
Introduction
Antibiotic resistance (referred to bacterial drug resistance here) has already developed into a leading problem in global public health [1]. The notorious New Delhi β-lactamase 1 (NDM-1)-producing super-bugs that was initially found in India, Pakistan and the UK [2] had ever pushed us on the cusp of post-antibiotics era. The prevalence of NDM-1 in the Gram-negative Enterobactereae including E. coli and Klebsiella pneumoniae (K. pneumoniae) confers the robust resistance to carbapenems and the expanded-spectrum (or third-generation) cephalosporins, the two extensively-used antibiotics for treatments of multidrug-resistant bacteria [3, 4]. The polymyxin E (colistin), a family of cationic polypeptide antibiotics, might represent the last line of defense against lethal infections by Gram-negative pathogens with pan-drug resistance [1]. However, it seems likely that this ultimate line of refuge antibiotics (polymyxin), has been challenged by the emergence of colistin resistance mechanisms [1, 5]. Among them, one refers to the chromosome-encoded machinery (two sets of bacterial two-component systems [pmrAB [6] and phoPQ [7]) and the regulator mgrB [7] are implicated in K. pneumoniae [7], and the other denotes the plasmid-mobilized colistin resistance (MCR-1) in certain species of Enterobacteriaceae, e.g.: E. coli [8]. The above two mechanisms are responsible for chemical modification of the lipid A on bacterial LPS, which consequently resulting in the reduced affinity for the polymyxin [8].
The mcr-1 gene defines a newly-emerging mechanism for plasmid-mediated transferable colistin resistance [8]. The mcr-1 protein product, MCR-1, is predicted to be an integral membrane protein with the catalytic activity of phosphoethanolamine transferases [9]. The MCR-1 enzyme modifies the chemical structure of lipid A moiety on bacterial LPS by the addition of phosphoethanolamine, which in turn reduces the binding affinity to colistin (i.e., producing the colistin resistance) [8, 9]. Since the first discovery of MCR-1 from Southern China, in the late of 2015, the new colistin resistance gene has spread to 5 of 7 continents [10]. A retrospective study by Shen et al. [11] reported that the mcr-1 gene is detected in three chicken E. coli isolates derived from the 1980s, when colisitin first started to be used in food-producing animals in China. It suggested that the emergence of the MCR-1 is much earlier than we anticipated [11]. To the best of our knowledge, no less than six species of Enterobacteriaceae (E. coli [9, 12-14], Enterobacter aerogenes [15], Enterobacter cloacae [15], K. pneumonia [16-18], Salmonella enterica [19-21] , and Shigella somnei [22] are recipient hosts for the mcr-1-harbouring plasmids [10]. In particular, the MCR-1-producing plasmids display unexpected diversity, indicating the complexity of the MCR-1 dissemination mechanism [9]. The whole genome sequences of diversified mcr-1-carrying plasmids allowed us to better understand the mechanisms for the origin, evolution, transfer and dissemination of the mcr-1 colistin resistance [13, 23-25].
Given the fact that 1) Guangdong province of China is the first place where the mcr-1 gene was discovered [8]; 2) We observed that unexpected diversity in the mcr-1-harbouring plasmid reservoirs is present in the gut microbiota from the diarrhea patients in Shenzhen city localized in the same province Guangdong [9]; 3) Guangdong is one of the largest province for pig production in China, we therefore attempted to address the possibility whether complex dissemination of the MCR-1 colistin resistance by the diversified mcr-1-bearing plasmids is widespread in E. coli with different sequence types from healthy pig populations. In this paper, we report that this is the case.
Results and Discussion
Occurrence of the mcr-1-positive E. coli isolates
In total, the bacterial samples were from three cities (and/or counties) of Guangdong Province (Figure 1A), which correspond to Yingde City, Huizhou City and Huidong County, respectively (Figure 1B). Each city hosted numbers of pig farms. In principle, nasal fluid and fecal samples were collected from piglets, fattening pigs and sows. The enteric bacteria were selectively screened on the MacConkey Agar plates, and the resultant single colonies were further subjected to the propagation in the liquid LB media (not shown). Subsequently, we conducted PCR screen for the presence of the mcr-1 gene amongst the bacterial species. As expected, the mcr-1-positive isolates were found in the above three cities/counties (Figure 1C). Direct DNA sequencing results revealed that the mcr-1 gene from hundreds of bacterial isolates is in 100% identity. Unlike the mcr-1.2, a variant (Q3L) of the mcr-1 gene [16], we indeed failed to observe any alleic variants. 16S rDNA-based phylogenetic analyses proved that all the mcr-1-positive isolates are E. coli (not shown). As a result, 302 of 1026 E. coli isolates from the 3 distant pig farms were confirmed to be positive for the mcr-1 gene in our trials. It suggested that the average positive rate of mcr-1 is around 30% in these pig-producing places. Also, 15 representative mcr-1-positive clinical isolates were applied for further functional tests using the LBA plates supplemented with colistin at various levels. Consistent with our recent observation with the human clinical isolates that produce MCR-1 [9], they consistently exhibited the appreciable level of colistin resistance in that the minimum inhibitory concentration (MIC) is up to 32 mg/L (not shown).
