
INTRODUCTION
Prostate cancer (PCa) recently became the second most frequently diagnosed malignant tumor, and the fifth leading cause of cancer death in men throughout the world [1]. Extensive evidence showed that obesity was implicated in the onset and progression of PCa [2, 3], and increasing researches were conducted to discover the connections between them. Of note, the aberrant secretion and signaling of various adipokines, including leptin and adiponectin, was proved to be one of the plausible mechanisms for carcinogenesis in obese man [4, 5].
Epidemiological evidence showed that high levels of serum leptin (LEP) were associated with onset and progression of various cancers [4, 6, 7]. In vitro studies revealed that leptin and its downstream signaling might induce cell proliferation [8], inhibit apoptosis [9], and mediate metabolism [10]. On the contrary, study showed that adiponectin (ADIPOQ) inhibited proliferation and invasion of PCa cells [11]. Moreover, the adiponectin level was proved lower in patients with metastatic PCa compared to those with organ-confined diseases [12]. Both leptin and adiponectin relied on binding to their receptors, leptin receptor (LEPR) or adiponectin receptor 1/2 (ADIPOR1/ADIPOR2), to activate downstream signaling and functioning [13, 14].
By modulating the levels and functioning of leptin and adiponectin, the genetic factors (single nucleotide polymorphisms and microsatellites in LEP, LEPR, ADIPOQ, ADIPOR1 and ADIPOR2) were demonstrated to affect the risk and aggressiveness of PCa [15–17]. Nevertheless, these studies were still insufficient and inconsistent. Therefore, we conducted an updated meta-analysis and pooled-review to better clarify the role of genetic variants in leptin, adiponectin and their receptors in PCa. The present study aimed to help developing better risk-assessment tools and targeted therapies in the field of PCa.
RESULTS
Study characteristics in the meta-analysis and pooled-review
As shown in Figure 1, a total of 133 published studies were identified from different database after initial search. We excluded 111 studies after title and abstract scanning, and enrolled two additional studies through manual reference searching. Then, a total of 24 studies were subject to full-text examination. Consequently, a total of 11 studies [18–28] were enrolled in the meta-analysis investigating the relationships between LEP/LEPR/ADIPOQ/ADIPOR1/ADIPOR2 variants and total PCa risk (Table 1), and ten studies [19, 23, 29–36] were included in the pooled-review discussing these polymorphisms’ impact on the aggressiveness of cancer (Table 2).

Figure 1: Flow chart of study selection and identification.
Table 1: Characteristics of studies included in the meta-analysis
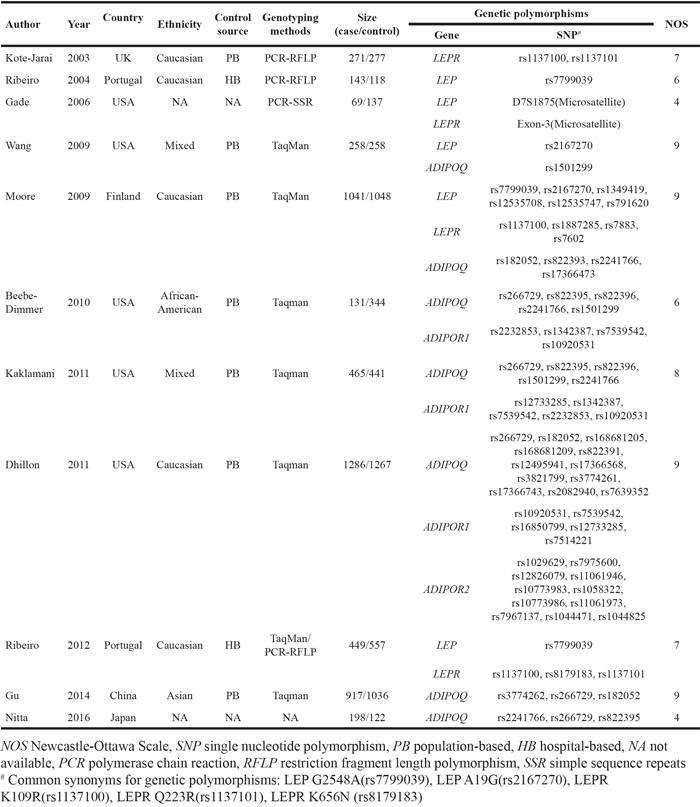
Table 2: Meta-analysis of multiple studies identifying the associations between LEP/LEPR/ADIPOQ/ADIPOR1 polymorphisms and prostate cancer risk
In the meta-analysis (Table 1), all studies were case-control design. Studies were conducted from 2003 to 2016. Among them, five were performed in USA, another two were performed in Portugal, and the rest four were performed in UK, Finland, China and Japan, respectively. The Newcastle-Ottawa Scale (NOS) scores ranged from 4 to 9. Polymorphisms with meta-analyzed statistics were presented in Table 2, and those with single supporting study were pooled in Table 3. The population number of case and control in each genetic variant and associated forest plots were also displayed (Supplemental Data S1, Supplemental Data S2). Besides, the genetic variants were found to be associated with PCa aggressiveness, e.g. pathological grade, castration resistance, recurrence and metastasis, and survival (Table 4).
