
Introduction
MiR, as an evolutionarily conserved gene expression regulator, participates in many fundamental physiological processes. In cancer development, miR functions as a tumor suppressor or an oncogene by targeting specific genes in a 3’UTR dependent manner. This mechanism occurs in a variety of tumors. It is well known that Wnt/β-catenin drives the carcinogenesis, cancer progression, and metastasis in many tumors. Emerging evidence has indicated the importance of interaction between the Wnt/β-catenin signaling pathway (Wnt pathway for short) and miR mediated gene regulation in cancer development. In this paper, we would like to summarize current understanding of the crosstalk between miRs and the Wnt pathway with respect to oncogenesis, cancer metastasis, and drug-resistance.
MiRs overview
MiRs are short (18-25 nucleotides) non-coding RNAs. They repress gene expression through interaction with the 3’UTR of target genes, either inhibiting mRNA translation and/or promoting mRNA degradation. One miR family is potentially able to target 500 genes on average [1]. Likewise, approximately 60% of the mRNAs can be targeted by one or multiple miRs [1]. For example, miR-34 was identified as targeting a number of genes including WNT1/3, β-catenin, LRP6, LEF1, AXIN2, TCF7 post-transcriptionally [2]. MiR-145, -133a and -133b were found to target gene FSCN1 to inhibit cell proliferation and invasion [3]. Thus, changing the expression of miRs will impact a cascade of modifications in gene expression.
MiRs in cancer
Genome-wide profiling has shown that miRs are frequently aberrantly expressed in human neoplasms including gastric cancer, hepatocellular carcinoma (HCC), breast cancer, glioblastoma, prostate cancer and colorectal cancer (CRC) [2]. Software prediction in combination with experimental validation showed that miRs are involved in tumorigenesis, angiogenesis, metastasis, and chemo-resistance by directly targeting specific oncogenes or tumor suppressors. For example, let-7c was shown to be down-regulated in gastric cancer, which was related to Helicobacter pylori-induced gastric carcinogenesis [4]. MiR-194 was identified to promote angiogenesis by suppressing the endogenous angiogenesis inhibitor thrombospondin1 in colon cancer [5]. MiR-21, one of the most well studied miRs, was found to be overexpressed in a wide variety of tumors including glioblastoma, colon, breast, and gastric cancers, leading to enhanced cell invasion and metastasis [6]. In addition, miRs can be used as diagnostic, therapeutic, and prognostic factors. Based on a meta-analysis, the circulating miR-21 expression is a useful noninvasive biological marker for the earlier detection of digestive system cancers such as CRC, HCC, gastric cancer, and a promising prognostic marker for digestive system cancers in the Asian population [7].
Canonical Wnt pathway overview
The Wnt pathway is critical for embryo development and adult tissue homeostasis. Without Wnt protein stimulation, β-catenin is anchored by a destruction complex comprised of APC, GSK3β and Axin [8]. β-catenin is phosphorylated by CK1α and GSK3β, followed by ubiquitination by β-TRCP, resulting in β-catenin proteasomal degradation. The activation of the pathway initiates from the Wnt proteins binding to the receptor Frizzled (FZD) and co-receptor LDL receptor-related protein (LRP) 5 or LRP6. Ligand binding to the receptor leads to the phosphorylation of Dishevelled (DVL). Phosphorylated DVL recruits AXIN and GSK3β to the cell membrane, thereby, inhibiting the phosphorylation of β-catenin and allowing β-catenin to dissociate from the destruction complex. Next, the β-catenin that accumulates localizes to the nucleus and interacts with the TCF/LEF to transactivate downstream gene expression including CyclinD, c-Myc, CD44. Wnt signaling can be suppressed by antagonist molecules such as Wnt inhibitory factor 1 (WIF1), secreted frizzled-related protein family (sFRP) and Dickkopf (DKK) (reviewed in [9]).
Canonical Wnt pathway in cancer
Aberrant activation of the Wnt pathway is highly associated with tumorigenesis and tumor metastasis. Wnt signaling components are commonly mutated in many human cancers. Exome-sequencing has revealed that most of CRC patients harbor loss of function mutations in APC [10]. In some CRC patients, Axin2 is mutated [11], or β-catenin has activating point mutations [12]. A rare but recurrent fusion between VTI1A and TCF7L2, the gene encoding TCF4, has also been found in CRC patients [13]. In CRC and a host of other cancers, Wnt signaling components are mutated. Oncogenic β-catenin mutations are present in melanoma, HCC, gastric, pancreas, ovarian, and endometrial cancers (reviewed in [14]). Inactivating mutations of Axin were observed in HCC [15]. In addition to germline/somatic mutations, aberrant activation of Wnt pathway can result from epigenetic modifications, such as miR regulation. We will discuss this in detail in later sections.
Crosstalk between miR and the Wnt pathway
Dysregulation of miR induces constitutively active Wnt signaling activity in cancer, while expression of miR, in turn, is intensely controlled by Wnt signaling. In order to identify miRs that regulate the Wnt pathway activity, 470 miRs were screened in a cell-based assay in human HEK293 cells and 38 candidate miRs were identified [16]. MiRs activate or repress the Wnt pathway at multiple levels by targeting Wnt ligand/receptor and ligand/receptor associated proteins, β-catenin, β-catenin interacting complex, Wnt pathway transcription factors, multiple Wnt signaling pathway components and components in other signaling pathways (Figure 1). Meanwhile, Wnt activation increases expression of miR through binding of β-catenin to TCF/LEF which then binds to promoter regions to activate transcription. Moreover, there are mutual feedback loops between some miRs and Wnt signaling components. In summary, Wnt activation and miR mediated gene regulation are reciprocal causation to drive oncogenesis.
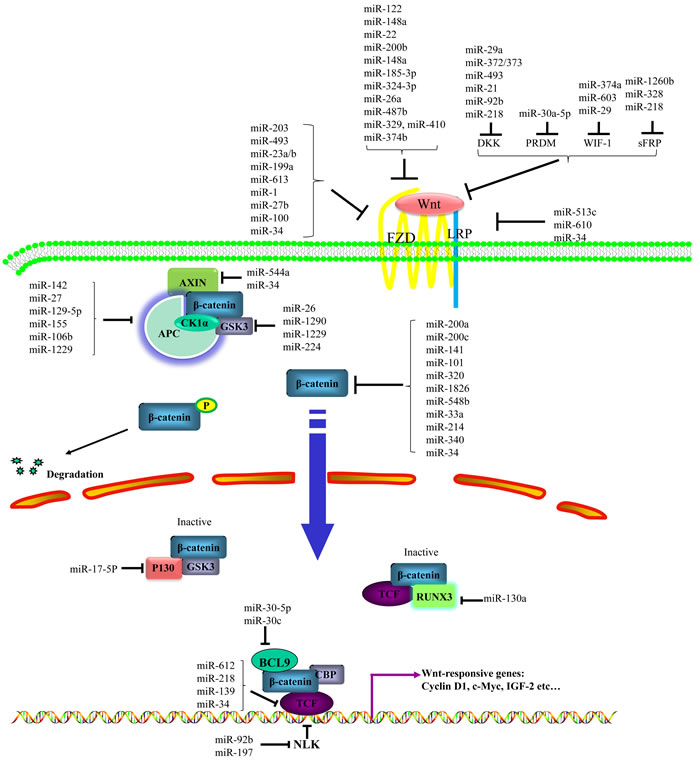
Figure 1: Regulation of miRs on Wnt/β-catenin signaling pathway. MiRs activate or inhibit canonical Wnt pathway at multiple levels by targeting Wnt ligands/receptors and ligands/receptors associated proteins, β-catenin, β-catenin interacting complex, Wnt pathway transcription factors, multiple Wnt signaling pathway components and components in other signaling pathways.
MiRs modulate the Wnt pathway
Targeting Wnt ligands/receptors and ligands/receptors associated inhibitory proteins
It is well established that Wnt activation initiates from the Wnt proteins binding to the FZD and LRP5 or LRP6. If miRs target the Wnt ligand/ receptor such as WNT, FZD and LRP, the pathway is repressed. Both miR-122 and -148a were found to be down-regulated in HCC and to repress the Wnt pathway by targeting WNT1. MiR-122 was shown to suppress cell proliferation and promote cell apoptosis [17]. MiR-148a was reported to inhibit the metastasis of HCCs by blocking EMT and cancer stem cell like properties [18]. Other miRs including miR-22, -200b, -185-3p, -324-3p, -26a, 487b, -329, -410 and -374b also target Wnt ligand to inhibit the Wnt/β-catenin signaling pathway [19-26] (Figure 1 and Table 1). Another group of miRs targeting FZD and LRP act like tumor suppressors. For example, miR-1 and -100 were identified to suppress breast cancer progression by inhibiting the Wnt pathway through targeting the FZD7 and FZD8 respectively [27, 28]. MiR-513c was found to suppress glioblastoma cell proliferation predominantly through direct suppression of the expression of LRP6 [29].
Table 1: MiRs targeting Wnt ligands/receptors and associated proteins
miR |
Target in Wnt signaling |
Effect |
Cell Type |
Reference |
Targeting Wnt ligands/receptors and associated proteins |
||||
miR-122 |
WNT1 |
Tumor suppressor |
HCC, Glioma |
|
miR-148a |
WNT1,WNT10B |
Tumor suppressor |
HCC, Endometrial cancer |
|
miR-22 |
WNT1 |
Tumor suppressor |
Gastric cancer |
[19] |
miR-200b |
WNT1, PKCα, ZEB2 |
Tumor suppressor |
Gastric cancer, Glimoma |
|
miR-185-3p |
WNT2B |
Tumor suppressor |
Nasopharyngeal carcinoma |
[21] |
miR-324-3p |
WNT2B |
Tumor suppressor |
Nasopharyngeal carcinoma |
[22] |
miR-26a |
WNT5A |
Tumor suppressor |
Prostate cancer |
[25] |
miR-487b |
WNT5A, SUZ12, BMI1, MYC, and KRAS |
Tumor suppressor |
Lung cancer |
[23] |
miR-329, miR-410 |
WNT7B |
Tumor suppressor |
Oral squamous cell carcinoma |
[24] |
miR-374b |
WNT16, AKT1 |
Tumor suppressor |
T-cell Lymphoblastic Lymphoma |
[26] |
miR-203 |
FZD2 |
Tumor suppressor |
Lung cancer |
[117] |
miR-493 |
FZD4 |
Tumor suppressor |
Bladder cancer |
[118] |
miR-23a/b |
FZD5, FZD7 |
Tumor suppressor |
Pancreatic cancer, Colon cancer |
|
miR-199a |
FZD7, E-cadherin |
Tumor suppressor/ Oncogene |
HCC, Gastric cancer |
|
miR-613 |
FZD7 |
Tumor suppressor |
Prostate cancer |
[123] |
miR-1 |
FZD7, TNKS2 |
Tumor suppressor |
Breast cancer |
[27] |
miR-27b |
FZD7 |
Tumor suppressor |
Gastric cancer |
[124] |
miR-100 |
FZD8 |
Tumor suppressor |
Breast cancer |
[28] |
miR-513c |
LRP6 |
Tumor suppressor |
Glioblastoma |
[29] |
miR-610 |
LRP6, TBL1X |
Tumor suppressor |
HCC |
[125] |
miR-218 |
LGR4 |
Tumor suppressor |
Prostate cancer |
[126] |
miR-374a |
WIF1, PTEN, WNT5A |
Oncogene |
Breast cancer |
|
miR-603 |
WIF1, CTNNBIP1 |
Oncogene |
Glioma |
[32] |
miR-29 |
demethylation of WIF1, N-myc interactor |
Tumor suppressor/ Oncogene |
NSCLC, Breast cancer |
|
miR-1260b |
sFRP1, DKK2, Smad4 |
Oncogene |
Renal cancer |
[33] |
miR-328 |
sFRP1 |
Oncogene |
Glioma |
[34] |
miR-29a |
DKK1, Kremen2, sFRP2 |
Oncogene |
Pancreatic cancer |
[35] |
miR-372/373 |
DKK1 |
Oncogene |
CRC, Breast Cancer |
[36] |
miR-493 |
DKK1 |
Oncogene |
Gastric cancer |
[129] |
miR-21 |
DKK2, TGFβR2 |
Oncogene |
Oral cancer, Colon cancer |
|
miR-92b |
DKK3,NLK |
Oncogene |
Glioma |
|
miR-218 |
SOST, DKK2, SFRP2 |
Oncogene |
Breast cancer |
[112] |
miR-30a-5p |
PRDM |
Oncogene |
Glioma |
[131] |
Conversely, Wnt signaling is activated when miRs target Wnt antagonists such as WIF, sFRP, DKK, and PR domain proteins (PRDM). MiR-374a [30, 31], -603 [32] -1260b [33], -328 [34] were discovered to activate Wnt/β-catenin signaling by targeting antagonists WIF1 or sFRP1. The DKK and PRDM protein families block Wnt signaling by binding to the LRP5/LRP6. MiR-29a [35], -372&373 [36], -21 [37] and -92b [38] were found to enhance the Wnt pathway activity via targeting DKK, which resulted in either accelerating cancer cell proliferation/invasion or inducing resistance to chemotherapy. Additional examples of miR regulating Wnt ligands/receptors and their associated inhibitory proteins are listed in Table 1 and Figure 1.