Table 1: Strains used in this study
Strains or plasmids |
Relevant characteristics |
Origin |
Strains |
||
MC1061 |
Wild type of E. coli K-12, Δlac |
[32] |
DH5a (λ-pir) |
Δlac host for pAH125 and its derivatives |
[32] |
GD97 |
E. coli carrying pGD97, the mcr-1-harbouring plasmid |
[12] |
WH13 |
E. coli carrying pWH13, the mcr-1-harbouring plasmid |
[12] |
E15017 |
E. coli carrying pE15017, the mcr-1-harbouring plasmid |
[9] |
A31-12 |
E. coli carrying the mcr-1-harbouring plasmid pA31-12 |
[25] |
MG1655 |
The wild type K-12 strain of E. coli |
Lab stock |
FYJ795 |
MG1655 carrying pBAD24::mcr-1 |
[9] |
FYJ796 |
MG1655 carrying pBAD24 |
[9] |
FYJ158 |
DH5α (λ-pir) carrying pAH-PfadD |
[31] |
FYJ846 |
DH5α (λ-pir) carrying pAH-Pmcr-1(pGD97) |
This work |
FYJ847 |
MC1061 with Pmcr-1_pWH09-lacZ transcriptional fusion at the chromosomal attB λ site |
This work |
FYJ848 |
DH5α (λ-pir) carrying pAH125-Pmcr-1(pWH13) |
This work |
FYJ849 |
MC1061 with Pmcr-1_pWH13-lacZ transcriptional fusion at the chromosomal attB λ site |
This work |
FYJ850 |
DH5α (λ-pir) carrying pAH125-Pmcr-1 (pE15017) |
This work |
FYJ851 |
MC1061 with Pmcr-1_pE15017-lacZ transcriptional fusion at the chromosomal attB λ site |
This work |
FYJ852 |
DH5α (λ-pir) carrying pAH125-mcr-1 (pA31-12) |
This work |
FYJ853 |
MC1061 with Pmcr-1_pA31-12-lacZ transcriptional fusion at the chromosomal attB λ site |
This work |
Plasmids |
||
pAH-PfadD |
pAH125 carrying the fadD promoter region, KanR |
[31] |
pAH-Pmcr-1(pGD97) |
pAH125 carrying the mcr-1 promoter region from pGD97), KanR |
This work |
pAH-Pmcr-1(pWH13) |
pAH125 carrying the mcr-1 promoter region from pWH13, KanR |
This work |
pAH-Pmcr-1(pE15017) |
pAH125 carrying the mcr-1 promoter region from pE15017, KanR |
This work |
pAH-Pmcr-1(pA31-12) |
pAH125 carrying the mcr-1 promoter region from pA31-12, KanR |
This work |
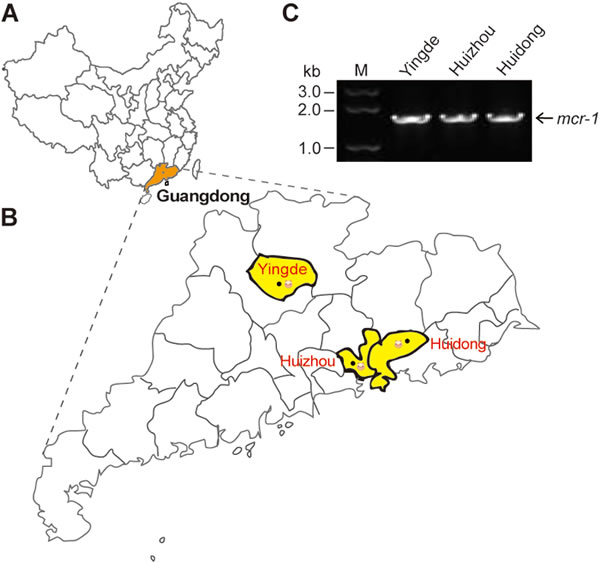
Figure 1: Distribution of the mcr-1 colistin resistance gene in Guangdong province. A. Location of Guangdong Province in China. Guangdong province is highlighted in orange. B. Locations of mcr-1-positive isolates from the swine gut microbiota in Guangdong Province. Locations are highlighted in yellow. C. The mcr-1-based screening of the isolates from the swine gut microbiota. The map is modified using adobe illustrator
Diversity in the mcr-1-harbouring plasmids
Given the fact the mcr-1 gene can be surrounded with different genetic environment of diversified plasmid backbones in the case of human clinical E. coli isolates [9], we hypothesized that similar scenario could already be present in the animal isolates in the healthy swine populations (Figure 2). To address this hypothesis, hundreds of mcr-1-carrying E. coli isolates were subjected to extensive analyses using the multiplex-PCR assays coupled with Sanger sequencing (Figure 3). In terms of the paradigm mcr-1-harbouring plasmid pHNSHP45 with known genome sequence, seven pairs of specific primers (Table 2) that target seven interested genes (including mcr-1 and nikB) were used in PCR assays to determine the various mcr-1-surrounding genetic environment (Figures 2 and 3). In addition to the mcr-1 gene (Figure 3C), the remaining six genes corresponded to nikB (Figure 3A), tnpA (Figure 3B), hp (Figure 3D), pilP (Figure 3E), virD4 (Figure 3F), and virB4 (Figure 3G), respectively. The result of PCR assays coupled with Sanger sequencing revealed unexpected diversity of the mcr-1-harbouring plasmids (Figures 2 and 3), which is much more complicated than the scenarios seen in the human and animal E. coli isolates [9, 26].