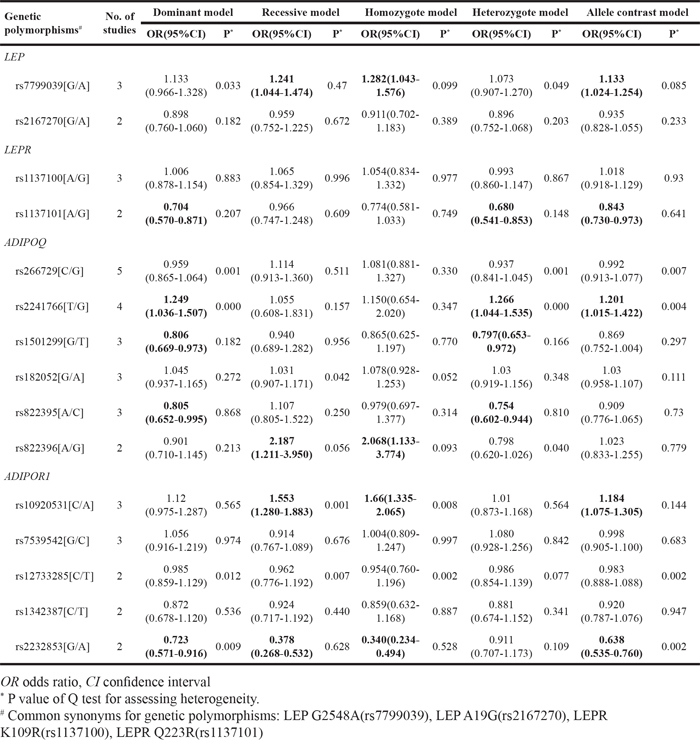
Table 3: Pooled-review of the studies investigating associations between LEP/LEPR/ADIPOQ/ADIPOR1/ADIPOR2 polymorphisms and prostate cancer risk
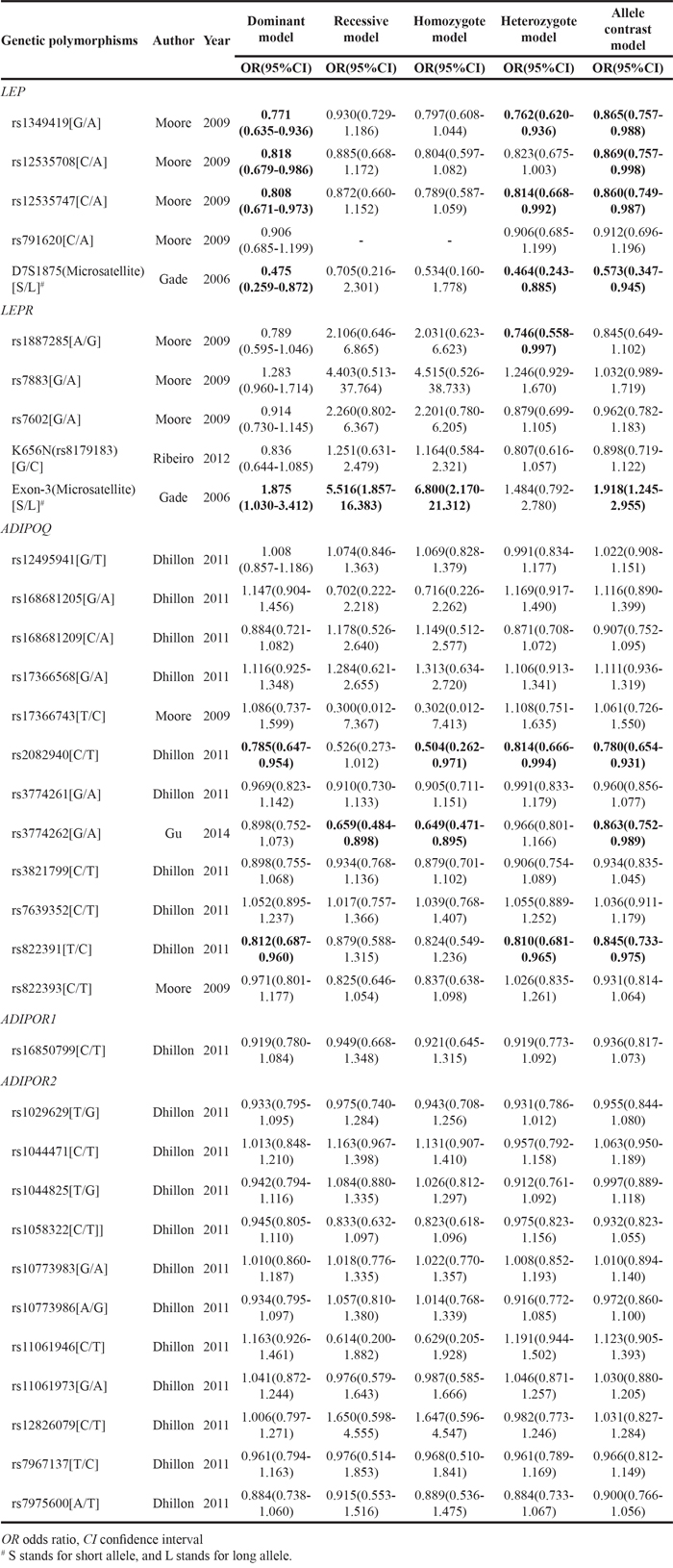
Table 4: Pooled-review of the studies investigating associations between genetic variants and prostate cancer aggressiveness
Genetic polymorphisms# |
Author |
Year |
Associations of minor allele of polymorphisms with PCa aggressiveness (unless otherwise stated) |
|---|---|---|---|
LEP |
|||
rs7799039[G/A] |
Ribeiro |
2004 |
Over represented in PCa patients with advanced disease (OR 1.91, 95%CI 1.24-2.59). |
rs10244329[A/T] |
Reese |
2010 |
Associated with PCa recurrence after definitive treatment (HR 0.49, 95%CI 0.28-0.84, P=0.010). |
LEPR |
|||
rs8179183[G/C] |
Monteiro |
2010 |
Associated with higher Gleason score (P=0.008). |
rs1137100[A/G] |
Monteiro |
2010 |
Lys carrier had lower time-to-bone metastasis in multivariate analysis (HR 0.37, 95% CI 0.14-0.95, P=0.039). |
Lin |
2011 |
Associated with a decrease in PCa specific mortality (HR 0.82, 95%CI 0.67-1.00, P=0.027) in the Swedish cohort. |
|
Kapustina |
2014 |
Distinguish PCa patients with high grade (Gleason score ≥7) or low grade (Gleason score <7) cancer. |
|
rs1137101[A/G] |
Monteiro |
2009 |
Associated with a higher Gleason score (P=0.022) and shorter time-to-relapse (P=0.006). |
Ribeiro |
2012 |
In multivariate model, AA was associated with high-grade PCa (Gleason score ≥7) (OR 1.56, 95%CI 1.15-2.12). |
|
ADIPOQ |
|||
rs1501299[G/T] |
Cunha |
2010 |
Associated with increased risk for higher Gleason score (OR 1.99, 95%CI 1.2-3.3, P=0.004), shorter time to hormonal castration resistance (TT vs. G, P=0.006). |
rs182052[G/A] |
Gu |
2015 |
Associated with increased risk of biochemical recurrence (AA vs GG, HR 2.44, 95%CI 1.57-3.79, P=6×10-5). |
ADIPOR2 |
|||
rs1044471[C/T] |
Stark |
2011 |
Associated with time to lethal PCa (CT vs CC, HR 0.6, 95%CI 0.4-0.9; TT vs CC, HR 0.8, 95%CI 0.6-1.2). |
OR odds ratio, CI confidence interval, PCa prostate cancer
# Common synonyms for genetic polymorphisms: LEP G2548A(rs7799039), LEPR K656N (rs8179183), LEPR K109R(rs1137100), LEPR Q223R(rs1137101)
LEP polymorphisms and PCa