The Wnt receptor is at the crux between extracellular ligands and intracellular responses such as survival or apoptosis, proliferation or growth arrest, drug susceptibility or drug resistance. Thus targeting the receptor and its associated proteins by miRs fundamentally influences signaling transduction at the initiation phase. As discussed, miR regulation of the Wnt/β-catenin cascade is multi-faceted with regards to receptor activation and warrants interest for possible therapeutic interventions.
Targeting β-catenin
β-catenin, the most important signaling factor in the Wnt pathway, transduces the signal by directly binding to TCF/LEF transcription factors and turning on downstream gene expressions which promote cell proliferation, migration, EMT, cancer cell metastasis and chemo-resistance [9]. A number of miRs have been found to suppress the Wnt pathway by modulating β-catenin. The well-studied miR-200 cluster is usually under-expressed in cancer cells and is identified to target β-catenin to inhibit cancer progression [39-45]. Five members are included in the family: miR-200a, -200b, -200c, -141 and -429. MiR-200a was identified to be reduced in meningioma, HCC, and gastric cancer [39-43]. This decrease blocked the Wnt pathway by two mechanisms: it targeted the 3’-UTR of β-catenin and the EMT transcription factors zinc finger E-box binding homeobox 1/2 (ZEB1/2). The resulting effect was the reduction of β-catenin and inhibition of Wnt signaling. MiR-200c was shown to repress Wnt pathway by targeting β-catenin directly in breast cancer [44]. MiR-200b was established to have suppressive effects on the proliferation, migration, invasion and EMT of glioma cells via targeting ZEB2 [46]. Moreover, miR-141 was also found to suppress β-catenin expression in breast cancer [45] and down-regulate SOX17 expression, causing activation of Wnt signaling in esophageal cancer [47].
Many other miRs target β-catenin as well, besides the miR-200 family. MiR-214 [48, 49], -320 [50], -101 [51], -1826 [52, 53], -548b [30], and -33a [54], were found to block Wnt pathway by targeting the 3’UTR of β-catenin in HCC, prostate, breast, colon, urological, and lung cancer, respectively (Figure 1 and Table 2).
Table 2: MiRs targeting β-catenin/TCF and associated proteins
miR |
Target in Wnt signaling |
Effect |
Cell Type |
Reference |
Targeting β-catenin |
||||
miR-200a |
β-catenin, ZEB1/2 |
Tumor suppressor |
Meningiomas, Gastric Cancer, HCC |
|
miR-200c |
β-catenin |
Tumor suppressor |
Breast cancer |
[44] |
miR-141 |
β-catenin, SOX17 |
Tumor suppressor |
Breast cancer, Esophageal cancer |
|
miR-101 |
β-catenin |
Tumor suppressor |
Colon cancer |
[51] |
miR-320 |
β-catenin,FOXM1 |
Tumor suppressor |
Prostate cancer, Colon cancer |
|
miR-1826 |
β-catenin |
Tumor suppressor |
Bladder cancer, Renal cancer |
|
miR-548b |
β-catenin |
Tumor suppressor |
Lung cancer |
[30] |
miR-33a |
β-catenin |
Tumor suppressor |
HCC |
[54] |
miR-214 |
β-catenin, EZH2 |
Tumor suppressor |
HCC |
|
miR-340 |
β-catenin, ROCK1, c-MYC |
Tumor suppressor |
Breast cancer |
[133] |
Targeting multiple Wnt signaling components |
||||
miR-34 |
WNT1/3, β-catenin, LRP6, LEF1, AXIN2,TCF7 |
Tumor suppressor |
Lung cancer, Breast cancer, CRC, Prostate cancer, HCC |
|
Targeting β-catenin interacting proteins |
||||
miR-142 |
APC |
Oncogene |
Breast cancer |
[135] |
miR-27 |
APC |
Oncogene |
Gastric cancer |
[136] |
miR-129-5p |
APC |
Oncogene |
Squamous cell carcinoma |
[137] |
miR-155 |
APC, CK1α, HBP1 |
Oncogene |
HCC, Papillary thyroid carcinoma, Liposarcoma, Glioma |
|
miR-106b |
APC |
Oncogene |
HCC |
[140] |
miR-1229 |
APC, GSK3β |
Oncogene |
Breast cancer |
[141] |
miR-26 |
GSK3β |
Oncogene |
Cholangiocarcinoma |
[63] |
miR-1290 |
GSK3β |
Oncogene |
Lung adenocarcinoma |
[64] |
miR-224 |
GSK3β, SFRP2 |
Oncogene |
CRC |
[142] |
miR-146a |
Numb |
Oncogene |
CRC |
[113] |
miR-544a |
E-cadherin, AXIN2 |
Oncogene |
Gastric cancer |
[65] |
miR-145 |
Catenin δ-1 |
Tumor suppressor |
Colon cancer |
[143] |
miR-490-3p |
FRAT1 |
Tumor suppressor |
CRC |
[144] |
Targeting Wnt signaling related transcription factors |
||||
miR-612 |
TCF/LEF |
Tumor suppressor |
HCC |
[145] |
miR-218 |
LEF1, BMI1 |
Tumor suppressor |
Glioblastoma, Glioma |
|
miR-139 |
TCF-4 |
Tumor suppressor |
HCC |
[67] |
miR-17-5P |
P130, HBP1 |
Oncogene |
CRC, Breast cancer |
|
miR-197 |
NLK |
Oncogene |
Ovarian cancer |
[75] |
miR-130a |
RUNX3 |
Oncogene |
HCC, Gastric Cancer |
|
miR-30-5p/ miR-30c |
BCL9 |
Tumor suppressor |
Myeloma, Prostate cancer |
|
miR-452 |
Sox7 |
Oncogene |
HCC |
[150] |
Certain miRs not only target β-catenin alone, but also directly suppress a set of multiple components of the Wnt signaling cascade to regulate Wnt activity such as miR-34. MiR-34 trans-activated by p53 inhibits the Wnt pathway by targeting the 3’-UTRs of a set of conserved targets which are elements of the Wnt pathway. These genes include WNT1/3, β-catenin, LRP6, AXIN2 and LEF1 [55-59]. The inactivating mutation of p53 turns on canonical Wnt signaling and enhances EMT by inducing miR-34 mediated reduction of target gene expression in human cancer.
β-catenin is the central and most well studied signaling molecule in the Wnt pathway. By targeting β-catenin, miRs directly repress the Wnt signaling activity (Figure 1 and Table 2). Moreover, miR-34s are able to target a number of Wnt signaling components, allowing efficient regulation of important physiological functions associated with cancer initiation and progression. This is a very appealing property for therapy development. As a result, a miR mimic targeting miR-34 was developed into a therapeutic (MRX34) and is in a phase 1 clinical trial for solid and hematological tumors [60].
Targeting β-catenin-interacting complex
Due to the significance of β-catenin in Wnt signaling, miRs targeting the β-catenin interacting proteins such as APC, GSKβ, AXIN and similar associates, consequently regulate Wnt signaling. APC is a scaffolding protein for the APC/GSK3β/CK1α/β-catenin/AXIN complex which mediates destruction of β-catenin. It is a common tumor suppressor, mutated in a variety of cancers. MiR-155 has been reported to negatively regulate APC in multiple carcinomas including HCC and papillary thyroid carcinoma [61, 62] to promote cell proliferation and tumorigenesis. These data suggest miR-155 as a promising target for diagnosis and treatment of HCC and papillary thyroid cancer.
In regard to the kinase in the “destruction complex”, GSK-3β was targeted by miR-26a as revealed in target prediction and validated in biological experiments. MiR-26a targets GSK-3β to promote cholangiocarcinoma growth through concomitant activation of Wnt/β-catenin [63]. GSK-3β is also a target of miR-1290 in lung cancer [64].
Another scaffolding protein in the “destruction complex”, AXIN2 is targeted by miR-544a. MiR-544a was identified as an EMT-inducing miR in a cell-based reporter system with a 328-miRs library. In gastric cancer, it targets E-cadherin and AXIN2 to promote β-catenin translocation to the nucleus, resulting in active Wnt signaling [65]. It suggests that miR-544a might be a potential target for treating advanced gastric cancer. Additional miRs targeting β-catenin associated factors are listed in Table 2 and Figure 1.
In summary, miRs regulate the Wnt pathway by targeting β-catenin directly and by inhibiting expression of β-catenin interacting proteins, revealing a regulatory network that is intertwined and complicated.
Targeting Wnt pathway transcription factors
The established action mechanism of β-catenin is activation of downstream gene expression by binding the transcriptional factor TCF/LEF. MiRs that target the TCF/LEF transcription factors and other associated co-activators/co-repressors regulate transcription transduced by Wnt signaling (Figure 1 and Table 2). For example, miR-218 inhibits glioblastoma cell invasiveness by suppressing the Wnt pathway via direct targeting of oncogenic transcription factor LEF1 [66]. MiR-139 attenuates the proliferative and invasive ability of HCC by inhibiting the Wnt pathway through TCF4 [67].