Table 2: Primers used in this study
Primers |
Primer sequence |
16S-F |
5'-AAA TTG AAG AGT TTG ATC ATG G-3' |
16S-R |
5'-GCT TCT TTA AGG TAA GGA GGT-3' |
mcr-1-F |
5'-ATG ATG CAG CAT ACT TCT GTG-3' |
mcr-1-R |
5'-TCA GCG GAT GAA TGC GGT G-3' |
nikB-F |
5'-GAT GAA CTT GAT CAT CGT GTT GT-3' |
nikB-R |
5'-GTA ATT CTG ACG AAA AAG AGG A-3' |
pilP-F |
5'-TTA AAG AAT AAG CTG GCG TTT C-3' |
pilP-R |
5'-ATG TTA AAA ATA ATT AAA CCA ACG-3' |
virD4-F |
5'-AAT GTC AAC ATG ATT GTT AC-3' |
virD4-R |
5'-GAA CAT AAC CCG GAC CTG AAA T-3' |
virB4-F |
5'-AAC TCT TTT TCA GTA AGC CCA AT-3' |
virB4-R |
5'-TTA ATG TTT GTT GTG GAT TAC AAC C-3' |
tnpA-F |
5'-GGT TTT CGG GCT TTT TAA GAG-3' |
tnpA-R |
5'-TAG CAC ATA GCG ATA CGA TG-3' |
hp-F |
5'-GAT AAG CAA ACT GGC ATC ACG-3' |
hp-R |
5'-GAA CCC TGT ATA TAG CCT GTC-3' |
adk-F |
5'-ATT CTG CTT GGC GCT CCG GG-3' |
adk-R |
5'-CCG TCA ACT TTC GCG TAT TT-3' |
fumC-F |
5'-TCA CAG GTC GCC AGC GCT TC-3' |
fumC-R |
5'-GTA CGC AGC GAA AAA GAT TC-3' |
gyrB-F |
5'-TCG GCG ACA CGG ATG ACG GC-3' |
gyrB-R |
5'-ATC AGG CCT TCA CGC GCA TC-3' |
icd-F |
5'-ATG GAA AGT AAA GTA GTT GTT CCG GCA CA-3' |
icd-R |
5'-GGA CGC AGC AGG ATC TGT T-3' |
mdh-F |
5'-ATG AAA GTC GCA GTC CTC GGC GCT GCT GGC GG-3' |
mdh-R |
5'-TTA ACG AAC TCC TGC CCC AGA GCG ATA TCT TTC TT-3' |
purA-F |
5'-CGC GCT GAT GAA AGA GAT GA-3' |
purA-R |
5'-CAT ACG GTA AGC CAC GCA GA-3' |
recA-F |
5'-CGC ATT CGC TTT ACC CTG ACC-3' |
recA-R |
5'-TCG TCG AAA TCT ACG GAC CGG A-3' |
Pmcr-1_A31-F(SalI) |
5'-ACGC GTCGAC GAT ATT CAA CAG GTG ATC AAT AAA-3' |
Pmcr-1_A31-R(EcoRI) |
5'-CCG GAATTC CAT GAG AAA CTA CTC AAA AAA TAA A-3' |
Pmcr-1_E15017-F(SalI) |
5'-ACGC GTCGAC CTT GGA AAA CAA TTT GTC CAG G-3' |
lacZ-R |
5'-CAG TGA ATC CGT AAT CAT GGT C-3' |
*The underlined sequences denote restriction enzyme sites
The integrated evidence accumulated here allowed us to temporarily divide them into eight groups (numbered with 1, 2, 3, … , 8) (Figure 2A). Type 1 plasmid is featuring with only PCR-positive for the mcr-1 gene (Figures 2A and 3C). Besides the mcr-1 gene, two more genes with expected size (nikB (Figure 3A) and tnpA (Figure 3B) are PCR-positive in Type 2 (Figure 2A). Unlike the type 4 plasmid whose PCR amplicons are identical to those of the paradigm mcr-1-containing plasmid pHNSHP45 (Figures 3A-3G) [8], Type 3 plasmid lacks the tnpA gene in front of the mcr-1 locus (Figures 2A and 3B). In relative to the type 3 plasmid, Type 5 plasmid unexpectedly has a longer version of hp locus (Figure 3D) that was subsequently found to encode an extra-insert sequence (IS1) with 97% similarity to the counterpart in Acinetobacter baumannii (Figure 2A). In the following three types of plasmids (Types 6, 7 and 8), the tnpA loci exhibited with PCR amplicons of various lengths (Figure 3B).
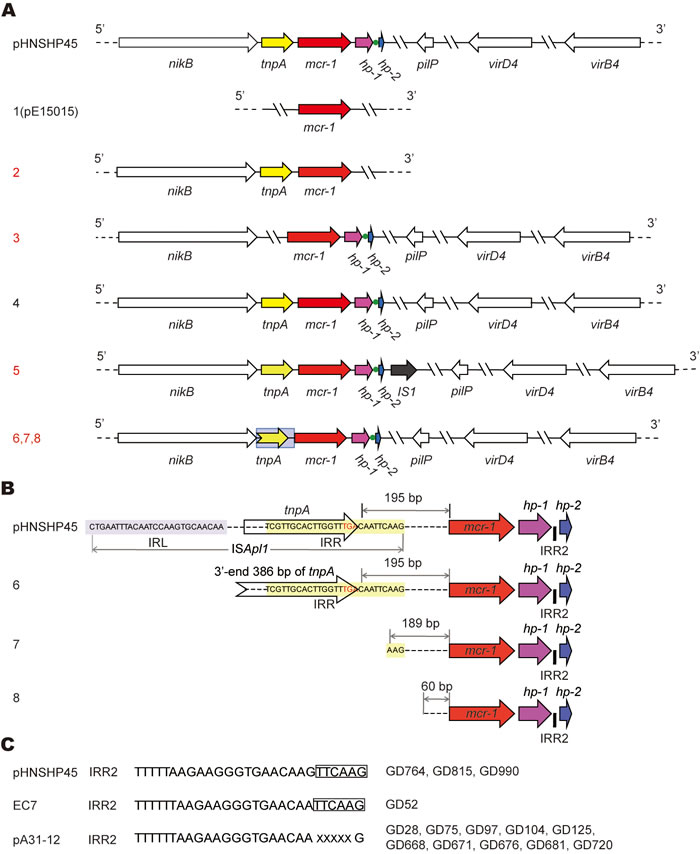
Figure 2: Genetic diversity in the mcr-1-harbouring plasmids from the swine gut microbiota. A. Scheme of the eight types of the mcr-1-harbouring plasmids. The plasmid pHNSHP45 is a prototype with known genome sequence [8], whereas the other eight types of plasmids are revealed after PCR-based sequencing of the 89 representative mcr-1-containing plasmids collected from three pig farms in Guangdong province of China, in 2016. Arrows denote the known (and/or putative) genes/loci. The mcr-1 gene is highlighted in red and 100% identical. The broken arrow marked in grey background denotes the partial sequence of tnpA gene at its 3’-end. The tnpA locus is PCR-negative in type 1 and 3. The green dot represents the IRR2 site. Apart from the type 1 and 2, all the other six types (3-8) are PCR-positive for the four genes (nikB, pilP, virD4 and virB4), as well as a hypothetical protein (hp), adjacent to the 3’-end of mcr-1. Unlike the other types with an intact tnpA adjacent to the 5’-end of mcr-1, types 6, 7 and 8 feature with the truncated versions of tnpA. In type 1 like pE15015 we recently reported, only the mcr-1 locus is PCR-positive. In type 2, two more loci (nikB and tnpA) can be detected in the PCR assays. In particular, an extra-insert IS1 transposase with 97% identity to the counterpart of Acinetobacter baumannii by BlastX (illustrated with a dark arrow) is closely present at the 3-end of hp in the type 5 of plasmid. In addition to the two known types (1 and 4) we reported very recently [9], we show six new plasmid types (2, 3, 5, 6, 7 and 8 labeled in red) in this study. B. Molecular insights into truncated versions of the mcr-1-containing ISApl1 mobile element. In the new plasmid types 6, 7 and 8, the tnpA transposase gene is variously featuring with truncated versions. Designations: IRL, Inverted Region Left; IRR, Inverted Region Right. C. Sequence feature of the IRR2 site . X denotes the variable nucleotide.