A total of eight LEP polymorphisms were enrolled, with key information outlined in Figure 2. LEP rs7799039 A allele was correlated with higher risk of PCa (allele contrast: OR 1.133, 95%CI 1.024-1.254, Table 2), which was also over represented in advanced diseases (Table 4). On the contrary, the variants of LEP rs1349419 (allele contrast: OR 0.865, 95%CI 0.757-0.988), LEP rs12535708 (allele contrast: OR 0.869, 95%CI 0.757-0.998), LEP rs12535747 (allele contrast: OR 0.860, 95%CI 0.749-0.987) and LEP D7S1875 (allele contrast: OR 0.573, 95%CI 0.347-0.945) were associated with decreased risk of total PCa (Table 3). Besides, LEP rs10244329 variant was associated with lower recurrence rate after definitive treatment (Table 4).
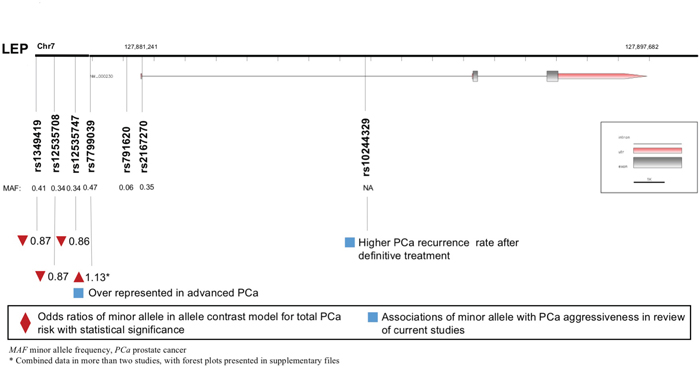
Figure 2: Diagram of LEP polymorphisms and prostate cancer.
LEPR polymorphisms and PCa
A total of eight LEPR polymorphisms were enrolled, with key data displayed in Figure 3. LEPR rs1137101 G allele was associated with lower PCa risk (allele contrast: OR 0.843, 95%CI 0.730-0.973, Table 2), while it was proved to be associated with worse pathological grade and prognosis (Table 4). LEPR Exon-3 long allele variant was proved to increase PCa risk (allele contrast: OR 1.918, 95%CI 1.245-2.955, Table 3). Besides, LEPR rs8179183 variant was associated with higher Gleason score, and LEPR rs1137100 polymorphism was related to both pathological grade and patient survival (Table 4).
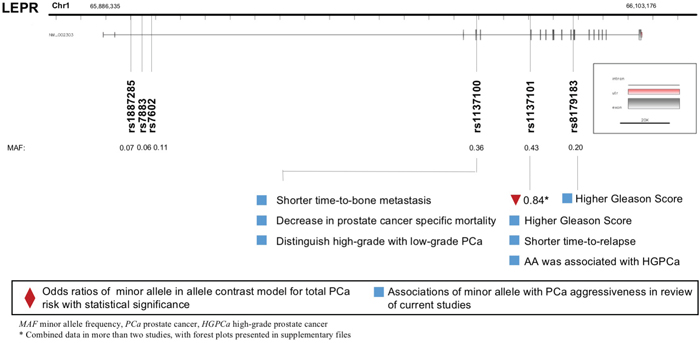
Figure 3: Diagram of LEPR polymorphisms and prostate cancer.
ADIPOQ polymorphisms and PCa
A total of 18 polymorphisms in gene ADIPOQ were enrolled, with key data displayed in Figure 4. ADIPOQ rs2241766 G allele was correlated with higher PCa risk (allele contrast: OR 1.201, 95%CI 1.015-1.422, Table 2). On the contrary, the variants of ADIPOQ rs2082940 (allele contrast: OR 0.780, 95%CI 0.654-0.931), ADIPOQ rs3774262 (allele contrast: OR 0.863, 95%CI 0.752-0.989) and ADIPOQ rs822391 (allele contrast: OR 0.845, 95%CI 0.733-0.975) were negatively associated with cancer risk (Table 3). ADIPOQ rs1501299 variant was reported to be associated with higher Gleason score and shorter time to castration resistance of PCa (Table 4). Besides, ADIPOQ rs182052 variant was associated with increased biochemical recurrence of PCa (Table 4).
