Mechanism of action of taxol. Trends in pharmacological sciences. 1992; 13:134-136.
51. Kobayashi Y, Seino K, Hosonuma S, Ohara T, Itamochi H, Isonishi S, Kita T, Wada H, Kojo S, Kiguchi K. Side population is increased in paclitaxel-resistant ovarian cancer cell lines regardless of resistance to cisplatin. Gynecologic oncology. 2011; 121:390-394.
52. Li J, Liu P, Mao H, Wanga A, Zhang X. Emodin sensitizes paclitaxel-resistant human ovarian cancer cells to paclitaxel-induced apoptosis in vitro. Oncology reports. 2009; 21:1605-1610.