Variation in inverted repeat right (IRR)
Generally, the ISApl1 insertion sequence is detected upstream of mcr-1 by recognizing its own IRL (terminal inverted repeat left) and the closest downstream sequence that resembled its IRR (terminal inverted repeat right) (Figure 2B). The IRR2 seemed a flexible/imperfect IRR-like sequence (Figure 2C) [25]. Further genetic dissection defined that i) the terminal inverted repeat right (IRR, “TCGTTGCACTTGGTTTGACAATTCAAG”) remains 195 bp ahead of the mcr-1 gene in Type 6 plasmid (Figure 2B); ii) the full sequence of ISApl1 is lost in the type 7 & 8 plasmids, remaining different relics with various DNA fragment left upstream of the mcr-1 locus (189 bp for Type 7, and 60 bp for Type 8) (Figure 2B); iii) the site of IRR2 (Figure 2B) is consistently present in the inter-genic region between hp-1 and hp-2 (Figure 2C). IRR2 of GD52 other than GD764, GD815, and GD990 is identical to that pHNSHP45 (TTTTTAAGAAGGGTGAACAAGTTCAAG). Intriguingly, the IRR2 (TTTTTTAAGAAGGGTGAACAAXXXXXG) of pA31-12 [25] was seen in ten strains/plasmids like GD28, GD75, GD97, etc. (Figure 2C).
Extensive analysis of 89 mcr-1-containing sequences suggested that 80% of plasmids belonged to Type 1, and the plasmid of Type 6/7/8 (15% in total) is next to Type 1 (Figure 3H). Unlike the plasmids of the prevalent type 1, the plasmids of Type 2 (or 3) are pretty rare (1-2%, Figure 3H). Given the fact that only Type 1 and 3 plasmids have ever been observed in our earlier investigation [9, 12, 13], our findings here extended significantly the proposal that the mcr-1 gene is carried by plasmids with diversified genetic environments.
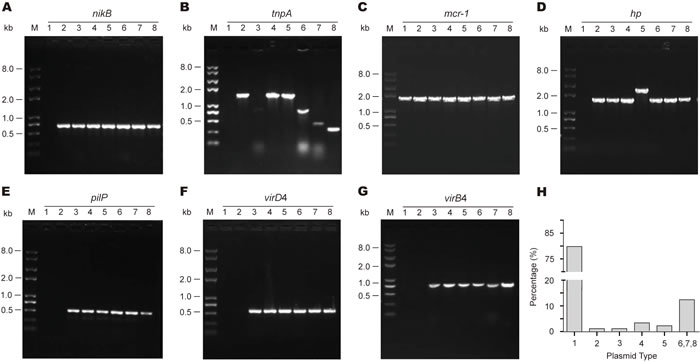
Figure 3: Genetic diversity of the mcr-1-harbouring plasmid backbones from the swine gut microbiota. A. PCR-based discovery of nikB, the relaxosome protein. B. Molecular detection of tnpA, the transposase-encoding gene. C. PCR screen for the mcr-1 gene. D. PCR detection of a putative protein (hp) gene that is supposed to be localized downstream of the mcr-1 locus. E. PCR assay of the pilP gene encoding the type IV pilus biosynthesis protein. Molecular probing of a type IV secretion system-encoding genes virD4. F. and virB4. G. As we recently reported [9], we applied eight pairs of specific primers that are supposed to target the eight unique genes/loci localized on the paradigm mcr-1-harbouring plasmid pHNSHP45 [8]. According to the different profile of PCR amplicon, 8 types of genetic environment in total (numbered from 1, 2 … 8) are assigned to these mcr-1-positive plasmids from the swine gut bacteria. M denotes Trans 2K Plus II DNA Ladder (TRANSGEN BIOTECH, Beijing, China), and kb refers to kilo-base pair. H. Estimated distribution of mcr-1-carrying plasmids featuring with different genetic environment. Totally, 89 representative strains were tested here.