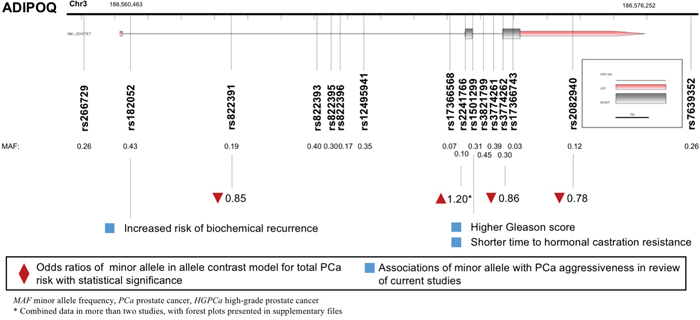
Figure 4: Diagram of ADIPOQ polymorphisms and prostate cancer.
ADIPOR1/ADIPOR2 polymorphisms and PCa
Both ADIPOR1 and ADIPOR2 encoded proteins that served as receptors for adiponectin. A total of six variants in gene ADIPOR1 and 11 variants in gene ADIPOR2 were enrolled and displayed in Figure 5. ADIPOR1 rs10920531 variant was correlated with higher PCa risk (allele contrast: OR 1.184, 95%CI 1.075-1.305, Table 2). However, a negative association was witnessed between ADIPOR1 rs2232853 variant and cancer risk (allele contrast: OR 0.638, 95%CI 0.535-0.760, Table 2). We failed to detect any associations between ADIPOR2 gene polymorphisms and risk of PCa. Meanwhile, ADIPOR2 rs1044471 variant was negatively associated with time to lethal PCa.
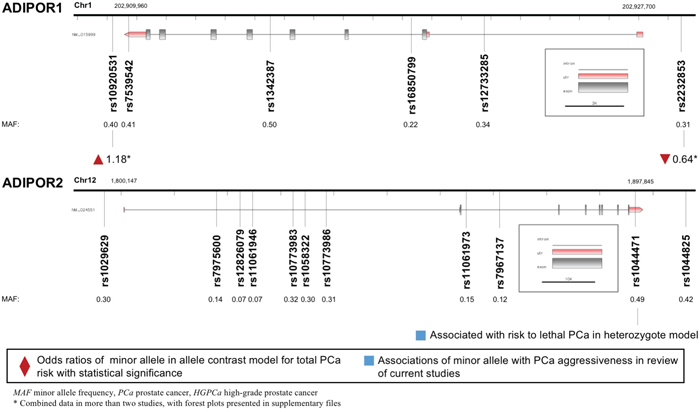
Figure 5: Diagram of ADIPOR1/ADIPOR2 polymorphisms and prostate cancer.
DISCUSSION
Various clinical and molecular studies were conducted to deliberate the associations among leptin, adiponectin and PCa, while controversies still existed and exact mechanisms remained largely unknown. From a genetic perspective, the present study conducted an updated meta-analysis and pooled-review. We enrolled 49 genetic variants in gene LEP, LEPR, ADIPOQ, ADIPOR1 and ADIPOR2, to investigate the relationships between these polymorphisms and PCa. The meta-analysis yielded five polymorphisms that associated with PCa risk according to allele contrast model. Meanwhile, the pooled-review outlined eight variants that affected cancer risk and another eight polymorphisms that affected cancer aggressiveness.
The LEP gene, encoded leptin that was associated with obesity and carcinogenesis [13]. In present study, we discovered that minor allele in LEP rs7799039 (G2548A) increased both the prevalence and aggressiveness of PCa. The increasing trend in cancer risk was in accordance with previous studies [37, 38]. The rs7799039 was located at promoter sequence of gene LEP. Several studies demonstrated that the variants might elevate the serum leptin level through transcriptional level, and increase human body weight as well [39, 40]. Higher leptin level, in turn, contributed to cancer growth and development via stimulating inflammation, angiogenesis, proliferation and epithelial-mesenchymal transition [10, 41].
Leptin exerted its downstream functioning through binding to leptin receptor (encoded by gene LEPR). Like LEP, LEPR was also highly polymorphic, with a number of single nucleotide polymorphisms identified. The present study revealed that LEPR rs1137101 (Q223R) variants were correlated with decreased PCa risk, whereas worse pathological grade and prognosis. The previous meta-analysis discovered that rs1137101 might increase the susceptibility to cancers in Asian and African patients, while decreasing the susceptibility in Caucasians, suggesting that race might explain for the heterogeneity and inconsistency among different studies [42]. On the other hand, the associations between rs1137101 and different cancers remained inconsistent [37, 42]. In the present study, the enrolled population were all Caucasians. Whether the negative associations with cancer risk contributed to Caucasian race or cancer type required further validation. It was well studied that rs1137101 mutation (A to G transversion) encoded a glutamine to arginine substitution in the N-terminal CRH1 domain [43], which was proved dispensable for a high affinity interaction with leptin binding [44]. However, the mutation might alter downstream signaling (e.g. STAT3, JAK2), surface expression, or receptor trafficking [44–46], which might partly underlie the mechanisms for worse pathological grade and prognosis of PCa in the present study.