Other transcriptional factors and co-activators/co-repressors besides the TCF family are modulated by miRs to affect Wnt signaling. P130 is a transcription factor associated with GSK3β which sequesters β-catenin in an inactive form as a P130/GSK3β/β-catenin complex in the cell nucleus [68]. It was found that P130 is targeted by miR-17-5p to activate the Wnt pathway in CRC to promote cancer advancement [69]. HBP1, a transcriptional repressor for the Wnt pathway, is also targeted by miR-17-5p to activate Wnt signaling to promote cell migration and invasion in breast cancer [70]. RUNX3 (Runt related transcription factor 3) forms a ternary complex with β-catenin/TCF4 to inhibit Wnt signaling activity [71]. Elevated expression of miR-130a may directly repress RUNX3 to activate Wnt/β-catenin signaling and subsequently lead to chemo-resistance in HCC cells [72].
NLK (Nemo-like kinase), an evolutionarily conserved protein kinase, is an inhibitor for the Wnt pathway by binding to and phosphorylating TCF/LEF-1 family proteins [73]. MiR-92b was identified to promote glioma proliferation and invasion by targeting NLK which resulted in active Wnt/β-catenin signaling [74]. More recently, miR-197 was reported to activate Wnt signaling by downregulation of NLK in ovarian cancer [75].
Collectively, research has documented the involvement of miRs in regulating the Wnt pathway at the last checkpoint-mRNA expression. Transcriptional regulation was observed by direct control of TCF/LEF expression and by inhibition of the formation of the active TCF/β-catenin complex. Thus, miR regulation of transcription adds another layer of fine tuning the signaling cascade.
Targeting components in other signaling pathways
More recently, the crosstalk between Wnt signaling and other signaling pathways has been described in more detail. A great number of miRs have been found to regulate the Wnt signaling pathway indirectly by targeting elements in other pathways (Table 3). The function of components from other signaling pathways varied from one to another. They can be classified into three major groups. The largest group comprises transcriptional factors such as E2F1 [76], smad [77] and ZEB [78]. The next set is the group of enzymes including Phosphatase and tensin homolog (PTEN) [79], and Zinc and Ring Finger 3 (ZNRF3) [80]. The final group involves the receptor associated proteins such as Roundabout 3 (ROBO3) [81] and Solute Carrier Family 34 Member 2 ( SLC34A2) [82].
Table 3: MiRs targeting other signaling pathway components
miR |
Target in Wnt signaling |
Effect |
Cell Type |
Reference |
Targeting other signaling pathway components |
||||
Targeting Transcription factors |
||||
miR-182-5p |
Smad4, RECK |
Oncogene |
Bladder cancer |
[77] |
miR-93 |
Smad7, ZNRF3 |
Tumor suppressor/ Oncogene |
CRC, Lung cancer |
|
miR-33b |
ZEB1 |
Tumor suppressor |
Lung adenocarcinoma |
[78] |
miR-376c |
LRH-1 |
Tumor suppressor |
NSCLC |
[151] |
miR-145 |
Oct4 |
Tumor suppressor |
Lung cancer |
[152] |
miR-191 |
WT1 |
Oncogene |
Lung cancer |
[153] |
miR-153 |
WWOX |
Oncogene |
HCC |
[154] |
miR-19b/20a/92a |
E2F1, HIPK1 |
Oncogene |
Gastric cancer |
[84] |
miR-19 |
MEF2D |
Tumor suppressor |
Gastric cancer |
[86] |
Targeting Enzyme |
||||
miR-301a |
PTEN |
Oncogene |
Breast cancer |
[79] |
miR-429 |
PTEN |
Oncogene |
HCC |
[88] |
miR-146b-5p |
ZNRF3 |
Oncogene |
Thyroid cancer, Osteosarcoma |
|
miR-506 |
EZH2 |
Tumor suppressor |
Colon cancer |
[92] |
miR-144 |
EZH2 |
Tumor suppressor |
Bladder cancer |
[93] |
miR-29c |
GNA13, PTP4A |
Tumor suppressor |
CRC |
[156] |
Targeting Receptor associated proteins |
||||
miR-576-5p |
ITGBL1 |
Oncogene |
NSCLC |
[157] |
miR-410 |
SLC34A2 |
Oncogene |
NSCLC |
[82] |
miR-483-5p |
RhoGDI1, ALCAM |
Oncogene |
Lung adenocarcinoma |
[158] |
miR-494 |
CXCR4 |
Tumor suppressor |
Breast cancer |
[159] |
miR-152 |
TNFRSF6B |
Tumor suppressor |
HCC |
[160] |
miR-383 |
ROBO3 |
Tumor suppressor |
Pancreatic cancer |
[81] |
Targeting transcription factors in other signaling pathways
Transcription factor E2F1 preferentially binds to retinoblastoma protein pRB to prevent cell-cycle progression. It mediates p53-dependent/independent apoptosis. E2F1 was shown to suppress the Wnt pathway by trans-activating Catenin Beta Interacting Protein 1 (CTNNBIP1) [76] or by up-regulating AXIN2 [83]. MiR-19b/20a/92a was found to sustain the stem cell self-renewal and promote cell proliferation in gastric cancer by increasing Wnt signaling through reduction of E2F1 and dishevelled-associated protein1 (HIPK1) expression [84].
Myocyte enhancer factor 2D (MEF2D) is a transcription factor of the MEF2 family. The family members were found to be involved in carcinogenesis and cancer progression [85]. MiR-19 inhibits cell proliferation in gastric cancer by targeting MEF2D. MEF2D inhibition leads to repression of the Wnt pathway. The miR-19/MEF2D/Wnt/β-catenin axis is critical for gastric cancer cell survival and proliferation, suggesting miR-19 as a potential therapeutic target for gastric cancer [86].
Take another transcription factor for example, smad4 is an important transcription factor for TGFβ signaling which participates in multiple biological processes including cell survival, apoptosis, growth and differentiation. For the TGFβ/Smad pathway, smad4, smad7 and TGFβ are targeted by miR-182-5p [77], miR-93 [87] and miR-21 [37] in bladder cancer, CRC, and colon cancer respectively.
Targeting enzymes in other signaling pathways
In addition to transcription factors, miR regulates Wnt signaling indirectly by targeting enzymes in other signaling pathways. PTEN, a critical enzymatic protein in PI3K/AKT signaling pathway, was found to be targeted by miR-301a [79] and miR-429 [88] in breast cancer and HCC respectively. It was shown that up-regulated miR-301a accelerates breast cancer progression by targeting PTEN, consequently resulting in Wnt signaling activation. The study suggests that miR-301a may be a potential therapeutic target for breast cancer. MiR-429 increases the metastatic capability of HCC by activating both Wnt and PI3K signaling through PTEN, suggesting miR-429 as a novel target for HCC treatment.
Enhancer of zeste homolog 2 (EZH2) catalyzes the methylation of histone H3 in target gene promoters to repress gene expression. It regulates cell proliferation, migration, metastasis, and chemo-resistance by silencing tumor suppressor genes such as APC [89-91]. EZH2 overexpression leads to activation of the Wnt pathway via APC reduction. EZH2 was reported to be targeted by miR-506 [92] and miR-144 [93] in colon and bladder cancer respectively. MiR-506 abrogates tumor proliferation and metastasis in colon cancer and miR-144 decreases cell growth in bladder cancer via Wnt pathway inhibition.
In another example, ZNRF3, an E3 ubiquitin ligase, inhibits Wnt pathway by increasing proteolysis of FZD and LRP6 [94]. It was reported that miR-146b-5p induces EMT and may promote papillary thyroid cancer metastasis through Wnt pathway activation by targeting ZNRF3. MiR-93 was also found to target ZNRF3 to promote lung carcinoma growth through Wnt signaling activation [95]. These data suggest ZNRF3 as a potential therapeutic target for thyroid and lung cancer.
Targeting receptor associated proteins in other signaling pathways
In addition to transcriptional factors and enzymes, miRs also regulate the Wnt pathway by targeting receptor associated proteins. For instance, ROBO receptors are members of the immunoglobulin superfamily of cell adhesion molecules (ICAMs). The ROBO pathway has been implicated in cancer development and progression [96]. The molecular mechanism by which dysregulated ROBO signaling promotes tumor progression is not well studied. ROBO3 was identified as a target of miR-383. ROBO3 increases Wnt pathway activity by sequestering sFRP to augment pancreatic cancer progression [81].
SLC34A2 is a plasma membrane protein that mediates sodium-dependent phosphate transport. SLC34A2 is involved in tumorigenesis. MiR-410 promotes the tumorigenesis and development of NSCLC by down-regulating SLC34A2 and activating the Wnt pathway. MiR-410 might be a new potential therapeutic target for NSCLC [82].
Integrin beta-like 1 (ITGBL1), is a beta-integrin similar protein. The function of ITGBL1 is not clear to date. ITGBL1, targeted by miR-576-5p, inhibits NSCLC progression by suppressing the Wnt pathway [97].
Additional examples of miRs that regulate Wnt by targeting elements in other signaling pathways are listed in Table 3. Wnt/β-catenin and other signaling pathways converge at numerous nodes of the cellular regulatory network, therefore, miRs regulate the Wnt pathway indirectly by targeting the node proteins with differential functions. Most of the targeted proteins are emerging novel oncogenes or tumor suppressors thereby the specific mechanisms by which they modulate Wnt/β-catenin signaling are not well understood. Additional studies revealing the regulation mechanisms will provide novel insight into the complex regulatory network that leads to tumorigenesis, cancer metastasis, and drug-resistance.
The Wnt pathway regulates the expression of miRs
Thus far, it has been shown that miRs regulate the Wnt pathway however, the reverse is true-Wnt pathway modulates the expression of miR positively and negatively in a variety of tumors to induce carcinogenesis, progression, and drug resistance (Table 4). The aberrant expression profile of miRs was in part due to the constitutive activation of Wnt pathway in a plethora of cancers. β-catenin enhances downstream target expression by binding to TCF in the promoter and recruiting a panel of co-activators to trans-activate gene transcription.
Table 4: MiRs up or down regulated by Wnt signaling.
Cancer Type |
Down |
Up |
Reference |
CRC |
miR-215,miR-137 |
miR-708, miR-31, miR-135b, miR-21, miR-145, miR-126, miR-139-5p, miR-574-3p, miR-30e, miR-150 |
|
Gastric cancer |
miR-1234-3p, miR-135b-5p, miR-210,miR-4739,miR-122a |
miR-20a-3p, miR-23b-5p, miR-335-3p, miR-423-5p,miR-455-3p |
|
HCC |
miR-375 |
miR-770 , miR-183/96/182 |
|
Breast cancer |
let-7 |
miR-182, miR-125b |
In colon cancer, Brian et al. have used quantitative PCR arrays as well as mathematical analysis to discover distinctively expressed miRs in tumors resulting from loss of function mutation of APC. MiR-215 and -137 are repressed and miR-708, -135b and -31 levels are increased in APC mutation induced tumors. Target prediction and pathway analysis suggest that these miRs control signaling pathways critical for transformation [98]. Conversely, the expression profile of miRs was explored in TCF activity disrupted cells [99]. The miR transcriptome was assessed with the TaqMan Array in DLD1 CRC cells expressing dominant negative (dn) TCF4 alleles. Fifty-one miRs were found to be upregulated and 9 downregulated by at least two fold in dnTCF4 cells. Sixteen of the increased miRNAs, such as miR-574-3p,-139-5p and -30e-3p, were shown to be markedly reduced in CRC tissue. Some selected miRs (miR-30e-3p -145, -139-5p and -126) significantly suppress cell proliferation. Consistently, miR-30e is trans-activated by β-catenin/TCF4 complex during intestinal cell differentiation in rat [100]. In another study, a bioinformatics approach with ChIP-PCR was utilized to discover that TCF4 trans-activates miR-21 by directly binding to its promoter in colon cancer cells [101]. Similarly, miR-21 is trans-activated by β-catenin in a STAT3 dependent manner to promote cell invasion in glioma [102]. Aberrant Wnt/β-catenin signaling is the driving force for CRC carcinogenesis and progression. These identified downstream miRs of the Wnt pathway reveal part of the mechanisms through which Wnt/β-catenin promotes CRC tumorigenesis.