Complexity in sequence types of the mcr-1-positive E. coli isolates
To test the genetic heterogeneity amongst the mcr-1-positive E. coli isolates, we thus performed the analyses of multi-locus sequence typing (MLST) using 15 representative E. coli isolates [23]. The MLST-based sequence typing showed that they comprise 10 different sequence types (STs) including ST98, ST10, ST20, ST218, ST165, ST641, ST93, ST1286, ST4656, ST3546, and a novel ST (n = 2) (Table 3). The novel ST (allelic profile: 10-11-5-10-8-236-2) is assigned to two E. coli isolates GD97 and GD676. Most of the STs we determined, were not previously related to the mcr-1 gene. But the ST10 E. coli in Belgium was ever found to carry ESBL-producing plasmids and associated with human infection [27]. By contrast, we recently observed co-production of MCR-1 and ESBL in the epidemic strain of ST648 E. coli [23]. Obviously, the mcr-1-positive E. coli isolates from the swine populations exhibited appreciably genetic heterogeneity in terms of diversified STs (Table 3).
Table 3: Sequence typing of the mcr-1-positive E. coli strains
Strains |
Alleles |
ST |
ST Complex |
||||||
adk |
fumC |
gyrB |
icd |
mdh |
purA |
recA |
|||
MG1655 |
10 |
11 |
4 |
39 |
8 |
8 |
2 |
ST98 |
ST10 Cplx |
GD28 |
10 |
11 |
4 |
8 |
8 |
8 |
2 |
ST10 |
ST10 Cplx |
GD428 |
10 |
11 |
4 |
8 |
8 |
8 |
2 |
ST10 |
ST10 Cplx |
GD703 |
10 |
11 |
4 |
8 |
8 |
8 |
2 |
ST10 |
ST10 Cplx |
GD75 |
6 |
4 |
3 |
18 |
7 |
7 |
6 |
ST20 |
ST20 Cplx |
GD104 |
6 |
4 |
3 |
18 |
7 |
7 |
6 |
ST20 |
ST20 Cplx |
GD125 |
6 |
4 |
3 |
18 |
7 |
7 |
6 |
ST20 |
ST20 Cplx |
GD66 |
10 |
11 |
4 |
12 |
8 |
8 |
2 |
ST218 |
ST10 Cplx |
GD668 |
10 |
27 |
5 |
10 |
12 |
8 |
2 |
ST165 |
ST165 Cplx |
GD788 |
9 |
6 |
33 |
131 |
24 |
8 |
7 |
ST641 |
ST86 Cplx |
GD811 |
6 |
11 |
4 |
10 |
7 |
8 |
6 |
ST93 |
ST168 Cplx |
GD720 |
10 |
174 |
4 |
8 |
8 |
8 |
2 |
ST1286 |
/ |
GD815 |
6 |
11 |
4 |
8 |
8 |
18 |
2 |
ST4656 |
/ |
GD819 |
10 |
11 |
57 |
8 |
8 |
8 |
20 |
ST3546 |
/ |
GD97 |
10 |
11 |
5 |
10 |
8 |
N/A |
2 |
New ST |
/ |
GD676 |
10 |
11 |
5 |
10 |
8 |
N/A |
2 |
New ST |
/ |
The mcr-1 promoter
The mcr-1 promoter regions from the two plasmids (pA31-12 and pGD97) were compared using multiple sequence alignment, indicating that it is not less conserved at 5’-end than that at 3’-end (Figure 4A). Prokaryotic promoter analysis by the Neutral Network Program of Promoter Prediction (http://www.fruitfly.org/seq_tools/promoter.html) suggested that the transcription start site of the mcr-1 gene is C at the position of 36 bp upstream of the translation initiation site “ATG” (Figure 4A), which is 1 bp upstream in comparison with the transcription start site “A’ revealed by Poirel et al. [28] with 5’-RACE. The minor difference might be due to mapping of the truncated version of mcr-1 transcript by 5’-RACE. To evaluate the promoter activity of the mcr-1 gene, the mcr-1 promoter regions (303 bp) of the two plasmids (pA31-12 and pGD97) were fused to the promoter-less lacZ gene, giving Pmcr-1-lacZ transcriptional fusions. Subsequent LacZ analyses showed that β-gal activity driven by the mcr-1 promoters of both pGD97 and pA31-12 is ~400 miller units (Figure 4B). It suggested that both pGD97 and pA31-12 has a mcr-1 promoter with comparable medium strength. Intriguingly, this speculation was proved by the fact the two mcr-1-positive strains (GD97 and A31-12) exhibited a similar level of colistin resistance to colistin (~16 mg/L) (Figure 4C). Given the fact that colistin resistance by the arabinose-induced expression of MCR-1 is similar to that of GD97 and A31-12, we therefore concluded that mcr-1 promoter is comparable to the arabinose-inducible promoter of pBAD24.