ADIPOQ gene was also highly polymorphic, with at least 620 reported polymorphisms. Some of the polymorphisms were studied for their associations with cancer risk [47]. In the present study, we demonstrated that ADIPOQ rs2241766 variant was associated with increased risk of PCa. Of note, the outcome was opposite to those found in colorectal cancer, where the variant allele was identified as a protective factor against cancer [14, 47]. The discrepancy remained unknown. To our knowledge, the rs2241766 variant, a synonymous polymorphism at exon 2, might alter mRNA levels by regulation of mRNA splicing or stability [48]. Recently, a meta-analysis discovered that G allele of rs2241766 increased the risk of metabolic syndrome [49]. Meanwhile, metabolic syndrome was closely associated with cancer development and progression [50], which provided a probable mechanism underlying the present outcomes.
Adiponectin receptor 1 and 2 were often differentially expressed in cancers [14]. In the present study, we found no correlation among ADIPOR1 rs7539542, rs12733285, rs1342387 and risk of PCa, which was in line with the previous study [17]. Of note, we discovered that ADIPOR1 rs10920531 polymorphism was associated with increased risk of PCa, while ADIPOR1 rs2232853 variant decreased the risk of PCa. These two polymorphisms were firstly reported that were associated with cancer risk. Since the limitation of related studies, the outcomes required more validation and the underlying mechanisms demand further exploration.
Several limitations existed in the present study. First, the number of studies addressing each polymorphism was limited, and therefore confined us from conducting more meaningful subgroup analyses, e.g. race. Second, the baseline body mass index, serum leptin and adiponectin level were not reported in each study, which might bias the outcomes and constrain us to draw more conclusions. Consequently, more well designed studies with different population were warranted to better identify these complexities.
In conclusion, the present study comprehensively summarized the associations of genetic polymorphisms in LEP, LEPR, ADIPOQ, ADIPOR1 and ADIPOR2 with the risk and aggressiveness of PCa. We discovered that LEP rs7799039, ADIPOQ rs2241766 and ADIPOR1 rs10920531 variants were correlated with increased risk of PCa, while LEPR rs1137101 and ADIPOR1 rs2232853 variants were associated with decreased risk of PCa in the meta-analysis. From the pooled-review, we further identified eight variants associated with cancer risk and another eight variants associated with cancer aggressiveness, respectively. These observations provided rich evidence that leptin and adiponectin, along with their receptors, were involved in the development and progression of PCa. We expected to help prediction of prevalence and prognosis of PCa, as well as generation of novel targeted therapy, especially in obese patients with impaired leptin and adiponectin signaling.
MATERIALS AND METHODS
Search strategy
We conducted a comprehensive literature search in PubMed and EMBASE database up to May 1st, 2016. Searching strategy involved “prostatic neoplasms OR prostatic cancer OR prostate cancer OR PCa”, “polymorphism* OR variant* OR SNP”, “leptin OR LEP OR LEPR OR adiponectin OR ADIPOQ OR ADIPOR*” and various combinations of these terms. The search was conducted without limitations of language. Reviews, meta-analyses, original articles, and other studies of interest were further examined to identify additional eligible studies.
Selection criteria
The eligible studies were included according to the following criteria: (1) studies investigating the associations between LEP/LEPR/ADIPOQ/ADIPOR1/ADIPOR2 polymorphisms and PCa risk or aggressiveness; (2) case-control or cohort studies; (3) sufficient data, including the accurate population number of specific alleles, genotypes and microsatellites; (4) genotype frequencies in control groups should be examined by the Hardy-Weinberg equilibrium (HWE). If serial studies of the same study population were reported, the most recent study or the study with the largest population should be adopted. The exclusion criteria included: (1) animal or molecular studies; (2) reviews, meta-analyses or case reports; (3) insufficient information published in the study; (4) duplicated studies.
Data extraction and quality assessment
Two investigators (Hu and Xu) independently evaluated the eligibility of all retrieved studies, and extracted key information. The collected data included: author’s name, year of publication, study population, study design, genotyping methods, case and control population, and polymorphisms investigated. NOS was adopted to evaluate the quality of all eligible studies [51]. The nine-star NOS was based on three dimensions: study group selection, comparability of cases and controls, and exposure of cases and controls. The disagreements during data extraction and quality assessment were resolved by consensus.
Meta-analysis and pooled review
In the meta-analysis, the pooled odds ratio (OR) with 95% confidence interval (95%CI) was calculated to evaluate the correlation between genetic polymorphisms and total PCa risk according to allele contrast, homozygote, heterozygote, dominant and recessive models. The inter-study heterogeneity was verified by Q and I2 statistics. Fixed-effect model was adopted to estimate the OR and 95%CI when heterogeneity was low, otherwise random-effect model was adopted. Besides, we conducted a pooled-review for studies discussing risk or aggressiveness of PCa that could not be included in the meta-analysis. All analyses were performed using the STATA/SE 12.0 software (StataCorp, College Station, TX, USA). Statistical significance was defined as a two-tailed p-value <0.05.
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest.
GRANT SUPPORT
This study was funded by National Natural Science Foundation of China (81272835), Outstanding Talents Cultivation Project of Shanghai Medical System (XBR2013092), Talents Development Foundation of Shanghai (201302) and Scientific Research Innovation Project of Shanghai Education Committee (14ZZ010). The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
REFERENCES
1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015; 65: 87–108. doi: 10.3322/caac.21262.
2. Hu M-B, Liu S-H, Jiang H-W, Bai P-D, Ding Q. Obesity Affects the Biopsy-Mediated Detection of Prostate Cancer, Particularly High-Grade Prostate Cancer: A Dose-Response Meta-Analysis of 29,464 Patients. Culig Z, editor. PLoS ONE. Public Library of Science; 2014; 9: e106677–10. doi: 10.1371/journal.pone.0106677.
3. Hu M-B, Xu H, Bai P-D, Jiang H-W, Ding Q. Obesity has multifaceted impact on biochemical recurrence of prostate cancer: a dose–response meta-analysis of 36,927 patients. Med Oncol. 2014; 31: 829–9. doi: 10.1007/s12032-013-0829-8.