Other than CRC, 30% of gastric cancers carry nuclear accumulation of β-catenin, highlighting the importance of the Wnt pathway in gastric oncogenesis. To identify the miRs regulated by Wnt signaling, a miR microarray was conducted with β-catenin interference in gastric cancer cells to detect expression differences in the miR transcriptome [103]. The expression of miRs-4739, -210, -135b-5p, and-1234-3p are significantly increased and that of miRs-23b-5p, -20a-3p, -423-5p, -455-3p and -335-3p are significantly decreased. β-catenin interference results in delayed cell proliferation, increased apoptosis, weakened invasion of gastric cancer cells, and increased chemo-sensitivity of cancer cells. Also in gastric cancer, Wang et al. have demonstrated that miR-122a, a new tumor suppressor, is down-regulated by Wnt/β-catenin signaling [104]. The down-regulation of miR-122a mediated by aberrant Wnt pathwaysignaling is critical for the pathogenesis of gastrointestinal cancer.
Additionally, several studies explored the underlying mechanisms of HCC progression from the perspective of Wnt signaling, as β-catenin is a major oncogene in HCC and is activated in 30-40% of cases [105]. Wnt/β-catenin signaling was shown to up-regulate miR-770 to promote cell proliferation in HCC [106]. Also in HCC, Wnt signaling elevates miR-183/96/182 levels to augment cell invasion. β-catenin was found to increase the miR-183/96/182 transcription by physically interacting with TCF complex on miR-183/96/182 promoter region [107]. Another study found a strong correlation between β-catenin-activating mutation and down-regulation of miR-375 expression in HCC according to miR profiling in hepatocellular tumors and normal liver samples [108].
In breast cancer, miR-182 is highly expressed and is up-regulated by the Wnt pathway. The overexpression of miR-182 increases tumorigenicity and invasiveness by repressing RECK [109]. In addition, the expression of miR-125b is elevated by overexpression of SNAIL in a β-catenin dependent manner to enhance cancer stem cell enrichment and chemo-resistance in breast cancer [110]. This suggests that miR-125 may serve as a novel target to overcome chemo-resistance in cancer cells. Lastly, let-7 miR was discovered to be repressed by Wnt pathway via trans-activating Lin28 in breast cancer stem cells [111]. It was demonstrated that the Wnt pathway induces Lin28 up-regulation and let-7 down-regulation to enhance breast cancer cell expansion.
Feedback loop between miRs and the Wnt pathway
There are positive feedback circuits between miR and Wnt signaling. MiR-218 and Wnt pathway form a positive feedback loop to increase osteoblast differentiation and abnormal expression of osteoblastic genes in breast cancer cells [112]. MiR-218 activates the Wnt pathway by targeting three Wnt signaling inhibitors (Sclerostin (SOST), DKK2, and sFRP2) during the process of osteogenesis. MiR-218 is also induced by active Wnt signaling, creating a positive feedback loop. MiR-146a expression is activated by SNAIL in a β-catenin dependent manner in CRC stem cells. In turn, miR-146a stabilizes β-catenin by targeting Numb [113]. A feedback loop is formed to direct symmetric cell division through activation of the Wnt pathway. MiR-372&373 induced by Wnt dependent transcription in turn, activates Wnt/β-catenin signaling by targeting Wnt signaling inhibitors including DKK1 [36].
Mutual inhibition between miR and Wnt pathway has also been observed. In glioma cells, miR-122 inhibits the Wnt pathway, which negatively regulates the expression of miR-122 [114]. In CRC, miR-101 and the Wnt pathway form a mutual inhibitory relationship [51]. The Wnt pathway activity represses the expression of miR-101 while overexpression of miR-101 strongly impairs β-catenin nuclear accumulation.
In addition, a negative feedback loop was also found between miR and the Wnt signaling pathway. MiR-483-3p targets β-catenin which induces the expression of miR-483-3p [115]. They form a negative feedback circuit in normal cells however, this negative feedback loop is inactivated if β-catenin carries an activating mutation.
In summary, miRs regulate the Wnt signaling pathway and Wnt signaling in turn modulates expression of miRs in human neoplasms. In addition, both form mutual feedback circuits, thereby increasing the connectivity and complexity of the regulatory network. Chartering this network will facilitate the development and advancement of miR-based clinical applications.
MiR-based therapeutics
Recently, miR-based therapeutics have been developed based on the regulatory network of miRs and Wnt/β-catenin. MiR-based anticancer therapeutic approaches have used several strategies such as miR sponges, anti-miR oligonucleotides, miR masks, and small molecule inhibitors (reviewed in [1]). MiR-34 is lost or down-regulated in different types of cancer such as glioblastoma, HCC, cervical, ovarian, colon, and lung cancer. The ability of miR-34 to regulate a set of multiple Wnt signaling components in WNT pathway (discussed above) makes it an excellent candidate for novel Wnt-targeted therapy. MRX34 is a liposomal mimic of miR-34. A multicenter Phase I clinical trial of MRX34 is currently recruiting patients (ClinicalTrials.gov; NCT01829971). MRX34 is used for patients with advanced HCC, other selected solid cancers and hematologic malignancies. The scope of the trial is to establish the safety, pharmacokinetics and pharmacodynamics of MRX34. The company planned to finish the phase 1 trial by the end of 2016 and start the phase 2 clinical trial in 2017.
Pioneering work has certain challenges that must be overcome before miR-based therapies can be used in a clinical setting. Low bioavailability, specific tissue delivery, off target side effects, miR instability, immunogenicity and tumorigenicity are major problems to solve before the application in human clinical trials [1]. Despite these challenges, targeting miRs in cancer to rewire signaling networks is an applicable and rational strategy with great potential for success.
Conclusions
Collectively, we have summarized the current understanding of crosstalk between miRs and the canonical Wnt signaling cascade in various types of tumors. Communication intersects in multiple planes to modulate cell proliferation, migration, cancer metastasis, and drug response. MiR-based therapeutics have entered clinical trials; a stage where opportunities and challenges co-exist for miR-based therapeutics for cancer. A better understanding of miRs and the Wnt signaling regulatory network will provide insights to further the development of miR-based remedies.
Acknowledgments
The work is supported by: National Natural Youth Science Foundation of China (31601028) and China Postdoctoral Science Foundation (2015M582417) to Y Peng; National Nature Science Foundation of China (81172282), the Planned Science and Technology Project of Shenzhen (GJHS20120621142654087 and JCYJ20140418095735574), the Shenzhen Peacock Plan (KQCX20130621101141669), the Key Laboratory Project of Shenzhen (ZDSY20130329101130496) to Z Jin; National Natural Youth Science Foundation of China (81302151), the Planned Science and Technology Project of Shenzhen (JCYJ20160422170722474) and Medical science and technology research foundation of Guangdong Province(A2016112)to X Zhang. Thank Dr. Adina Stanculescu for editing the manuscript.
Conflicts of interest
The authors disclose no potential conflicts of interest.
References
1. Garzon R, Marcucci G and Croce CM. Targeting microRNAs in cancer: rationale, strategies and challenges. Nature reviews Drug discovery. 2010; 9:775-789.
2. Sun X, He Y, Huang C, Ma TT and Li J. Distinctive microRNA signature associated of neoplasms with the Wnt/beta-catenin signaling pathway. Cellular signalling. 2013; 25:2805-2811.
3. Kano M, Seki N, Kikkawa N, Fujimura L, Hoshino I, Akutsu Y, Chiyomaru T, Enokida H, Nakagawa M and Matsubara H. miR-145, miR-133a and miR-133b: Tumor-suppressive miRNAs target FSCN1 in esophageal squamous cell carcinoma. International journal of cancer. 2010; 127:2804-2814.
4. Fassan M, Saraggi D, Balsamo L, Cascione L, Castoro C, Coati I, De Bernard M, Farinati F, Guzzardo V, Valeri N, Zambon CF and Rugge M. Let-7c down-regulation in Helicobacter pylori-related gastric carcinogenesis. Oncotarget. 2016; 7:4915-24. doi: 10.18632/oncotarget.6642.
5. Sundaram P, Hultine S, Smith LM, Dews M, Fox JL, Biyashev D, Schelter JM, Huang Q, Cleary MA, Volpert OV and Thomas-Tikhonenko A. p53-responsive miR-194 inhibits thrombospondin-1 and promotes angiogenesis in colon cancers. Cancer research. 2011; 71:7490-7501.
6. Krichevsky AM and Gabriely G. miR-21: a small multi-faceted RNA. Journal of cellular and molecular medicine. 2009; 13:39-53.
7. Yin C, Zhou X, Dang Y, Yan J and Zhang G. Potential Role of Circulating MiR-21 in the Diagnosis and Prognosis of Digestive System Cancer: A Systematic Review and Meta-Analysis. Medicine. 2015; 94:e2123.
8. Song JL, Nigam P, Tektas SS and Selva E. microRNA regulation of Wnt signaling pathways in development and disease. Cellular signalling. 2015; 27:1380-1391.
9. Clevers H and Nusse R. Wnt/beta-catenin signaling and disease. Cell. 2012; 149:1192-1205.
10. Wood LD, Parsons DW, Jones S, Lin J, Sjoblom T, Leary RJ, Shen D, Boca SM, Barber T, Ptak J, Silliman N, Szabo S, Dezso Z, Ustyanksky V, Nikolskaya T, Nikolsky Y, et al. The genomic landscapes of human breast and colorectal cancers. Science. 2007; 318:1108-1113.
11. Liu W, Dong X, Mai M, Seelan RS, Taniguchi K, Krishnadath KK, Halling KC, Cunningham JM, Boardman LA, Qian C, Christensen E, Schmidt SS, Roche PC, Smith DI and Thibodeau SN. Mutations in AXIN2 cause colorectal cancer with defective mismatch repair by activating beta-catenin/TCF signalling. Nature genetics. 2000; 26:146-147.
12. Morin PJ, Sparks AB, Korinek V, Barker N, Clevers H, Vogelstein B and Kinzler KW. Activation of beta-catenin-Tcf signaling in colon cancer by mutations in beta-catenin or APC. Science. 1997; 275:1787-1790.