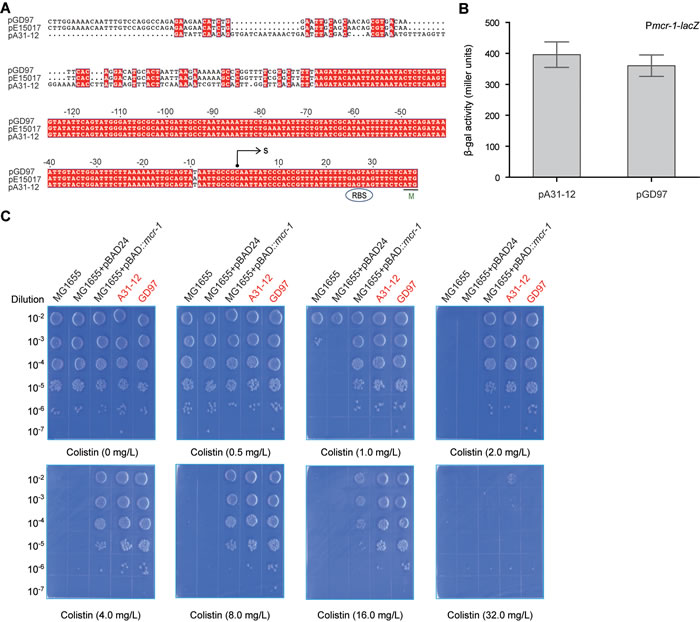
Figure 4: The mcr-1 promoter activity determines the level of the E. coli resistance to colistin. A. Bioinformatics analyses of the mcr-1 promoter. The putative mcr-1 promoter sequences were derived from the two Guangdong’s mcr-1-positive plasmids (pA31-12 [25] and pGD97, a representative of plasmids we report here). The nucleotide sequences were aligned with ClustalW2 (http://www.ebi.ac.uk/Tools/clustalw2/index.html), and the resultant output was generated with ESPript 2.2 (http://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi) [34, 35]. Identical residues are white letters with red background, similar residues are black letters, and gaps are denoted with dots. Designations: S, transcription start site; M, methionine and translation initiation site; RBS, ribosome binding site. B. Measurement of β-gal activity driven by the mcr-1 promoter. To measure bacterial β-gal activity, mid-log phase cultures of E. coli MC1061 with the Pmcr-1-lacZ transcriptional fusion on chromosome were collected by spinning and suspended in Z-buffer [36]. The data was recorded in triplicate in three independent experiments. The two strains used here include FYJ853 (Pmcr-1_pA31-12-lacZ for the mcr-1 promoter of A31-12 strain) and FYJ 851 (Pmcr-1_pGD97-lacZ for the mcr-1 promoter of GD97 strain). Note: the mcr-1 promoter sequences (~300 bp region upstream the transcription start site C) are identical. C. Visualization of the colistin resistance of E. coli conferred by the mcr-1-harbouring plasmids. The E. coli strain MG1655 with/without the pBAD24 vector acts as the negative control, whereas the MG1655 with the arabinose-driven MCR-1 expression refers to the positive control. The remaining two (A31-12 and GD97) are clinical mcr-1-positive isolates (highlighted in red). The minimum inhibitory concentration (MIC) of the colistin was determined with the method of plating in series of dilution. Briefly, the mid-log phase cultures (OD600 = 0.8) in serial dilution were spotted on LBA plates with different level of colistin (0, 0.5, 1.0, 2.0, 4.0, 8.0, 16.0 and 32.0 mg/L) and 0.2% arabinose. The LBA plates were maintained overnight at 37oC.
Conclusions
Colistin is a last defense against lethal infections by Gram-negative pathogens with multiple drug resistance. The MCR-1 mediates the transferable resistance to colistin, raising an old, but newly-emerging threat to public health [10]. In the past six months since the first discovery of the mcr-1 colistin resistance gene in Southern China [8], the literatures regarding to the mcr-1/colistin resistance are increasingly-accumulated (http://www.ncbi.nlm.nih.gov/pubmed/?term = mcr+1+colistin). Among them, most of studies concentrated on molecular epidemiology of the mcr-1 transmission/dissemination. In particular, unexpected complexity in the multi-drug resistance was assigned to the mcr-1-positive enteric bacteria [14, 23]. More seriously, the MCR-1 was found to co-localize with two types of notorious drug resistant genes (ESBL [20, 23, 26] and NDM-1 (and/or its mutants) [29, 30]. We are first to report the diversified mcr-1-harbouring plasmids from clinical E. coli isolates of diarrhea patients in China [9]. The similar scenarios were also seen in the E. coli isolates from the swine populations [12].
In this study, we screened over 1000 pieces of E. coli samples (from Guangdong Province, China) for the presence of the mcr-1 gene (Figure 1). It allowed us to gain insights into genetic environment of the mcr-1-bearing plasmids. In addition to the two known plasmid types (types 1 and 3), we observed four more kinds of plasmid types (Figure 2), suggesting the unprecedented complexity in the mcr-1-carrying plasmids. Also, the variation is present in the IRR2 motif. It is not surprise for us to notice nearly 10 different sequence types can be assigned to these mcr-1-positive E. coli isolates. To the best of our knowledge, it might represent an example of complicated genetic diversity in both mcr-1-carrying plasmids and E. coli hosts originating from the healthy swine populations in China. Because that the IRR sites are involved in the transposon-like transferable events of the mcr-1 gene, the various versions of IRR2 might imply difference/diversity among the MCR-1 transfer. Given the fact that the high prevalence and complexity of the MCR-1 colistin resistance in the healthy swine populations in China, it is reasonable that enhanced surveillance efforts is warranted to monitor and/or control the spread of the mcr-1 resistance gene, esp. the possible dissemination of food chain. Additionally, we are first to showe in vitro evidence that the level of colistin resistance in various clinical strains is determined by the promoter activity of the mcr-1 gene (Figure 4B).
In summary, it seems likely that complex dissemination of the diversified mcr-1-harbouring plasmids occurs amongst the various ST E. coli inhabiting the healthy swine populations, in Southern China. In particular, our findings highlighted the urgent need to reconsider the efficacy (safety) of colistin in the veterinary use, and formulate a comprehensive strategy to fight against the diversified plasmid-mediated mcr-1 colistin resistance in pan-drug-resistant Gram-negative bacteria.
Materials and Methods
Bacterial isolations and identification
All the bacterial strains were E. coli derivatives (Table 1). Fecal samples were routinely collected from three different pig farms in Guangdong province, China, in 2016. The nasal fluid and feces of pigs were sampled. MacConkey solid agar plates were applied to isolate the Enterobacterial species. The resultant bacteria were subjected to colony PCR assays for the presence of the mcr-1 gene (Table 2).
DNA manipulations
To address the genetic context surrounding the mcr-1 gene, all of the mcr-1-positive E. coli isolates were subjected to the multiplex-PCR with six pairs of specific primers (Table 2) [9]. The resultant PCR products were determined with Sanger sequencing. To probe the genetic heterogeneity of the mcr-1-positive isolates, MLST was carried out as we described [23]. Seven house-keeping genes examined here correspond to adk, fumC, gyrB, icd, mdh, purA and recA, respectively. The relevant seven pairs of primers were available in the server of MLST ( http://mlst.warwick.ac.uk/mlst/dbs/Ecoli) .