4. Kato S, Abarzua-Catalan L, Trigo C, Delpiano A, Sanhueza C, García K, Ibañez C, Hormazábal K, Diaz D, Brañes J, Castellón E, Bravo E, Owen G, et al. Leptin stimulates migration and invasion and maintains cancer stem-like properties in ovarian cancer cells: an explanation for poor outcomes in obese women. Oncotarget. 2015; 6: 21100–19. doi: 10.18632/oncotarget.4228.
5. Mistry T, Digby JE, Desai KM, Randeva HS. Obesity and prostate cancer: a role for adipokines. Eur Urol. 2007; 52: 46–53. doi: 10.1016/j.eururo.2007.03.054.
6. Oba J, Wei W, Gershenwald JE, Johnson MM, Wyatt CM, Ellerhorst JA, Grimm EA. Elevated Serum Leptin Levels are Associated With an Increased Risk of Sentinel Lymph Node Metastasis in Cutaneous Melanoma. Medicine (Baltimore). 2016; 95: e3073. doi: 10.1097/MD.0000000000003073.
7. Alshaker H, Krell J, Frampton AE, Waxman J, Blyuss O, Zaikin A, Winkler M, Stebbing J, Yagüe E, Pchejetski D. Leptin induces upregulation of sphingosine kinase 1 in oestrogen receptor-negative breast cancer via Src family kinase-mediated, janus kinase 2-independent pathway. Breast Cancer Res. BioMed Central; 2014; 16: 426. doi: 10.1186/s13058-014-0426-6.
8. Andò S, Barone I, Giordano C, Bonofiglio D, Catalano S. The Multifaceted Mechanism of Leptin Signaling within Tumor Microenvironment in Driving Breast Cancer Growth and Progression. Front Oncol. Frontiers; 2014; 4: 340. doi: 10.3389/fonc.2014.00340.
9. Dubois V, Jardé T, Delort L, Billard H, Bernard-Gallon D, Berger E, Geloen A, Vasson M-P, Caldefie-Chezet F. Leptin induces a proliferative response in breast cancer cells but not in normal breast cells. Nutr Cancer. 2014; 66: 645–55. doi: 10.1080/01635581.2014.894104.
10. Vansaun MN. Molecular pathways: adiponectin and leptin signaling in cancer. Clin Cancer Res. American Association for Cancer Research; 2013; 19: 1926–32. doi: 10.1158/1078-0432.CCR-12-0930.
11. Tan W, Wang L, Ma Q, Qi M, Lu N, Zhang L, Han B. Adiponectin as a potential tumor suppressor inhibiting epithelial-to-mesenchymal transition but frequently silenced in prostate cancer by promoter methylation. Prostate. 2015; 75: 1197–205. doi: 10.1002/pros.23002.
12. Allott EH, Masko EM, Freedland SJ. Obesity and prostate cancer: weighing the evidence. Eur Urol. 2013; 63: 800–9. doi: 10.1016/j.eururo.2012.11.013.
13. Alshaker H, Sacco K, Alfraidi A, Muhammad A, Winkler M, Pchejetski D. Leptin signalling, obesity and prostate cancer: molecular and clinical perspective on the old dilemma. Oncotarget. 2015; 6: 35556–63. doi: 10.18632/oncotarget.5574.
14. Katira A, Tan PH. Adiponectin and its receptor signaling: an anti-cancer therapeutic target and its implications for anti-tumor immunity. Expert Opin Ther Targets. 2015; 19: 1105–25. doi: 10.1517/14728222.2015.1035710.
15. Liu Y. Genetic association between leptin-2548G/A polymorphism and risk of cancer: A meta analysis. International Journal of Clinical and Experimental Medicine. 2015; 8: 448–55.
16. Liu P, Shi H, Huang C, Shu H, Liu R, Yang Y, Gong J, Yang Y, Cai M. Association of LEP A19G polymorphism with cancer risk: a systematic review and pooled analysis. Tumour Biol. Springer Netherlands; 2014; 35: 8133–41. doi: 10.1007/s13277-014-2088-5.
17. Ye J, Jiang L, Wu C, Liu A, Mao S, Ge L. Three ADIPOR1 Polymorphisms and Cancer Risk: A Meta-Analysis of Case-Control Studies. Devaney J, editor. PLoS ONE. Public Library of Science; 2015; 10: e0127253. doi: 10.1371/journal.pone.0127253.
18. Kote-Jarai Z, Singh R, Durocher F, Easton D, Edwards SM, Ardern-Jones A, Dearnaley DP, Houlston R, Kirby R, Eeles R. Association between leptin receptor gene polymorphisms and early-onset prostate cancer. BJU Int. 2003; 92: 109–12.
19. Ribeiro R, Vasconcelos A, Costa S, Pinto D, Morais A, Oliveira J, Lobo F, Lopes C, Medeiros R. Overexpressing leptin genetic polymorphism (-2548 G/A) is associated with susceptibility to prostate cancer and risk of advanced disease. Prostate. Wiley Subscription Services, Inc., A Wiley Company; 2004; 59: 268–74. doi: 10.1002/pros.20004.
20. Gade-Andavolu R, Cone LA, Shu S, Morrow A, Kowshik B, Andavolu MVS. Molecular interactions of leptin and prostate cancer. Cancer J. 2006; 12: 201–6.
21. Moore SC, Leitzmann MF, Albanes D, Weinstein SJ, Snyder K, Virtamo J, Ahn J, Mayne ST, Yu H, Peters U, Gunter MJ. Adipokine genes and prostate cancer risk. Int J Cancer. 2009; 124: 869–76. doi: 10.1002/ijc.24043.