13. Bass AJ, Lawrence MS, Brace LE, Ramos AH, Drier Y, Cibulskis K, Sougnez C, Voet D, Saksena G, Sivachenko A, Jing R, Parkin M, Pugh T, Verhaak RG, Stransky N, Boutin AT, et al. Genomic sequencing of colorectal adenocarcinomas identifies a recurrent VTI1A-TCF7L2 fusion. Nature genetics. 2011; 43:964-968.
14. Rosenbluh J, Wang X and Hahn WC. Genomic insights into WNT/beta-catenin signaling. Trends in pharmacological sciences. 2014; 35:103-109.
15. Rubinfeld B, Robbins P, El-Gamil M, Albert I, Porfiri E and Polakis P. Stabilization of beta-catenin by genetic defects in melanoma cell lines. Science. 1997; 275:1790-1792.
16. Anton R, Chatterjee SS, Simundza J, Cowin P and Dasgupta R. A systematic screen for micro-RNAs regulating the canonical Wnt pathway. PloS one. 2011; 6:e26257.
17. Xu J, Zhu X, Wu L, Yang R, Yang Z, Wang Q and Wu F. MicroRNA-122 suppresses cell proliferation and induces cell apoptosis in hepatocellular carcinoma by directly targeting Wnt/beta-catenin pathway. Liver international. 2012; 32:752-760.
18. Yan H, Dong X, Zhong X, Ye J, Zhou Y, Yang X, Shen J and Zhang J. Inhibitions of epithelial to mesenchymal transition and cancer stem cells-like properties are involved in miR-148a-mediated anti-metastasis of hepatocellular carcinoma. Molecular carcinogenesis. 2014; 53:960-969.
19. Tang H, Kong Y, Guo J, Tang Y, Xie X, Yang L, Su Q and Xie X. Diallyl disulfide suppresses proliferation and induces apoptosis in human gastric cancer through Wnt-1 signaling pathway by up-regulation of miR-200b and miR-22. Cancer letters. 2013; 340:72-81.
20. Wang Z, Humphries B, Xiao H, Jiang Y and Yang C. MicroRNA-200b suppresses arsenic-transformed cell migration by targeting protein kinase Calpha and Wnt5b-protein kinase Calpha positive feedback loop and inhibiting Rac1 activation. The Journal of biological chemistry. 2014; 289:18373-18386.
21. Li G, Wang Y, Liu Y, Su Z, Liu C, Ren S, Deng T, Huang D, Tian Y and Qiu Y. miR-185-3p regulates nasopharyngeal carcinoma radioresistance by targeting WNT2B in vitro. Cancer science. 2014; 105:1560-1568.
22. Li G, Liu Y, Su Z, Ren S, Zhu G, Tian Y and Qiu Y. MicroRNA-324-3p regulates nasopharyngeal carcinoma radioresistance by directly targeting WNT2B. European journal of cancer. 2013; 49:2596-2607.
23. Xi S, Xu H, Shan J, Tao Y, Hong JA, Inchauste S, Zhang M, Kunst TF, Mercedes L and Schrump DS. Cigarette smoke mediates epigenetic repression of miR-487b during pulmonary carcinogenesis. The Journal of clinical investigation. 2013; 123:1241-1261.
24. Shiah SG, Hsiao JR, Chang WM, Chen YW, Jin YT, Wong TY, Huang JS, Tsai ST, Hsu YM, Chou ST, Yen YC, Jiang SS, Shieh YS, Chang IS, Hsiao M and Chang JY. Downregulated miR329 and miR410 promote the proliferation and invasion of oral squamous cell carcinoma by targeting Wnt-7b. Cancer research. 2014; 74:7560-7572.
25. Zhao S, Ye X, Xiao L, Lian X, Feng Y, Li F and Li L. MiR-26a inhibits prostate cancer progression by repression of Wnt5a. Tumour biology. 2014; 35:9725-9733.
26. Qian D, Chen K, Deng H, Rao H, Huang H, Liao Y, Sun X, Lu S, Yuan Z, Xie D and Cai Q. MicroRNA-374b Suppresses Proliferation and Promotes Apoptosis in T-cell Lymphoblastic Lymphoma by Repressing AKT1 and Wnt-16. Clinical cancer research. 2015.
27. Liu T, Hu K, Zhao Z, Chen G, Ou X, Zhang H, Zhang X, Wei X, Wang D, Cui M and Liu C. MicroRNA-1 down-regulates proliferation and migration of breast cancer stem cells by inhibiting the Wnt/beta-catenin pathway. Oncotarget. 2015; 6:41638-49. doi: 10.18632/oncotarget.5873.
28. Jiang Q, He M, Guan S, Ma M, Wu H, Yu Z, Jiang L, Wang Y, Zong X, Jin F and Wei M. MicroRNA-100 suppresses the migration and invasion of breast cancer cells by targeting FZD-8 and inhibiting Wnt/beta-catenin signaling pathway. Tumour biology. 2015.
29. Xu J, Sun T and Hu X. microRNA-513c suppresses the proliferation of human glioblastoma cells by repressing low-density lipoprotein receptor-related protein 6. Molecular medicine reports. 2015; 12:4403-4409.
30. Wang Y, Xia H, Zhuang Z, Miao L, Chen X and Cai H. Axl-altered microRNAs regulate tumorigenicity and gefitinib resistance in lung cancer. Cell death & disease. 2014; 5:e1227.
31. Cai J, Guan H, Fang L, Yang Y, Zhu X, Yuan J, Wu J and Li M. MicroRNA-374a activates Wnt/beta-catenin signaling to promote breast cancer metastasis. The Journal of clinical investigation. 2013; 123:566-579.
32. Guo M, Zhang X, Wang G, Sun J, Jiang Z, Khadarian K, Yu S, Zhao Y, Xie C, Zhang K, Zhu M, Shen H, Lin Z, Jiang C, Shen J and Zheng Y. miR-603 promotes glioma cell growth via Wnt/beta-catenin pathway by inhibiting WIF1 and CTNNBIP1. Cancer letters. 2015; 360:76-86.
33. Hirata H, Ueno K, Nakajima K, Tabatabai ZL, Hinoda Y, Ishii N and Dahiya R. Genistein downregulates onco-miR-1260b and inhibits Wnt-signalling in renal cancer cells. British journal of cancer. 2013; 108:2070-2078.
34. Delic S, Lottmann N, Stelzl A, Liesenberg F, Wolter M, Gotze S, Zapatka M, Shiio Y, Sabel MC, Felsberg J, Reifenberger G and Riemenschneider MJ. MiR-328 promotes glioma cell invasion via SFRP1-dependent Wnt-signaling activation. Neuro-oncology. 2014; 16:179-190.
35. Nagano H, Tomimaru Y, Eguchi H, Hama N, Wada H, Kawamoto K, Kobayashi S, Mori M and Doki Y. MicroRNA-29a induces resistance to gemcitabine through the Wnt/beta-catenin signaling pathway in pancreatic cancer cells. International journal of oncology. 2013; 43:1066-1072.
36. Zhou AD, Diao LT, Xu H, Xiao ZD, Li JH, Zhou H and Qu LH. beta-Catenin/LEF1 transactivates the microRNA-371-373 cluster that modulates the Wnt/beta-catenin-signaling pathway. Oncogene. 2012; 31:2968-2978.
37. Yu Y, Kanwar SS, Patel BB, Oh PS, Nautiyal J, Sarkar FH and Majumdar AP. MicroRNA-21 induces stemness by downregulating transforming growth factor beta receptor 2 (TGFbetaR2) in colon cancer cells. Carcinogenesis. 2012; 33:68-76.
38. Li Q, Shen K, Zhao Y, Ma C, Liu J and Ma J. MiR-92b inhibitor promoted glioma cell apoptosis via targeting DKK3 and blocking the Wnt/beta-catenin signaling pathway. Journal of translational medicine. 2013; 11:302.
39. Tian Y, Pan Q, Shang Y, Zhu R, Ye J, Liu Y, Zhong X, Li S, He Y, Chen L, Zhao J, Chen W, Peng Z and Wang R. MicroRNA-200 (miR-200) cluster regulation by achaete scute-like 2 (Ascl2): impact on the epithelial-mesenchymal transition in colon cancer cells. The Journal of biological chemistry. 2014; 289:36101-36115.
40. Liu J, Ruan B, You N, Huang Q, Liu W, Dang Z, Xu W, Zhou T, Ji R, Cao Y, Li X, Wang D, Tao K and Dou K. Downregulation of miR-200a induces EMT phenotypes and CSC-like signatures through targeting the beta-catenin pathway in hepatic oval cells. PloS one. 2013; 8:e79409.
41. Cong N, Du P, Zhang A, Shen F, Su J, Pu P, Wang T, Zjang J, Kang C and Zhang Q. Downregulated microRNA-200a promotes EMT and tumor growth through the wnt/beta-catenin pathway by targeting the E-cadherin repressors ZEB1/ZEB2 in gastric adenocarcinoma. Oncology reports. 2013; 29:1579-1587.
42. Su J, Zhang A, Shi Z, Ma F, Pu P, Wang T, Zhang J, Kang C and Zhang Q. MicroRNA-200a suppresses the Wnt/beta-catenin signaling pathway by interacting with beta-catenin. International journal of oncology. 2012; 40:1162-1170.
43. Saydam O, Shen Y, Wurdinger T, Senol O, Boke E, James MF, Tannous BA, Stemmer-Rachamimov AO, Yi M, Stephens RM, Fraefel C, Gusella JF, Krichevsky AM and Breakefield XO. Downregulated microRNA-200a in meningiomas promotes tumor growth by reducing E-cadherin and activating the Wnt/beta-catenin signaling pathway. Molecular and cellular biology. 2009; 29:5923-5940.
44. Ahmad A, Sarkar SH, Bitar B, Ali S, Aboukameel A, Sethi S, Li Y, Bao B, Kong D, Banerjee S, Padhye SB and Sarkar FH. Garcinol regulates EMT and Wnt signaling pathways in vitro and in vivo, leading to anticancer activity against breast cancer cells. Molecular cancer therapeutics. 2012; 11:2193-2201.
45. Abedi N, Mohammadi-Yeganeh S, Koochaki A, Karami F and Paryan M. miR-141 as potential suppressor of beta-catenin in breast cancer. Tumour biology. 2015.
46. Li J, Yuan J, Yuan X, Zhao J, Zhang Z, Weng L and Liu J. MicroRNA-200b inhibits the growth and metastasis of glioma cells via targeting ZEB2. International journal of oncology. 2016; 48:541-550.
47. Jia Y, Yang Y, Zhan Q, Brock MV, Zheng X, Yu Y, Herman JG and Guo M. Inhibition of SOX17 by microRNA 141 and methylation activates the WNT signaling pathway in esophageal cancer. The Journal of molecular diagnostics. 2012; 14:577-585.
48. Xia H, Ooi LL and Hui KM. MiR-214 targets beta-catenin pathway to suppress invasion, stem-like traits and recurrence of human hepatocellular carcinoma. PloS one. 2012; 7:e44206.
49. Wang X, Chen J, Li F, Lin Y, Zhang X, Lv Z and Jiang J. MiR-214 inhibits cell growth in hepatocellular carcinoma through suppression of beta-catenin. Biochemical and biophysical research communications. 2012; 428:525-531.