Plasmids and genetic manipulations
The mcr-1 promoters from the following plasmids (pA31-12, pE15017 and pGD97) were amplified with PCR and directly cloned into pAH125, giving the lacZ transcriptional fusions, Pmcr-1-lacZ (Table 1). As we described earlier [31], the resultant recombinant plasmids like pAH-Pmcr-1(pA31-12), were transformed into MC1061, and screened on Luria-Bertani Agar (LBA) plates containing kanamycin and x-gal to give the chromosomal mcr-1 transcriptional lacZ fusion strains like FYJ853 (Table 1).
β-Galactosidase assays
The strains of E. coli carrying the chromosome Pmcr-1-lacZ transcriptional fusion (e.g., FYJ853) were grown in LB media, and mid-log phase cultures were collected assayed for β-galactosidase activity following lysis with SDS (sodium dodecylsulfate)-chloroform [31, 32]. The data were recorded in triplicate from three independent experiments.
Determination of the colistin resistance/tolerance
Antibiotic susceptibility experiments were conducted by the agar dilution method [33] recommended by the Clinical and Laboratory Standards Institute guidelines (CLSI M100-S25). The EUCAST breakpoint for Enterobacteriaceae was applied for colistin resistance (European Committee on Antimicrobial Susceptibility Testing 2015). Here, strain FYJ795 refers to the positive control, whereas the two strains (MG1655 and FYJ796) are the negative controls. All strains including A31-12 and GD97 were collected in Mid-log-phase, then bacterial cultures in a dilution series were spotted on LBA plates supplemented with various levels of colistin (0, 0.5, 1.0, 2.0, 4.0, 8.0, 16.0, and 32.0 mg/liter) and maintained at 37°C overnight. When necessary, 0.2% arabinose was added into the LBA plates for induction of MCR-1 expression [9].
Acknowledgments
Dr. Feng is a recipient of the “Young 1000 Talents” Award.
Conflicts of Interests
The authors declare no conflict of interests.
Fundings
This work was supported by the Basic Research Program of China (2016YFC1200100).
References
1. Paterson DL and Harris PN. Colistin resistance: a major breach in our last line of defence. Lancet Infect Dis. 2016; 16(2):132-133.
2. Kumarasamy KK, Toleman MA, Walsh TR, Bagaria J, Butt F, Balakrishnan R, Chaudhary U, Doumith M, Giske CG, Irfan S, Krishnan P, Kumar AV, Maharjan S, et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: a molecular, biological, and epidemiological study. Lancet Infect Dis. 2010; 10(9):597-602.
3. Poirel L, Hombrouck-Alet C, Freneaux C, Bernabeu S and Nordmann P. Global spread of New Delhi metallo-beta-lactamase 1. Lancet Infect Dis. 2010; 10(12):832.
4. Goel N and Wattal C. New Delhi metallo-beta-lactamase 1. Lancet Infect Dis. 2010; 10(11):751; author reply 752-754.
5. Nation RL, Li J, Cars O, Couet W, Dudley MN, Kaye KS, Mouton JW, Paterson DL, Tam VH, Theuretzbacher U, Tsuji BT and Turnidge JD. Framework for optimisation of the clinical use of colistin and polymyxin B: the Prato polymyxin consensus. Lancet Infec Dis. 2015; 15(2):225-234.
6. Gunn JS. The Salmonella PmrAB regulon: lipopolysaccharide modifications, antimicrobial peptide resistance and more. Trends Microbiol. 2008; 16(6):284-290.
7. Cannatelli A, D’Andrea MM, Giani T, Di Pilato V, Arena F, Ambretti S, Gaibani P and Rossolini GM. In vivo emergence of colistin resistance in Klebsiella pneumoniae producing KPC-type carbapenemases mediated by insertional inactivation of the PhoQ/PhoP mgrB regulator. Antimicrob Agents Chemother. 2013; 57(11):5521-5526.
8. Liu YY, Wang Y, Walsh TR, Yi LX, Zhang R, Spencer J, Doi Y, Tian G, Dong B, Huang X, Yu LF, Gu D, Ren H, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016; 16(2):161-168.
9. Ye H, Li Y, Li Z, Gao R, Zhang H, Wen R, Gao GF, Hu Q and Feng Y. Diversified mcr-1-harbouring plasmid reservoirs confer resistance to colistin in human gut microbiota. mBio. 2016; 7(2):e00177.
10. Schwarz S and Johnson AP. Transferable resistance to colistin: a new but old threat. Journal Antimicrob Chemother. 2016; 71(8):2066-2070.
11. Shen Z, Wang Y, Shen Y, Shen J and Wu C. Early emergence of mcr-1 in Escherichia coli from food-producing animals. Lancet Infect Dis. 2016; 16(3):293.
12. Li Z, Tan C, Lin J and Feng Y. Diversified variants of the mcr-1-carrying plasmid reservoir in the swine lung microbiota. Sci China Life Sci. 2016; 59(9):971-973.
13. Gao R, Wang Q, Li P, Li Z and Feng Y. Genome sequence and characteristics of plasmid pWH12, a variant of the mcr-1-harbouring plasmid pHNSHP45, from the multi-drug resistant E. coli. Virulence. 2016:1-4.
14. Gao R, Li Y, Lin J, Tan C and Feng Y. Unexpected complexity of multidrug resistance in the mcr-1-harbouring Escherichia coli. Sci China Life Sci. 2016; 59(7):732-734.
15. Zeng KJ, Doi Y, Patil S, Huang X and Tian GB. Emergence of the Plasmid-Mediated mcr-1 Gene in Colistin-Resistant Enterobacter aerogenes and Enterobacter cloacae. Antimicrob Agents Chemother. 2016; 60(6):3862-3863.