22. Wang M-H, Helzlsouer KJ, Smith MW, Hoffman-Bolton JA, Clipp SL, Grinberg V, De Marzo AM, Isaacs WB, Drake CG, Shugart YY, Platz EA. Association of IL10 and other immune response- and obesity-related genes with prostate cancer in CLUE II. Prostate. 2009; 69: 874–85. doi: 10.1002/pros.20933.
23. Ribeiro RJT, Monteiro CPD, Azevedo ASM, Cunha VFM, Ramanakumar AV, Fraga AM, Pina FM, Lopes CMS, Medeiros RM, Franco EL. Performance of an adipokine pathway-based multilocus genetic risk score for prostate cancer risk prediction. Scheurer M, editor. PLoS ONE. Public Library of Science; 2012; 7: e39236. doi: 10.1371/journal.pone.0039236.
24. Beebe-Dimmer JL, Zuhlke KA, Ray AM, Lange EM, Cooney KA. Genetic variation in adiponectin (ADIPOQ) and the type 1 receptor (ADIPOR1), obesity and prostate cancer in African Americans. Prostate Cancer Prostatic Dis. Nature Publishing Group; 2010; 13: 362–8. doi: 10.1038/pcan.2010.27.
25. Kaklamani V, Yi N, Zhang K, Sadim M, Offit K, Oddoux C, Ostrer H, Mantzoros C, Pasche B. Polymorphisms of ADIPOQ and ADIPOR1 and prostate cancer risk. Metab Clin Exp. Elsevier; 2011; 60: 1234–43. doi: 10.1016/j.metabol.2011.01.005.
26. Dhillon PK, Penney KL, Schumacher F, Rider JR, Sesso HD, Pollak M, Fiorentino M, Finn S, Loda M, Rifai N, Mucci LA, Giovannucci E, Stampfer MJ, et al. Common polymorphisms in the adiponectin and its receptor genes, adiponectin levels and the risk of prostate cancer. Cancer Epidemiol Biomarkers Prev. American Association for Cancer Research; 2011; 20: 2618–27. doi: 10.1158/1055-9965.EPI-11-0434.
27. Gu C-Y, Li Q-X, Zhu Y, Wang M-Y, Shi T-Y, Yang Y-Y, Wang J-C, Jin L, Wei Q-Y, Ye D-W. Genetic variations of the ADIPOQgene and risk of prostate cancer in Chinese Han men. Asian Journal of Andrology. Medknow Publications; 2014; 16: 878–83. doi: 10.4103/1008-682X.129939.
28. Nitta T, Koike H, Mastui H, Miyao T, Kurihara S, Miyazawa Y, Kato H, Sekine Y, Shibata Y, Ito K, Suzuki K. MP04-03 ROLE OF ADIPONECTIN IN PROSTATE CANCER RISK AND THE PROLIFERATION OF PROSTATE CANCER. The Journal of Urology. 2016; 195: e28. doi: 10.1016/j.juro.2016.02.1938.
29. Reese A, Cheng I, Plummer S, Mefford J, Casey G, Witte J. GENETIC VARIANTS IN LEPTIN AND PPARγ INFLUENCE DEVELOPMENT AND RECURRENCE OF ADVANCED PROSTATE CANCER. The Journal of Urology. Elsevier; 2010; 183: e830. doi: 10.1016/j.juro.2010.02.2223.
30. Monteiro C, Ribeiro R, Azevedo A, Cunha V, Fraga A, Pina F, Calais da Silva F, Lobo F, Medeiros R. Non-synonym leptin receptor genetic variants, prostate cancer susceptibility and aggressiveness. European Journal of Cancer Supplements. Elsevier; 2010; 8: 25–6. doi: 10.1016/S1359-6349(10)70905-8.
31. Lin DW, FitzGerald LM, Fu R, Kwon EM, Zheng SL, Kolb S, Wiklund F, Stattin P, Isaacs WB, Xu J, Ostrander EA, Feng Z, Grönberg H, et al. Genetic variants in the LEPR, CRY1, RNASEL, IL4, and ARVCF genes are prognostic markers of prostate cancer-specific mortality. Cancer Epidemiol Biomarkers Prev. 2011; 20: 1928–36. doi: 10.1158/1055-9965.EPI-11-0236.
32. Kapustina Ž, Valaitienė G, Daniūnaitė K, Jarmalaitė S, Jankevičius F, Laurinavičius A, Lazutka JR. Analysis of single nucleotide polymorphisms in RNASEL, LEPR, CRY1, IL4 and CHI3L2 genes in a cohort of Lithuanian prostate cancer and benign prostatic hyperplasia patients. European Urology Supplements. 2014; 13: e1149. doi: 10.1016/S1569-9056(14)50003-6.
33. Monteiro C, Ribeiro R, Azevedo A, Cunha V, Francisco N, Fraga A, Pina F, Calais-da-Silva E, Lobo F, Medeiros R. Leptin receptor genetic variants are associated with prostate cancer development, aggressiveness and the time to biochemical relapse. European Journal of Cancer Supplements. 2009; 7: 412–3. doi: 10.1016/S1359-6349(09)71399-0.
34. Cunha V, Ribeiro R, Azevedo A, Monteiro C, Pina F, Fraga A, Calais da Silva F, Lobo F, Medeiros R. 96 Adiponectin functional polymorphisms and haplotype are associated with prostate cancer aggressiveness and to hormonal castration resistance. European Journal of Cancer Supplements. Elsevier; 2010; 8: 25. doi: 10.1016/S1359-6349(10)70904-6.
35. Gu C, Qu Y, Zhang G, Sun L, Zhu Y, Ye D. A single nucleotide polymorphism in ADIPOQ predicts biochemical recurrence after radical prostatectomy in localized prostate cancer. Oncotarget. 2015; 6: 32205–11. doi: 10.18632/oncotarget.4980.