50. Hsieh IS, Chang KC, Tsai YT, Ke JY, Lu PJ, Lee KH, Yeh SD, Hong TM and Chen YL. MicroRNA-320 suppresses the stem cell-like characteristics of prostate cancer cells by downregulating the Wnt/beta-catenin signaling pathway. Carcinogenesis. 2013; 34:530-538.
51. Strillacci A, Valerii MC, Sansone P, Caggiano C, Sgromo A, Vittori L, Fiorentino M, Poggioli G, Rizzello F, Campieri M and Spisni E. Loss of miR-101 expression promotes Wnt/beta-catenin signalling pathway activation and malignancy in colon cancer cells. The Journal of pathology. 2013; 229:379-389.
52. Hirata H, Hinoda Y, Ueno K, Shahryari V, Tabatabai ZL and Dahiya R. MicroRNA-1826 targets VEGFC, beta-catenin (CTNNB1) and MEK1 (MAP2K1) in human bladder cancer. Carcinogenesis. 2012; 33:41-48.
53. Hirata H, Hinoda Y, Ueno K, Nakajima K, Ishii N and Dahiya R. MicroRNA-1826 directly targets beta-catenin (CTNNB1) and MEK1 (MAP2K1) in VHL-inactivated renal cancer. Carcinogenesis. 2012; 33:501-508.
54. Fang Y, Feng Y, Wu T, Srinivas S, Yang W, Fan J, Yang C and Wang S. Aflatoxin B1 negatively regulates Wnt/beta-catenin signaling pathway through activating miR-33a. PloS one. 2013; 8:e73004.
55. Zhu L, Gao J, Huang K, Luo Y, Zhang B and Xu W. miR-34a screened by miRNA profiling negatively regulates Wnt/beta-catenin signaling pathway in Aflatoxin B1 induced hepatotoxicity. Scientific reports. 2015; 5:16732.
56. Liang J, Li Y, Daniels G, Sfanos K, De Marzo A, Wei J, Li X, Chen W, Wang J, Zhong X, Melamed J, Zhao J and Lee P. LEF1 Targeting EMT in Prostate Cancer Invasion Is Regulated by miR-34a. Molecular cancer research. 2015; 13:681-688.
57. Kim NH, Cha YH, Kang SE, Lee Y, Lee I, Cha SY, Ryu JK, Na JM, Park C, Yoon HG, Park GJ, Yook JI and Kim HS. p53 regulates nuclear GSK-3 levels through miR-34-mediated Axin2 suppression in colorectal cancer cells. Cell cycle. 2013; 12:1578-1587.
58. Cha YH, Kim NH, Park C, Lee I, Kim HS and Yook JI. MiRNA-34 intrinsically links p53 tumor suppressor and Wnt signaling. Cell cycle. 2012; 11:1273-1281.
59. Kim NH, Kim HS, Li XY, Lee I, Choi HS, Kang SE, Cha SY, Ryu JK, Yoon D, Fearon ER, Rowe RG, Lee S, Maher CA, Weiss SJ and Yook JI. A p53/miRNA-34 axis regulates Snail1-dependent cancer cell epithelial-mesenchymal transition. The Journal of cell biology. 2011; 195:417-433.
60. Shah MY and Calin GA. MicroRNAs as therapeutic targets in human cancers. Wiley interdisciplinary reviews RNA. 2014; 5:537-548.
61. Zhang X, Li M, Zuo K, Li D, Ye M, Ding L, Cai H, Fu D, Fan Y and Lv Z. Upregulated miR-155 in papillary thyroid carcinoma promotes tumor growth by targeting APC and activating Wnt/beta-catenin signaling. The Journal of clinical endocrinology and metabolism. 2013; 98:E1305-1313.
62. Zhang Y, Wei W, Cheng N, Wang K, Li B, Jiang X and Sun S. Hepatitis C virus-induced up-regulation of microRNA-155 promotes hepatocarcinogenesis by activating Wnt signaling. Hepatology. 2012; 56:1631-1640.
63. Zhang J, Han C and Wu T. MicroRNA-26a promotes cholangiocarcinoma growth by activating beta-catenin. Gastroenterology. 2012; 143:246-256 e248.
64. Wu L, Liu T, Xiao Y, Li X, Zhu Y, Zhao Y, Bao J and Wu C. Polygonatum odoratum lectin induces apoptosis and autophagy by regulation of microRNA-1290 and microRNA-15a-3p in human lung adenocarcinoma A549 cells. International journal of biological macromolecules. 2015.
65. Yanaka Y, Muramatsu T, Uetake H, Kozaki K and Inazawa J. miR-544a induces epithelial-mesenchymal transition through the activation of WNT signaling pathway in gastric cancer. Carcinogenesis. 2015; 36:1363-1371.
66. Liu Y, Yan W, Zhang W, Chen L, You G, Bao Z, Wang Y, Wang H, Kang C and Jiang T. MiR-218 reverses high invasiveness of glioblastoma cells by targeting the oncogenic transcription factor LEF1. Oncology reports. 2012; 28:1013-1021.
67. Gu W, Li X and Wang J. miR-139 regulates the proliferation and invasion of hepatocellular carcinoma through the WNT/TCF-4 pathway. Oncology reports. 2014; 31:397-404.
68. Petrov N, Zhidkova O, Serikov V, Zenin V and Popov B. Induction of Wnt/beta-catenin signaling in mouse mesenchymal stem cells is associated with activation of the p130 and E2f4 and formation of the p130/Gsk3beta/beta-catenin complex. Stem cells and development. 2012; 21:589-597.
69. Ma Y, Zhang P, Wang F, Zhang H, Yang Y, Shi C, Xia Y, Peng J, Liu W, Yang Z and Qin H. Elevated oncofoetal miR-17-5p expression regulates colorectal cancer progression by repressing its target gene P130. Nature communications. 2012; 3:1291.
70. Li H, Bian C, Liao L, Li J and Zhao RC. miR-17-5p promotes human breast cancer cell migration and invasion through suppression of HBP1. Breast cancer research and treatment. 2011; 126:565-575.
71. Ito K, Lim AC, Salto-Tellez M, Motoda L, Osato M, Chuang LS, Lee CW, Voon DC, Koo JK, Wang H, Fukamachi H and Ito Y. RUNX3 attenuates beta-catenin/T cell factors in intestinal tumorigenesis. Cancer cell. 2008; 14:226-237.
72. Xu N, Shen C, Luo Y, Xia L, Xue F, Xia Q and Zhang J. Upregulated miR-130a increases drug resistance by regulating RUNX3 and Wnt signaling in cisplatin-treated HCC cell. Biochemical and biophysical research communications. 2012; 425:468-472.
73. Yamada M, Ohnishi J, Ohkawara B, Iemura S, Satoh K, Hyodo-Miura J, Kawachi K, Natsume T and Shibuya H. NARF, an nemo-like kinase (NLK)-associated ring finger protein regulates the ubiquitylation and degradation of T cell factor/lymphoid enhancer factor (TCF/LEF). The Journal of biological chemistry. 2006; 281:20749-20760.
74. Wang K, Wang X, Zou J, Zhang A, Wan Y, Pu P, Song Z, Qian C, Chen Y, Yang S and Wang Y. miR-92b controls glioma proliferation and invasion through regulating Wnt/beta-catenin signaling via Nemo-like kinase. Neuro-oncology. 2013; 15:578-588.
75. Zou D, Wang D, Li R, Tang Y, Yuan L, Long X and Zhou Q. MiR-197 induces Taxol resistance in human ovarian cancer cells by regulating NLK. Tumour biology. 2015; 36:6725-6732.
76. Wu Z, Zheng S, Li Z, Tan J and Yu Q. E2F1 suppresses Wnt/beta-catenin activity through transactivation of beta-catenin interacting protein ICAT. Oncogene. 2011; 30:3979-3984.
77. Hirata H, Ueno K, Shahryari V, Tanaka Y, Tabatabai ZL, Hinoda Y and Dahiya R. Oncogenic miRNA-182-5p targets Smad4 and RECK in human bladder cancer. PloS one. 2012; 7:e51056.
78. Qu J, Li M, An J, Zhao B, Zhong W, Gu Q, Cao L, Yang H and Hu C. MicroRNA-33b inhibits lung adenocarcinoma cell growth, invasion, and epithelial-mesenchymal transition by suppressing Wnt/beta-catenin/ZEB1 signaling. International journal of oncology. 2015.
79. Ma F, Zhang J, Zhong L, Wang L, Liu Y, Wang Y, Peng L and Guo B. Upregulated microRNA-301a in breast cancer promotes tumor metastasis by targeting PTEN and activating Wnt/beta-catenin signaling. Gene. 2014; 535:191-197.
80. Deng X, Wu B, Xiao K, Kang J, Xie J, Zhang X and Fan Y. MiR-146b-5p promotes metastasis and induces epithelial-mesenchymal transition in thyroid cancer by targeting ZNRF3. Cellular physiology and biochemistry. 2015; 35:71-82.
81. Han S, Cao C, Tang T, Lu C, Xu J, Wang S, Xue L, Zhang X and Li M. ROBO3 promotes growth and metastasis of pancreatic carcinoma. Cancer letters. 2015; 366:61-70.
82. Zhang X, Ke X, Pu Q, Yuan Y, Yang W, Luo X, Jiang Q, Hu X, Gong Y, Tang K, Su X, Liu L, Zhu W and Wei Y. MicroRNA-410 acts as oncogene in NSCLC through downregulating SLC34A2 via activating Wnt/beta-catenin pathway. Oncotarget. 2016; 7:14569-85. doi: 10.18632/oncotarget.7538.
83. Hughes TA and Brady HJ. Regulation of axin2 expression at the levels of transcription, translation and protein stability in lung and colon cancer. Cancer letters. 2006; 233:338-347.
84. Wu Q, Yang Z, Wang F, Hu S, Yang L, Shi Y and Fan D. MiR-19b/20a/92a regulates the self-renewal and proliferation of gastric cancer stem cells. Journal of cell science. 2013; 126:4220-4229.
85. Pon JR and Marra MA. MEF2 transcription factors: developmental regulators and emerging cancer genes. Oncotarget. 2016; 7:2297-2312. doi: 10.18632/oncotarget.6223.
86. Xu K and Zhao YC. MEF2D/Wnt/beta-catenin pathway regulates the proliferation of gastric cancer cells and is regulated by microRNA-19. Tumour biology. 2016.
87. Tang Q, Zou Z, Zou C, Zhang Q, Huang R, Guan X, Li Q, Han Z, Wang D, Wei H, Gao X and Wang X. MicroRNA-93 suppress colorectal cancer development via Wnt/beta-catenin pathway downregulating. Tumour biology. 2015; 36:1701-1710.
88. Tang J, Li L, Huang W, Sui C, Yang Y, Lin X, Hou G, Chen X, Fu J, Yuan S, Li S, Wen W, Tang S, Cao D, Wu M, Chen L, et al. MiR-429 increases the metastatic capability of HCC via regulating classic Wnt pathway rather than epithelial-mesenchymal transition. Cancer letters. 2015; 364:33-43.
89. Qi W, Chan H, Teng L, Li L, Chuai S, Zhang R, Zeng J, Li M, Fan H, Lin Y, Gu J, Ardayfio O, Zhang JH, Yan X, Fang J, Mi Y, et al. Selective inhibition of Ezh2 by a small molecule inhibitor blocks tumor cells proliferation. Proceedings of the National Academy of Sciences of the United States of America. 2012; 109:21360-21365.