16. Di Pilato V, Arena F, Tascini C, Cannatelli A, Henrici De Angelis L, Fortunato S, Giani T, Menichetti F and Rossolini GM. MCR-1.2: a new MCR variant encoded by a transferable plasmid from a colistin-resistant KPC carbapenemase-producing Klebsiella pneumoniae of sequence type 512. Antimicrob Agents Chemother. 2016; 60(9):5612-5615.
17. Gu DX, Huang YL, Ma JH, Zhou HW, Fang Y, Cai JC, Hu YY and Zhang R. Detection of Colistin Resistance Gene mcr-1 in Hypervirulent Klebsiella pneumoniae and Escherichia coli Isolates from an Infant with Diarrhea in China. Antimicrob Agents Chemother. 2016; 60(8):5099-5100.
18. Stoesser N, Mathers AJ, Moore CE, Day NP and Crook DW. Colistin resistance gene mcr-1 and pHNSHP45 plasmid in human isolates of Escherichia coli and Klebsiella pneumoniae. Lancet Infect Dis. 2016; 16(3):285-286.
19. Doumith M, Godbole G, Ashton P, Larkin L, Dallman T, Day M, Day M, Muller-Pebody B, Ellington MJ, de Pinna E, Johnson AP, Hopkins KL and Woodford N. Detection of the plasmid-mediated mcr-1 gene conferring colistin resistance in human and food isolates of Salmonella enterica and Escherichia coli in England and Wales. J Antimicrob Chemother. 2016; 71(8):2300-2305.
20. Yang YQ, Zhang AY, Ma SZ, Kong LH, Li YX, Liu JX, Davis MA, Guo XY, Liu BH, Lei CW and Wang HN. Co-occurrence of mcr-1 and ESBL on a single plasmid in Salmonella enterica. J Antimicrob Chemother. 2016; 71(8):2336-2338.
21. Quesada A, Ugarte-Ruiz M, Iglesias MR, Porrero MC, Martinez R, Florez-Cuadrado D, Campos MJ, Garcia M, Piriz S, Saez JL and Dominguez L. Detection of plasmid mediated colistin resistance (MCR-1) in Escherichia coli and Salmonella enterica isolated from poultry and swine in Spain. Re Vet Sci. 2016; 105:134-135.
22. Pham Thanh D, Thanh Tuyen H, Nguyen Thi Nguyen T, Chung The H, Wick RR, Thwaites GE, Baker S and Holt KE. Inducible colistin resistance via a disrupted plasmid-borne mcr-1 gene in a 2008 Vietnamese Shigella sonnei isolate. J Antimicrob Chemother. 2016; 71(8):2314-2317.
23. Zhang H, Seward CH, Wu Z, Ye H and Feng Y. Genomic insights into the ESBL and MCR-1-producing ST648 Escherichia coli with multi-drug resistance. Sci Bull (Beijing). 2016; 61:875-878.
24. Xavier BB, Lammens C, Butaye P, Goossens H and Malhotra-Kumar S. Complete sequence of an IncFII plasmid harbouring the colistin resistance gene mcr-1 isolated from Belgian pig farms. J Antimicrob Chemother. 2016; 71(8):2342-2344.
25. Sun J, Li XP, Yang RS, Fang LX, Huo W, Li SM, Jiang P, Liao XP and Liu YH. Complete nucleotide sequence of an IncI2 plasmid coharboring blaCTX-M-55 and mcr-1. Antimicrob Agents Chemother. 2016; 60(8):5014-5017.
26. Li A, Yang Y, Miao M, Chavda KD, Mediavilla JR, Xie X, Feng P, Tang YW, Kreiswirth BN, Chen L and Du H. Complete sequences of mcr-1-harboring plasmids from extended-spectrum-beta-lactamase- and carbapenemase-producing Enterobacteriaceae. Antimicrob Agents Chemother. 2016; 60(7):4351-4354.
27. Leverstein-van Hall MA, Dierikx CM, Cohen Stuart J, Voets GM, van den Munckhof MP, van Essen-Zandbergen A, Platteel T, Fluit AC, van de Sande-Bruinsma N, Scharinga J, Bonten MJ, Mevius DJ and National Esg. Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin Microbiol Infect. 2011; 17(6):873-880.
28. Poirel L, Kieffer N, Brink A, Coetze J, Jayol A and Nordmann P. Genetic features of MCR-1-producing colistin-resistant Escherichia coli isolates in South Africa. Antimicrob Agents Chemother. 2016; 60(7):4394-4397.
29. Delgado-Blas JF, Ovejero CM, Abadia Patino L and Gonzalez-Zorn B. Coexistence of mcr-1 and blaNDM-1 in Escherichia coli from Venezuela. Antimicrob Agents Chemother. 2016; 60(10):6356-6358.
30. Yao X, Doi Y, Zeng L, Lv L and Liu JH. Carbapenem-resistant and colistin-resistant Escherichia coli co-producing NDM-9 and MCR-1. Lancet Infect Dis. 2016; 16(3):288-289.
31. Feng Y and Cronan JE. Crosstalk of Escherichia coli FadR with global regulators in expression of fatty acid transport genes. PLoS One. 2012; 7(9):e46275.
32. Feng Y and Cronan JE. Overlapping repressor binding sites result in additive regulation of Escherichia coli FadH by FadR and ArcA. J Bacteriol. 2010; 192(17):4289-4299.
33. Standards NCfCL. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically—Ninth Edition: Approved standard M7-A9. 2012.
34. Feng Y and Cronan JE. The Vibrio cholerae fatty acid regulatory protein, FadR, represses transcription of plsB, the gene encoding the first enzyme of membrane phospholipid biosynthesis. Mol Microbiol. 2011; 81(4):1020-1033.
35. Robert X and Gouet P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014; 42(Web Server issue):W320-324.
36. Gao R, Lin J, Zhang H and Feng Y. Transcriptional repression of the VC2105 protein by Vibrio FadR suggests that it is a new auxiliary member of the fad regulon. Appl Environ Microbiol. 2016; 82(9):2819-2832.