36. Stark JR, Finn SP, Ma J, Sinnott JA, Schumacher F, Lis R, Penney KL, Kasperzyk JL, Sesso HD, Stampfer MJ, Giovannucci EL, Loda M, Mucci LA. Adiponectin receptor 2 expression predicts lethal prostate cancer. Lab Invest. 2011; 91: 226A.
37. He J, Xi B, Ruiter R, Shi T-Y, Zhu M-L, Wang M-Y, Li Q-X, Zhou X-Y, Qiu L-X, Wei Q-Y. Association of LEP G2548A and LEPR Q223R polymorphisms with cancer susceptibility: evidence from a meta-analysis. Huang Q, editor. PLoS ONE. 2013; 8: e75135–12. doi: 10.1371/journal.pone.0075135.
38. Yang Y, Liu P, Guo F, Liu R, Yang Y, Huang C, Shu H, Gong J, Cai M. Genetic G2548A polymorphism of leptin gene and risk of cancer: a meta-analysis of 6860 cases and 7956 controls. J BUON. 2014; 19: 1096–104.
39. Shabana, Hasnain S. Leptin promoter variant G2548A is associated with serum leptin and HDL-C levels in a case control observational study in association with obesity in a Pakistani cohort. J Biosci. 2016; 41: 251–5.
40. Boumaiza I, Omezzine A, Rejeb J, Rebhi L, Ouedrani A, Ben Rejeb N, Nabli N, Ben Abdelaziz A, Bouslama A. Relationship between leptin G2548A and leptin receptor Q223R gene polymorphisms and obesity and metabolic syndrome risk in Tunisian volunteers. Genet Test Mol Biomarkers. Mary Ann Liebert, Inc. 140 Huguenot Street, 3rd Floor New Rochelle, NY 10801 USA; 2012; 16: 726–33. doi: 10.1089/gtmb.2011.0324.
41. Noda T, Kikugawa T, Tanji N, Miura N, Asai S, Higashiyama S, Yokoyama M. Long-term exposure to leptin enhances the growth of prostate cancer cells. Int J Oncol. Spandidos Publications; 2015; 46: 1535–42. doi: 10.3892/ijo.2015.2845.
42. Liu P, Shi H, Liu R, Yang Y, Yang Y, Huang C, Shu H, Gong J, Cai M. Lack of association between LEPR Q223R polymorphisms and cancer susceptibility: evidence from a meta-analysis. J BUON. 2014; 19: 855–62.
43. Stratigopoulos G, LeDuc CA, Matsuoka N, Gutman R, Rausch R, Robertson SA, Myers MG, Chung WK, Chua SC, Leibel RL. Functional consequences of the human leptin receptor (LEPR) Q223R transversion. Obesity (Silver Spring). Blackwell Publishing Ltd; 2009; 17: 126–35. doi: 10.1038/oby.2008.489.
44. Peelman F, Van Beneden K, Zabeau L, Iserentant H, Ulrichts P, Defeau D, Verhee A, Catteeuw D, Elewaut D, Tavernier J. Mapping of the leptin binding sites and design of a leptin antagonist. Journal of Biological Chemistry. American Society for Biochemistry and Molecular Biology; 2004; 279: 41038–46. doi: 10.1074/jbc.M404962200.
45. Kimber W, Peelman F, Prieur X, Wangensteen T, O'Rahilly S, Tavernier J, Farooqi IS. Functional characterization of naturally occurring pathogenic mutations in the human leptin receptor. Endocrinology. Endocrine Society; 2008; 149: 6043–52. doi: 10.1210/en.2008-0544.
46. Zabeau L, Defeau D, Van der Heyden J, Iserentant H, Vandekerckhove J, Tavernier J. Functional analysis of leptin receptor activation using a Janus kinase/signal transducer and activator of transcription complementation assay. Mol Endocrinol. Endocrine Society; 2004; 18: 150–61. doi: 10.1210/me.2003-0078.
47. Li Q, Ma Y, Sang W, Cui W, Li X, Liu X, Zhang W. Five common haplotype-tagging variants of adiponectin (ADIPOQ) and cancer susceptibility: a meta-analysis. Genet Test Mol Biomarkers. Mary Ann Liebert, Inc. 140 Huguenot Street, 3rd Floor New Rochelle, NY 10801 USA; 2014; 18: 417–24. doi: 10.1089/gtmb.2013.0493.
48. Mahmoudi T, Karimi K, Karimi N, Farahani H, Nobakht H, Dabiri R, Vahedi M, Zali MR. Association of adiponectin receptor 1 gene - 106 C > T variant with susceptibility to colorectal cancer. Meta Gene. 2016; 9: 210–4. doi: 10.1016/j.mgene.2016.07.008.
49. Zhou JM, Zhang M, Wang S, Wang BY, Han CY, Ren YC, Zhang L, Zhang HY, Yang XY, Zhao Y, Hu DS. Association of the ADIPOQ Rs2241766 and Rs266729 Polymorphisms with Metabolic Syndrome in the Chinese Population: A Meta-analysis. Biomed Environ Sci. 2016; 29: 505–15. doi: 10.3967/bes2016.066.
50. Rhee H, Vela I, Chung E. Metabolic Syndrome and Prostate Cancer: a Review of Complex Interplay Amongst Various Endocrine Factors in the Pathophysiology and Progression of Prostate Cancer. Horm Cancer. 2016; 7: 75–83. doi: 10.1007/s12672-015-0238-x.
51. Stang A. Critical evaluation of the Newcastle-Ottawa scale for the assessment of the quality of nonrandomized studies in meta-analyses. Eur J Epidemiol. Springer Netherlands; 2010; 25: 603–5. doi: 10.1007/s10654-010-9491-z.