90. Ren G, Baritaki S, Marathe H, Feng J, Park S, Beach S, Bazeley PS, Beshir AB, Fenteany G, Mehra R, Daignault S, Al-Mulla F, Keller E, Bonavida B, de la Serna I and Yeung KC. Polycomb protein EZH2 regulates tumor invasion via the transcriptional repression of the metastasis suppressor RKIP in breast and prostate cancer. Cancer research. 2012; 72:3091-3104.
91. Zhang Y, Liu G, Lin C, Liao G and Tang B. Silencing the EZH2 gene by RNA interference reverses the drug resistance of human hepatic multidrug-resistant cancer cells to 5-Fu. Life sciences. 2013; 92:896-902.
92. Zhang Y, Lin C, Liao G, Liu S, Ding J, Tang F, Wang Z, Liang X, Li B, Wei Y, Huang Q, Li X and Tang B. MicroRNA-506 suppresses tumor proliferation and metastasis in colon cancer by directly targeting the oncogene EZH2. Oncotarget. 2015; 6:32586-32601. doi: 10.18632/oncotarget.5309.
93. Guo Y, Ying L, Tian Y, Yang P, Zhu Y, Wang Z, Qiu F and Lin J. miR-144 downregulation increases bladder cancer cell proliferation by targeting EZH2 and regulating Wnt signaling. The FEBS journal. 2013; 280:4531-4538.
94. Jiang X, Charlat O, Zamponi R, Yang Y and Cong F. Dishevelled promotes Wnt receptor degradation through recruitment of ZNRF3/RNF43 E3 ubiquitin ligases. Molecular cell. 2015; 58:522-533.
95. Shi J, Jiang X, Yu Z, He G, Ning H, Wu Z, Cai Y, Yu H and Chen A. ZNRF3 contributes to the growth of lung carcinoma via inhibiting Wnt/beta-catenin pathway and is regulated by miR-93. Tumour biology. 2015.
96. Gara RK, Kumari S, Ganju A, Yallapu MM, Jaggi M and Chauhan SC. Slit/Robo pathway: a promising therapeutic target for cancer. Drug discovery today. 2015; 20:156-164.
97. Gan X, Liu Z, Tong B and Zhou J. Epigenetic downregulated ITGBL1 promotes non-small cell lung cancer cell invasion through Wnt/PCP signaling. Tumour biology. 2015.
98. Necela BM, Carr JM, Asmann YW and Thompson EA. Differential expression of microRNAs in tumors from chronically inflamed or genetic (APC(Min/+)) models of colon cancer. PloS one. 2011; 6:e18501.
99. Schepeler T, Holm A, Halvey P, Nordentoft I, Lamy P, Riising EM, Christensen LL, Thorsen K, Liebler DC, Helin K, Orntoft TF and Andersen CL. Attenuation of the beta-catenin/TCF4 complex in colorectal cancer cells induces several growth-suppressive microRNAs that target cancer promoting genes. Oncogene. 2012; 31:2750-2760.
100. Liao Y and Lonnerdal B. Beta-catenin/TCF4 transactivates miR-30e during intestinal cell differentiation. Cellular and molecular life sciences. 2010; 67:2969-2978.
101. Lan F, Yue X, Han L, Shi Z, Yang Y, Pu P, Yao Z and Kang C. Genome-wide identification of TCF7L2/TCF4 target miRNAs reveals a role for miR-21 in Wnt-driven epithelial cancer. International journal of oncology. 2012; 40:519-526.
102. Han L, Yue X, Zhou X, Lan FM, You G, Zhang W, Zhang KL, Zhang CZ, Cheng JQ, Yu SZ, Pu PY, Jiang T and Kang CS. MicroRNA-21 expression is regulated by beta-catenin/STAT3 pathway and promotes glioma cell invasion by direct targeting RECK. CNS neuroscience & therapeutics. 2012; 18:573-583.
103. Dong L, Deng J, Sun ZM, Pan AP, Xiang XJ, Zhang L, Yu F, Chen J, Sun Z, Feng M and Xiong JP. Interference with the beta-catenin gene in gastric cancer induces changes to the miRNA expression profile. Tumour biology. 2015; 36:6973-6983.
104. Wang X, Lam EK, Zhang J, Jin H and Sung JJ. MicroRNA-122a functions as a novel tumor suppressor downstream of adenomatous polyposis coli in gastrointestinal cancers. Biochemical and biophysical research communications. 2009; 387:376-380.
105. Levrero M and Zucman-Rossi J. Mechanisms of HBV-induced hepatocellular carcinoma. Journal of hepatology. 2016; 64:S84-S101.
106. Wu WJ, Shi J, Hu G, Yu X, Lu H, Yang ML, Liu B and Wu ZX. Wnt/beta-catenin signaling inhibits FBXW7 expression by upregulation of microRNA-770 in hepatocellular carcinoma. Tumour biology. 2015.
107. Leung WK, He M, Chan AW, Law PT and Wong N. Wnt/beta-Catenin activates MiR-183/96/182 expression in hepatocellular carcinoma that promotes cell invasion. Cancer letters. 2015; 362:97-105.
108. Ladeiro Y, Couchy G, Balabaud C, Bioulac-Sage P, Pelletier L, Rebouissou S and Zucman-Rossi J. MicroRNA profiling in hepatocellular tumors is associated with clinical features and oncogene/tumor suppressor gene mutations. Hepatology. 2008; 47:1955-1963.
109. Chiang CH, Hou MF and Hung WC. Up-regulation of miR-182 by beta-catenin in breast cancer increases tumorigenicity and invasiveness by targeting the matrix metalloproteinase inhibitor RECK. Biochimica et biophysica acta. 2013; 1830:3067-3076.
110. Liu Z, Liu H, Desai S, Schmitt DC, Zhou M, Khong HT, Klos KS, McClellan S, Fodstad O and Tan M. miR-125b functions as a key mediator for snail-induced stem cell propagation and chemoresistance. The Journal of biological chemistry. 2013; 288:4334-4345.
111. Cai WY, Wei TZ, Luo QC, Wu QW, Liu QF, Yang M, Ye GD, Wu JF, Chen YY, Sun GB, Liu YJ, Zhao WX, Zhang ZM and Li BA. The Wnt-beta-catenin pathway represses let-7 microRNA expression through transactivation of Lin28 to augment breast cancer stem cell expansion. Journal of cell science. 2013; 126:2877-2889.
112. Hassan MQ, Maeda Y, Taipaleenmaki H, Zhang W, Jafferji M, Gordon JA, Li Z, Croce CM, van Wijnen AJ, Stein JL, Stein GS and Lian JB. miR-218 directs a Wnt signaling circuit to promote differentiation of osteoblasts and osteomimicry of metastatic cancer cells. The Journal of biological chemistry. 2012; 287:42084-42092.
113. Hwang WL, Jiang JK, Yang SH, Huang TS, Lan HY, Teng HW, Yang CY, Tsai YP, Lin CH, Wang HW and Yang MH. MicroRNA-146a directs the symmetric division of Snail-dominant colorectal cancer stem cells. Nature cell biology. 2014; 16:268-280.
114. Wang G, Zhao Y and Zheng Y. MiR-122/Wnt/beta-catenin regulatory circuitry sustains glioma progression. Tumour biology. 2014; 35:8565-8572.
115. Veronese A, Visone R, Consiglio J, Acunzo M, Lupini L, Kim T, Ferracin M, Lovat F, Miotto E, Balatti V, D’Abundo L, Gramantieri L, Bolondi L, Pekarsky Y, Perrotti D, Negrini M, et al. Mutated beta-catenin evades a microRNA-dependent regulatory loop. Proceedings of the National Academy of Sciences of the United States of America. 2011; 108:4840-4845.
116. Aprelikova O, Palla J, Hibler B, Yu X, Greer YE, Yi M, Stephens R, Maxwell GL, Jazaeri A, Risinger JI, Rubin JS and Niederhuber J. Silencing of miR-148a in cancer-associated fibroblasts results in WNT10B-mediated stimulation of tumor cell motility. Oncogene. 2013; 32:3246-3253.
117. Mine M, Yamaguchi K, Sugiura T, Chigita S, Yoshihama N, Yoshihama R, Hiyake N, Kobayashi Y and Mori Y. miR-203 Inhibits Frizzled-2 Expression via CD82/KAI1 Expression in Human Lung Carcinoma Cells. PloS one. 2015; 10:e0131350.
118. Ueno K, Hirata H, Majid S, Yamamura S, Shahryari V, Tabatabai ZL, Hinoda Y and Dahiya R. Tumor suppressor microRNA-493 decreases cell motility and migration ability in human bladder cancer cells by downregulating RhoC and FZD4. Molecular cancer therapeutics. 2012; 11:244-253.
119. Listing H, Mardin WA, Wohlfromm S, Mees ST and Haier J. MiR-23a/-24-induced gene silencing results in mesothelial cell integration of pancreatic cancer. British journal of cancer. 2015; 112:131-139.
120. Zhang H, Hao Y, Yang J, Zhou Y, Li J, Yin S, Sun C, Ma M, Huang Y and Xi JJ. Genome-wide functional screening of miR-23b as a pleiotropic modulator suppressing cancer metastasis. Nature communications. 2011; 2:554.
121. Zhao X, He L, Li T, Lu Y, Miao Y, Liang S, Guo H, Bai M, Xie H, Luo G, Zhou L, Shen G, Guo C, Bai F, Sun S, Wu K, et al. SRF expedites metastasis and modulates the epithelial to mesenchymal transition by regulating miR-199a-5p expression in human gastric cancer. Cell death and differentiation. 2014; 21:1900-1913.
122. Song J, Gao L, Yang G, Tang S, Xie H, Wang Y, Wang J, Zhang Y, Jin J, Gou Y, Yang Z, Chen Z, Wu K, Liu J and Fan D. MiR-199a regulates cell proliferation and survival by targeting FZD7. PloS one. 2014; 9:e110074.
123. Ren W, Li C, Duan W, Du S, Yang F, Zhou J and Xing J. MicroRNA-613 represses prostate cancer cell proliferation and invasion through targeting Frizzled7. Biochemical and biophysical research communications. 2016; 469:633-638.
124. Geng Y, Lu X, Wu X, Xue L, Wang X and Xu J. MicroRNA-27b suppresses Helicobacter pylori-induced gastric tumorigenesis through negatively regulating Frizzled7. Oncology reports. 2016; 35:2441-2450.
125. Zeng XC, Liu FQ, Yan R, Yi HM, Zhang T, Wang GY, Li Y and Jiang N. Downregulation of miR-610 promotes proliferation and tumorigenicity and activates Wnt/beta-catenin signaling in human hepatocellular carcinoma. Molecular cancer. 2014; 13:261.
126. Li F, Gu C, Tian F, Jia Z, Meng Z, Ding Y and Yang J. miR-218 impedes IL-6-induced prostate cancer cell proliferation and invasion via suppression of LGR4 expression. Oncology reports. 2016; 35:2859-2865.
127. Tan M, Wu J and Cai Y. Suppression of Wnt signaling by the miR-29 family is mediated by demethylation of WIF-1 in non-small-cell lung cancer. Biochemical and biophysical research communications. 2013; 438:673-679.
128. Rostas JW, 3rd, Pruitt HC, Metge BJ, Mitra A, Bailey SK, Bae S, Singh KP, Devine DJ, Dyess DL, Richards WO, Tucker JA, Shevde LA and Samant RS. microRNA-29 negatively regulates EMT regulator N-myc interactor in breast cancer. Molecular cancer. 2014; 13:200.
129. Jia X, Li N, Peng C, Deng Y, Wang J, Deng M, Lu M, Yin J, Zheng G, Liu H and He Z. miR-493 mediated DKK1 down-regulation confers proliferation, invasion and chemo-resistance in gastric cancer cells. Oncotarget. 2016; 7:7044-54. doi: 10.18632/oncotarget.6951.
130. Kawakita A, Yanamoto S, Yamada S, Naruse T, Takahashi H, Kawasaki G and Umeda M. MicroRNA-21 promotes oral cancer invasion via the Wnt/beta-catenin pathway by targeting DKK2. Pathology oncology research. 2014; 20:253-261.
131. Wang X, Wang K, Han L, Zhang A, Shi Z, Zhang K, Zhang H, Yang S, Pu P, Shen C, Yu C and Kang C. PRDM1 is directly targeted by miR-30a-5p and modulates the Wnt/beta-catenin pathway in a Dkk1-dependent manner during glioma growth. Cancer letters. 2013; 331:211-219.
132. Wan LY, Deng J, Xiang XJ, Zhang L, Yu F, Chen J, Sun Z, Feng M and Xiong JP. miR-320 enhances the sensitivity of human colon cancer cells to chemoradiotherapy in vitro by targeting FOXM1. Biochemical and biophysical research communications. 2015; 457:125-132.
133. Mohammadi-Yeganeh S, Paryan M, Arefian E, Vasei M, Ghanbarian H, Mahdian R, Karimipoor M and Soleimani M. MicroRNA-340 inhibits the migration, invasion, and metastasis of breast cancer cells by targeting Wnt pathway. Tumour biology. 2016.
134. Chen WY, Liu SY, Chang YS, Yin JJ, Yeh HL, Mouhieddine TH, Hadadeh O, Abou-Kheir W and Liu YN. MicroRNA-34a regulates WNT/TCF7 signaling and inhibits bone metastasis in Ras-activated prostate cancer. Oncotarget. 2015; 6:441-457. doi: 10.18632/oncotarget.2690.
135. Isobe T, Hisamori S, Hogan DJ, Zabala M, Hendrickson DG, Dalerba P, Cai S, Scheeren F, Kuo AH, Sikandar SS, Lam JS, Qian D, Dirbas FM, Somlo G, Lao K, Brown PO, et al. miR-142 regulates the tumorigenicity of human breast cancer stem cells through the canonical WNT signaling pathway. eLife. 2014; 3.
136. Zhang Z, Liu S, Shi R and Zhao G. miR-27 promotes human gastric cancer cell metastasis by inducing epithelial-to-mesenchymal transition. Cancer genetics. 2011; 204:486-491.
137. Li M, Tian L, Wang L, Yao H, Zhang J, Lu J, Sun Y, Gao X, Xiao H and Liu M. Down-regulation of miR-129-5p inhibits growth and induces apoptosis in laryngeal squamous cell carcinoma by targeting APC. PloS one. 2013; 8:e77829.
138. Yan Z, Che S, Wang J, Jiao Y, Wang C and Meng Q. miR-155 contributes to the progression of glioma by enhancing Wnt/beta-catenin pathway. Tumour biology. 2015; 36:5323-5331.
139. Zhang P, Bill K, Liu J, Young E, Peng T, Bolshakov S, Hoffman A, Song Y, Demicco EG, Terrada DL, Creighton CJ, Anderson ML, Lazar AJ, Calin GG, Pollock RE and Lev D. MiR-155 is a liposarcoma oncogene that targets casein kinase-1alpha and enhances beta-catenin signaling. Cancer research. 2012; 72:1751-1762.
140. Shen G, Jia H, Tai Q, Li Y and Chen D. miR-106b downregulates adenomatous polyposis coli and promotes cell proliferation in human hepatocellular carcinoma. Carcinogenesis. 2013; 34:211-219.
141. Tan Z, Zheng H, Liu X, Zhang W, Zhu J, Wu G, Cao L, Song J, Wu S, Song L and Li J. MicroRNA-1229 overexpression promotes cell proliferation and tumorigenicity and activates Wnt/beta-catenin signaling in breast cancer. Oncotarget. 2016; 7:24076-87. doi: 10.18632/oncotarget.8119.
142. Li T, Lai Q, Wang S, Cai J, Xiao Z, Deng D, He L, Jiao H, Ye Y, Liang L, Ding Y and Liao W. MicroRNA-224 sustains Wnt/beta-catenin signaling and promotes aggressive phenotype of colorectal cancer. Journal of experimental & clinical cancer research. 2016; 35:21.
143. Yamada N, Noguchi S, Mori T, Naoe T, Maruo K and Akao Y. Tumor-suppressive microRNA-145 targets catenin delta-1 to regulate Wnt/beta-catenin signaling in human colon cancer cells. Cancer letters. 2013; 335:332-342.
144. Zheng K, Zhou X, Yu J, Li Q, Wang H, Li M, Shao Z, Zhang F, Luo Y, Shen Z, Chen F, Shi F, Cui C, Zhao D, Lin Z, Zheng W, et al. Epigenetic silencing of miR-490-3p promotes development of an aggressive colorectal cancer phenotype through activation of the Wnt/beta-catenin signaling pathway. Cancer letters. 2016; 376:178-187.
145. Tang J, Tao ZH, Wen D, Wan JL, Liu DL, Zhang S, Cui JF, Sun HC, Wang L, Zhou J, Fan J and Wu WZ. MiR-612 suppresses the stemness of liver cancer via Wnt/beta-catenin signaling. Biochemical and biophysical research communications. 2014; 447:210-215.
146. Tu Y, Gao X, Li G, Fu H, Cui D, Liu H, Jin W and Zhang Y. MicroRNA-218 inhibits glioma invasion, migration, proliferation, and cancer stem-like cell self-renewal by targeting the polycomb group gene Bmi1. Cancer research. 2013; 73:6046-6055.
147. Jiang H, Yu WW, Wang LL and Peng Y. miR-130a acts as a potential diagnostic biomarker and promotes gastric cancer migration, invasion and proliferation by targeting RUNX3. Oncology reports. 2015; 34:1153-1161.
148. Zhao JJ, Lin J, Zhu D, Wang X, Brooks D, Chen M, Chu ZB, Takada K, Ciccarelli B, Admin S, Tao J, Tai YT, Treon S, Pinkus G, Kuo WP, Hideshima T, et al. miR-30-5p functions as a tumor suppressor and novel therapeutic tool by targeting the oncogenic Wnt/beta-catenin/BCL9 pathway. Cancer research. 2014; 74:1801-1813.
149. Ling XH, Chen ZY, Luo HW, Liu ZZ, Liang YK, Chen GX, Jiang FN and Zhong WD. BCL9, a coactivator for Wnt/beta-catenin transcription, is targeted by miR-30c and is associated with prostate cancer progression. Oncology letters. 2016; 11:2001-2008.
150. Zheng Z, Liu J, Yang Z, Wu L, Xie H, Jiang C, Lin B, Chen T, Xing C, Liu Z, Song P, Yin S, Zheng S and Zhou L. MicroRNA-452 promotes stem-like cells of hepatocellular carcinoma by inhibiting sox7 involving wnt/beta-catenin signaling pathway. Oncotarget. 2016; 7:28000-12. doi: 10.18632/oncotarget.8584.
151. Jiang W, Tian Y, Jiang S, Liu S, Zhao X and Tian D. MicroRNA-376c suppresses non-small-cell lung cancer cell growth and invasion by targeting LRH-1-mediated Wnt signaling pathway. Biochemical and biophysical research communications. 2016; 473:980-986.
152. Ling DJ, Chen ZS, Zhang YD, Liao QD, Feng JX, Zhang XY and Shi TS. MicroRNA-145 inhibits lung cancer cell metastasis. Molecular medicine reports. 2015; 11:3108-3114.
153. Xu W, Ji J, Xu Y, Liu Y, Shi L, Liu Y, Lu X, Zhao Y, Luo F, Wang B, Jiang R, Zhang J and Liu Q. MicroRNA-191, by promoting the EMT and increasing CSC-like properties, is involved in neoplastic and metastatic properties of transformed human bronchial epithelial cells. Molecular carcinogenesis. 2015; 54 Suppl 1:E148-161.
154. Hua HW, Jiang F, Huang Q, Liao Z and Ding G. MicroRNA-153 promotes Wnt/beta-catenin activation in hepatocellular carcinoma through suppression of WWOX. Oncotarget. 2015; 6:3840-3847. doi: 10.18632/oncotarget.2927.
155. Xu E, Zhao J, Ma J, Wang C, Zhang C, Jiang H, Cheng J, Gao R and Zhou X. miR-146b-5p promotes invasion and metastasis contributing to chemoresistance in osteosarcoma by targeting zinc and ring finger 3. Oncology reports. 2016; 35:275-283.
156. Zhang JX, Mai SJ, Huang XX, Wang FW, Liao YJ, Lin MC, Kung HF, Zeng YX and Xie D. MiR-29c mediates epithelial-to-mesenchymal transition in human colorectal carcinoma metastasis via PTP4A and GNA13 regulation of beta-catenin signaling. Annals of oncology. 2014; 25:2196-2204.
157. Gan X, Liu Z, Tong B and Zhou J. Epigenetic downregulated ITGBL1 promotes non-small cell lung cancer cell invasion through Wnt/PCP signaling. Tumour biology. 2016; 37:1663-1669.
158. Song Q, Xu Y, Yang C, Chen Z, Jia C, Chen J, Zhang Y, Lai P, Fan X, Zhou X, Lin J, Li M, Ma W, Luo S and Bai X. miR-483-5p promotes invasion and metastasis of lung adenocarcinoma by targeting RhoGDI1 and ALCAM. Cancer research. 2014; 74:3031-3042.
159. Song L, Liu D, Wang B, He J, Zhang S, Dai Z, Ma X and Wang X. miR-494 suppresses the progression of breast cancer in vitro by targeting CXCR4 through the Wnt/beta-catenin signaling pathway. Oncology reports. 2015; 34:525-531.
160. Dang YW, Zeng J, He RQ, Rong MH, Luo DZ and Chen G. Effects of miR-152 on cell growth inhibition, motility suppression and apoptosis induction in hepatocellular carcinoma cells. Asian Pacific journal of cancer prevention. 2014; 15:4969-4976.
161. Guo YH, Wang LQ, Li B, Xu H, Yang JH, Zheng LS, Yu P, Zhou AD, Zhang Y, Xie SJ, Liang ZR, Zhang CM, Zhou H and Qu LH. Wnt/beta-catenin pathway transactivates microRNA-150 that promotes EMT of colorectal cancer cells by suppressing CREB signaling. Oncotarget. 2016; 7:42513-42526. doi: 10.18632/oncotarget.9893.